
Changes in the Cardiotoxic Effects of Lead Intoxication in Rats Induced by Muscular Exercise

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Development of Lead Intoxication and the Effect of Muscle Exercise on It
2.2. Biochemical and Systemic Physiological Indices Associated with the Condition of the Heart, Blood Vessels and Breathing Function
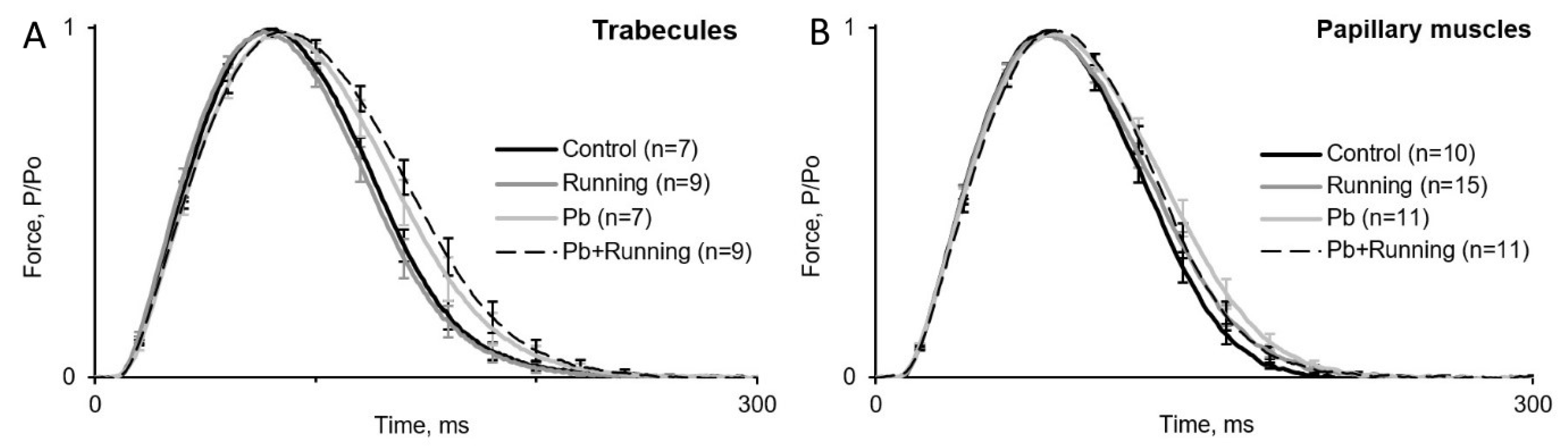
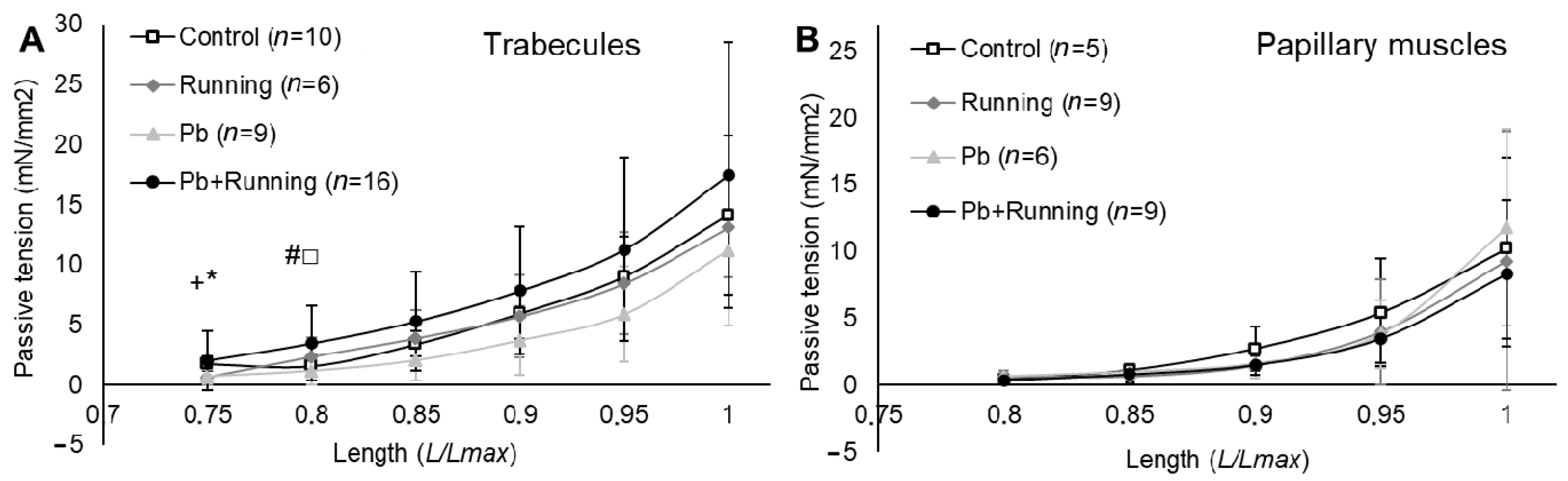
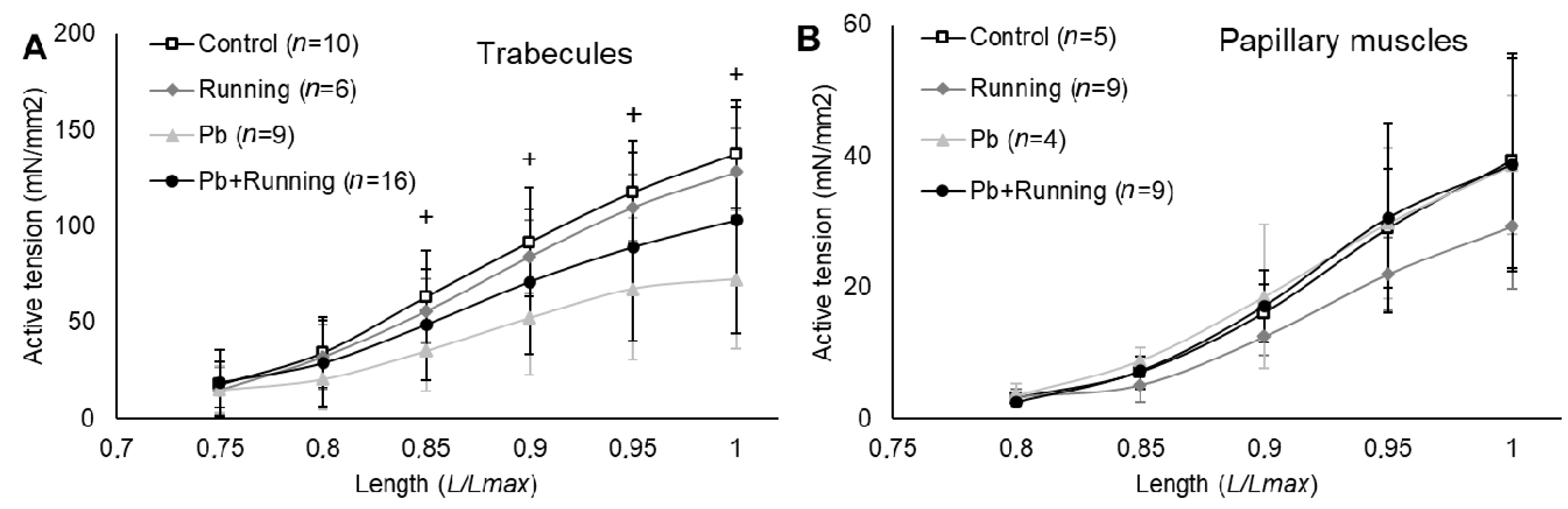
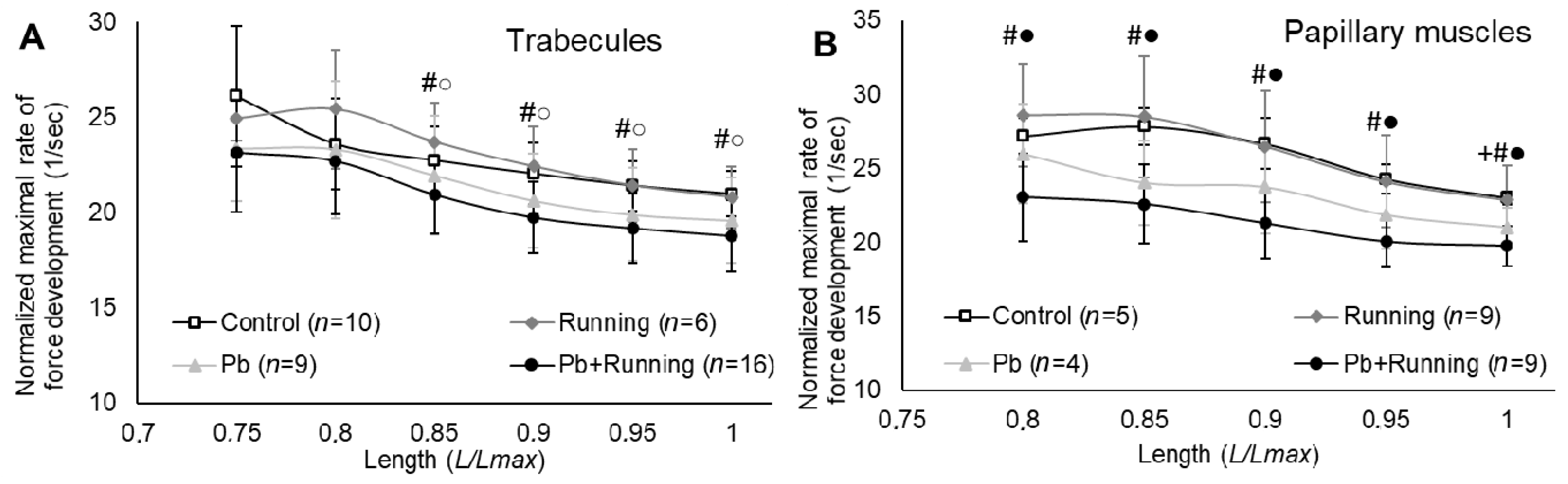
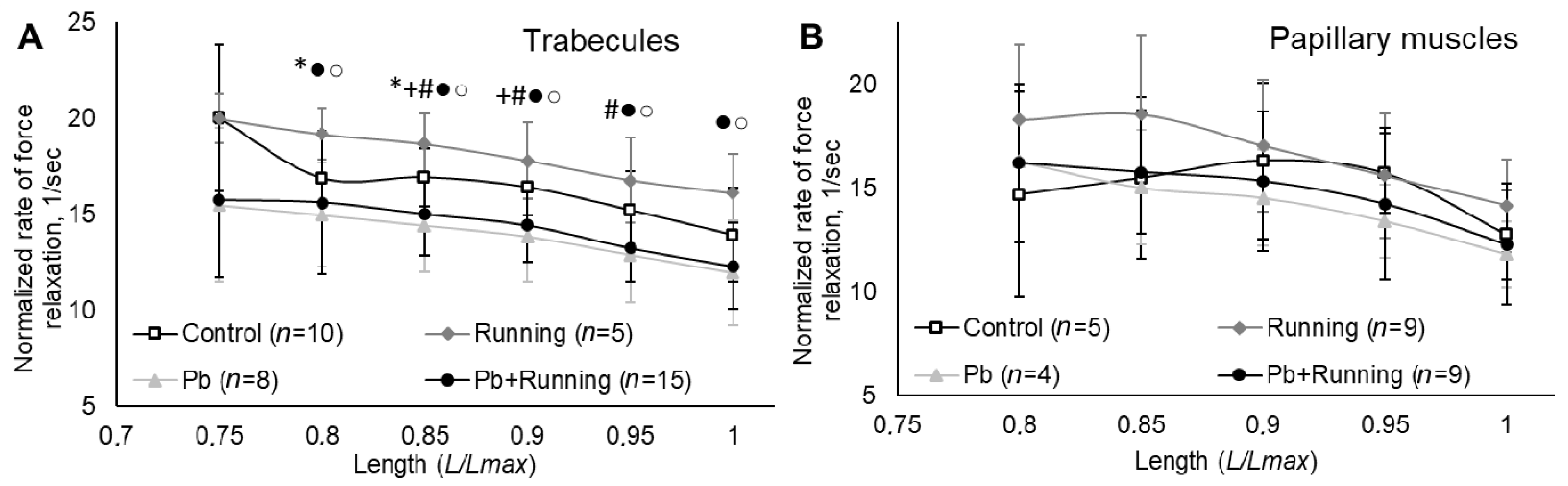
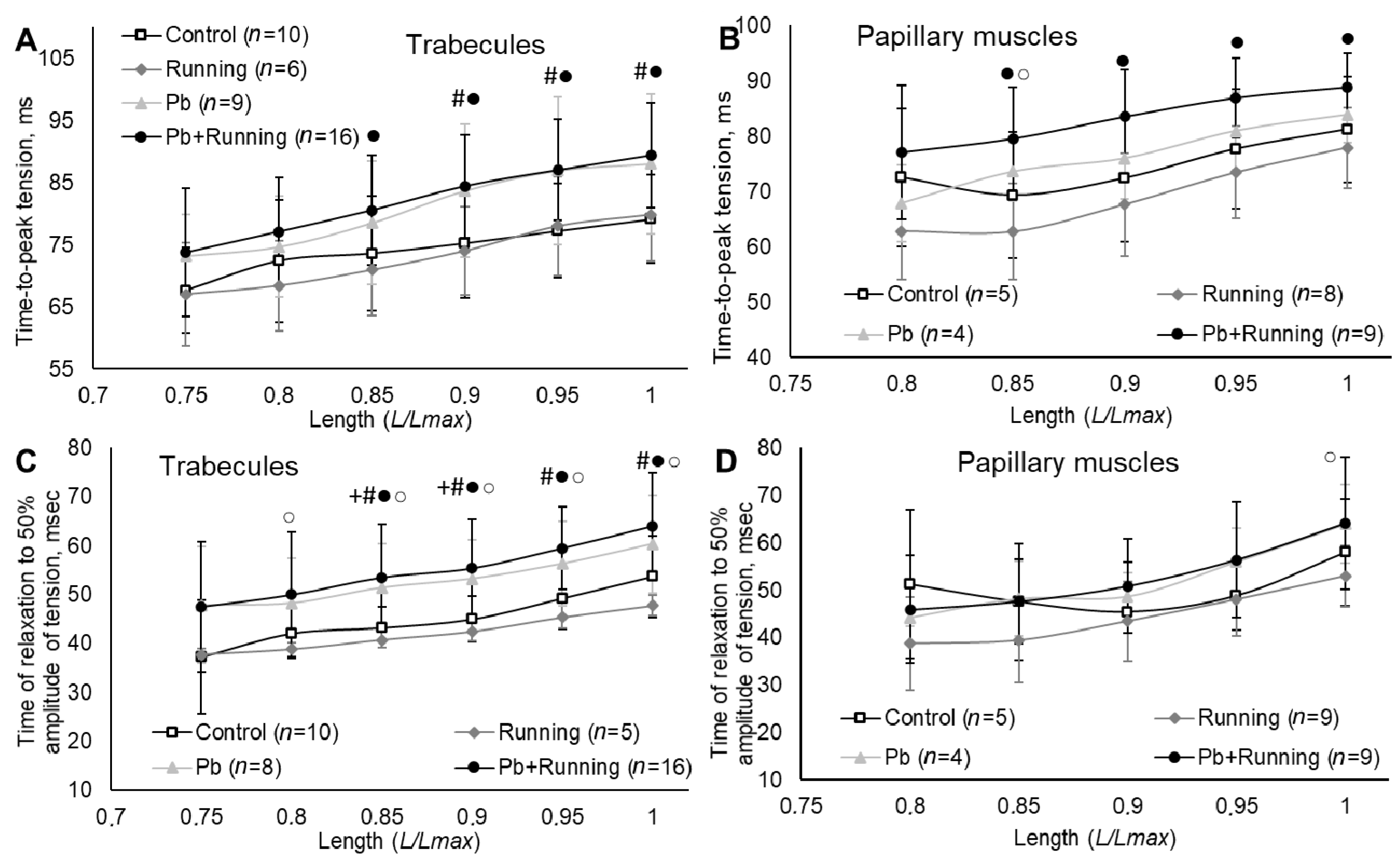
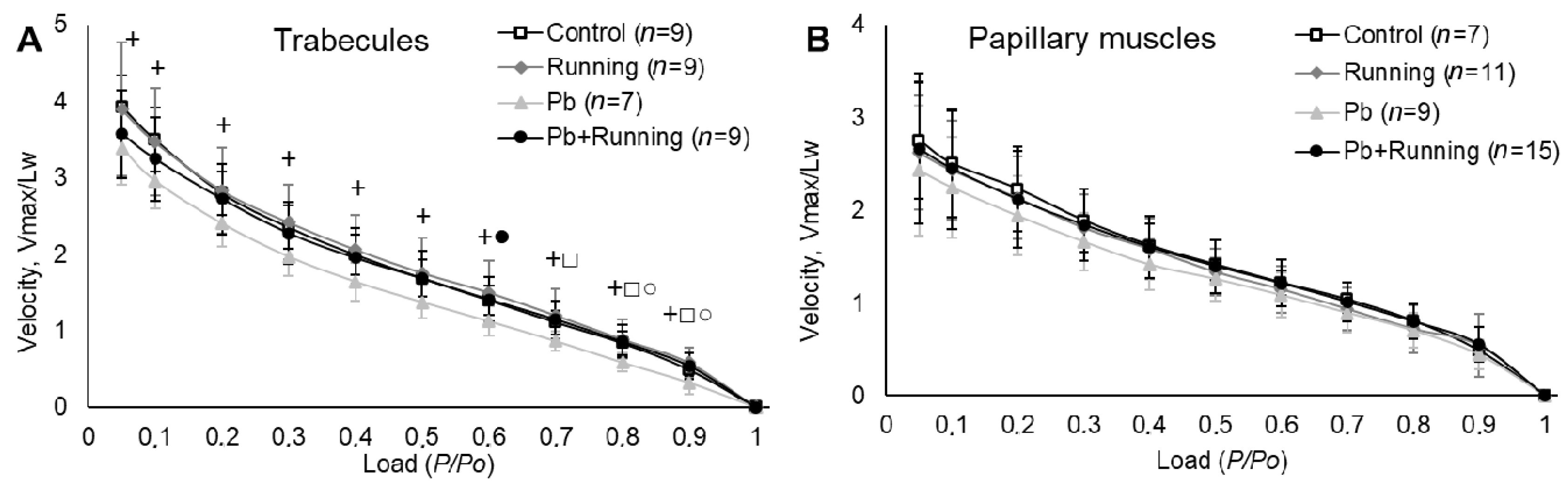
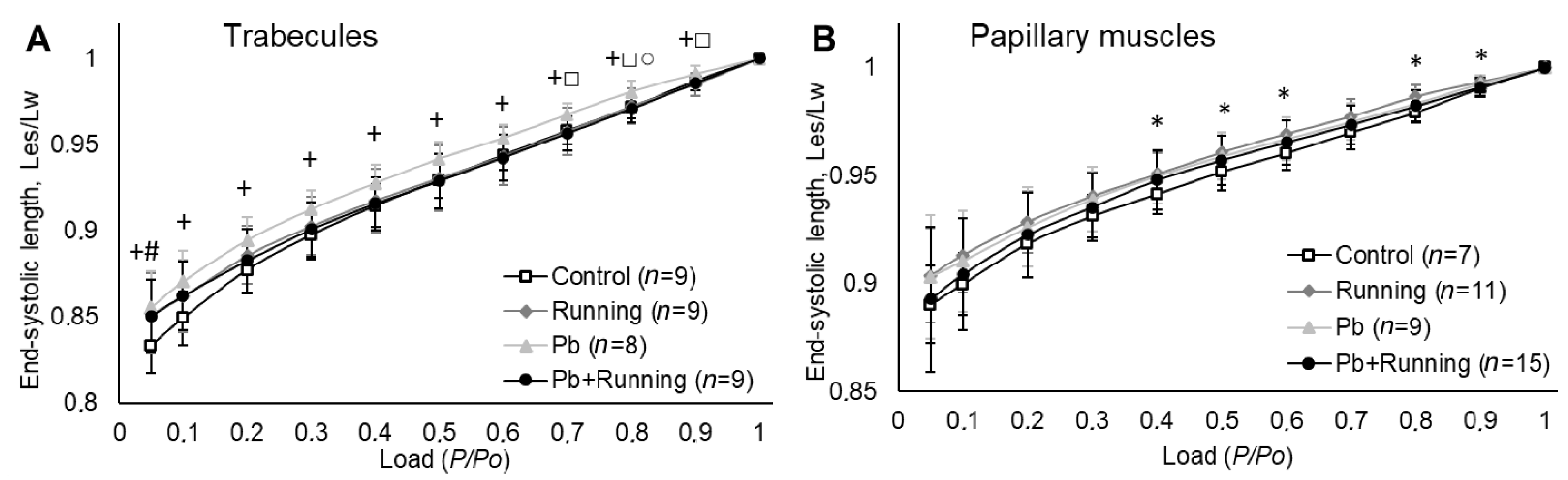
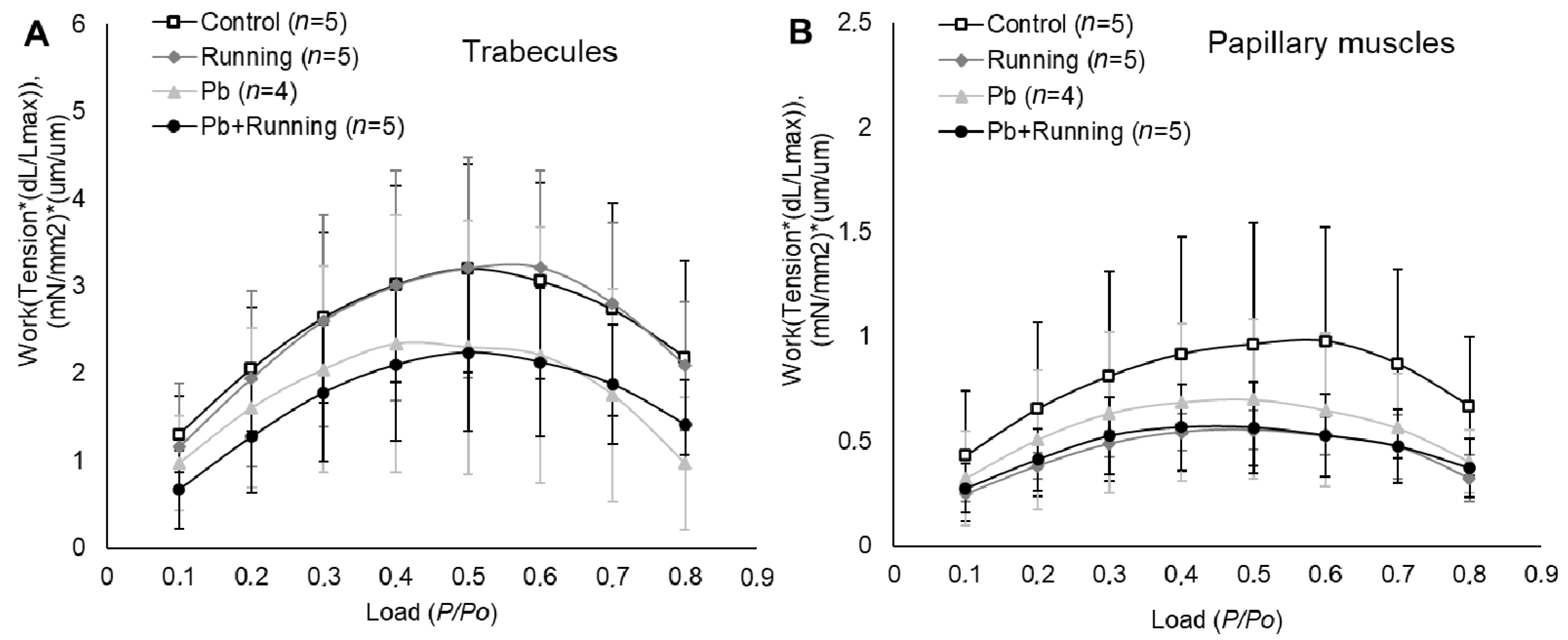
2.3. Contractility of Isolated Myocardial Preparations
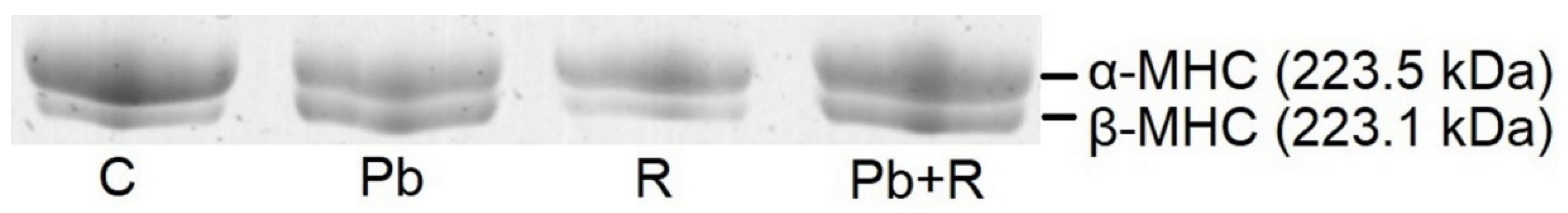
2.4. Myosins
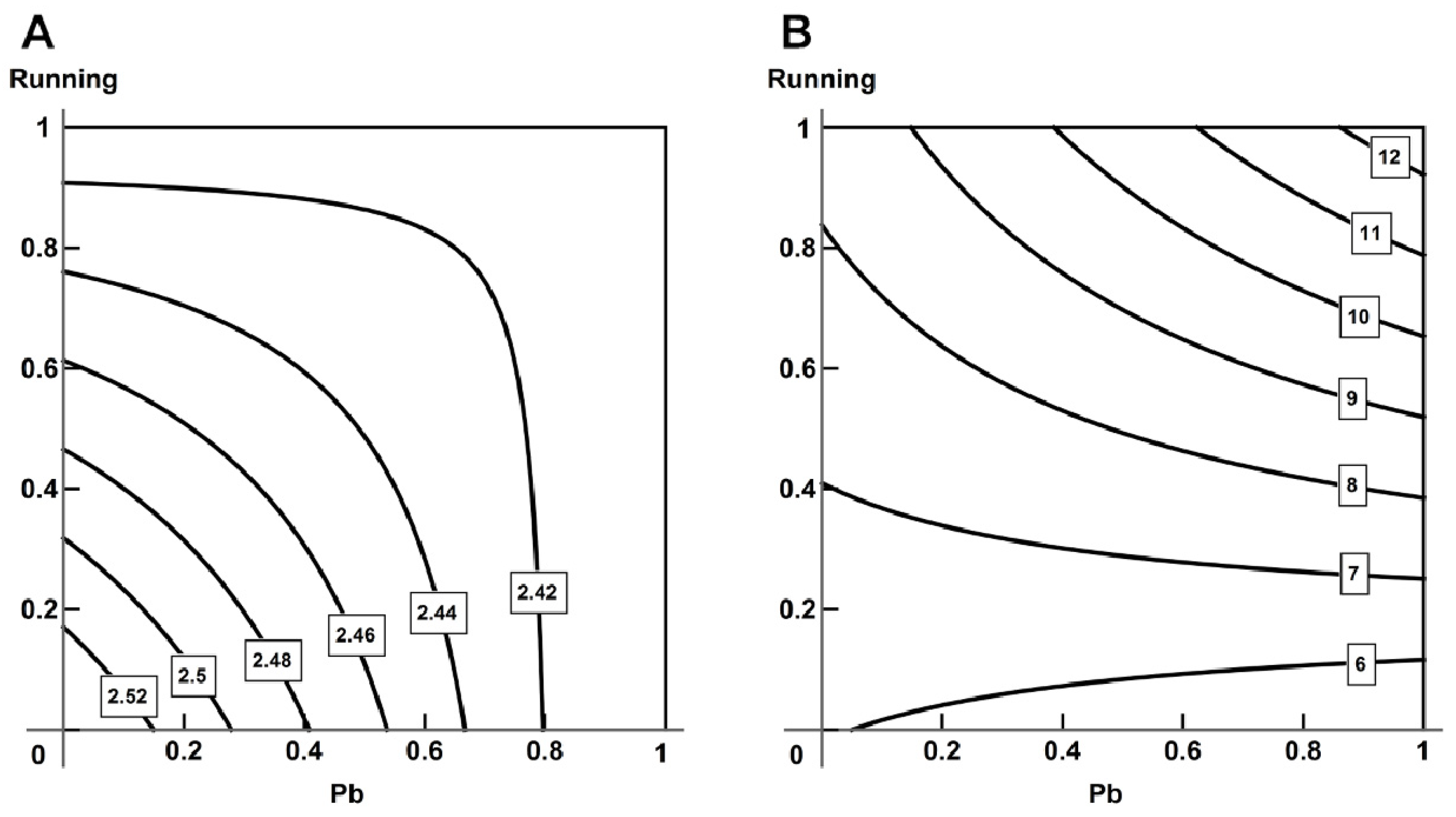
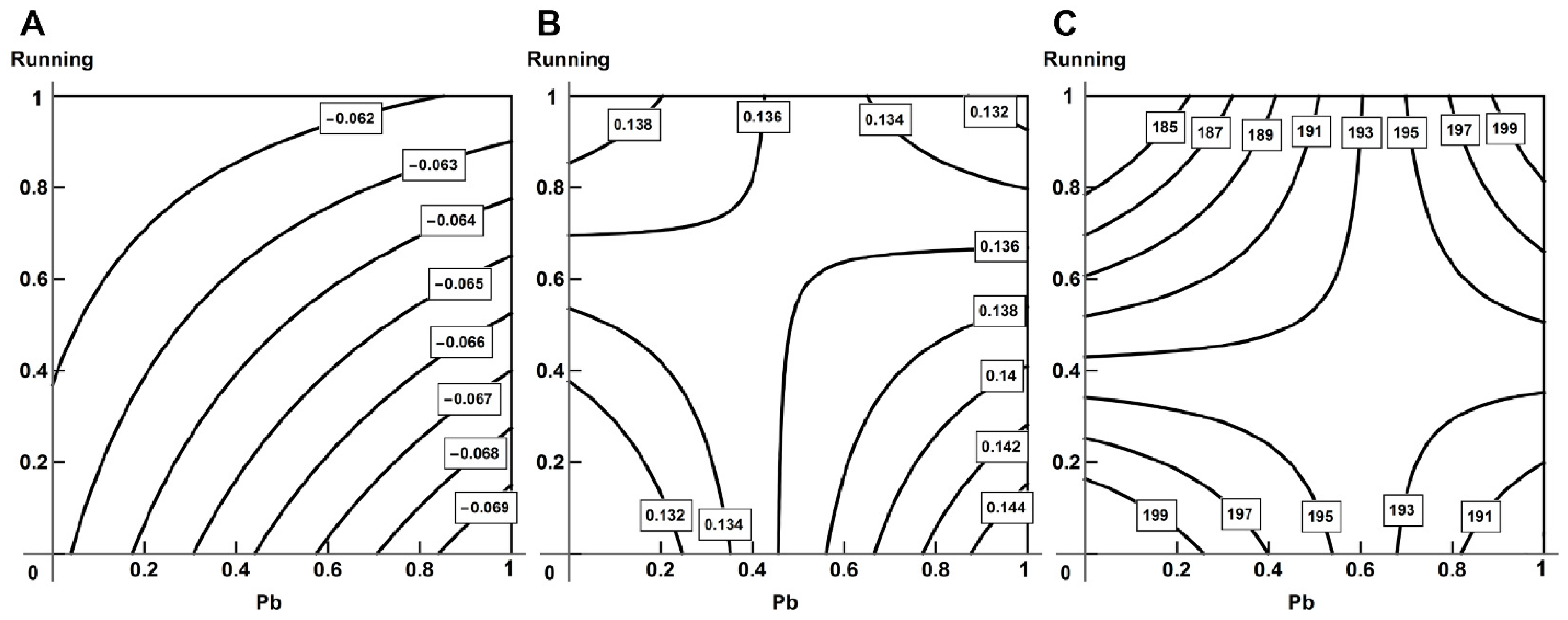
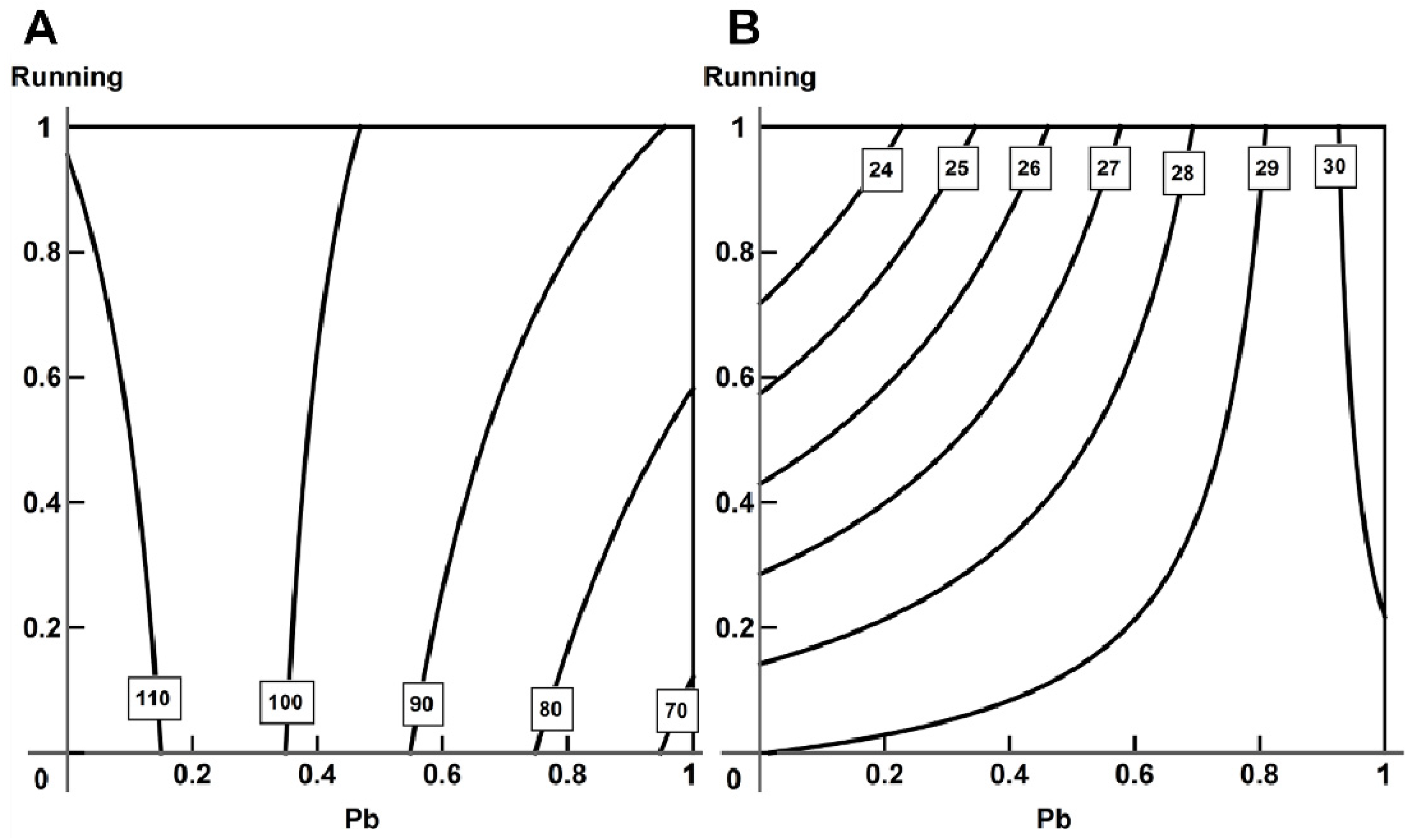
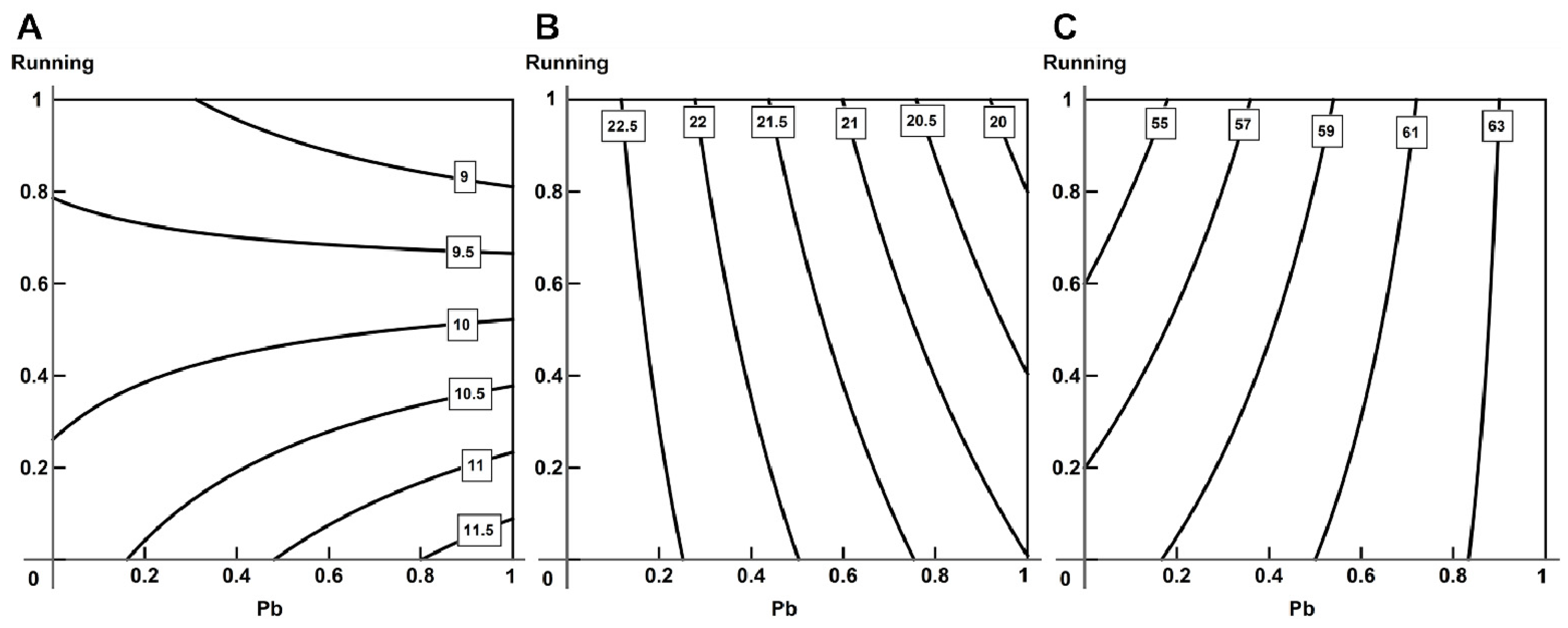
2.5. Analysis of Combined Action with the Help of RSM
3. Discussion
4. Materials and Methods
4.1. Experimental Animals, Exposures and Toxicological Indices
4.2. Determination of α– and β–Cardiac Myosin Heavy Chain (MHC) Ratio
4.3. Assessment of the Mechanical Characteristics of Actin–Myosin Interaction by In Vitro Motility Assay
4.4. Measuring the Contractile Response of Isolated Myocardial Preparations
4.5. Mathematical Processing and Analysis
5. Conclusions
- Under moderate subchronic intoxication with lead, we observed the following:
- 2.
- In the context of moderate lead intoxication in rats, the repeated short-term exercise applied in this experiment generally produced a positive effect on the cardiovascular system, as follows:
- 3.
- Using the response surface methodology, we confirmed the typological diversity of combined actions produced by lead and physical load on the cardiovascular system of rats, depending on the effect by which it is assessed, the ratio of doses and the type of object studied.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Katsnelson, B.A.; Polzik, E.V.; Privalova, L.I. Some aspects of the problem of individual predisposition to silicosis. Environ. Health Perspect. 1986, 68, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Katsnelson, B. Some mechanisms of combined action playing a role in the pathogenesis of silicosis. In The Combined Effect of Chemical and Physical Factors of the Industrial Environment; Publishing House of Labor Hygiene: Sverdlovsk, Russia, 1972; pp. 1–10. [Google Scholar]
- UNEP. Key Scientific Findings for Lead; UNEP: Nairobi, Kenya, 2010. [Google Scholar]
- ITG. Information and Technical Guide to the Best Available Technologies. ITS 13-2020 Production of Lead, Zinc and Cadmium; Bureau NTD: Moskow, Russia, 2020. [Google Scholar]
- State Report on the State of Sanitary and Epidemiological Well-Being of the Population in the Russian Federation in 2019; Federal Service for Supervision of Consumer Rights Protection and Human Welfare: Moskow, Russia, 2020.
- Alissa, E.M.; Ferns, G.A. Heavy metal poisoning and cardiovascular disease. J. Toxicol. 2011, 2011, 870125. [Google Scholar] [CrossRef] [PubMed]
- Solenkova, N.V.; Newman, J.D.; Berger, J.S.; Thurston, G.; Hochman, J.S.; Lamas, G.A. Metal pollutants and cardiovascular disease: Mechanisms and consequences of exposure. Am. Heart J. 2014, 168, 812–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, G.A.; Navas-Acien, A.; Mark, D.B.; Lee, K.L. Heavy metals, cardiovascular disease, and the unexpected benefits of chelation therapy. J. Am. Coll. Cardiol. 2016, 67, 2411–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.Y.; Zhang, Z.Y.; Thijs, L.; Cauwenberghs, N.; Wei, F.F.; Jacobs, L.; Luttun, A.; Verhamme, P.; Kuznetsova, T.; Nawrot, T.S.; et al. Left ventricular structure and function in relation to environmental exposure to lead and cadmium. J. Am. Heart Assoc. 2017, 6, e004692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GBD Compare|IHME Viz Hub. Available online: https://vizhub.healthdata.org/gbd-compare/ (accessed on 28 December 2020).
- Maynar-Mariño, M.; Llerena, F.; Bartolomé, I.; Crespo, C.; Muñoz, D.; Robles, M.C.; Caballero, M.J. Effect of long-term aerobic, anaerobic and aerobic-anaerobic physical training in seric toxic minerals concentrations. J. Trace Elem. Med. Biol. 2018, 45, 136–141. [Google Scholar] [CrossRef] [PubMed]
- LLerena, F.; Maynar, M.; Barrientos, G.; Palomo, R.; Robles, M.C.; Caballero, M.J. Comparison of urine toxic metals concentrations in athletes and in sedentary subjects living in the same area of Extremadura (Spain). Eur. J. Appl. Physiol. 2012, 112, 3027–3031. [Google Scholar] [CrossRef]
- Roshan, V.D.; Assali, M.; Moghaddam, A.H.; Hosseinzadeh, M.; Myers, J. Exercise training and antioxidants: Effects on rat heart tissue exposed to lead acetate. Int. J. Toxicol. 2011, 30, 190–196. [Google Scholar] [CrossRef]
- Shahandeh, M.; Roshan, V.D.; Hosseinzadeh, S.; Mahjoub, S.; Sarkisian, V. Chronic exercise training versus acute endurance exercise in reducing neurotoxicity in rats exposed to lead acetate. Neural Regen. Res. 2013, 8, 714–722. [Google Scholar] [CrossRef]
- Abadin, H.; Ashizawa, A.; Stevens, Y.-W.; Llados, F.; Diamond, G.; Sage, G.; Citra, M.; Quinones, A.; Bosch, S.J.; Swarts, S.G. Toxicological Profile for Lead; Syracuse Research Corporation: Syracuse, NY, USA, 2020; 582p. [Google Scholar]
- Rius, R.A.; Govoni, S.; Trabucchi, M. Regional modification of brain calcium antagonist binding after in vivo chronic lead exposure. Toxicology 1986, 40, 191–197. [Google Scholar] [CrossRef]
- Savchenko, O.V.; Sgrebneva, M.N.; Kiselev, V.I.; Khotimchenko, Y.S. Lead removal in rats using calcium alginate. Environ. Sci. Pollut. Res. 2014, 22, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Kohrt, W.M.; Wherry, S.J.; Wolfe, P.; Sherk, V.D.; Wellington, T.; Swanson, C.M.; Weaver, C.M.; Boxer, R.S. Maintenance of Serum Ionized Calcium During Exercise Attenuates Parathyroid Hormone and Bone Resorption Responses. J. Bone Miner. Res. 2018, 33, 1326–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardi, G.; Ziemann, E.; Banfi, G.; Corbetta, S. Physical activity-dependent regulation of parathyroid hormone and calcium-phosphorous metabolism. Int. J. Mol. Sci. 2020, 21, 5388. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N. Role of vascular endothelial growth factor in the regulation of angiogenesis. Kidney Int. 1999, 56, 794–814. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Saha, K.; Hypoxia, S. Lead Toxicities and Oxidative Stress: Cell Signaling, Molecular Interactions and Antioxidant (Vitamin C) Defense. Curr. Signal Transduct. Ther. 2015, 9, 113–122. [Google Scholar] [CrossRef]
- Fiorim, J.; Ribeiro Junior, R.F.; Silveira, E.A.; Padilha, A.S.; Vescovi, M.V.A.; de Jesus, H.C.; Stefanon, I.; Salaices, M.; Vassallo, D.V. Low-Level Lead Exposure Increases Systolic Arterial Pressure and Endothelium-Derived Vasodilator Factors in Rat Aortas. PLoS ONE 2011, 6, e17117. [Google Scholar] [CrossRef] [Green Version]
- Silveira, E.A.; Siman, F.D.M.; De Oliveira Faria, T.; Vescovi, M.V.A.; Furieri, L.B.; Lizardo, J.H.F.; Stefanon, I.; Padilha, A.S.; Vassallo, D.V. Low-dose chronic lead exposure increases systolic arterial pressure and vascular reactivity of rat aortas. Free Radic. Biol. Med. 2014, 67, 366–376. [Google Scholar] [CrossRef]
- Krüger, M.; Linke, W.A. Titin-based mechanical signalling in normal and failing myocardium. J. Mol. Cell. Cardiol. 2009, 46, 490–498. [Google Scholar] [CrossRef]
- Allen, D.G.; Kentish, J.C. The cellular basis of the length-tension relation in cardiac muscle. J. Mol. Cell. Cardiol. 1985, 17, 821–840. [Google Scholar] [CrossRef]
- de Tombe, P.P.; ter Keurs, H.E.D.J. Cardiac muscle mechanics: Sarcomere length matters. J. Mol. Cell. Cardiol. 2016, 91, 148–150. [Google Scholar] [CrossRef] [Green Version]
- Fujita, H.; Labeit, D.; Gerull, B.; Labeit, S.; Granzier, H.L. Titin isoform-dependent effect of calcium on passive myocardial tension. Am. J. Physiol.-Heart Circ. Physiol. 2004, 287, 2528–2534. [Google Scholar] [CrossRef] [PubMed]
- Stuyvers, B.D.; Miura, M.; Jin, J.P.; Ter Keurs, H.E.D.J. Ca2+-dependence of diastolic properties of cardiac sarcomeres: Involvement of titin. Prog. Biophys. Mol. Biol. 1998, 69, 425–443. [Google Scholar] [CrossRef]
- Linke, W.A. Titin Gene and Protein Functions in Passive and Active Muscle. Annu. Rev. Physiol. 2018, 80, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Vassallo, D.V.; Lebarch, E.C.; Moreira, C.M.; Wiggers, G.A.; Stefanon, I. Lead reduces tension development and the myosin ATPase activity of the rat right ventricular myocardium. Brazilian J. Med. Biol. Res. 2008, 41, 789–795. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.A.S.C.; de Oliveira, T.F.; Almenara, C.C.P.; Broseghini-Filho, G.B.; Vassallo, D.V.; Padilha, A.S.; Silveira, E.A. Exposure to a Low Lead Concentration Impairs Contractile Machinery in Rat Cardiac Muscle. Biol. Trace Elem. Res. 2015, 167, 280–287. [Google Scholar] [CrossRef]
- Carmignani, M.; Boscolo, P.; Poma, A.; Volpe, A.R. Kininergic system and arterial hypertension following chronic exposure to inorganic lead. Immunopharmacology 1999, 44, 105–110. [Google Scholar] [CrossRef]
- Carmignani, M.; Volpe, A.R.; Boscolo, P.; Qiao, N.; Di Gioacchino, M.; Grilli, A.; Felaco, M. Catcholamine and nitric oxide systems as targets of chronic lead exposure in inducing selective functional impairment. Life Sci. 2000, 68, 401–415. [Google Scholar] [CrossRef]
- Lookin, O.; Balakin, A.; Kuznetsov, D.; Protsenko, Y. The length-dependent activation of contraction is equally impaired in impuberal male and female rats in monocrotaline-induced right ventricular failure. Clin. Exp. Pharmacol. Physiol. 2015, 42, 1198–1206. [Google Scholar] [CrossRef]
- Protsenko, Y.L.; Katsnelson, B.A.; Klinova, S.V.; Lookin, O.N.; Balakin, A.A.; Nikitina, L.V.; Gerzen, O.P.; Minigalieva, I.A.; Privalova, L.I.; Gurvich, V.B.; et al. Effects of subchronic lead intoxication of rats on the myocardium contractility. Food Chem. Toxicol. 2018, 120, 378–389. [Google Scholar] [CrossRef]
- Protsenko, Y.L.; Klinova, S.V.; Gerzen, O.P.; Privalova, L.I.; Minigalieva, I.A.; Balakin, A.A.; Lookin, O.N.; Lisin, R.V.; Butova, K.A.; Nabiev, S.R.; et al. Changes in rat myocardium contractility under subchronic intoxication with lead and cadmium salts administered alone or in combination. Toxicol. Rep. 2020, 7, 433–442. [Google Scholar] [CrossRef]
- Klinova, S.V.; Minigalieva, I.A.; Privalova, L.I.; Valamina, I.E.; Makeyev, O.H.; Shuman, E.A.; Korotkov, A.A.; Panov, V.G.; Sutunkova, M.P.; Ryabova, J.V.; et al. Further verification of some postulates of the combined toxicity theory: New animal experimental data on separate and joint adverse effects of lead and cadmium. Food Chem. Toxicol. 2020, 136, 110971. [Google Scholar] [CrossRef] [PubMed]
- Protsenko, Y.L.; Katsnelson, B.A.; Klinova, S.V.; Lookin, O.N.; Balakin, A.A.; Nikitina, L.V.; Gerzen, O.P.; Nabiev, S.R.; Minigalieva, I.A.; Privalova, L.I.; et al. Further analysis of rat myocardium contractility changes associated with a subchronic lead intoxication. Food Chem. Toxicol. 2019, 125, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Katsnelson, B.A.; Klinova, S.V.; Gerzen, O.P.; Balakin, A.A.; Lookin, O.N.; Lisin, R.V.; Nabiev, S.R.; Privalova, L.I.; Minigalieva, I.A.; Panov, V.G.; et al. Force-velocity characteristics of isolated myocardium preparations from rats exposed to subchronic intoxication with lead and cadmium acting separately or in combination. Food Chem. Toxicol. 2020, 144, 111641. [Google Scholar] [CrossRef] [PubMed]
- Pardee, J.D.; Spudich, J.A. Purification of muscle actin. InMethods in cell biology. Methods Cell Biol. 1982, 24, 271–289. [Google Scholar]
- Potter, J.D. Preparation of troponin and its subnits. Methods Enzymol. 1982, 85, 241–263. [Google Scholar]
- Matyushenko, A.M.; Artemova, N.V.; Shchepkin, D.V.; Kopylova, G.V.; Bershitsky, S.Y.; Tsaturyan, A.K.; Sluchanko, N.N.; Levitsky, D.I. Structural and functional effects of two stabilizing substitutions, D137L and G126R, in the middle part of α-tropomyosin molecule. FEBS J. 2014, 281, 2004–2016. [Google Scholar] [CrossRef] [Green Version]
- Gordon, A.M.; Homsher, E.; Regnier, M. Regulation of contraction in striated muscle. Physiol. Rev. 2000, 80, 853–924. [Google Scholar] [CrossRef]
- Harris, S.P.; Shaffer, J.F.; Wong, P.; Bezold, K.L. Functional differences between the N-terminal domains of mouse and human myosin binding protein-C. J. Biomed. Biotechnol. 2010, 2010, 789798. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Nikitina, L.V.; Kopylova, G.V.; Shchepkin, D.V.; Katsnelson, L.B. Study of the interaction between rabbit cardiac contractile and regulatory proteins. An in vitro motility assay. Biochemistry 2008, 73, 178–184. [Google Scholar] [CrossRef]
- Mashanov, G.I.; Molloy, J.E. Automatic detection of single fluorophores in live cells. Biophys. J. 2007, 92, 2199–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hisano, R.; Cooper, I.V.G. Correlation of force-length area with oxygen consumption in ferret papillary muscle. Circ. Res. 1987, 61, 318–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mast, F.; Elzinga, G. Heat Released During Relaxation Equals Force-Length Area in Isometric Contractions of Rabbit Papillary Muscle. Circ. Res. 1990, 67, 893–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panov, V.G.; Varaksin, A.N. Identification of combined action types in experiments with two toxicants: A response surface linear model with a cross term. Toxicol. Mech. Methods 2016, 26, 139–150. [Google Scholar] [CrossRef]
- Katsnelson, B.A.; Privalova, L.I.; Sutunkova, M.P.; Minigalieva, I.A.; Gurvich, V.B.; Shur, V.Y.; Shishkina, E.V.; Makeyev, O.H.; Valamina, I.E.; Varaksin, A.N.; et al. Experimental Research into Metallic and Met-al Oxide Nanoparticle Toxicity in vivo. In Bioactivity of Engineered Nanoparticles; Yan, B., Zhou, H., Gardea-Torresdey, J., Eds.; Springer: Singapore, 2017; pp. 259–319. [Google Scholar]
- Medinger, J.; Nedyalkova, M.; Lattuada, M. Solvothermal synthesis combined with design of experiments—Optimization approach for magnetite nanocrystal clusters. Nanomaterials 2021, 11, 360. [Google Scholar] [CrossRef]
- Box, G.E.P.; Draper, N.R. Response Surfaces, Mixtures and Ridge Analyses, 2nd ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2007. [Google Scholar]
- Myers, R.H.; Montgomery, D.C.; Anderson-Cook, C.M. Response Surface Methodology: Process and Product Optimization Using Designed Experiments (Wiley Series in Probability and Statistics), 4th ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2016. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indices | Groups of Rats Exposed | |||
|---|---|---|---|---|
| Normal Saline (Control) | Pb | Running | Pb + Running | |
| Number of Rats | 9 | 10 | 10 | 15 |
| Total Ca, mmol/L | 2.487 ± 0.042 | 2.353 ± 0.029 * | 2.368 ± 0.031 * | 2.388 ± 0.028 |
| CK-MB, U/L | 1337.13 ± 123.30 | 1358.82 ± 105.66 | 1398.46 ± 106.53 | 1750.23 ± 122.78 *#● |
| ACE, U/L | 143.00 ± 14.74 | 236.36 ± 21.95 * | 166.64 ± 13.44 # | 135.91 ± 15.22 # |
| VEGF, 106 U/mL | 5.67 ± 0.40 | 5.13 ± 0.31 | 8.10 ± 0.74 *# | 12.58 ± 1.68 *#● |
| Indices | Groups of Rats Exposed | |||
|---|---|---|---|---|
| Normal Saline (Control) | Pb | Running | Pb + Running | |
| ECG 2nd Lead | ||||
| Number of Rats | 10 | 10 | 10 | 10 |
| RR interval, ms | 156.87 ± 4.01 | 161.71 ± 2.88 | 148.62 ± 4.61 # | 163.13 ± 5.77 |
| Heart rate, bpm | 378.52 ± 8.97 | 373.46 ± 6.77 | 408.45 ± 12.57 # | 372.00 ± 13.04 |
| Isoelectric line, mV | −0.06082 ± 0.00091 | −0.0713 ± 0.0025 * | −0.0596 ± 0.0032 # | −0.0635 ± 0.0051 |
| PQ interval, ms | 41.76 ± 0.74 | 44.21 ± 0.93 | 44.24 ± 0.96 | 44.92 ± 1.14 * |
| QRS interval, ms | 24.79 ± 0.45 | 25.43 ± 0.20 | 24.83 ± 0.73 | 24.66 ± 0.58 |
| QT interval, ms | 64.64 ± 1.78 | 66.89 ± 0.60 | 63.89 ± 1.80 | 64.86 ± 0.28 # |
| T amplitude, mV | 0.1250 ± 0.0028 | 0.1506 ± 0.0074 * | 0.138 ± 0.011 | 0.136 ± 0.011 |
| Respiratory flow | ||||
| Number of rats | 10 | 11 | 11 | 12 |
| Respiratory cycle duration (peak to peak), ms | 437.97 ± 15.48 | 459.79 ± 33.09 | 381.89 ± 9.01 *# | 406.01 ± 16.12 |
| Respiratory rate (peak to peak), bpm | 138.51 ± 4.80 | 136.13 ± 8.85 | 157.83 ± 3.79 *# | 150.14 ± 6.02 |
| Inspiratory time, ms | 200.38 ± 6.08 | 187.91 ± 3.98 | 177.81 ± 5.30 * | 201.05 ± 6.86 ● |
| Peak inspiratory flow, ml/s | 10.61 ± 1.17 | 7.96 ± 1.52 | 12.37 ± 1.57 | 7.95 ± 1.41 *● |
| Expiratory volume/inspiratory volume ratio | −1.41 ± 0.58 | −0.63 ± 0.33 | 0.090 ± 0.138 * | −0.58 ± 0.32 |
| End-inspiratory pause, ms | 8.20 ± 0.50 | 7.24 ± 0.27 | 6.85 ± 0.14 * | 7.13 ± 0.24 |
| Relaxation time, ms | 189.63 ± 11.42 | 174.07 ± 14.99 | 166.74 ± 7.76 | 158.73 ± 6.73 * |
| End-expiratory pause, ms | 21.90 ± 0.31 | 23.96 ± 0.58 * | 21.45 ± 0.39 # | 23.51 ± 0.44 *● |
| penH | −0.356 ± 0.054 | −0.65 ± 0.18 | −0.311 ± 0.017 | −0.503 ± 0.064 ● |
| Indices | Groups of Rats Exposed | |||
|---|---|---|---|---|
| Normal Saline (Control) | Pb | Running | Pb + Running | |
| Number of Rats | 10 | 10 | 10 | 10 |
| Vmax, µm/s | 6.19 ± 0.16 | 4.70 ± 0.10 * | 6.26 ± 0.17 # | 5.77 ± 0.16 *#● |
| α-MHC, % | 85 ± 5 | 53 ± 4 * | 82 ± 3 # | 68 ± 4 *#● |
| β-MHC, % | 15 ± 5 | 47 ± 4 * | 18 ± 3 # | 32 ± 4 *#● |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klinova, S.V.; Minigalieva, I.A.; Protsenko, Y.L.; Sutunkova, M.P.; Gurvich, V.B.; Ryabova, J.V.; Valamina, I.E.; Gerzen, O.P.; Nabiev, S.R.; Balakin, A.A.; et al. Changes in the Cardiotoxic Effects of Lead Intoxication in Rats Induced by Muscular Exercise. Int. J. Mol. Sci. 2022, 23, 4417. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23084417
Klinova SV, Minigalieva IA, Protsenko YL, Sutunkova MP, Gurvich VB, Ryabova JV, Valamina IE, Gerzen OP, Nabiev SR, Balakin AA, et al. Changes in the Cardiotoxic Effects of Lead Intoxication in Rats Induced by Muscular Exercise. International Journal of Molecular Sciences. 2022; 23(8):4417. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23084417
Chicago/Turabian StyleKlinova, Svetlana V., Ilzira A. Minigalieva, Yuri L. Protsenko, Marina P. Sutunkova, Vladimir B. Gurvich, Julia V. Ryabova, Irene E. Valamina, Oksana P. Gerzen, Salavat R. Nabiev, Alexander A. Balakin, and et al. 2022. "Changes in the Cardiotoxic Effects of Lead Intoxication in Rats Induced by Muscular Exercise" International Journal of Molecular Sciences 23, no. 8: 4417. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23084417