
Aluminium Nanoparticles as Efficient Adjuvants Compared to Their Microparticle Counterparts: Current Progress and Perspectives

 ,
,  ,
,
Abstract
:1. Introduction
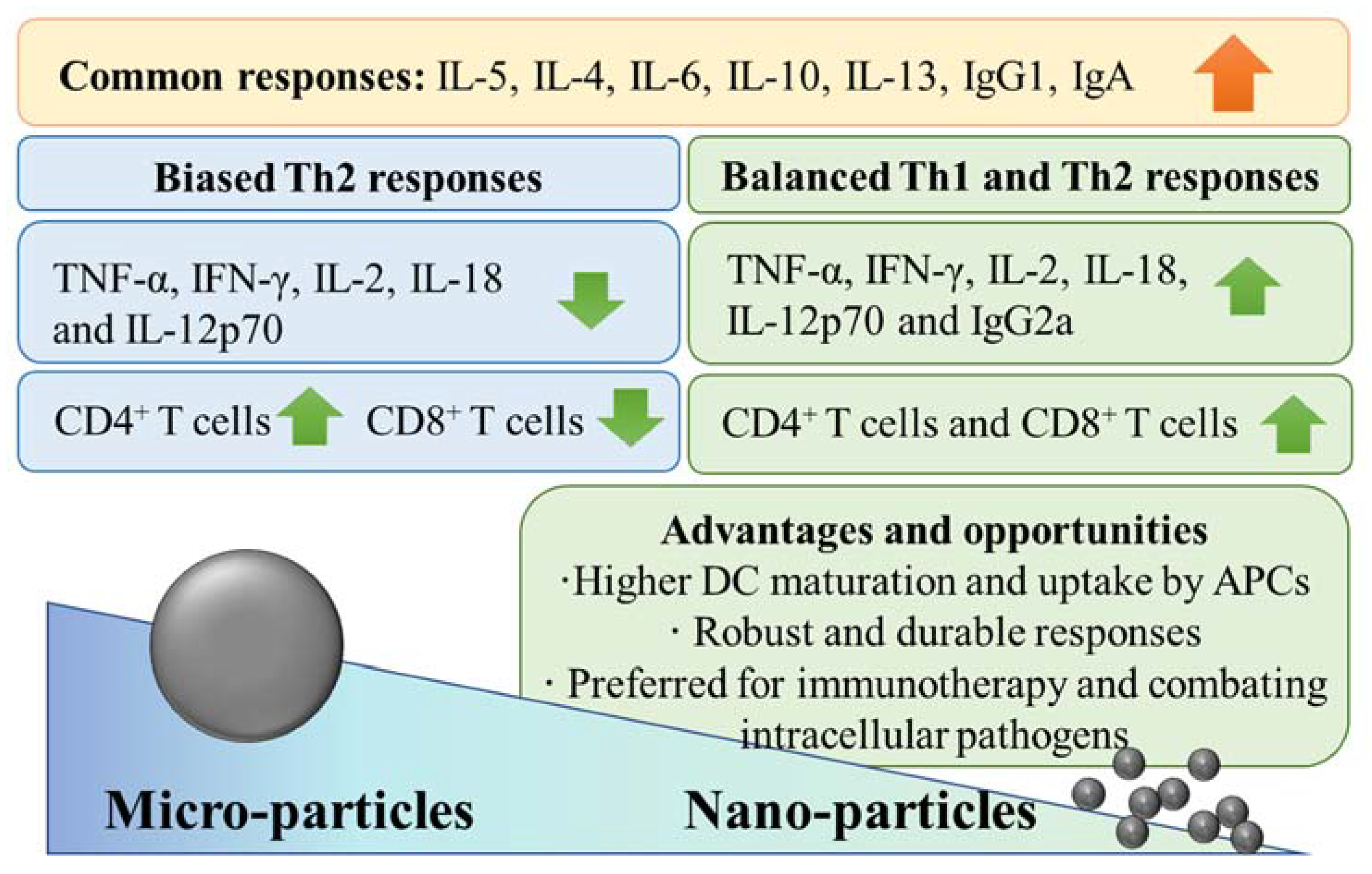
2. Al-NPs Show Different Adjuvant Properties Than Al-MPs
3. Reformulating Conventional Aluminium Microadjuvants into Nanoadjuvants
4. Mechanisms of Al-NP Adjuvanticity
5. Antitumour Immune Responses to Al-NPs

{kind=link}
| Nanostructure | Study Mode | Principal Findings | Ref |
|---|---|---|---|
| OVA and two different adjuvants (CpG and 3pRNA †) were co-loaded on Al(OH)3-NPs (overall size: ~120 nm, diameter). The nanoformulation was used for vaccination. | DC2.4, Raw264.7, and BMDCs were used to assess uptake. BMDCs were used to assess antigen cross-presentation (measured as expression of H2-Kb-SIINFEKL complexes on BMDC surface). C57BL/6 was injected on days 0, 7, and 21 with the nanoformulation, and 7 days after the last injection, anti-OVA antibody levels were measured. C57BL/6 mice bearing B16-OVA tumours were used to assess anticancer effects of prophylactic intra-footpad vaccine injections given 26, 19, and 5 days before tumour inoculation or therapeutic intra-footpad vaccine injection 7 and 14 days after establishing the tumour. | The NP was internalized (~ 55–80%) by all 3 cell lines and enhanced cross-presentation of OVA. Additionally, it increased anti-OVA IgG levels. Whereas the formulation containing both 3pRNA and CpG induced the strongest IgG2a response, the formulation containing only 3pRNA induced the strongest IgG1 response. Vaccination also increased IFN-γ secretion in the spleen. Consistently, the population of IL-4+ CD4+ T cells and IFN-γ + CD8+ T cells were abundant in the spleen. Both prophylactic and therapeutic vaccines delayed tumour growth and prolonged mouse survival. | [56] |
| AlPO4 NPs (~50 nm) loaded with CpG, then coated with B16F10 cell membranes (overall size: ~60 nm). The nanoformulation was used for vaccination. | L929, DC2.4, and Raw264.7 cells were used to assess viability and uptake. BMDCs were used to assess maturation (measured as expression of CD80, CD86, and CCR7). C57BL/6 mice bearing B16F10 tumours were used to assess anticancer effects of prophylactic or therapeutic subcutaneous vaccine injections given 7, 14, and 21 days before tumour inoculation or after tumour establishment. | Cell viability was 90–100% (at concentrations up to 50 µg/mL) and >80% of DC2.4 and Raw264.7 cells for the NP formulation, which also induced maturation of BMDCs. Vaccines increased IFN-γ- and IL-4-expressing CD4+ T cells and IFN-γ-expressing CD8+ T cells in spleen and lymph nodes and concentrations of IL-6, IFN-γ, and TNF-α in culture supernatants of cell suspension from spleen or lymph nodes. The NP formulation induced mild skin inflammation at the injection site and no adverse histopathological effects in heart, liver, spleen, lung, or kidney. In contrast, mice injected with the MP had local skin inflammation and lymph node hyperplasia. Both prophylactic and therapeutic vaccines delayed tumour growth and prolonged mouse survival. | [13] |
| OVA and CpG were loaded on AlO(OH) NPs coated with polymer (PEG) (overall size: ~90 nm). The nanoformulation was used for vaccination. | Raw264.7, DC2.4, or BMDCs were used to assess uptake. BMDCs were used to assess lysosomal escape, antigen cross-presentation (measured as expression of H2-Kb-SIINFEKL complexes on BMDC surface), and DC maturation (measured as CD40, CD80, and CD86 expression, as well as TNF-α and IL-12p70 secretion). C57BL/6 mice bearing either B16-OVA or B16F10 tumours were used to assess biodistribution and anticancer effects of therapeutic intra-footpad vaccine injections given 7 and 14 days after tumour inoculation. | NP formulation was internalized (~100%) by all cell lines and enhanced cross-presentation of OVA. It was retained in draining lymph nodes, leading to an increase in the APC population and maturation compared to commercial microadjuvant. The NP (prime at day 0 and boosted at day 7) induced potent IgG1 and IgG2a responses, but the MP induced Th2-skewed immunity. IFN-γ + CD4+ cells and CTL populations, as well as TNF-α and IFN-γ levels, were higher in the culture (supernatants) of cell suspension from spleen isolated from NP-immunized mice compared to the MP-immunized cohort. The NP vaccine delayed tumour growth and prolonged mouse survival. | [16] |
| Rehydragel® (Al(OH)3@heparanase, LV@HPA) was coated with polyethyleneimine (PEI) to synthesize LV@HPA/PEI nanoadjuvant. Then, OVA or tumour-derived autophagosomes (DRibbles) were adsorbed on the nanoadjuvant. The vaccine was formulated using LV@HPA/PEI-DRibble-DCs. The nanoformulation was used for vaccination ‡. | Murine DCs were used to assess viability and uptake. B3Z, BMDCs, and DCs were used to assess cross-presentation and DC maturation (measured as IL-12 secretion and CD80 and CD86 expression). C57BL/6 mice bearing PancO2 tumours were used to assess anticancer effects of therapeutic vaccine injections given subcutaneously 7 days and in the intra-inguinal lymph nodes 14 days after tumour inoculation. OT-1 mice were used to isolate splenocytes. | Cell viability was ~100% at concentrations up to 10 µg/mL. The NP promoted OVA uptake by the DCs (free OVA uptake: 7% vs. LV@HPA/PEI-OVA: 25.5%), DC maturation, and cross-presentation of OVA. The NP increased secretion of IFN-γ by CTLs isolated from tumour-bearing mice. No obvious body weight loss or abnormality were noticed in the immunized mice during the study course (54 days). Vaccination supressed tumour growth and prolonged mouse survival. | [57] |
| Aminophenol-functionalized α-Al2O3 NPs (~60 nm) conjugated with OVA aminophenol-functionalized α-Al2O3 NPs (~60 nm) conjugated with autophagosomes derived from 3LL cells. The nanoformulations were used for vaccination ^. | BMDCs were used to assess uptake. BMDCs, DC2.4, and OT-I T cells were used to assess cross-presentation (measured as expression of Kb-SIINFEKL on DC surface). Naïve C57BL/6 mice bearing B16–OVA tumours were used to assess anticancer effects of therapeutic vaccine injections given subcutaneously 7 days after tumour inoculation. C57BL/6 mice intravenously injected with 3LL lung tumour cells were used to assess anti-lung metastatic effects of therapeutic vaccine injections given subcutaneously 7 days after tumour inoculation. | NPs were internalized by DCs, which enhanced antigen cross-presentation and stimulation of naïve OVA-specific CD8+ T cells, leading to secretion of IFN-γ and IL-2. Animals immunized with NPs completely rejected tumours and remained tumour-free for >40 days, whereas the MP cohort succumbed to tumour burden. Subcutaneous injection of α-Al2O3-autophagosomes significantly suppressed lung metastases compared to the naked autophagosomes. The combination of vaccine and anti-OX40 antibody led to zero metastases in three of five mice, but no effect was observed in mice treated with anti-OX40 antibody alone. | [58] |
| Al sulphate and chlorin e6 (Ce6) were incorporated into bovine serum albumin (overall size: ~25 nm). The nanoformulation was used for photoimmunotherapy *. | B16F10 cells were used to assess uptake and photodynamic cytotoxicity. BMDCs were used to assess maturation (measured as expression of CD80, CD86, and CD40 and secretion of cytokines IL-6, IL-12/p70, and TNF-α). C57BL/6 mice bearing B16F10 tumours were used to assess biodistribution and anticancer effects of the nanoformulation intravenously injected 8 days after tumour inoculation. | NPs increased Ce6 uptake compared to free Ce6 without reducing cell viability (90–100%). NPs also enhanced cell-killing effects of irradiation and maturation of BMDCs. Following a single intravenous injection, NPs accumulated in tumours at 4-fold higher rate than free Ce6. Compared to locally injected commercial MPs, the NPs significantly reduced growth of primary tumours and metastatic foci and prolonged survival of the animals without causing substantial toxicity to other organs. | [14] |
6. Concluding Remarks and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosen, F.S.; Steiner, L.; Unanue, E. Macmillan Dictionary of Immunology; The MacMillan Press Ltd.: New York, NY, USA, 1989. [Google Scholar]
- Apostólico, J.D.S.; Lunardelli, V.A.S.; Coirada, F.C.; Boscardin, S.B.; Rosa, D.S. Adjuvants: Classification, Modus Operandi, and Licensing. J. Immunol. Res. 2016, 2016, 1459394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glenny, A.; Pope, C.; Waddington, H.; Wallace, U. Immunological notes. xvii–xxiv. J. Pathol. Bacteriol. 1926, 29, 31–40. [Google Scholar] [CrossRef]
- Petrovsky, N. Comparative Safety of Vaccine Adjuvants: A Summary of Current Evidence and Future Needs. Drug Saf. 2015, 38, 1059–1074. [Google Scholar] [CrossRef] [PubMed]
- Danielsson, R.; Eriksson, H. Aluminium adjuvants in vaccines—A way to modulate the immune response. Semin. Cell Dev. Biol. 2021, 115, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Cheever, M.A.; Higano, C.S. PROVENGE (Sipuleucel-T) in Prostate Cancer: The First FDA-Approved Therapeutic Cancer Vaccine. Clin. Cancer. Res. 2011, 17, 3520–3526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.-B.; Xu, J. Better Adjuvants for Better Vaccines: Progress in Adjuvant Delivery Systems, Modifications, and Adjuvant–Antigen Codelivery. Vaccines 2020, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowen, W.S.; Svrivastava, A.K.; Batra, L.; Barsoumian, H.; Shirwan, H. Current challenges for cancer vaccine adjuvant development. Expert Rev. Vaccines 2018, 17, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Ding, B.B.; Teng, B.; Ma, P.A.; Lin, J. Understanding Structure-Function Relationships of Nanoadjuvants for Enhanced Cancer Vaccine Efficacy. Adv. Funct. Mater. 2022, 32, 2111670. [Google Scholar] [CrossRef]
- Garg, A.; Dewangan, H.K. Nanoparticles as Adjuvants in Vaccine Delivery. Crit. Rev. Ther. Drug Carr. Syst. 2020, 37, 183–204. [Google Scholar] [CrossRef]
- Cuzzubbo, S.; Mangsbo, S.; Nagarajan, D.; Habra, K.; Pockley, A.G.; McArdle, S.E. Cancer vaccines: Adjuvant potency, importance of age, lifestyle, and treatments. Front. Immunol. 2021, 11, 3850. [Google Scholar] [CrossRef]
- Gupta, A.; Das, S.; Schanen, B.; Seal, S. Adjuvants in micro-to nanoscale: Current state and future direction. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2016, 8, 61–84. [Google Scholar] [CrossRef] [PubMed]
- Gan, J.; Du, G.; He, C.; Jiang, M.; Mou, X.; Xue, J.; Sun, X. Tumor cell membrane enveloped aluminum phosphate nanoparticles for enhanced cancer vaccination. J. Control. Release 2020, 326, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xue, J.; Chen, W.; Bai, S.; Zheng, T.; He, C.; Guo, Z.; Jiang, M.; Du, G.; Sun, X. Albumin-biomineralized nanoparticles to synergize phototherapy and immunotherapy against melanoma. J. Control. Release 2020, 322, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Aldayel, A.M.; Cui, Z. Aluminum hydroxide nanoparticles show a stronger vaccine adjuvant activity than traditional aluminum hydroxide microparticles. J. Control. Release 2014, 173, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Wang, Q.; Li, L.; Zeng, Q.; Li, H.; Gong, T.; Zhang, Z.; Sun, X. Turning the Old Adjuvant from Gel to Nanoparticles to Amplify CD8+ T Cell Responses. Adv. Sci. 2018, 5, 1700426. [Google Scholar] [CrossRef]
- Orr, M.T.; Khandhar, A.P.; Seydoux, E.; Liang, H.; Gage, E.; Mikasa, T.; Beebe, E.L.; Rintala, N.D.; Persson, K.H.; Ahniyaz, A.; et al. Reprogramming the adjuvant properties of aluminum oxyhydroxide with nanoparticle technology. npj Vaccines 2019, 4, 1. [Google Scholar] [CrossRef]
- Shah, R.R.; O’Hagan, D.T.; Amiji, M.M.; Brito, L.A. The impact of size on particulate vaccine adjuvants. Nanomedicine 2014, 9, 2671–2681. [Google Scholar] [CrossRef]
- Cain, D.W.; Sanders, S.E.; Cunningham, M.M.; Kelsoe, G. Disparate adjuvant properties among three formulations of “alum”. Vaccine 2013, 31, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Hogenesch, H. Mechanism of immunopotentiation and safety of aluminum adjuvants. Front. Immunol. 2012, 3, 406. [Google Scholar] [CrossRef] [Green Version]
- Abkar, M.; Alamian, S.; Sattarahmady, N. A comparison between adjuvant and delivering functions of calcium phosphate, aluminum hydroxide and chitosan nanoparticles, using a model protein of Brucella melitensis Omp31. Immunol. Lett. 2019, 207, 28–35. [Google Scholar] [CrossRef]
- Bungener, L.; Geeraedts, F.; ter Veer, W.; Medema, J.; Wilschut, J.; Huckriede, A. Alum boosts TH2-type antibody responses to whole-inactivated virus influenza vaccine in mice but does not confer superior protection. Vaccine 2008, 26, 2350–2359. [Google Scholar] [CrossRef] [PubMed]
- Chéramy, M.; Skoglund, C.; Johansson, I.; Ludvigsson, J.; Hampe, C.S.; Casas, R. GAD-alum treatment in patients with type 1 diabetes and the subsequent effect on GADA IgG subclass distribution, GAD65 enzyme activity and humoral response. Clin. Immunol. 2010, 137, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebre, F.; Pedroso de Lima, M.C.; Lavelle, E.C.; Borges, O. Mechanistic study of the adjuvant effect of chitosan-aluminum nanoparticles. Int. J. Pharm. 2018, 552, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Frey, A.; Neutra, M.R.; Robey, F.A. Peptomer Aluminum Oxide Nanoparticle Conjugates as Systemic and Mucosal Vaccine Candidates: Synthesis and Characterization of a Conjugate Derived from the C4 Domain of HIV-1MN Gp120. Bioconj. Chem. 1997, 8, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Frey, A.; Mantis, N.; Kozlowski, P.A.; Quayle, A.J.; Bajardi, A.; Perdomo, J.J.; Robey, F.A.; Neutra, M.R. Immunization of mice with peptomers covalently coupled to aluminum oxide nanoparticles. Vaccine 1999, 17, 3007–3019. [Google Scholar] [CrossRef]
- Tang, C.; Huang, X.; Yang, F.; Li, M.; Fan, G.; Yue, H. Adjuvant effect of aluminum hydroxide nanoparticles on Newcastle diseases antigen in chichens. Chin. Vet. Sci. 2008, 38, 1060–1064. [Google Scholar]
- Ruwona, T.B.; Xu, H.; Li, X.; Taylor, A.N.; Shi, Y.-C.; Cui, Z. Toward understanding the mechanism underlying the strong adjuvant activity of aluminum salt nanoparticles. Vaccine 2016, 34, 3059–3067. [Google Scholar] [CrossRef] [Green Version]
- Alshanqiti, F.M.; Al-Masaudi, S.B.; Al-Hejin, A.M.; El-Baky, N.A.; Redwan, E.M. Development of nanoparticle adjuvants to potentiate the immune response against diphtheria toxoid. Hum. Antibodies 2017, 26, 75–85. [Google Scholar] [CrossRef]
- Amini, Y.; Moradi, B.; Fasihi-Ramandi, M. Aluminum hydroxide nanoparticles show strong activity to stimulate Th-1 immune response against tuberculosis. Artif. Cells Nanomed. Biotechnol. 2017, 45, 1331–1335. [Google Scholar] [CrossRef] [Green Version]
- Gogoi, H.; Mani, R.; Aggarwal, S.; Malik, A.; Munde, M.; Bhatnagar, R. Crystalline and Amorphous Preparation of Aluminum Hydroxide Nanoparticles Enhances Protective Antigen Domain 4 Specific Immunogenicity and Provides Protection Against Anthrax. Int. J. Nanomed. 2020, 15, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Gogoi, H.; Mani, R.; Malik, A.; Sehrawat, P.; Bhatnagar, R. Co-Administration of Aluminium Hydroxide Nanoparticles and Protective Antigen Domain 4 Encapsulated Non-Ionic Surfactant Vesicles Show Enhanced Immune Response and Superior Protection against Anthrax. Vaccines 2020, 8, 571. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wei, C.; Zhang, Z.; Liu, T.; Wang, T. Aluminum Nanoparticles Acting as a Pulmonary Vaccine Adjuvant-Delivery System (VADS) Able to Safely Elicit Robust Systemic and Mucosal Immunity. J. Inorg. Organomet. Polym. Mater. 2020, 30, 4203–4217. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Qiu, C.; Chen, M.; Liu, T.; Wang, T. Covering Aluminum Oxide Nanoparticles with Biocompatible Materials to Efficiently Deliver Subunit Vaccines. Vaccines 2019, 7, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Ji, Z.; Liao, Y.-P.; Chang, C.H.; Wang, X.; Ku, J.; Xue, C.; Mirshafiee, V.; Xia, T. Enhanced Immune Adjuvant Activity of Aluminum Oxyhydroxide Nanorods through Cationic Surface Functionalization. ACS Appl. Mater. Interfaces 2017, 9, 21697–21705. [Google Scholar] [CrossRef]
- Li, X.; Hufnagel, S.; Xu, H.; Valdes, S.A.; Thakkar, S.G.; Cui, Z.; Celio, H. Aluminum (Oxy)Hydroxide Nanosticks Synthesized in Bicontinuous Reverse Microemulsion Have Potent Vaccine Adjuvant Activity. ACS Appl. Mater. Interfaces 2017, 9, 22893–22901. [Google Scholar] [CrossRef]
- Wang, T.; Zhen, Y.; Ma, X.; Wei, B.; Wang, N. Phospholipid Bilayer-Coated Aluminum Nanoparticles as an Effective Vaccine Adjuvant-Delivery System. ACS Appl. Mater. Interfaces 2015, 7, 6391–6396. [Google Scholar] [CrossRef]
- Bilyy, R.; Paryzhak, S.; Turcheniuk, K.; Dumych, T.; Barras, A.; Boukherroub, R.; Wang, F.; Yushin, G.; Szunerits, S. Aluminum oxide nanowires as safe and effective adjuvants for next-generation vaccines. Mater. Today 2019, 22, 58–66. [Google Scholar] [CrossRef]
- Hilgers, L.T.; Ghenne, L.; Nicolas, I.; Fochesato, M.; Lejeune, G.; Boon, B. Alkyl–polyacrylate esters are strong mucosal adjuvants. Vaccine 2000, 18, 3319–3325. [Google Scholar] [CrossRef]
- Khandhar, A.P.; Liang, H.; Simpson, A.C.; Reed, S.G.; Carter, D.; Fox, C.B.; Orr, M.T. Physicochemical structure of a polyacrylic acid stabilized nanoparticle alum (nanoalum) adjuvant governs TH1 differentiation of CD4+ T cells. Nanoscale 2020, 12, 2515–2523. [Google Scholar] [CrossRef]
- Vrieling, H.; Espitia Ballestas, M.; Hamzink, M.; Willems, G.-J.; Soema, P.; Jiskoot, W.; Kersten, G.; Metz, B. Stabilised aluminium phosphate nanoparticles used as vaccine adjuvant. Colloids Surf. B. Biointerfaces 2019, 181, 648–656. [Google Scholar] [CrossRef]
- Liang, Z.; Yang, Y.; Yu, G.; Zhu, H.; Xia, X.; Chen, C.; Fu, D.; Li, M.; Cheng, G.; Xue, C.; et al. Engineering aluminum hydroxyphosphate nanoparticles with well-controlled surface property to enhance humoral immune responses as vaccine adjuvants. Biomaterials 2021, 275, 120960. [Google Scholar] [CrossRef] [PubMed]
- Simón-Vázquez, R.; Lozano-Fernández, T.; Dávila-Grana, A.; González-Fernández, A. Metal oxide nanoparticles interact with immune cells and activate different cellular responses. Int. J. Nanomed. 2016, 11, 4657–4668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Li, X.; Cui, Z. Toward understanding the mechanism underlying the strong adjuvant activity of aluminum salt nanoparticles. Ruwona TB, Xu H, Li X, Taylor AN, Shi Y, Cui Z. Vaccine 2016;34:3059–67. Vaccine 2017, 35, 1102–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mold, M.; Eriksson, H.; Siesjö, P.; Darabi, A.; Shardlow, E.; Exley, C. Unequivocal identification of intracellular aluminium adjuvant in a monocytic THP-1 cell line. Sci. Rep. 2014, 4, srep06287. [Google Scholar] [CrossRef] [Green Version]
- Mold, M.; Shardlow, E.; Exley, C. Insight into the cellular fate and toxicity of aluminium adjuvants used in clinically approved human vaccinations. Sci. Rep. 2016, 6, 31578. [Google Scholar] [CrossRef] [Green Version]
- Kesavardhana, S.; Kanneganti, T.-D. Mechanisms governing inflammasome activation, assembly and pyroptosis induction. Int. Immunol. 2017, 29, 201–210. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, H.; Kouadir, M.; Song, H.; Shi, F. Recent advances in the mechanisms of NLRP3 inflammasome activation and its inhibitors. Cell Death Dis. 2019, 10, 128. [Google Scholar] [CrossRef] [Green Version]
- Hamarsheh, S.A.; Zeiser, R. NLRP3 Inflammasome Activation in Cancer: A Double-Edged Sword. Front. Immunol. 2020, 11, 1444. [Google Scholar] [CrossRef]
- Thakkar, S.G.; Xu, H.; Li, X.; Cui, Z. Uric acid and the vaccine adjuvant activity of aluminium (oxy)hydroxide nanoparticles. J. Drug Target. 2018, 26, 474–480. [Google Scholar] [CrossRef]
- Sun, B.; Ji, Z.; Liao, Y.P.; Wang, M.; Wang, X.; Dong, J.; Chang, C.H.; Li, R.; Zhang, H.; Nel, A.E.; et al. Engineering an effective immune adjuvant by designed control of shape and crystallinity of aluminum oxyhydroxide nanoparticles. ACS Nano 2013, 7, 10834–10849. [Google Scholar] [CrossRef] [Green Version]
- Mesa, C.; Fernández, L.E. Challenges facing adjuvants for cancer immunotherapy. Immunol. Cell Biol. 2004, 82, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Wang, W.; Wang, R.; Duan, J.; Hu, Y.; Ma, J.; Zhou, J.; Xie, S.; Lu, X.; Zhu, Z.; et al. Aluminum nanoparticles enhance anticancer immune response induced by tumor cell vaccine. Cancer Nanotechnol. 2010, 1, 63–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.; Rolfe, B.E.; Zhang, B.; Mohammed, Y.H.; Gu, W.; Xu, Z.P. Polarized immune responses modulated by layered double hydroxides nanoparticle conjugated with CpG. Biomaterials 2014, 35, 9508–9516. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, X.; Wen, Y.; Fu, J.; Wang, H.; Ma, Z.; Shi, Y.; Wang, B. Suppression of established hepatocarcinoma in adjuvant only immunotherapy: Alum triggers anti-tumor CD8+ T cell response. Sci. Rep. 2015, 5, 17695. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, Y.; Tang, Y.; Zhou, Z.; Tan, L.; Gong, T.; Zhang, L.; Sun, X. Co-delivery of antigen and dual adjuvants by aluminum hydroxide nanoparticles for enhanced immune responses. J. Control. Release 2020, 326, 120–130. [Google Scholar] [CrossRef]
- Dong, H.; Wen, Z.-F.; Chen, L.; Zhou, N.; Liu, H.; Dong, S.; Hu, H.; Mou, Y. Polyethyleneimine modification of aluminum hydroxide nanoparticle enhances antigen transportation and cross-presentation of dendritic cells. Int. J. Nanomed. 2018, 13, 3353–3365. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, Y.; Jiao, J.; Hu, H.-M. Alpha-alumina nanoparticles induce efficient autophagy-dependent cross-presentation and potent antitumour response. Nat. Nanotech. 2011, 6, 645–650. [Google Scholar] [CrossRef]
- Lozhkomoev, A.S.; Mikhaylov, G.; Turk, V.; Turk, B.; Vasiljeva, O. Application of Crumpled Aluminum Hydroxide Nanostructures for Cancer Treatment. In Multiscale Biomechanics and Tribology of Inorganic and Organic Systems; Springer: Cham, Switzerland, 2021; pp. 211–223. [Google Scholar]
- Hao, G.; Xu, Z.P.; Li, L. Manipulating extracellular tumour pH: An effective target for cancer therapy. RSC Adv. 2018, 8, 22182–22192. [Google Scholar] [CrossRef] [Green Version]
- Lozhkomoev, A.S. Alkalization of tumor microenvironment for cancer treatment. In AIP Conference Proceedings; AIP Publishing LLC: Melville, NY, USA, 2017; Volume 1882, p. 020046. [Google Scholar]
- Lozhkomoev, A.S.; Bakina, O.V.; Fomenko, A.N.; Avgustinovich, A.V.; Afanasyev, S.G.; Dobrodeev, A.Y.; Spirina, L.V.; Tarasova, A.S.; Urmonov, U.B. The use of nanostructured metal oxides for directional pH changes of tumor cells microenvironment. Sib. J. Oncol. 2019, 18, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-Y.; Xu, L.; Li, L.-F.; Liu, X.-X.; Gao, J.-X.; Bai, Y.-R. Inhibiting the CD8+ T cell infiltration in the tumor microenvironment after radiotherapy is an important mechanism of radioresistance. Sci. Rep. 2018, 8, 11934. [Google Scholar] [CrossRef]
- Kempson, I. Mechanisms of nanoparticle radiosensitization. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2021, 13, e1656. [Google Scholar] [CrossRef] [PubMed]
- Bakina, O.V.; Svarovskaya, N.V.; Miller, A.A.; Lozhkomoev, A.S.; Avgustinovich, A.V.; Dobrodeev, A.Y.; Spirina, L.V.; Afanasyev, S.G. Synergistic effect of antitumor activity of doxorubicin and bicomponent nanostructures based on aluminum oxide. Sib. J. Oncol. 2020, 19, 82–89. [Google Scholar] [CrossRef]
- Lerner, M.I.; Mikhaylov, G.; Tsukanov, A.A.; Lozhkomoev, A.S.; Gutmanas, E.; Gotman, I.; Bratovs, A.; Turk, V.; Turk, B.; Psakhye, S.G.; et al. Crumpled Aluminum Hydroxide Nanostructures as a Microenvironment Dysregulation Agent for Cancer Treatment. Nano Lett. 2018, 18, 5401–5410. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Kovochich, M.; Brant, J.; Hotze, M.; Sempf, J.; Oberley, T.; Sioutas, C.; Yeh, J.I.; Wiesner, M.R.; Nel, A.E. Comparison of the abilities of ambient and manufactured nanoparticles to induce cellular toxicity according to an oxidative stress paradigm. Nano Lett. 2006, 6, 1794–1807. [Google Scholar] [CrossRef]

| Adjuvant (Particle Size) | Vaccine Formulation | Principal Findings | Ref |
|---|---|---|---|
| Al(OH)3-NPs (~112 nm) vs. Al(OH)3-MPs (~9.3 μm) | Ovalbumin (OVA) and Bacillus anthracis protective antigen (PA) were adsorbed on the adjuvants. | OVA had more affinity to bind to the NPs than the MPs due to larger total surface area and more positive zeta potential of the NP. At equal OVA levels adsorbed on the particles, the NP induced higher anti-OVA IgG levels than the MP. The NP also induced higher anti-PA IgG levels than the MP 4 weeks after the second immunization. APCs internalized significantly higher levels of OVA adsorbed on the NP than the MP. | [15] |
| Al hydroxyphosphate NPs (<100 nm) vs. Al hydroxyphosphate MPs (~8–13 μm) | Egg lysozyme was adsorbed on the adjuvants. | The NP induced significantly higher antigen-specific IgG levels than the MP. | [28] |
| The sizes of Al(OH)3 (~0.99–1.96 μm) and AlPO4 (~1 μm) particles were reduced by applying high shear forces, then compared to Alhydrogel® and a commercially available vaccine (TETAVAX). | Diphtheria toxoid was adsorbed on the adjuvants. | The size reduction improved protein loading capacity, boosted antidiphtheria antibody titration, and induced stronger Th2 antibody isotypes (IgG1 and IgA). Size-reduced Al(OH)3 adjuvant also induced stronger Th2 cytokines (IL-5, IL-6, IL-10 and IL-13). | [29] |
| Al(OH)3-NPs (~141.1 nm) vs. Bacillus Calmette–Guerin (BCG) vaccine | Mycobacterium tuberculosis ESAT-6-like protein EsxV was adsorbed on the Al(OH)3-NPs. | The NP stimulated secretion of Th1 cytokines, e.g., IFN-γ comparable to BCG. | [30] |
| Amorphous and crystalline forms of Al(OH)3-NPs (150–200 nm) vs. Alhydrogel® | B. anthracis protective antigen domain 4 (D4) was adsorbed on the adjuvants. | The NPs enhanced antigen uptake by THP-1 cells, induced more robust and durable Th1/Th2 responses evidenced by higher IgG1 and IgG2a levels compared to Alhydrogel®, and induced higher Th1 cytokine levels (IL-2 and IFN-γ). Conversely, Alhydrogel® induced comparable or higher Th2 cytokine levels (IL-4 and IL-10). NPs prolonged survival of anthrax spore-challenged mice. The crystalline NP had moderate binding affinity compared to its amorphous counterpart, resulting in moderate antigen release (almost equal to Alhydrogel®). | [31] |
| Crystalline Al(OH)3-NPs (150–200 nm) vs. Alhydrogel® | D4 was encapsulated by non-ionic surfactant-based vesicles and adsorbed on the adjuvants. | The NP induced higher antigen-specific antibody titres (anti-D4 IgG) and IgG isotypes (IgG1 and IgG2a) than Alhydrogel®. It also stimulated splenocytes to produce both Th1 (IL-2, IFN-γ, and TNF-α) and Th2 (IL-4, IL-6, and IL-10) cytokines. The NP induced superior protection against anthrax spore challenge. | [32] |
| Al2O3-NPs (~30 nm) as a pulmonary vaccine adjuvant-delivery system vs. AlPO4-MP (2 μm) | OVA was adsorbed on the adjuvants. | NPs had significantly higher uptake by bone-marrow-derived dendritic cells (BMDCs) and promoted DC maturation to a higher degree, measured as CD40, CD80, and CD86 surface expression. NPs did not influence Raw264.7 (macrophage) cell viability at concentrations as high as 200 µg/mL. The NP induced more balanced Th1/Th2 responses, measured as anti-OVA IgG, mucosal IgA, and cytokine secretion (IFN-γ and IL-4), with only mild pulmonary inflammation. | [33] |
| Rod-shaped Al(OH)3-NPs stabilized with PAA (~60 nm) vs. Alhydrogel® | ID93 (M. tuberculosis vaccine antigen) or recombinant rHA (seasonal influenza hemagglutinin) were adsorbed on the adjuvants. | Unlike Alhydrogel®, the NP increased splenic IFN-γ-secreting CD4+ T cells and levels of Th1 cytokines, IL-18, and IL-12p70. The NP induced more robust and durable ID93-specific IgG1 and IgG2c antibodies, whereas Alhydrogel® induced IgG1 antibody and was biased toward a Th2 response. The NP induced superior protection against lethal influenza challenge. | [17] |
| Al2O3-NPs (~28 nm), phospholipid bilayer-coated Al2O3-NPs (PLANs, ~33 nm) and the PEGylated PLANs (PEG-PLANs, ~31 nm) vs. AlPO4-MPs (~2 μm) | OVA was adsorbed on the adjuvants. | BMDC uptake of formulations ranked in the order of AlPO4-MPs<Al2O3-NPs<PEG- PLANs<PLANs. The microparticle reduced cell (Raw264.7) viability. NPs did not show cytotoxicity and promoted cell growth. NPs and, more specifically, PLANs promoted DC maturation, measured as CD40, CD80, and CD86 surface expression. PEG-PLANs accumulated in draining lymph nodes at significantly higher levels. Whereas PLANs and PEG-PLANs elicited stronger humoral responses than AlPO4-MPs, Al2O3-NPs did not. NPs induced Th1 responses (IgG2a> IgG1), and conversely, the MP induced Th2 responses. NPs increase IL-4 and IFN-γ levels, as well as CD8+ T cells, in spleen compared to the MP. PEG-PLANs were the most effective adjuvant. | [34] |
| AlOOH nanorods (ALNRs) functionalized with (3-aminopropyl) triethoxysilane (ALNR-NH2) or 3-(trihydroxysilyl)- 1-propanesulfonic acid (ALNR-SO3H) (diameter: 20 nm, length: 150–200 nm) vs. Imject® | OVA was adsorbed on the adjuvants. | THP-1 cell uptake of formulations ranked in the order of ALNR-NH2 = ALNR-SO3H< Imject®. Moreover, IL-1β secretion by THP-1 cells ranked in the order of ALNR-SO3H≤Imject®<ALNR-NH2. Cellular oxidative stress (measured as glutathione level) of formulations ranked in the order of ALNR-SO3H <alum<ALNR-NH2. ALNR-SO3H and Imject® had the same capacity to induce anti-OVA IgG1 and IgE, whereas ALNR-NH2 induced significantly higher levels of the antibodies. | [35] |
| Amorphous AlOOH nanosticks (diameter: ~8 nm, length: ~80 nm) vs. Alhydrogel® (~900 nm) | OVA was adsorbed on the adjuvants. | J774A.1 macrophage uptake of NPs was higher than that of Alhydrogel®. THP-1 cells treated with NPs released higher levels of IL-1β than Alhydrogel®. NPs induced higher levels of serum anti-OVA IgG and IgG1 than Alhydrogel®. Al nanosticks and Alhydrogel® induced local subcutaneous nodule and granuloma formation, although the site injected with the Al nanosticks had a relatively lower cellularity. | [36] |
| Al(OH)3-NPs (~40 nm), phospholipid bilayer-coated Al(OH)3-NPs (PLAlOH3: ~50 nm) vs. Al(OH)3-MPs (~10 μm) | OVA was adsorbed on the adjuvants. | BMDC uptake of formulations ranked in the order of Al(OH)3-MPs<Al(OH)3-NPs<PLAl(OH)3. NPs induced more durable and higher anti-OVA IgG and IgA than the MP. PLAl(OH)3 induced balanced IgG2a>IgG1, contrary to the MP which induced biased Th2 response (very high level of IgG1). Whereas PLAl(OH)3 elevated both IL-4 and IFN-γ in serum and supernatant of splenocytes, the MP increased only the IL-4 level. The MP increased only CD4+ T-cell populations in the spleen, but the PLAl(OH)3 elevated both CD4+ T and CD8+ T-cell populations. The stimulation index for splenocyte proliferation was ~2-fold higher for PLAl(OH)3 than the MP. Following subcutaneous injection into a forelimb, PLAl(OH)3 was accumulated in axillary lymph nodes and taken up by DCs. Following intramuscular injection, neither NP induced local inflammation, but the MP induced severe inflammatory reactions. | [37] |
| Al2O3-nanowire (diameter: ~20–40 nm, length: ~20–60 µm) vs. Al2O3-MPs (20 µm scale) and Alhydrogel® (2 µm) | OVA was adsorbed on the adjuvants. | All formulations were non-toxic to HeLa and THP-1 cells (up to 200 µg/mL); however, the nanowire was slightly toxic to U87MG cells (viability: ~70%) compared to Al2O3-MPs (viability: ~80%) and Alhydrogel® (viability: ~85%) at the mentioned concentration. The nanowire induced higher levels of anti-OVA IgG than MPs 11 days after the second immunization. Cellular immune response, measured as delayed-type hypersensitivity, was stronger in nanowire-treated mice than the Al2O3-MP-injected cohort at 6–24 h after antigen exposure. Following injection into an air sac in the flank, the nanowire induced a lower degree of microvascular damage and oedema than Alhydrogel®. | [38] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazarizadeh, A.; Staudacher, A.H.; Wittwer, N.L.; Turnbull, T.; Brown, M.P.; Kempson, I. Aluminium Nanoparticles as Efficient Adjuvants Compared to Their Microparticle Counterparts: Current Progress and Perspectives. Int. J. Mol. Sci. 2022, 23, 4707. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094707
Nazarizadeh A, Staudacher AH, Wittwer NL, Turnbull T, Brown MP, Kempson I. Aluminium Nanoparticles as Efficient Adjuvants Compared to Their Microparticle Counterparts: Current Progress and Perspectives. International Journal of Molecular Sciences. 2022; 23(9):4707. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094707
Chicago/Turabian StyleNazarizadeh, Ali, Alexander H. Staudacher, Nicole L. Wittwer, Tyron Turnbull, Michael P. Brown, and Ivan Kempson. 2022. "Aluminium Nanoparticles as Efficient Adjuvants Compared to Their Microparticle Counterparts: Current Progress and Perspectives" International Journal of Molecular Sciences 23, no. 9: 4707. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094707