
Biophysical Modulation of Mesenchymal Stem Cell Differentiation in the Context of Skeletal Repair


Abstract
:1. Introduction
1.1. Influence of the Biophysical Environment and Cell Reception
1.2. Mechanical and Electrical Influences on Lineage Expression of MSCs
2. Response to Mesenchymal Cells to Biophysical Regulation and Mechanical Strain Associated Events In Vitro
3. Experimental Differentiation of Mesenchymal Cells to Chondrocytes and Osteocytes by PEMF In Vivo
3.1. Electromagnetic Stimulation of In Vivo Mesenchymal Cell Differentiation, Chondrogenesis, and Osteogenesis
3.2. A Potential Intermediary Mechanism of Tissue Stimulation by Electromagnetic Exposure
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Aaron, R.K.; Ciombor, D.M.; Wang, S.; Simon, B. Clinical Biophysics: The Promotion of Skeletal Repair by Physical Forces. Ann. N. Y. Acad. Sci. 2006, 1068, 513–531. [Google Scholar] [CrossRef] [PubMed]
- Jaalouk, D.E.; Lammerding, J. Mechanotransduction gone awry. Nat. Rev. Mol. Cell Biol. 2009, 10, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, F.; Perestrelo, A.R.; Vinarský, V.; Pagliari, S.; Forte, G. Cellular Mechanotransduction: From Tension to Function. Front. Physiol. 2018, 9, 824. [Google Scholar] [CrossRef] [PubMed]
- Minguell, J.J.; Erices, A.; Conget, P. Mesenchymal stem cells. Exp. Biol. Med. 2001, 226, 507–520. [Google Scholar] [CrossRef]
- Discher, D.E.; Janmey, P.; Wang, Y.-L. Tissue Cells Feel and Respond to the Stiffness of Their Substrate. Science 2005, 310, 1139–1143. [Google Scholar] [CrossRef] [Green Version]
- Gavazzo, P.; Viti, F.; Donnelly, H.; Oliva, M.A.G.; Salmeron-Sanchez, M.; Dalby, M.J.; Vassalli, M. Biophysical phenotyping of mesenchymal stem cells along the osteogenic differentiation pathway. Cell Biol. Toxicol. 2021, 37, 915–933. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Marie, P.J. Transcription factors controlling osteoblastogenesis. Arch. Biochem. Biophys. 2008, 473, 98–105. [Google Scholar] [CrossRef]
- Govey, P.M.; Loiselle, A.E.; Donahue, H.J. Biophysical Regulation of Stem Cell Differentiation. Curr. Osteoporos. Rep. 2013, 11, 83–91. [Google Scholar] [CrossRef]
- Fu, J.; Wang, Y.-K.; Yang, M.T.; Desai, R.; Yu, X.; Liu, Z.; Chen, C. Mechanical regulation of cell function with geometrically modulated elastomeric substrates. Nat. Methods 2010, 7, 733–736. [Google Scholar] [CrossRef]
- Cun, X.; Hosta-Rigau, L. Topography: A Biophysical Approach to Direct the Fate of Mesenchymal Stem Cells in Tissue Engineering Applications. Nanomaterials 2020, 10, 2070. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, O.; Gu, L.; Klumpers, D.; Darnell, M.; Bencherif, S.A.; Weaver, J.C.; Huebsch, N.; Lee, H.-P.; Lippens, E.; Duda, G.N.; et al. Hydrogels with tunable stress relaxation regulate stem cell fate and activity. Nat. Mater. 2015, 15, 326–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-P.; Gu, L.; Mooney, D.J.; Levenston, M.E.; Chaudhuri, O. Mechanical confinement regulates cartilage matrix formation by chondrocytes. Nat. Mater. 2017, 16, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Mow, V.C.; Wang, C.C.; Hung, C.T. The extracellular matrix, interstitial fluid and ions as a mechanical signal transducer in articular cartilage. Osteoarthr. Cartil. 1999, 7, 41–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, D.R.; Lindsey, D.P.; Li, K.W.; Tummala, P.; Chandran, S.E.; Smith, R.L.; Longaker, M.T.; Carter, D.R.; Beaupre, G.S. Hydrostatic Pressure Enhances Chondrogenic Differentiation of Human Bone Marrow Stromal Cells in Osteochondrogenic Medium. Ann. Biomed. Eng. 2008, 36, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.-J.; Seedhom, B.B. Light and low-frequency pulsatile hydrostatic pressure enhances extracellular matrix formation by bone marrow mesenchymal cells: An in-vitro study with special reference to cartilage repair. Proc. Inst. Mech. Eng. Part H J. Eng. Med. 2007, 221, 499–507. [Google Scholar] [CrossRef]
- Haudenschild, A.K.; Hsieh, A.H.; Kapila, S.; Lotz, J.C. Pressure and Distortion Regulate Human Mesenchymal Stem Cell Gene Expression. Ann. Biomed. Eng. 2009, 37, 492–502. [Google Scholar] [CrossRef] [Green Version]
- Tummala, P.; Arnsdorf, E.J.; Jacobs, C.R. The Role of Primary Cilia in Mesenchymal Stem Cell Differentiation: A Pivotal Switch in Guiding Lineage Commitment. Cell. Mol. Bioeng. 2010, 3, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Hoey, D.A.; Tormey, S.; Ramcharan, S.; O’Brien, F.J.; Jacobs, C.R. Primary Cilia-Mediated Mechanotransduction in Human Mesenchymal Stem Cells. Stem Cells 2012, 30, 2561–2570. [Google Scholar] [CrossRef] [Green Version]
- Wann, A.K.T.; Zuo, N.; Haycraft, C.J.; Jensen, C.G.; Poole, C.A.; McGlashan, S.R.; Knight, M.M. Primary cilia mediate mechanotransduction through control of ATP-induced Ca2+ signaling in compressed chondrocytes. FASEB J. 2011, 26, 1663–1671. [Google Scholar] [CrossRef] [Green Version]
- Deren, M.E.; Yang, X.; Guan, Y.; Chen, Q. Biological and Chemical Removal of Primary Cilia Affects Mechanical Activation of Chondrogenesis Markers in Chondroprogenitors and Hypertrophic Chondrocytes. Int. J. Mol. Sci. 2016, 17, 188. [Google Scholar] [CrossRef] [PubMed]
- González-Cruz, R.D.; Fonseca, V.C.; Darling, E.M. Cellular mechanical properties reflect the differentiation potential of adipose-derived mesenchymal stem cells. Proc. Natl. Acad. Sci. USA 2012, 109, E1523–E1529. [Google Scholar] [CrossRef] [Green Version]
- Ofek, G.; Willard, V.P.; Koay, E.J.; Hu, J.C.; Lin, P.; Athanasiou, K.A. Mechanical Characterization of Differentiated Human Embryonic Stem Cells. J. Biomech. Eng. 2009, 131, 061011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, S.-J.; Driscoll, T.; Thorpe, S.D.; Nerurkar, N.L.; Baker, B.; Yang, M.T.; Chen, C.; Lee, D.; Mauck, R.L. Differentiation alters stem cell nuclear architecture, mechanics, and mechano-sensitivity. ELife 2016, 5, e18207. [Google Scholar] [CrossRef] [PubMed]
- Alegre-Aguarón, E.; Sampat, S.R.; Xiong, J.C.; Colligan, R.M.; Bulinski, J.C.; Cook, J.L.; Ateshian, G.A.; Brown, L.M.; Hung, C.T. Growth Factor Priming Differentially Modulates Components of the Extracellular Matrix Proteome in Chondrocytes and Synovium-Derived Stem Cells. PLoS ONE 2014, 9, e88053. [Google Scholar] [CrossRef] [Green Version]
- Oswald, E.S.; Brown, L.M.; Bulinski, J.C.; Hung, C.T. Label-Free Protein Profiling of Adipose-Derived Human Stem Cells under Hyperosmotic Treatment. J. Proteome Res. 2011, 10, 3050–3059. [Google Scholar] [CrossRef] [Green Version]
- Oswald, E.S.; Ahmed, H.S.; Kramer, S.P.; Bulinski, J.C.; Ateshian, G.A.; Hung, C.T. Effects of Hypertonic (NaCl) Two-Dimensional and Three-Dimensional Culture Conditions on the Properties of Cartilage Tissue Engineered from an Expanded Mature Bovine Chondrocyte Source. Tissue Eng. Part C Methods 2011, 17, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Sampat, S.R.; Dermksian, M.V.; Oungoulian, S.R.; Winchester, R.J.; Bulinski, J.C.; Ateshian, G.A.; Hung, C.T. Applied osmotic loading for promoting development of engineered cartilage. J. Biomech. 2013, 46, 2674–2681. [Google Scholar] [CrossRef] [Green Version]
- Sampat, S.R.; O’Connell, G.D.; Fong, J.V.; Alegre-Aguarón, E.; Ateshian, G.A.; Hung, C.T. Growth Factor Priming of Synovium-Derived Stem Cells for Cartilage Tissue Engineering. Tissue Eng. Part A 2011, 17, 2259–2265. [Google Scholar] [CrossRef] [Green Version]
- Freeman, F.E.; Schiavi, J.; Brennan, M.; Owens, P.; Layrolle, P.; McNamara, L.M. Mimicking the Biochemical and Mechanical Extracellular Environment of the Endochondral Ossification Process to Enhance the In Vitro Mineralization Potential of Human Mesenchymal Stem Cells. Tissue Eng. Part A 2017, 23, 1466–1478. [Google Scholar] [CrossRef]
- Zhao, Z.; Watt, C.; Karystinou, A.; Roelofs, A.; McCaig, C.; Gibson, I.; De Bari, C. Directed migration of human bone marrow mesenchymal stem cells in a physiological direct current electric field. Eur. Cells Mater. 2011, 22, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.; Alegre-Aguarón, E.; O’Connell, G.; VandenBerg, C.; Aaron, R.; Vunjak-Novakovic, G.; Bulinski, J.C.; Ateshian, G.; Hung, C. Passage-dependent relationship between mesenchymal stem cell mobilization and chondrogenic potential. Osteoarthr. Cartil. 2015, 23, 319–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, J.H.; van der Jagt, O.P.; Punt, B.J.; Verhaar, J.A.; van Leeuwen, J.P.; Weinans, H.; Jahr, H. Stimulation of osteogenic differentiation in human osteoprogenitor cells by pulsed electromagnetic fields: An in vitro study. BMC Musculoskelet. Disord. 2010, 11, 188. [Google Scholar] [CrossRef] [Green Version]
- Parate, D.; Franco-Obregón, A.; Fröhlich, J.; Beyer, C.; Abbas, A.A.; Kamarul, T.; Hui, J.H.P.; Yang, Z. Enhancement of mesenchymal stem cell chondrogenesis with short-term low intensity pulsed electromagnetic fields. Sci. Rep. 2017, 7, 9421. [Google Scholar] [CrossRef] [PubMed]
- Parate, D.; Kadir, N.D.; Celik, C.; Lee, E.H.; Hui, J.H.P.; Franco-Obregón, A.; Yang, Z. Pulsed electromagnetic fields potentiate the paracrine function of mesenchymal stem cells for cartilage regeneration. Stem Cell Res. Ther. 2020, 11, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, A.; Farrell, M.J.; Mauck, R.L. Mechanics and mechanobiology of mesenchymal stem cell-based engineered cartilage. J. Biomech. 2010, 43, 128–136. [Google Scholar] [CrossRef] [Green Version]
- Huang, A.H.; Farrell, M.J.; Kim, M.; Mauck, R.L. Long-term dynamic loading improves the mechanical properties of chondrogenic mesenchymal stem cell-laden hydrogel. Eur. Cells Mater. 2010, 19, 72–85. [Google Scholar] [CrossRef]
- Mauck, R.L.; Hung, C.T.; Ateshian, G.A. Modeling of Neutral Solute Transport in a Dynamically Loaded Porous Permeable Gel: Implications for Articular Cartilage Biosynthesis and Tissue Engineering. J. Biomech. Eng. 2003, 125, 602–614. [Google Scholar] [CrossRef] [Green Version]
- Simmons, C.A.; Matlis, S.; Thornton, A.J.; Chen, S.; Wang, C.Y.; Mooney, D.J. Cyclic strain enhances matrix mineralization by adult human mesenchymal stem cells via the extracellular signal-regulated kinase (ERK1/2) signaling pathway. J. Biomech. 2003, 36, 1087–1096. [Google Scholar] [CrossRef]
- Carroll, S.F.; Buckley, C.; Kelly, D.J. Cyclic Tensile Strain Can Play a Role in Directing both Intramembranous and Endochondral Ossification of Mesenchymal Stem Cells. Front. Bioeng. Biotechnol. 2017, 5, 73. [Google Scholar] [CrossRef]
- Mauney, J.R.; Sjostorm, S.; Blumberg, J.; Horan, R.; O’Leary, J.P.; Vunjak-Novakovic, G.; Volloch, V.; Kaplan, D.L. Mechanical Stimulation Promotes Osteogenic Differentiation of Human Bone Marrow Stromal Cells on 3-D Partially Demineralized Bone Scaffolds In Vitro. Calcif. Tissue Res. 2004, 74, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Sumanasinghe, R.D.; Bernacki, S.H.; Loboa, E.G. Osteogenic differentiation of human mesenchymal stem cells in collagen matrices: Effect of uniaxial cyclic tensile strain on bone morphogenetic protein (BMP-2) mRNA expression. Tissue Eng. 2006, 12, 3459–3465. [Google Scholar] [CrossRef] [PubMed]
- Schreivogel, S.; Kuchibhotla, V.; Knaus, P.; Duda, G.N.; Petersen, A. Load-induced osteogenic differentiation of mesenchymal stromal cells is caused by mechano-regulated autocrine signaling. J. Tissue Eng. Regen. Med. 2019, 13, 1992–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stavenschi, E.; Corrigan, M.A.; Johnson, G.P.; Riffault, M.; Hoey, D.A. Physiological cyclic hydrostatic pressure induces osteogenic lineage commitment of human bone marrow stem cells: A systematic study. Stem Cell Res. Ther. 2018, 9, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.-H.; Lv, X.; Liu, Y.-L.; Zhao, Y.; Li, Q.; Chen, Y.-J.; Zhang, M. Hydrostatic pressure promotes the proliferation and osteogenic/chondrogenic differentiation of mesenchymal stem cells: The roles of RhoA and Rac. Stem Cell Res. 2015, 14, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Kisiday, J.D.; Frisbie, D.D.; McIlwraith, C.W.; Grodzinsky, A.J. Dynamic Compression Stimulates Proteoglycan Synthesis by Mesenchymal Stem Cells in the Absence of Chondrogenic Cytokines. Tissue Eng. Part A 2009, 15, 2817–2824. [Google Scholar] [CrossRef] [Green Version]
- Grayson, W.L.; Bhumiratana, S.; Cannizzaro, C.; Vunjak-Novakovic, G. Bioreactor Cultivation of Functional Bone Grafts. In Mesenchymal Stem Cell Assays and Applications; Humana Press: Totowa, NJ, USA, 2011; Volume 698, pp. 231–241. [Google Scholar] [CrossRef]
- Grayson, W.; Bhumiratana, S.; Chao, P.G.; Hung, C.; Vunjak-Novakovic, G. Spatial regulation of human mesenchymal stem cell differentiation in engineered osteochondral constructs: Effects of pre-differentiation, soluble factors and medium perfusion. Osteoarthr. Cartil. 2010, 18, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Estell, E.G.; Murphy, L.A.; Silverstein, A.M.; Tan, A.R.; Shah, R.P.; Ateshian, G.A.; Hung, C.T. Fibroblast-like synoviocyte mechanosensitivity to fluid shear is modulated by interleukin-1α. J. Biomech. 2017, 60, 91–99. [Google Scholar] [CrossRef]
- Nombela-Arrieta, C.; Ritz, J.; Silberstein, L.E. The elusive nature and function of mesenchymal stem cells. Nat. Rev. Mol. Cell Biol. 2011, 12, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.-T.; Li, W.-J.; Tuan, R.S.; Chang, W.H. Modulation of osteogenesis in human mesenchymal stem cells by specific pulsed electromagnetic field stimulation. J. Orthop. Res. 2009, 27, 1169–1174. [Google Scholar] [CrossRef] [Green Version]
- Esposito, M.; Lucariello, A.; Riccio, I.; Riccio, V.; Esposito, V.; Riccardi, G. Differentiation of human osteoprogenitor cells increases after treatment with pulsed electromagnetic fields. Vivo 2012, 26, 299–304. [Google Scholar]
- Schwartz, Z.; Simon, B.J.; Duran, M.A.; Barabino, G.; Chaudhri, R.; Boyan, B.D. Pulsed electromagnetic fields enhance BMP-2 dependent osteoblastic differentiation of human mesenchymal stem cells. J. Orthop. Res. 2008, 26, 1250–1255. [Google Scholar] [CrossRef] [PubMed]
- Mayer-Wagner, S.; Passberger, A.; Sievers, B.; Aigner, J.; Summer, B.; Schiergens, T.S.; Jansson, V.; Müller, P.E. Effects of low frequency electromagnetic fields on the chondrogenic differentiation of human mesenchymal stem cells. Bioelectromagnetics 2011, 32, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Varani, K.; Vincenzi, F.; Pasquini, S.; Blo, I.; Salati, S.; Cadossi, M.; De Mattei, M. Pulsed Electromagnetic Field Stimulation in Osteogenesis and Chondrogenesis: Signaling Pathways and Therapeutic Implications. Int. J. Mol. Sci. 2021, 22, 809. [Google Scholar] [CrossRef]
- Ongaro, A.; Pellati, A.; Bagheri, L.; Fortini, C.; Setti, S.; De Mattei, M. Pulsed electromagnetic fields stimulate osteogenic differentiation in human bone marrow and adipose tissue derived mesenchymal stem cells. Bioelectromagnetics 2014, 35, 426–436. [Google Scholar] [CrossRef]
- Poh, P.S.P.; Seeliger, C.; Unger, M.; Falldorf, K.; Balmayor, E.R.; Van Griensven, M. Osteogenic Effect and Cell Signaling Activation of Extremely Low-Frequency Pulsed Electromagnetic Fields in Adipose-Derived Mesenchymal Stromal Cells. Stem Cells Int. 2018, 2018, 5402853. [Google Scholar] [CrossRef]
- Brighton, C.T.; Wang, W.; Seldes, R.; Zhang, G.; Pollack, S.R. Signal Transduction in Electrically Stimulated Bone Cells. J. Bone Jt. Surg. 2001, 83, 1514–1523. [Google Scholar] [CrossRef] [Green Version]
- Walleczek, J. Electromagnetic field effects on cells of the immune system: The role of calcium signaling. FASEB J. 1992, 6, 3177–3185. [Google Scholar] [CrossRef]
- Varani, K.; Gessi, S.; Merighi, S.; Iannotta, V.; Cattabriga, E.; Spisani, S.; Cadossi, R.; Borea, P.A. Effect of low frequency electromagnetic fields on A2A adenosine receptors in human neutrophils. Br. J. Pharmacol. 2002, 136, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Varani, K.; Gessi, S.; Merighi, S.; Iannotta, V.; Cattabriga, E.; Pancaldi, C.; Cadossi, R.; Borea, P.A. Alteration of A3 adenosine receptors in human neutrophils and low frequency electromagnetic fields. Biochem. Pharmacol. 2003, 66, 1897–1906. [Google Scholar] [CrossRef]
- Fini, M.; Pagani, S.; Giavaresi, G.; De Mattei, M.; Ongaro, A.; Varani, K.; Vincenzi, F.; Massari, L.; Cadossi, M. Functional tissue engineering in articular cartilage repair: Is there a role for electromagnetic biophysical stimulation? Tissue Eng. Part B Rev. 2013, 19, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Maziarz, A.; Kocan, B.; Bester, M.; Budzik, S.; Cholewa, M.; Ochiya, T.; Banas, A. How electromagnetic fields can influence adult stem cells: Positive and negative impacts. Stem Cell Res. Ther. 2016, 7, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aaron, R.K.; Ciombor, D.M.; Jolly, G. Stimulation of experimental endochondral ossification by low-energy pulsing electromagnetic fields. J. Bone Miner. Res. 2009, 4, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Ciombor, D.M.; Lester, G.; Aaron, R.K.; Neame, P.; Caterson, B. Low frequency EMF regulates chondrocyte differentiation and expression of matrix proteins. J. Orthop. Res. 2002, 20, 40–50. [Google Scholar] [CrossRef]
- Aaron, R.K.; Mc Ciombor, D.K. Acceleration of experimental endochondral ossification by biophysical stimulation of the progenitor cell pool. J. Orthop. Res. 1996, 14, 582–589. [Google Scholar] [CrossRef]
- Aaron, R.K.; Wang, S.; Ciombor, D.M. Upregulation of basal TGFß1 levels by EMF coincident with chondrogenesis—Implications of skeletal repair and tissue engineering. J. Orthop. Res. 2002, 20, 233–240. [Google Scholar] [CrossRef]
- Cadossi, R.; Massari, L.; Racine-Avila, J.; Aaron, R.K. Pulsed Electromagnetic Field Stimulation of Bone Healing and Joint Preservation: Cellular Mechanisms of Skeletal Response. JAAOS Glob. Res. Rev. 2020, 4, e19.00155. [Google Scholar] [CrossRef]
- Aaron, R.K.; Boyan, B.D.; Ciombor, D.M.; Schwartz, Z.; Simon, B.J. Stimulation of Growth Factor Synthesis by Electric and Electromagnetic Fields. Clin. Orthop. Relat. Res. 2004, 419, 30–37. [Google Scholar] [CrossRef]
- Owen-Woods, C.; Kusumbe, A. Fundamentals of bone vasculature: Specialization, interactions and functions. Semin. Cell Dev. Biol. 2021, 123, 36–47. [Google Scholar] [CrossRef]
- Chen, J.; Hendriks, M.; Chatzis, A.; Ramasamy, S.K.; Kusumbe, A.P. Bone Vasculature and Bone Marrow Vascular Niches in Health and Disease. J. Bone Miner. Res. 2020, 35, 2103–2120. [Google Scholar] [CrossRef]
- Ramasamy, S.; Kusumbe, A.P.; Adams, R.H. Regulation of tissue morphogenesis by endothelial cell-derived signals. Trends Cell Biol. 2015, 25, 148–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, L.; Fu, C.; Wang, L.; Zhang, Q.; Liang, Z.; He, C.; Wei, Q. The Effect of Pulsed Electromagnetic Fields on Angiogenesis. Bioelectromagnetics 2021, 42, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Tepper, O.M.; Callaghan, M.J.; Chang, E.I.; Galiano, R.D.; Bhatt, K.A.; Baharestani, S.; Gan, J.; Simon, B.; Hopper, R.A.; Levine, J.P.; et al. Electromagnetic fields increase in vitro and in vivo angiogenesis through endothelial release of FGF-2. FASEB J. 2004, 18, 1231–1233. [Google Scholar] [CrossRef] [Green Version]
- Goto, T.; Fujioka, M.; Ishida, M.; Kuribayashi, M.; Ueshima, K.; Kubo, T. Noninvasive up-regulation of angiopoietin-2 and fibroblast growth factor-2 in bone marrow by pulsed electromagnetic field therapy. J. Orthop. Sci. 2010, 15, 661–665. [Google Scholar] [CrossRef]
- Wang, D.; Wang, J.; Zhou, J.; Zheng, X. The Role of Adenosine Receptor A2A in the Regulation of Macrophage Exosomes and Vascular Endothelial Cells during Bone Healing. J. Inflamm. Res. 2021, 14, 4001–4017. [Google Scholar] [CrossRef] [PubMed]
- Roelofs, A.J.; Kania, K.; Rafipay, A.J.; Sambale, M.; Kuwahara, S.T.; Collins, F.L.; Smeeton, J.; A Serowoky, M.; Rowley, L.; Wang, H.; et al. Identification of the skeletal progenitor cells forming osteophytes in osteoarthritis. Ann. Rheum. Dis. 2020, 79, 1625–1634. [Google Scholar] [CrossRef]
- Roelofs, A.J.; Zupan, J.; Riemen, A.; Kania, K.; Ansboro, S.; White, N.; Clark, S.M.; De Bari, C. Joint morphogenetic cells in the adult mammalian synovium. Nat. Commun. 2017, 8, 15040. [Google Scholar] [CrossRef] [Green Version]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.-W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A Perivascular Origin for Mesenchymal Stem Cells in Multiple Human Organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Caplan, A.I. All MSCs are pericytes? Cell Stem Cell 2008, 3, 229–230. [Google Scholar] [CrossRef] [Green Version]
- Moore, E.M.; Maestas, D.R., Jr.; Comeau, H.Y.; Elisseeff, J.H. The Immune System and Its Contribution to Variability in Regenerative Medicine. Tissue Eng. Part B Rev. 2021, 27, 39–47. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stucky, E.C.; Schloss, R.S.; Yarmush, M.L.; Shreiber, D.I. Alginate micro-encapsulation of mesenchymal stromal cells enhances modulation of the neuro-inflammatory response. Cytotherapy 2015, 17, 1353–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Chiu, S.M.; Motan, D.A.L.; Zhang, Z.; Chen, L.; Ji, H.-L.; Tse, H.-F.; Fu, Q.-L.; Lian, Q. Mesenchymal stem cells and immunomodulation: Current status and future prospects. Cell Death Dis. 2016, 7, e2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, J.R.; Teixeira, G.Q.; Santos, S.G.; Barbosa, M.A.; Almeida-Porada, G.; Goncalves, R.M. Mesenchymal Stromal Cell Secretome: Influencing Therapeutic Potential by Cellular Pre-conditioning. Front. Immunol. 2018, 9, 2837. [Google Scholar] [CrossRef]
- Vizoso, F.J.; Eiro, N.; Costa, L.; Esparza, P.; Landin, M.; Diaz-Rodriguez, P.; Schneider, J.; Pérez, L.C. Mesenchymal Stem Cells in Homeostasis and Systemic Diseases: Hypothesis, Evidences, and Therapeutic Opportunities. Int. J. Mol. Sci. 2019, 20, 3738. [Google Scholar] [CrossRef] [Green Version]
- Madrigal, M.; Rao, K.S.; Riordan, N.H. A review of therapeutic effects of mesenchymal stem cell secretions and induction of secretory modification by different culture methods. J. Transl. Med. 2014, 12, 260. [Google Scholar] [CrossRef] [Green Version]
- Gazdic, M.; Volarevic, V.; Arsenijevic, N.; Stojkovic, M. Mesenchymal Stem Cells: A Friend or Foe in Immune-Mediated Diseases. Stem Cell Rev. Rep. 2015, 11, 280–287. [Google Scholar] [CrossRef]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal Stem Cell Secretome: Toward Cell-Free Therapeutic Strategies in Regenerative Medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef] [Green Version]
- Maguire, G. Stem cell therapy without the cells. Commun. Integr. Biol. 2013, 6, e26631. [Google Scholar] [CrossRef]
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal stem cells in health and disease. Nat. Rev. Immunol. 2008, 8, 726–736. [Google Scholar] [CrossRef]
- Ha, C.-W.; Park, Y.-B.; Kim, S.H.; Lee, H.-J. Intra-articular Mesenchymal Stem Cells in Osteoarthritis of the Knee: A Systematic Review of Clinical Outcomes and Evidence of Cartilage Repair. Arthrosc. J. Arthrosc. Relat. Surg. 2019, 35, 277–288.e2. [Google Scholar] [CrossRef]
- Bianchi, F.; Maioli, M.; Leonardi, E.; Olivi, E.; Pasquinelli, G.; Valente, S.; Mendez, A.J.; Ricordi, C.; Raffaini, M.; Tremolada, C.; et al. A New Nonenzymatic Method and Device to Obtain a Fat Tissue Derivative Highly Enriched in Pericyte-Like Elements by Mild Mechanical Forces from Human Lipoaspirates. Cell Transplant. 2013, 22, 2063–2077. [Google Scholar] [CrossRef]
- Jones, I.A.; Wilson, M.; Togashi, R.; Han, B.; Mircheff, A.K.; Vangsness, T., Jr. A randomized, controlled study to evaluate the efficacy of intra-articular, autologous adipose tissue injections for the treatment of mild-to-moderate knee osteoarthritis compared to hyaluronic acid: A study protocol. BMC Musculoskelet. Disord. 2018, 19, 383. [Google Scholar] [CrossRef]
- McIntyre, J.A.; Jones, I.; Danilkovich, A.; Vangsness, C.T. The Placenta: Applications in Orthopaedic Sports Medicine. Am. J. Sports Med. 2018, 46, 234–247. [Google Scholar] [CrossRef]
- Tan, A.R.; Hung, C.T. Concise Review: Mesenchymal Stem Cells for Functional Cartilage Tissue Engineering: Taking Cues from Chondrocyte-Based Constructs. Stem Cells Transl. Med. 2017, 6, 1295–1303. [Google Scholar] [CrossRef]
- Nixon, A.J.; Rickey, E.; Butler, T.J.; Scimeca, M.S.; Moran, N.; Matthews, G.L. A chondrocyte infiltrated collagen type I/III membrane (MACI(R) implant) improves cartilage healing in the equine patellofemoral joint model. Osteoarthr. Cartil. 2015, 23, 648–660. [Google Scholar] [CrossRef] [Green Version]
- FDA. FDA Approves First Autologous Cellularized Scaffold for the Repair of Cartilage Defects of the Knee; FDA: Silver Spring, MD, USA, 2016.
- Chan, B.P.; Leong, K.W. Scaffolding in tissue engineering: General approaches and tissue-specific considerations. Eur. Spine J. 2008, 17, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Cook, J.; Mendelson, A.; Moioli, E.K.; Yao, H.; Mao, J.J. Regeneration of the articular surface of the rabbit synovial joint by cell homing: A proof of concept study. Lancet 2010, 376, 440–448. [Google Scholar] [CrossRef] [Green Version]
- Brunger, J.M.; Huynh, N.P.T.; Guenther, C.M.; Perez-Pinera, P.; Moutos, F.T.; Sanchez-Adams, J.; Gersbach, C.A.; Guilak, F. Scaffold-mediated lentiviral transduction for functional tissue engineering of cartilage. Proc. Natl. Acad. Sci. USA 2014, 111, E798–E806. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.R.; Rodeo, S.; Bhutani, N.; Goodrich, L.R.; Huard, J.; Irrgang, J.; LaPrade, R.F.; Lattermann, C.; Lu, Y.; Mandelbaum, B.; et al. Optimizing Clinical Use of Biologics in Orthopaedic Surgery: Consensus Recommendations from the 2018 AAOS/NIH U-13 Conference. J. Am. Acad. Orthop. Surg. 2019, 27, e50–e63. [Google Scholar] [CrossRef]
- AAOS. Available online: https://www.aaos.org/globalassets/advocacy/issues/aaos-letter-to-the-fda-on-biologics-regulation_final_signed_111521.pdf (accessed on 19 March 2022).
- Shive, M.S.; Hoemann, C.D.; Restrepo, A.; Hurtig, M.B.; Duval, N.; Ranger, P.; Stanish, W.; Buschmann, M.D. BST-CarGel: In Situ ChondroInduction for Cartilage Repair. Oper. Tech. Orthop. 2006, 16, 271–278. [Google Scholar] [CrossRef]
- Pot, M.W.; Gonzales, V.K.; Buma, P.; IntHout, J.; Van Kuppevelt, T.H.; De Vries, R.B.; Daamen, W.F. Improved cartilage regeneration by implantation of acellular biomaterials after bone marrow stimulation: A systematic review and meta-analysis of animal studies. PeerJ 2016, 4, e2243. [Google Scholar] [CrossRef]
- Sumant, U.; Rupali, S. Bone Growth Stimulators Market Size by Type: Bone Growth Stimulation Devices GMI1904. 2021. Available online: https://www.gminsights.com/industry-analysis/bone-growth-stimulators-market (accessed on 23 March 2022).
- Guo, X.E.; Hung, C.T.; Sandell, L.J.; Silva, M.J. Musculoskeletal mechanobiology: A new era for MechanoMedicine. J. Orthop. Res. 2018, 36, 531–532. [Google Scholar] [CrossRef] [Green Version]
- Glatt, V.; Evans, C.H.; Stoddart, M.J. Regenerative rehabilitation: The role of mechanotransduction in orthopaedic regenerative medicine. J. Orthop. Res. 2019, 37, 1263–1269. [Google Scholar] [CrossRef]
- O’Conor, C.J.; Leddy, H.A.; Benefield, H.C.; Liedtke, W.B.; Guilak, F. TRPV4-mediated mechanotransduction regulates the metabolic response of chondrocytes to dynamic loading. Proc. Natl. Acad. Sci. USA 2014, 111, 1316–1321. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
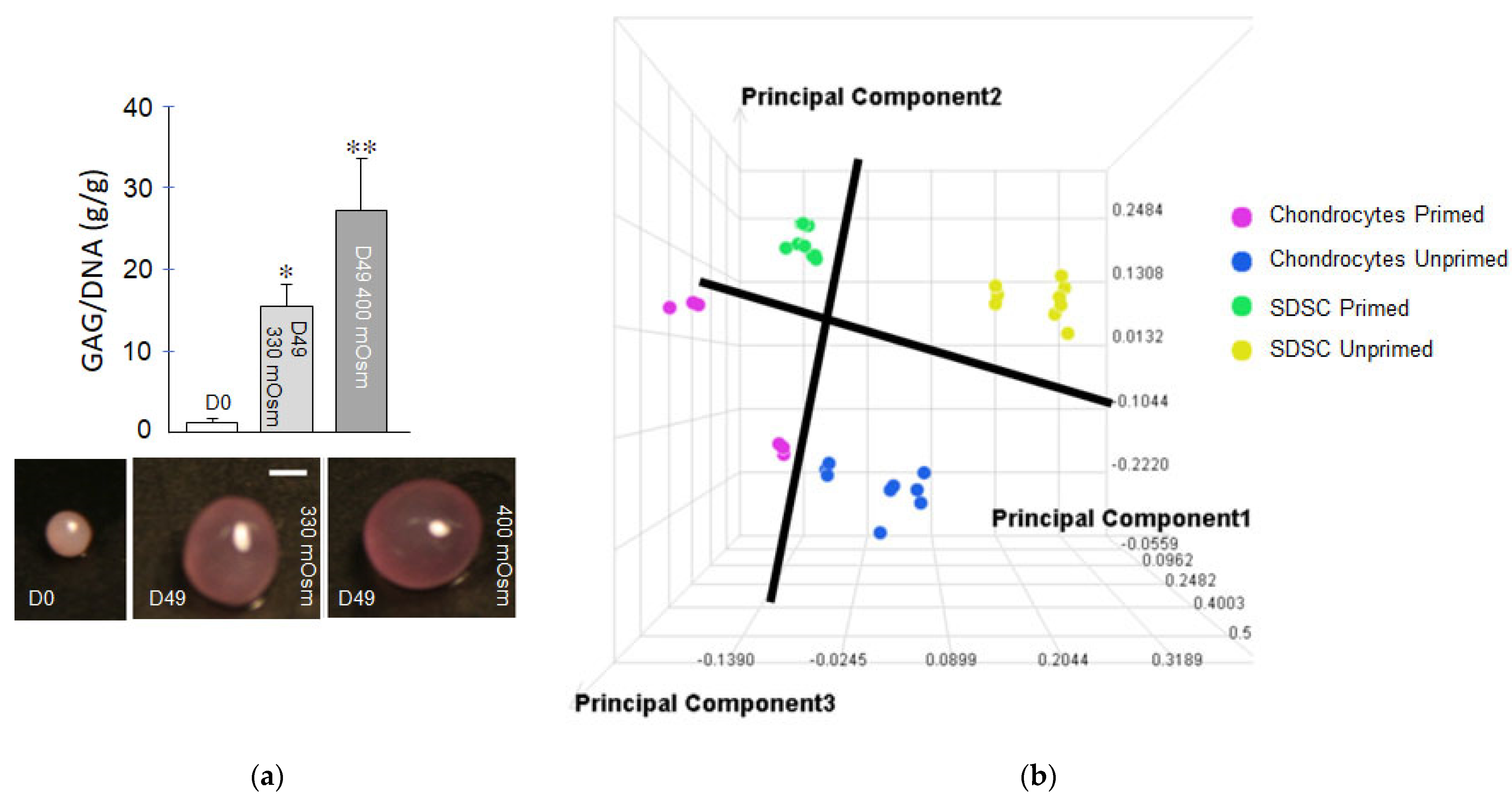{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
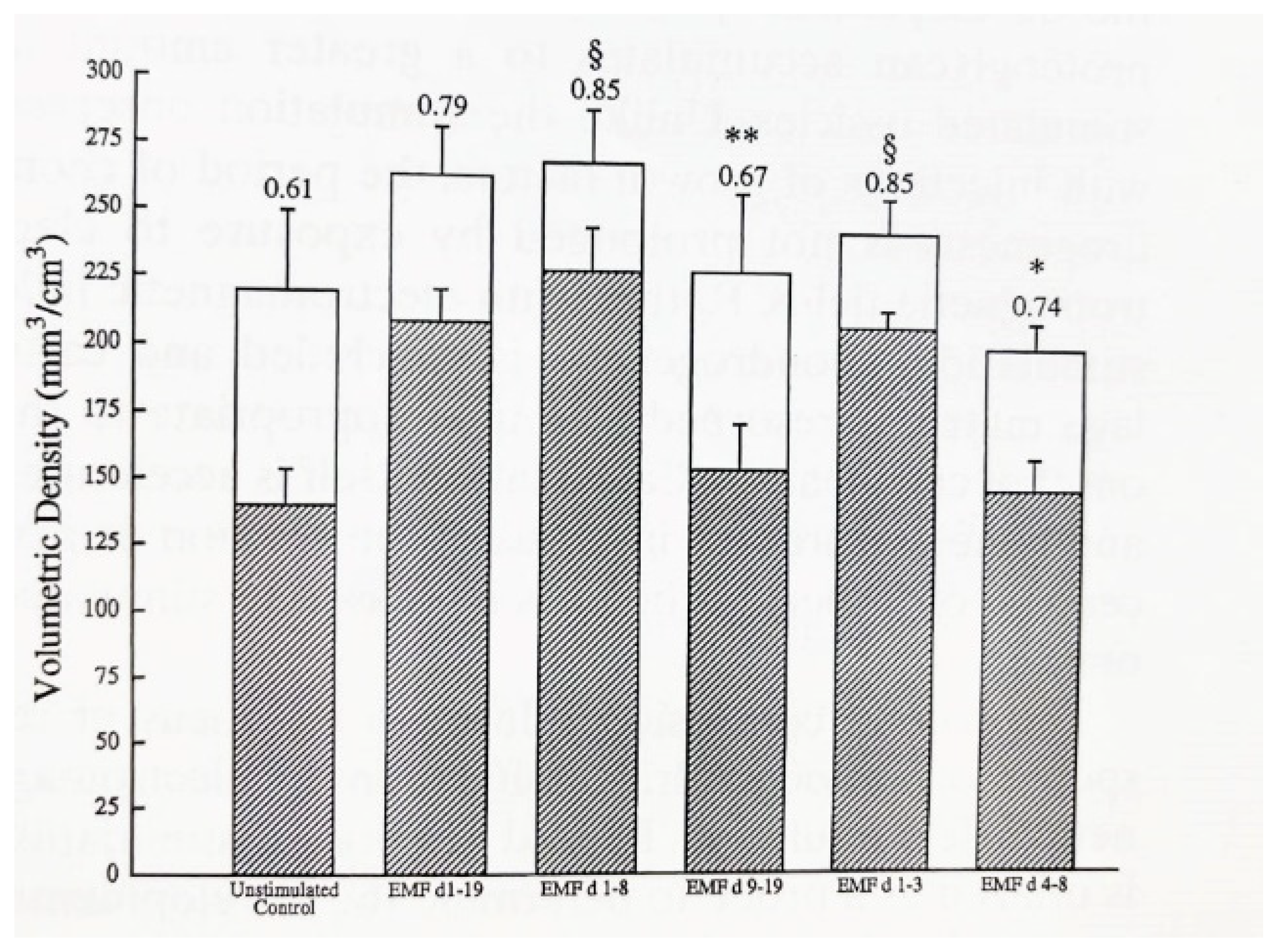{kind=link}
| Control | PEMF | Percent | p | |
|---|---|---|---|---|
| mRNA aggrecan | 6.1 | 22.5 | 269 | 0.02 |
| mRNA type II collagen | 11.8 | 21.9 | 86 | 0.05 |
| 35SO4 incorporation (cpm/mg) | 2166 ± 387 | 4448 ± 293 | 105 | 0.005 |
| GAG content (µg/mg) | 1.4 ± 0.2 | 2.5 ± 0.2 | 79 | 0.01 |
| Cartilage area (mm2) | 24 ± 2.1 | 148 ± 11.7 | 517 | 0.001 |
| Chondrocytes (n) | 701 ± 227 | 3582 ± 675 | 411 | 0.005 |
| Chondrocytes/cartilage | 29.2 | 24.2 | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hung, C.T.; Racine-Avila, J.; Pellicore, M.J.; Aaron, R. Biophysical Modulation of Mesenchymal Stem Cell Differentiation in the Context of Skeletal Repair. Int. J. Mol. Sci. 2022, 23, 3919. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073919
Hung CT, Racine-Avila J, Pellicore MJ, Aaron R. Biophysical Modulation of Mesenchymal Stem Cell Differentiation in the Context of Skeletal Repair. International Journal of Molecular Sciences. 2022; 23(7):3919. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073919
Chicago/Turabian StyleHung, Clark T., Jennifer Racine-Avila, Matthew J. Pellicore, and Roy Aaron. 2022. "Biophysical Modulation of Mesenchymal Stem Cell Differentiation in the Context of Skeletal Repair" International Journal of Molecular Sciences 23, no. 7: 3919. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073919