
The Plasmid pEX18Gm Indirectly Increases Caenorhabditis elegans Fecundity by Accelerating Bacterial Methionine Synthesis

Abstract
:1. Introduction
2. Results
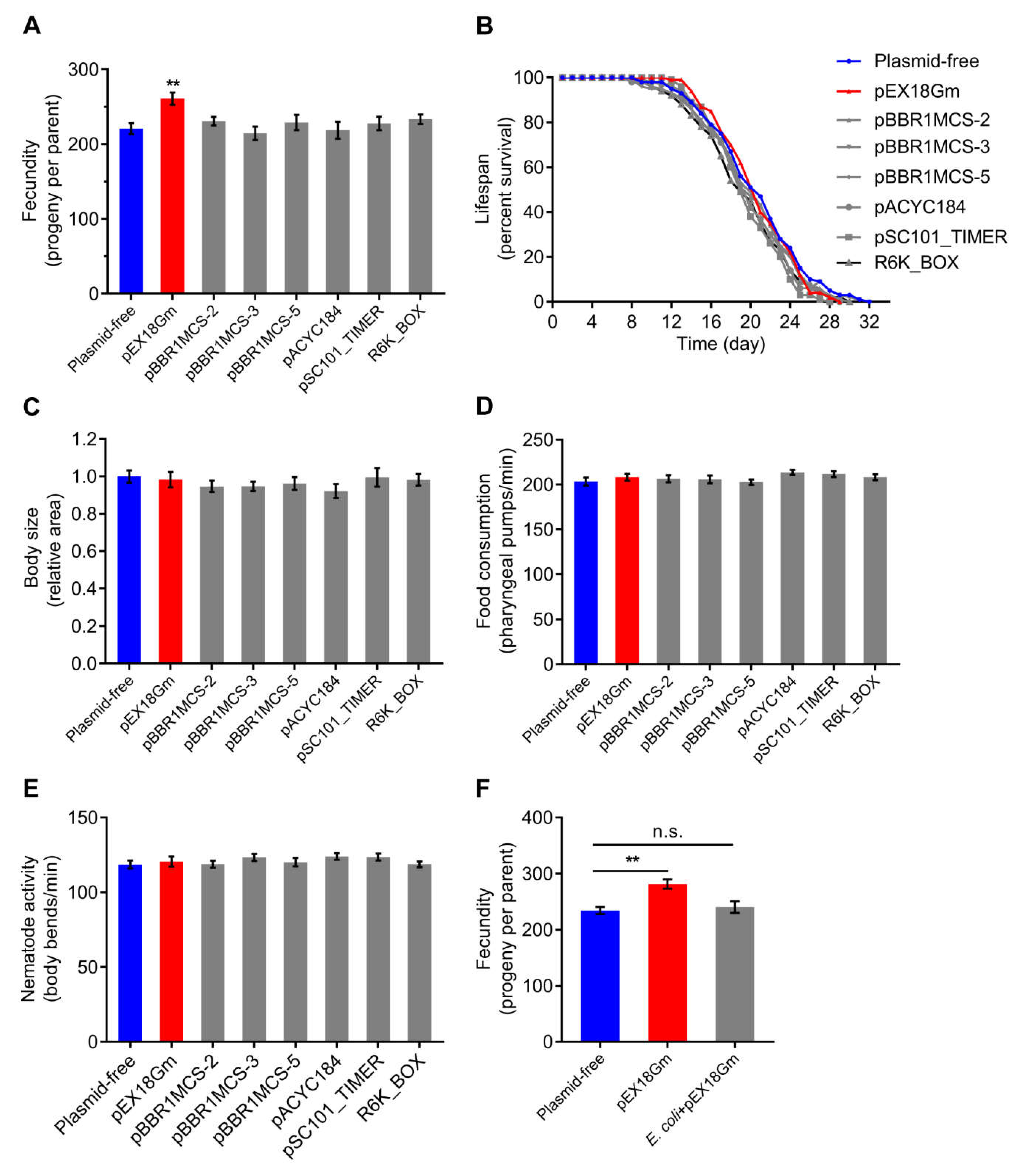
2.1. E. coli OP50 Harboring pEX18Gm Increases C. elegans Fecundity
2.2. RNA-seq Analysis of E. coli Strains
2.3. The Bacterial Methionine Synthase Gene metH Is Necessary for pEX18Gm-Induced Increase in C. elegans Fecundity
2.4. Methionine Is a Metabolite of pEX18Gm-Induced Increase in C. elegans Fecundity
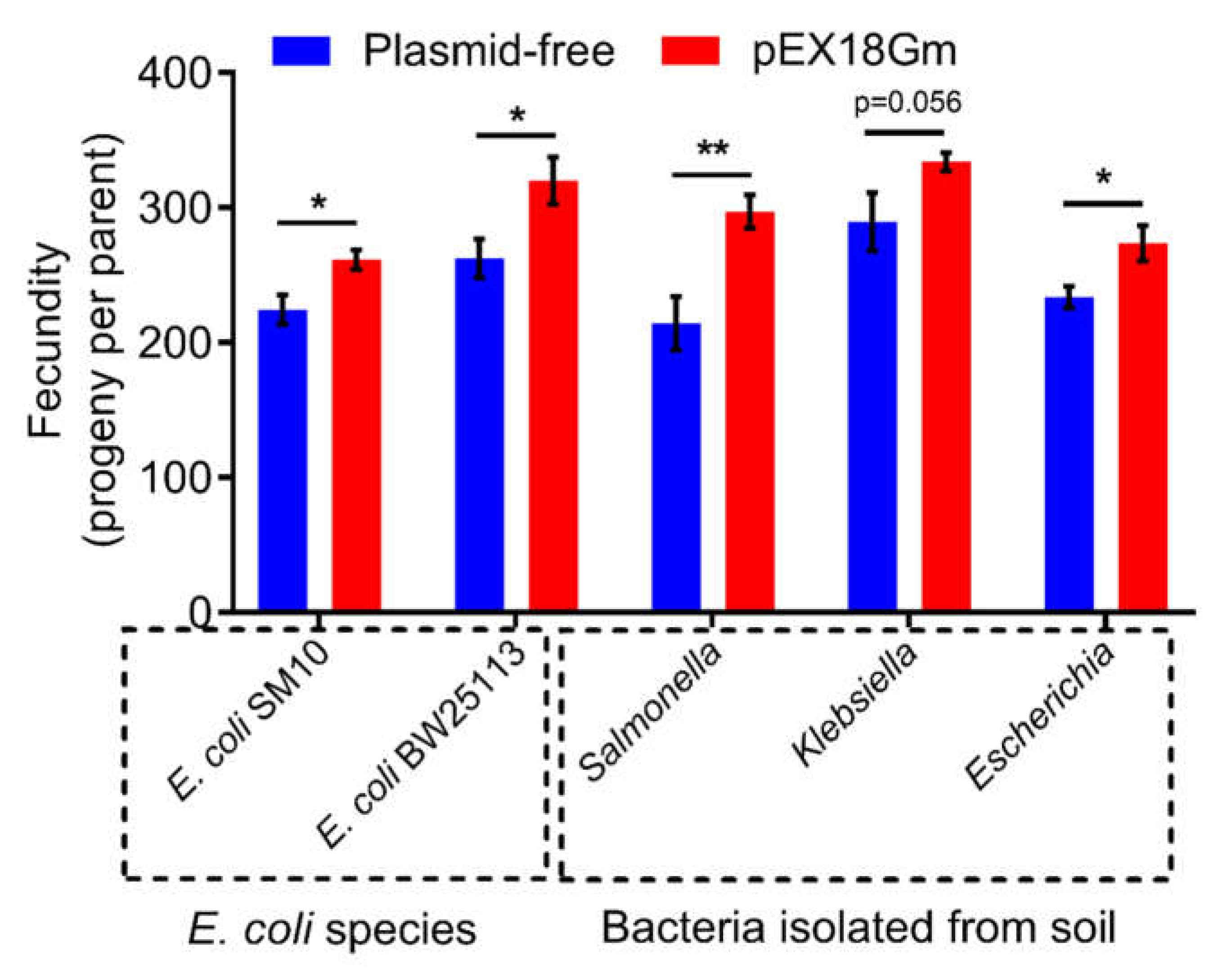
2.5. pEX18Gm-Induced Increase in C. elegans Fecundity Is Not Limited to the E. coli OP50 Species
3. Discussion
4. Materials and Methods
4.1. Plasmids and Chemicals
4.2. Strains and Growth Conditions
4.3. Competent Cells Preparation and Plasmid Introduction
4.4. Phenotypic Analysis of C. elegans Life History Traits
4.4.1. Fecundity
4.4.2. Lifespan
4.4.3. Body Size
4.4.4. Pharyngeal Pump Rates
4.4.5. Body Bends
4.5. Bacterial Growth Rate
4.6. RNA-seq and Analysis
4.7. qRT-PCR
4.8. E. coli Gene Knockout
4.9. Methionine and Vitamin B12 Supplement Assay
4.10. Methionine Determination in Bacteria and C. elegans by LC-MS/MS
4.10.1. Bacteria Sample Preparation
4.10.2. C. elegans Sample Preparation
4.10.3. LC-MS/MS Analysis
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lederberg, J. Plasmid (1952–1997). Plasmid 1998, 39, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mitragotri, S.; Burke, P.A.; Langer, R. Overcoming the challenges in administering biopharmaceuticals: Formulation and delivery strategies. Nat. Rev. Drug Discov. 2014, 13, 655–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, S.P.; Perutka, J.; Powell, J.E.; Geng, P.; Richhart, D.D.; Byrom, M.; Kar, S.; Davies, B.W.; Ellington, A.D.; Moran, N.A.; et al. Genetic engineering of bee gut microbiome bacteria with a toolkit for modular assembly of broad-host-range plasmids. ACS Synth. Biol. 2018, 7, 1279–1290. [Google Scholar] [CrossRef]
- Takahashi, Y.; Shintani, M.; Takase, N.; Kazo, Y.; Kawamura, F.; Hara, H.; Nishida, H.; Okada, K.; Yamane, H.; Nojiri, H. Modulation of primary cell function of host Pseudomonas bacteria by the conjugative plasmid pCAR1. Environ. Microbiol. 2015, 17, 134–155. [Google Scholar] [CrossRef] [PubMed]
- Tester, M.; Langridge, P. Breeding technologies to increase crop production in a changing world. Science 2010, 327, 818–822. [Google Scholar] [CrossRef]
- San Millan, A.; MacLean, R.C. Fintess costs of plasmids: A limit to plasmid transmission. Microbiol. Spectr. 2017, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Vogwill, T.; MacLean, R.C. The genetic basis of the fitness costs of antimicrobial resistance: A meta-analysis approach. Evol. Appl. 2015, 8, 284–295. [Google Scholar] [CrossRef]
- Anthony Mason, C.; Bailey, J. Effects of plasmid presence on growth and enzyme activity of Escherichia coli DH5α. Appl. Microbiol. Biotechnol. 1989, 32, 54–60. [Google Scholar] [CrossRef]
- Chernin, L.S.; Goldberg, G.I.; Goldfarb, D.M. DNA synthesis in F- cells treated with filtrates of male strains of E. coli K12. Mol. Gen. Genet. MGG 1970, 108, 376–377. [Google Scholar] [CrossRef]
- Goldfarb, D.M.; Chernin, L.S.; Fomitchev, Y.K. Inhibition of nitrogen mustard after-effect in F minus-cells by the factor produced by E. coli K-12 fertile strains. Mol. Gen. Genet. MGG 1969, 104, 107–115. [Google Scholar] [CrossRef]
- Howarth, S. Resistance to the bactericidal effect of ultraviolet radiation conferred on Enterobacteria by the colicine factor coli. J. Gen. Microbiol. 1965, 40, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Howarth, S. Increase in frequency of ultraviolet-induced mutation brought about by the colicine factor, colI in Salmonella typhimurium. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 1966, 3, 129–134. [Google Scholar] [CrossRef]
- Billane, K.; Harrison, E.; Cameron, D.; Brockhurst, M.A. Why do plasmids manipulate the expression of bacterial phenotypes? Phil. Trans. R. Soc. B 2021, 377, 20200461. [Google Scholar] [CrossRef] [PubMed]
- Buckner, M.M.C.; Saw, H.T.H.; Osagie, R.N.; McNally, A.; Ricci, V.; Wand, M.E.; Woodford, N.; Ivens, A.; Webber, M.A.; Piddock, L.J.V.; et al. Clinically relevant plasmid-host interactions indicate that transcriptional and not genomic modifications ameliorate fitness costs of Klebsiella pneumoniae carbapenemase-carrying plasmids. mBio 2018, 9, e02303-17. [Google Scholar] [CrossRef] [Green Version]
- Lang, K.S.; Johnson, T.J. Transcriptome modulations due to A/C2 plasmid acquisition. Plasmid 2015, 80, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Shintani, M.; Takahashi, Y.; Tokumaru, H.; Kadota, K.; Hara, H.; Miyakoshi, M.; Naito, K.; Yamane, H.; Nishida, H.; Nojiri, H. Response of the Pseudomonas host chromosomal transcriptome to carriage of the IncP-7 plasmid pCAR1. Environ. Microbiol. 2010, 12, 1413–1426. [Google Scholar]
- Harr, B.; Schlotterer, C. Gene expression analysis indicates extensive genotype-specific crosstalk between the conjugative F-plasmid and the E. coli chromosome. BMC Microbiol. 2006, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- San Millan, A.; Toll-Riera, M.; Qi, Q.; Betts, A.; Hopkinson, R.J.; McCullagh, J.; MacLean, R.C. Integrative analysis of fitness and metabolic effects of plasmids in Pseudomonas aeruginosa PAO1. ISME J. 2018, 12, 3014–3024. [Google Scholar] [CrossRef]
- Diot, C.; Garcia-Gonzalez, A.P.; Walhout, A.J.M. C. elegans and its bacterial diet: An interspecies model to explore the effects of microbiota on drug response. Drug Discov. Today Dis. Models 2019, 28, 21–26. [Google Scholar] [CrossRef]
- Zhang, J.; Holdorf, A.D.; Walhout, A.J. C. elegans and its bacterial diet as a model for systems-level understanding of host-microbiota interactions. Curr. Opin. Biotechnol. 2017, 46, 74–80. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Gusarov, I.; Gautier, L.; Smolentseva, O.; Shamovsky, I.; Eremina, S.; Mironov, A.; Nudler, E. Bacterial nitric oxide extends the lifespan of C. elegans. Cell 2013, 152, 818–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, X.; Olmedo, M.; Holdorf, A.D.; Shang, Y.; Artal-Sanz, M.; Yilmaz, L.S.; Walhout, A.J.M. A delicate balance between bacterial iron and reactive oxygen species supports optimal C. elegans development. Cell Host Microbe 2019, 26, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Watson, E.; MacNeil, L.T.; Ritter, A.D.; Yilmaz, L.S.; Rosebrock, A.P.; Caudy, A.A.; Walhout, A.J. Interspecies systems biology uncovers metabolites affecting C. elegans gene expression and life history traits. Cell 2014, 156, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Virk, B.; Correia, G.; Dixon, D.P.; Feyst, I.; Jia, J.; Oberleitner, N.; Briggs, Z.; Hodge, E.; Edwards, R.; Ward, J.; et al. Excessive folate synthesis limits lifespan in the C. elegans E. coli aging model. BMC Biol. 2012, 10, 67. [Google Scholar] [CrossRef] [Green Version]
- Ahmadzadeh, L.; Hosseinkhani, A.; Daghigh Kia, H. Effect of supplementing a diet with monensin sodium and Saccharomyces Cerevisiae on reproductive performance of Ghezel ewes. Anim. Reprod. Sci. 2018, 188, 93–100. [Google Scholar] [CrossRef]
- Bass, T.M.; Weinkove, D.; Houthoofd, K.; Gems, D.; Partridge, L. Effects of resveratrol on lifespan in Drosophila melanogaster and Caenorhabditis elegans. Mech. Ageing Dev. 2007, 128, 546–552. [Google Scholar] [CrossRef]
- Beaulieu, M.; Bischofberger, I.; Lorenz, I.; Scheelen, L.; Fischer, K. Reproducing butterflies do not increase intake of antioxidants when they could benefit from them. Biol. Lett. 2016, 12, 20150941. [Google Scholar] [CrossRef]
- Grandison, R.C.; Piper, M.D.; Partridge, L. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature 2009, 462, 1061–1064. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.C.; Kaya, A.; Ma, S.; Kim, G.; Gerashchenko, M.V.; Yim, S.H.; Hu, Z.; Harshman, L.G.; Gladyshev, V.N. Methionine restriction extends lifespan of Drosophila melanogaster under conditions of low amino-acid status. Nat. Commun. 2014, 5, 3592. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.G.; Rogina, B.; Lavu, S.; Howitz, K.; Helfand, S.L.; Tatar, M.; Sinclair, D. Sirtuin activators mimic caloric restriction and delay ageing in metazoans. Nature 2004, 430, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.Y.; Maxon, M.E.; Redfield, B.; Glass, R.; Brot, N.; Weissbach, H. Methionine synthesis in Escherichia coli: Effect of the MetR protein on metE and metH expression. Proc. Natl. Acad. Sci. USA 1989, 86, 4407–4411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helliwell, K.E.; Wheeler, G.L.; Leptos, K.C.; Goldstein, R.E.; Smith, A.G. Insights into the evolution of vitamin B12 Auxotrophy from sequenced algal genomes. Mol. Biol. Evol. 2011, 28, 2921–2933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabreiro, F.; Au, C.; Leung, K.Y.; Vergara-Irigaray, N.; Cocheme, H.M.; Noori, T.; Weinkove, D.; Schuster, E.; Greene, N.D.; Gems, D. Metformin retards aging in C. elegans by altering microbial folate and methionine metabolism. Cell 2013, 153, 228–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redondo-Salvo, S.; Fernandez-Lopez, R.; Ruiz, R.; Vielva, L.; Toro, M.; Rocha, E.P.C.; Garcillan-Barcia, M.P.; Cruz, F. Pathways for horizontal gene transfer in bacteria revealed by a global map of their plasmid. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Partridge, L.; Gems, D.; Withers, D.J. Sex and death: What is the connection? Cell 2005, 120, 461–472. [Google Scholar] [CrossRef]
- Adler, M.I.; Cassidy, E.J.; Fricke, C.; Bonduriansky, R. The lifespan-reproduction trade-off under dietary restriction is sex-specific and context-dependent. Exp. Gerontol. 2013, 48, 539–548. [Google Scholar] [CrossRef]
- Santos, J.; Leitao-Correia, F.; Sousa, M.J.; Leao, C. Diety restriction and nutrient balance in aging. Oxidative Gedicine Cell. Longev. 2016, 2016. [Google Scholar]
- Jiang, Y.; Yan, F.; Lazarovici, P.; Zheng, W. Signaling network of forkhead family of transcription factors (FOXO) in dietary restriction. Cells 2020, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Aguiar-Oliveira, M.H.; Bartke, A. Groeth hormone deficiency: Health and longevity. Endocr. Rev. 2019, 40, 575–601. [Google Scholar] [CrossRef]
- Lushchak, O.V.; Gospodaryov, D.V.; Rovenko, B.M.; Yurkevych, I.S.; Perkhulyn, N.V.; Lushchak, V.I. Specific dietary carbohydrates differentially influence the life span and fecundity of Drosophila melanogaster. J. Gerontol. Ser. A 2014, 69, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, J.M.; Hibshman, J.D.; Webster, A.K.; Kaplan, R.E.W.; Leinroth, A.; Guzman, R.; Maxwell, C.S.; Chitrakar, R.; Bowman, E.A.; Fry, A.L.; et al. Insulin/IGF signaling and vitellogenin provisioning mediate intergenerational adaptation to nutrient stress. Curr. Biol. 2019, 29, 2380–2388. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Li, P.W.; Goldstein, B.; Cai, W.; Thomas, E.L.; Chen, F.; Hubbard, A.E.; Melov, S.; Kapahi, P. Germline signaling mediates the synergistically prolonged longevity produced by double mutations in daf-2 and rsks-1 in C. elegans. Cell Rep. 2013, 5, 1600–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regan, J.C.; Froy, H.; Walling, C.A.; Moatt, J.P.; Nussey, D.H. Dietary restriction and insulin-like signalling pathways as adaptive plasticity: A synthesis and re-evaluatuin. Funct. Ecol. 2020, 34, 107–128. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Lee, H.; Yu, M.; Yeom, E.; Lee, J.; Yoon, A.; Lee, K.; Min, K. Extension of Drosophila lifespan by Korean red ginseng through a mechanism dependent on dSir2 and insulin/IGF-1 signaling. Aging 2019, 11, 9369–9387. [Google Scholar] [CrossRef]
- Hoang, T.T.; Karkhoff-Schweizer, R.R.; Kutchma, A.J.; Schweizer, H.P. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: Application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 1998, 212, 77–86. [Google Scholar] [CrossRef]
- Liu, Z.; Gao, X.; Wang, H.; Fang, H.; Yan, Y.; Liu, L.; Chen, R.; Zhou, D.; Yang, R.; Han, Y. Plasmid pPCP1-derived sRNA HmsA promotes biofilm formation of Yersinia pestis. BMC Microbiol. 2016, 16, 176. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, F.C.; Kalscheur, K.F.; Drackley, J.K. Symposium review: Nutrition strategies for improved health, production, and fertility during the transition period. J. Dairy Sci. 2020, 103, 5684–5693. [Google Scholar] [CrossRef]
- Reda, F.M.; Swelum, A.A.; Hussein, E.O.S.; Elnesr, S.S.; Alhimaidi, A.R.; Alagawany, M. Effects of varying dietary DL-methionine levels on productive and reproductive performance, egg quality, and blood biochemical parameters of quail breeders. Animals 2020, 10, 1839. [Google Scholar] [CrossRef]
- Allai, L.; Benmoula, A.; Maia, S.M.; Nasser, B.; Amiri, B.E. Supplementation of ram semen extender to improve seminal quality and fertility rate. Anim. Reprod. Sci. 2018, 192, 6–17. [Google Scholar] [CrossRef]
- Gao, J.; Guo, H.; Sun, Y.; Ge, F. Differential accumulation of leucine and methionine in red and green pea aphids leads to different fecundity in response to nitrogen fertilization. Pest Manag. Sci. 2018, 74, 1779–1789. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.E.; Elzer, P.H.; Steven Hill, D.; Robertson, G.T.; Farris, M.A.; Roop, R.M.; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- Rose, R.E. The nucleotide sequence of pACYC184. Nucleic Acids Res. 1988, 16, 355. [Google Scholar] [CrossRef] [PubMed]
- Claudi, B.; Sprote, P.; Chirkova, A.; Personnic, N.; Zankl, J.; Schurmann, N.; Schmidt, A.; Bumann, D. Phenotypic variation of Salmonella in host tissues delays eradication by antimicrobial chemotherapy. Cell 2014, 158, 722–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szybalski, W.; Bryson, V. Genetic studies on microbial cross resistance to toxic agents. I. Cross resistance of Escherichia coli to fifteen antibiotics. J. Bacteriol. 1952, 64, 489–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, Y. Quick knowout of aroK and aroL genes from the chromosome of E. coli BW25113 by using Red system. Bull. Acad. Mil. Med. Sci. 2007, 31, 308–311. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Size | Antibiotic Resistance | Replicon | Copy Number Level |
|---|---|---|---|---|
| pEX18Gm | 5831 bp | Gm | pMB1 | high |
| pBBR1MCS-2 | 5144 bp | Km | P15A | high |
| pBBR1MCS-3 | 5228 bp | Tc | P15A | high |
| pBBR1MCS-5 | 4768 bp | Gm | P15A | high |
| pACYC184 | 4245 bp | Cm, Tc | P15A | low |
| pSC101_TIMER | 4946 bp | Km | pSC101 | low |
| R6K_BOX | 1801 bp | Km | R6K | high |
| Strain | Source | Identifier | |
|---|---|---|---|
| Bacterial strain | E. coli OP50 | Caenorhabditis Genetics Center | N/A |
| E. coli OP50 ΔmetH | This work | N/A | |
| E. coli DH5α | Vazyme | Cat#: C502 | |
| E. coli SM10 | Miaolingbio | Cat#: S0049 | |
| E. coli BW25113 | Dharmacon | Cat#: OEC4988 | |
| Salmonella | This work | N/A | |
| Klebsiella | This work | N/A | |
| Escherichia | This work | N/A | |
| Nematode strain | C. elegans N2 | Caenorhabditis Genetics Center | N/A |
| C. elegans Δmetr-1 | C. elegans Gene Knock-out Consortium | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, R.; Li, G.; Lu, L.; Sun, S.; Liu, T.; Li, M.; Zheng, Y.; Walhout, A.J.M.; Wu, J.; Li, H. The Plasmid pEX18Gm Indirectly Increases Caenorhabditis elegans Fecundity by Accelerating Bacterial Methionine Synthesis. Int. J. Mol. Sci. 2022, 23, 5003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095003
Guo R, Li G, Lu L, Sun S, Liu T, Li M, Zheng Y, Walhout AJM, Wu J, Li H. The Plasmid pEX18Gm Indirectly Increases Caenorhabditis elegans Fecundity by Accelerating Bacterial Methionine Synthesis. International Journal of Molecular Sciences. 2022; 23(9):5003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095003
Chicago/Turabian StyleGuo, Rui, Gen Li, Leilei Lu, Shan Sun, Ting Liu, Mengsha Li, Yong Zheng, Albertha J. M. Walhout, Jun Wu, and Huixin Li. 2022. "The Plasmid pEX18Gm Indirectly Increases Caenorhabditis elegans Fecundity by Accelerating Bacterial Methionine Synthesis" International Journal of Molecular Sciences 23, no. 9: 5003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095003