
Molecular Mechanisms of Flavonoids against Tumor Gamma-Herpesviruses and Their Correlated Cancers—A Focus on EBV and KSHV Life Cycles and Carcinogenesis



Abstract
:1. Introduction
2. Flavonoids: An Overview, Anti-Herpesvirus, and Anticancer Properties
3. Flavonoids Target EBV and Its Correlated Cancers
3.1. Flavonoids with Anti-EBV Properties
3.2. Therapeutic Effects of Flavonoids against EBV-Associated Cancers
3.2.1. Flavones and Prenylated Flavonols
3.2.2. Isoflavones
3.2.3. Flavanones
3.2.4. Flavonols
3.2.5. Dihydroflavonols
3.2.6. Chalcones
3.2.7. Other Flavonoid-Type Compounds
4. Flavonoids Target KSHV and Its Correlated Cancers
4.1. Flavonoids with Anti-KSHV Properties
4.2. Therapeutic Effects of Flavonoids against KSHV-Associated Cancers
4.2.1. Flavones
4.2.2. Flavonols
4.2.3. Catechins
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Münz, C. The Role of Lytic Infection for Lymphomagenesis of Human γ-Herpesviruses. Front. Cell. Infect. Microbiol. 2021, 11, 605258. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.W.; Wang, L.; Menke, J.R.; Damania, B. Cancers associated with human gammaherpesviruses. FEBS J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Šudomová, M.; Berchová-Bímová, K.; Marzocco, S.; Liskova, A.; Kubatka, P.; Hassan, S.T. Berberine in Human Oncogenic Herpesvirus Infections and Their Linked Cancers. Viruses 2021, 13, 1014. [Google Scholar] [CrossRef] [PubMed]
- Soldan, S.S.; Lieberman, P.M. Epstein-Barr virus infection in the development of neurological disorders. Drug Discov. Today Dis. Model. 2020, 32, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Bharucha, T.; Houlihan, C.F.; Breuer, J. Herpesvirus Infections of the Central Nervous System. Semin. Neurol. 2019, 39, 369–382. [Google Scholar] [CrossRef]
- Adler, B.; Sattler, C.; Adler, H. Herpesviruses and Their Host Cells: A Successful Liaison. Trends Microbiol. 2017, 25, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, M. Pathogenesis of gammaherpesvirus infections. Vet. Microbiol. 2006, 113, 211–222. [Google Scholar] [CrossRef]
- Azab, W.; Osterrieder, K. Initial Contact: The First Steps in Herpesvirus Entry. Adv. Anat. Embryol. Cell. Biol. 2017, 223, 1–27. [Google Scholar] [CrossRef]
- Connolly, S.A.; Jardetzky, T.S.; Longnecker, R. The structural basis of herpesvirus entry. Nat. Rev. Microbiol. 2021, 19, 110–121. [Google Scholar] [CrossRef]
- Jarosinski, K.W. Interindividual Spread of Herpesviruses. Adv. Anat. Embryol. Cell. Biol. 2017, 223, 195–224. [Google Scholar] [CrossRef]
- Cohen, J.I. Herpesvirus latency. J. Clin. Investig. 2020, 130, 3361–3369. [Google Scholar] [CrossRef] [PubMed]
- Speck, S.H.; Ganem, D. Viral Latency and Its Regulation: Lessons from the γ-Herpesviruses. Cell Host Microbe 2010, 8, 100–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenzie, J.; El-Guindy, A. Epstein-Barr Virus Lytic Cycle Reactivation. Curr. Top. Microbiol. Immunol. 2015, 391, 237–261. [Google Scholar] [CrossRef] [PubMed]
- Broussard, G.; Damania, B. Regulation of KSHV Latency and Lytic Reactivation. Viruses 2020, 12, 1034. [Google Scholar] [CrossRef]
- Jondle, C.N.; Tarakanova, V.L. Innate immunity and alpha/gammaherpesviruses: First impressions last a lifetime. Curr. Opin. Virol. 2020, 44, 81–89. [Google Scholar] [CrossRef]
- Lange, P.T.; White, M.C.; Damania, B. Activation and Evasion of Innate Immunity by Gammaherpesviruses. J. Mol. Biol. 2021, 434, 167214. [Google Scholar] [CrossRef]
- Wołącewicz, M.; Becht, R.; Grywalska, E.; Niedźwiedzka-Rystwej, P. Herpesviruses in Head and Neck Cancers. Viruses 2020, 12, 172. [Google Scholar] [CrossRef] [Green Version]
- Poole, C.L.; James, S.H. Antiviral Therapies for Herpesviruses: Current Agents and New Directions. Clin. Ther. 2018, 40, 1282–1298. [Google Scholar] [CrossRef] [Green Version]
- Kłysik, K.; Pietraszek, A.; Karewicz, A.; Nowakowska, M. Acyclovir in the Treatment of Herpes Viruses—A Review. Curr. Med. Chem. 2020, 27, 4118–4137. [Google Scholar] [CrossRef]
- Chen, E.Y.; Raghunathan, V.; Prasad, V. An Overview of Cancer Drugs Approved by the US Food and Drug Administration Based on the Surrogate End Point of Response Rate. JAMA Intern. Med. 2019, 179, 915–921. [Google Scholar] [CrossRef]
- Vasan, N.; Baselga, J.; Hyman, D.M. A view on drug resistance in cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šudomová, M.; Hassan, S. Nutraceutical Curcumin with Promising Protection against Herpesvirus Infections and Their Associated Inflammation: Mechanisms and Pathways. Microorganisms 2021, 9, 292. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; International Natural Product Sciences Taskforce; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.T.S.; Šudomová, M.; Mazurakova, A.; Kubatka, P. Insights into Antiviral Properties and Molecular Mechanisms of Non-Flavonoid Polyphenols against Human Herpesviruses. Int. J. Mol. Sci. 2022, 23, 13891. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.G.A.; Silva, A.M.S. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Petrussa, E.; Braidot, E.; Zancani, M.; Peresson, C.; Bertolini, A.; Patui, S.; Vianello, A. Plant Flavonoids—Biosynthesis, Transport and Involvement in Stress Responses. Int. J. Mol. Sci. 2013, 14, 14950–14973. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Teng, H.; Chen, L. Polyphenols and bioavailability: An update. Crit. Rev. Food Sci. Nutr. 2019, 59, 2040–2051. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A concise overview on the chemistry, occurrence, and human health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.; Kumar, A.; Malik, A.K. Flavonoids biosynthesis in plants and its further analysis by capillary electrophoresis. Electrophoresis 2017, 38, 820–832. [Google Scholar] [CrossRef]
- Treutter, D. Significance of Flavonoids in Plant Resistance and Enhancement of Their Biosynthesis. Plant Biol. 2005, 7, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.; Fang, X.; Yang, J.; Yao, Y.; Nandakumar, K.S.; Salem, M.L.; Cheng, K. Recent Research on Flavonoids and their Biomedical Applications. Curr. Med. Chem. 2021, 28, 1042–1066. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.T.S.; Masarčíková, R.; Berchová-Bímová, K. Bioactive natural products with anti-herpes simplex virus properties. J. Pharm. Pharmacol. 2015, 67, 1325–1336. [Google Scholar] [CrossRef]
- Treml, J.; Gazdová, M.; Šmejkal, K.; Šudomová, M.; Kubatka, P.; Hassan, S.T.S. Natural Products-Derived Chemicals: Breaking Barriers to Novel Anti-HSV Drug Development. Viruses 2020, 12, 154. [Google Scholar] [CrossRef] [Green Version]
- Šudomová, M.; Berchová-Bímová, K.; Mazurakova, A.; Šamec, D.; Kubatka, P.; Hassan, S.T.S. Flavonoids Target Human Herpesviruses That Infect the Nervous System: Mechanisms of Action and Therapeutic Insights. Viruses 2022, 14, 592. [Google Scholar] [CrossRef]
- Liskova, A.; Samec, M.; Koklesova, L.; Brockmueller, A.; Zhai, K.; Abdellatif, B.; Siddiqui, M.; Biringer, K.; Kudela, E.; Pec, M.; et al. Flavonoids as an effective sensitizer for anti-cancer therapy: Insights into multi-faceted mechanisms and applicability towards individualized patient profiles. EPMA J. 2021, 12, 155–176. [Google Scholar] [CrossRef]
- Samec, M.; Liskova, A.; Koklesova, L.; Mersakova, S.; Strnadel, J.; Kajo, K.; Pec, M.; Zhai, K.; Smejkal, K.; Mirzaei, S.; et al. Flavonoids Targeting HIF-1: Implications on Cancer Metabolism. Cancers 2021, 13, 130. [Google Scholar] [CrossRef]
- Liskova, A.; Koklesova, L.; Samec, M.; Smejkal, K.; Samuel, S.M.; Varghese, E.; Abotaleb, M.; Biringer, K.; Kudela, E.; Danko, J.; et al. Flavonoids in Cancer Metastasis. Cancers 2020, 12, 1498. [Google Scholar] [CrossRef] [PubMed]
- Bisol, Â.; De Campos, P.S.; Lamers, M.L. Flavonoids as anticancer therapies: A systematic review of clinical trials. Phytother. Res. 2020, 34, 568–582. [Google Scholar] [CrossRef] [PubMed]
- Nowalk, A.; Green, M. Epstein-Barr Virus. Microbiol. Spectr. 2016, 4, 127–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciccarese, G.; Trave, I.; Herzum, A.; Parodi, A.; Drago, F. Dermatological manifestations of Epstein-Barr virus systemic infection: A case report and literature review. Int. J. Dermatol. 2020, 59, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Tsao, S.W.; Tsang, C.M.; Lo, K.W. Epstein–Barr virus infection and nasopharyngeal carcinoma. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160270. [Google Scholar] [CrossRef] [Green Version]
- Farrell, P.J. Epstein–Barr Virus and Cancer. Annu. Rev. Pathol. 2019, 14, 29–53. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Z.; Zeng, B.; Hu, G.; Gan, R. Epstein–Barr virus-associated gastric cancer: A distinct subtype. Cancer Lett. 2020, 495, 191–199. [Google Scholar] [CrossRef]
- Murray, P.G.; Young, L.S. An etiological role for the Epstein-Barr virus in the pathogenesis of classical Hodgkin lymphoma. Blood 2019, 134, 591–596. [Google Scholar] [CrossRef]
- Naughton, P.; Healy, M.; Enright, F.; Lucey, B. Infectious Mononucleosis: Diagnosis and clinical interpretation. Br. J. Biomed. Sci. 2021, 78, 107–116. [Google Scholar] [CrossRef]
- Münz, C. Latency and lytic replication in Epstein–Barr virus-associated oncogenesis. Nat. Rev. Microbiol. 2019, 17, 691–700. [Google Scholar] [CrossRef]
- Yin, H.; Qu, J.; Peng, Q.; Gan, R. Molecular mechanisms of EBV-driven cell cycle progression and oncogenesis. Med Microbiol. Immunol. 2019, 208, 573–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosemarie, Q.; Sugden, B. Epstein–Barr Virus: How Its Lytic Phase Contributes to Oncogenesis. Microorganisms 2020, 8, 1824. [Google Scholar] [CrossRef] [PubMed]
- Kutok, J.; Wang, F. SPECTRUM OF EPSTEIN-BARR VIRUS–ASSOCIATED DISEASES. Annu. Rev. Pathol. 2006, 1, 375–404. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, S.; Hu, J.; Luo, X.; Li, N.; Bode, A.M.; Cao, Y. Epstein-Barr virus lytic reactivation regulation and its pathogenic role in carcinogenesis. Int. J. Biol. Sci. 2016, 12, 1309–1318. [Google Scholar] [CrossRef] [Green Version]
- Tsai, Y.-C.; Hohmann, J.; El-Shazly, M.; Chang, L.-K.; Dankó, B.; Kúsz, N.; Hsieh, C.-T.; Hunyadi, A.; Chang, F.-R. Bioactive constituents of Lindernia crustacea and its anti-EBV effect via Rta expression inhibition in the viral lytic cycle. J. Ethnopharmacol. 2020, 250, 112493. [Google Scholar] [CrossRef]
- Wu, C.-C.; Fang, C.-Y.; Hsu, H.-Y.; Chen, Y.-J.; Chou, S.-P.; Huang, S.-Y.; Cheng, Y.-J.; Lin, S.-F.; Chang, Y.; Tsai, C.-H.; et al. Luteolin inhibits Epstein-Barr virus lytic reactivation by repressing the promoter activities of immediate-early genes. Antivir. Res. 2016, 132, 99–110. [Google Scholar] [CrossRef]
- Wu, C.-C.; Fang, C.-Y.; Cheng, Y.-J.; Hsu, H.-Y.; Chou, S.-P.; Huang, S.-Y.; Tsai, C.-H.; Chen, J.-Y. Inhibition of Epstein-Barr virus reactivation by the flavonoid apigenin. J. Biomed. Sci. 2017, 24, 2. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Koike, R.; Yamamoto, A.; Ukiya, M.; Fukatsu, M.; Banno, N.; Miura, M.; Motohashi, S.; Tokuda, H.; Akihisa, T. Glycosidic Inhibitors of Melanogenesis from Leaves of Passiflora edulis. Chem. Biodivers. 2013, 10, 1851–1865. [Google Scholar] [CrossRef]
- Tung, C.-P.; Chang, F.-R.; Wu, Y.-C.; Chuang, D.-W.; Hunyadi, A.; Liu, S.-T. Inhibition of the Epstein–Barr virus lytic cycle by protoapigenone. J. Gen. Virol. 2011, 92, 1760–1768. [Google Scholar] [CrossRef]
- Vágvölgyi, M.; Girst, G.; Kúsz, N.; Ötvös, S.B.; Fülöp, F.; Hohmann, J.; Servais, J.-Y.; Seguin-Devaux, C.; Chang, F.-R.; Chen, M.S.; et al. Less Cytotoxic Protoflavones as Antiviral Agents: Protoapigenone 1′-O-isopropyl ether Shows Improved Selectivity Against the Epstein–Barr Virus Lytic Cycle. Int. J. Mol. Sci. 2019, 20, 6269. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, W.-F.; Xu, J.; Kitdamrongtham, W.; Manosroi, A.; Manosroi, J.; Tokuda, H.; Abe, M.; Akihisa, T.; Feng, F. Potential cancer chemopreventive and anticancer constituents from the fruits of Ficus hispida L.f. (Moraceae). J. Ethnopharmacol. 2018, 214, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Ito, C.; Matsui, T.; Miyabe, K.; Hasan, C.M.; Rashid, M.A.; Tokuda, H.; Itoigawa, M. Three isoflavones from Derris scandens (Roxb.) Benth and their cancer chemopreventive activity and in vitro antiproliferative effects. Phytochemistry 2020, 175, 112376. [Google Scholar] [CrossRef] [PubMed]
- Jakhmola, S.; Hazarika, Z.; Jha, A.N.; Jha, H.C. In silico analysis of antiviral phytochemicals efficacy against Epstein–Barr virus glycoprotein H. J. Biomol. Struct. Dyn. 2022, 40, 5372–5385. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.T.; Seca, H.; Palmeira, A.; Fernandes, M.X.; Castro, F.; Correia-Da-Silva, M.; Nascimento, M.S.J.; Sousa, M.E.; Pinto, M.; Vasconcelos, M.H. Sulfated Small Molecules Targeting EBV in Burkitt Lymphoma: FromIn SilicoScreening to the Evidence ofIn VitroEffect on Viral Episomal DNA. Chem. Biol. Drug Des. 2013, 81, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Akazawa, H.; Kohno, H.; Tokuda, H.; Suzuki, N.; Yasukawa, K.; Kimura, Y.; Manosroi, A.; Manosroi, J.; Akihisa, T. Anti-Inflammatory and Anti-Tumor-Promoting Effects of 5-Deprenyllupulonol C and Other Compounds from Hop (Humulus lupulus L.). Chem. Biodivers. 2012, 9, 1045–1054. [Google Scholar] [CrossRef]
- Akihisa, T.; Motoi, T.; Seki, A.; Kikuchi, T.; Fukatsu, M.; Tokuda, H.; Suzuki, N.; Kimura, Y. Cytotoxic Activities and Anti-Tumor-Promoting Effects of Microbial Transformation Products of Prenylated Chalcones from Angelica keiskei. Chem. Biodivers. 2012, 9, 318–330. [Google Scholar] [CrossRef]
- Liu, S.; Li, H.; Chen, L.; Yang, L.; Li, L.; Tao, Y.; Li, W.; Li, Z.; Liu, H.; Tang, M.; et al. (-)-Epigallocatechin-3-gallate inhibition of Epstein-Barr virus spontaneous lytic infection involves ERK1/2 and PI3-K/Akt signaling in EBV-positive cells. Carcinogenesis 2013, 34, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Li, H.; Tang, M.; Cao, Y. (-)-Epigallocatechin-3-gallate inhibition of Epstein-Barr virus spontaneous lytic infection involves downregulation of latent membrane protein 1. Exp. Ther. Med. 2018, 15, 1105–1112. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, Y.; Hu, J.; Liu, S.; Luo, X.; Tang, M.; Bode, A.M.; Dong, Z.; Liu, X.; Liao, W.; et al. ()-Epigallocatechin-3-Gallate Inhibits EBV Lytic Replication via Targeting LMP1-Mediated MAPK Signal Axes. Oncol. Res. 2021, 28, 763–778. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Tsai, H.-L.; Peng, C.-W. EGCG debilitates the persistence of EBV latency by reducing the DNA binding potency of nuclear antigen 1. Biochem. Biophys. Res. Commun. 2012, 417, 1093–1099. [Google Scholar] [CrossRef]
- Jakhmola, S.; Jonniya, N.A.; Sk, F.; Rani, A.; Kar, P.; Jha, H.C. Identification of Potential Inhibitors against Epstein–Barr Virus Nuclear Antigen 1 (EBNA1): An Insight from Docking and Molecular Dynamic Simulations. ACS Chem. Neurosci. 2021, 12, 3060–3072. [Google Scholar] [CrossRef] [PubMed]
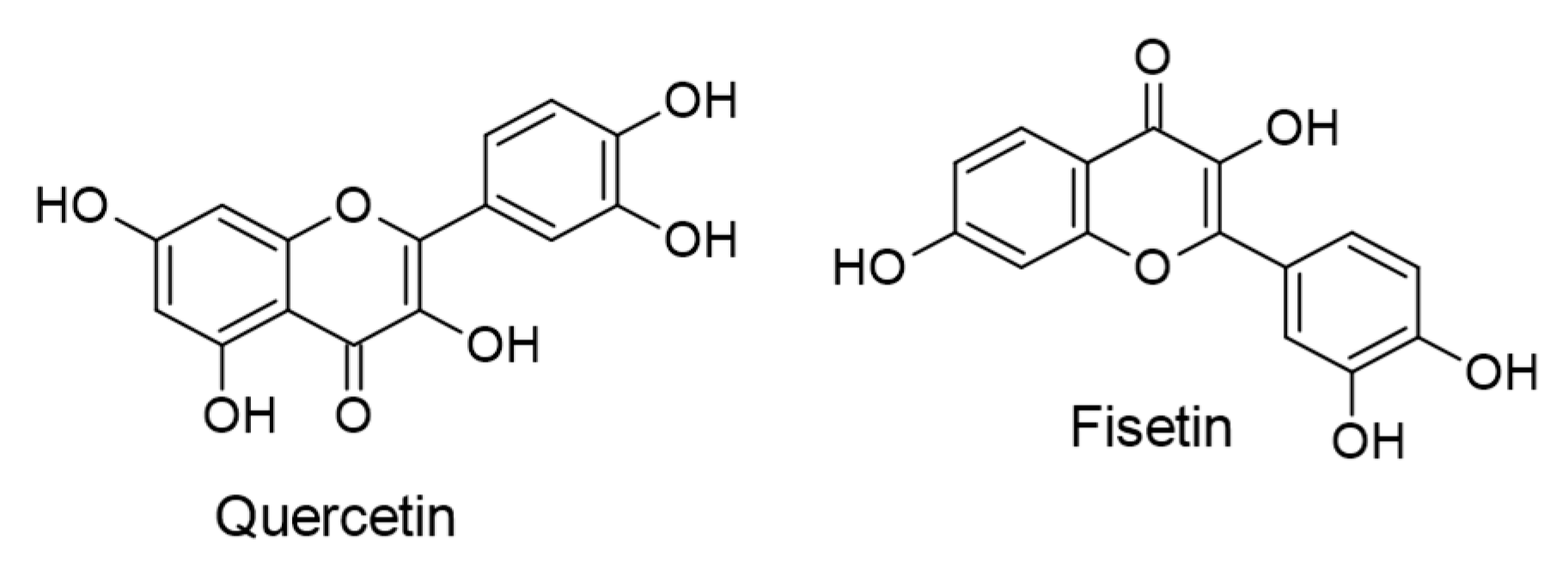
- Lee, M.; Son, M.; Ryu, E.; Shin, Y.S.; Kim, J.G.; Kang, B.W.; Sung, G.-H.; Cho, H.; Kang, H. Quercetin-induced apoptosis prevents EBV infection. Oncotarget 2015, 6, 12603–12624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, S.; Lee, S.; Choi, S.J.; Wu, Z.; Cho, J.-H.; Kim, L.; Shin, Y.S.; Kang, B.W.; Kim, J.G.; Liu, K.; et al. Quercetin Synergistically Inhibit EBV-Associated Gastric Carcinoma with Ganoderma lucidum Extracts. Molecules 2019, 24, 3834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, C.; Itoigawa, M.; Tan, H.T.; Tokuda, H.; Mou, X.Y.; Mukainaka, T.; Ishikawa, T.; Nishino, H.; Furukawa, H. Anti-tumor-promoting effects of isoflavonoids on Epstein–Barr virus activation and two-stage mouse skin carcinogenesis. Cancer Lett. 2000, 152, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Zhao, Y.; Rasheed, T.; Ahmed, I.; Hassan, S.T.; Nawaz, M.Z.; Iqbal, H.M. Biogenic Nanoparticle-Chitosan Conjugates with Antimicrobial, Antibiofilm, and Anticancer Potentialities: Development and Characterization. Int. J. Environ. Res. Public Health 2019, 16, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, R.; Tsujii, H.; Yamada, T.; Kajimoto, T.; Tokuda, H.; Arai, T.; Suzuki, N.; Hasegawa, J.; Hamashima, Y.; Node, M. Conjugates of 3α-methoxyserrat-14-en-21β-ol (PJ-1) and 3β-methoxyserrat-14-en-21β-ol (PJ-2) as cancer chemopreventive agents. Eur. J. Med. Chem. 2011, 46, 3368–3375. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Kong, S.; Song, F.; Li, L.; Jiang, H. Pharmacokinetic study of luteolin, apigenin, chrysoeriol and diosmetin after oral administration of Flos Chrysanthemi extract in rats. Fitoterapia 2012, 83, 1616–1622. [Google Scholar] [CrossRef] [PubMed]
- Fasoulakis, Z.; Koutras, A.; Syllaios, A.; Schizas, D.; Garmpis, N.; Diakosavvas, M.; Angelou, K.; Tsatsaris, G.; Pagkalos, A.; Ntounis, T.; et al. Breast Cancer Apoptosis and the Therapeutic Role of Luteolin. Chirurgia 2021, 116, 170–177. [Google Scholar] [CrossRef]
- Fan, W.; Qian, S.; Qian, P.; Li, X. Antiviral activity of luteolin against Japanese encephalitis virus. Virus Res. 2016, 220, 112–116. [Google Scholar] [CrossRef]
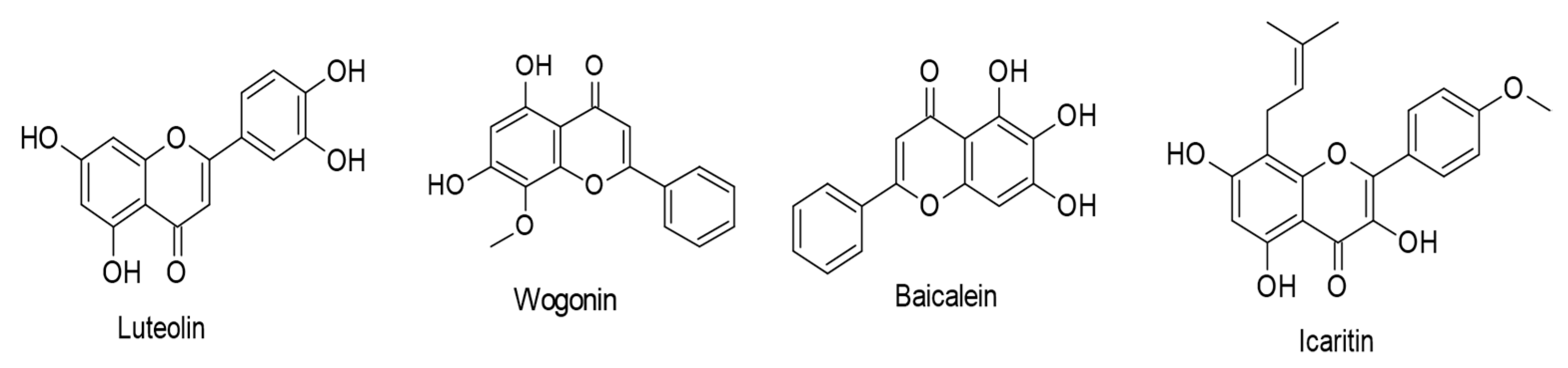
- Wu, C.-C.; Fang, C.-Y.; Hsu, H.-Y.; Chuang, H.-Y.; Cheng, Y.-J.; Chen, Y.-J.; Chou, S.-P.; Huang, S.-Y.; Lin, S.-F.; Chang, Y.; et al. EBV reactivation as a target of luteolin to repress NPC tumorigenesis. Oncotarget 2016, 7, 18999–19017. [Google Scholar] [CrossRef]
- Lo, A.K.-F.; Dawson, C.W.; Lung, H.L.; Wong, K.-L.; Young, L.S. The Role of EBV-Encoded LMP1 in the NPC Tumor Microenvironment: From Function to Therapy. Front. Oncol. 2021, 11, 640207. [Google Scholar] [CrossRef] [PubMed]
- Lo, A.K.-F.; Lung, R.W.-M.; Dawson, C.W.; Young, L.S.; Ko, C.-W.; Yeung, W.W.; Kang, W.; To, K.-F.; Lo, K.-W. Activation of sterol regulatory element-binding protein 1 (SREBP1)-mediated lipogenesis by the Epstein-Barr virus-encoded latent membrane protein 1 (LMP1) promotes cell proliferation and progression of nasopharyngeal carcinoma. J. Pathol. 2018, 246, 180–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Liu, P.; Zhang, H.; Li, Y.; Salmani, J.M.M.; Wang, F.; Yang, K.; Fu, R.; Chen, Z.; Chen, B. Wogonin as a targeted therapeutic agent for EBV (+) lymphoma cells involved in LMP1/NF-κB/miR-155/PU.1 pathway. BMC Cancer 2017, 17, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, H.; Liu, Y.; Wang, C.; Wang, J.; Long, C.; Guo, W.; Sun, X. Baicalein inhibits growth of Epstein-Barr virus-positive nasopharyngeal carcinoma by repressing the activity of EBNA1 Q-promoter. Biomed. Pharmacother. 2018, 102, 1003–1014. [Google Scholar] [CrossRef]
- Bin Park, G.; Kim, Y.S.; Lee, H.-K.; Yang, J.W.; Kim, D.; Hur, D.Y. ASK1/JNK-mediated TAp63 activation controls the cell survival signal of baicalein-treated EBV-transformed B cells. Mol. Cell. Biochem. 2016, 412, 247–258. [Google Scholar] [CrossRef]
- Wu, T.; Wang, S.; Wu, J.; Lin, Z.; Sui, X.; Xu, X.; Shimizu, N.; Chen, B.; Wang, X. Icaritin induces lytic cytotoxicity in extranodal NK/T-cell lymphoma. J. Exp. Clin. Cancer Res. 2015, 34, 17. [Google Scholar] [CrossRef] [Green Version]
- David, A.V.A.; Arulmoli, R.; Parasuraman, S. Overviews of biological importance of quercetin: A bioactive flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Farias, M.; Carrasco-Pozo, C. The Anti-Cancer Effect of Quercetin: Molecular Implications in Cancer Metabolism. Int. J. Mol. Sci. 2019, 20, 3177. [Google Scholar] [CrossRef] [Green Version]
- Di Petrillo, A.; Orrù, G.; Fais, A.; Fantini, M.C. Quercetin and its derivates as antiviral potentials: A comprehensive review. Phytother. Res. 2022, 36, 266–278. [Google Scholar] [CrossRef]
- Lee, H.H.; Lee, S.; Shin, Y.S.; Cho, M.; Kang, H.; Cho, H. Anti-Cancer Effect of Quercetin in Xenograft Models with EBV-Associated Human Gastric Carcinoma. Molecules 2016, 21, 1286. [Google Scholar] [CrossRef]
- Granato, M.; Rizzello, C.; Romeo, M.A.; Yadav, S.; Santarelli, R.; D’Orazi, G.; Faggioni, A.; Cirone, M. Concomitant reduction of c-Myc expression and PI3K/AKT/mTOR signaling by quercetin induces a strong cytotoxic effect against Burkitt’s lymphoma. Int. J. Biochem. Cell Biol. 2016, 79, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Daker, M.; Bhuvanendran, S.; Ahmad, M.; Takada, K.; Khoo, A.S.-B. Deregulation of lipid metabolism pathway genes in nasopharyngeal carcinoma cells. Mol. Med. Rep. 2013, 7, 731–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iranshahi, M.; Sahebkar, A.; Hosseini, S.; Takasaki, M.; Konoshima, T.; Tokuda, H. Cancer chemopreventive activity of diversin from Ferula diversivittata in vitro and in vivo. Phytomedicine 2010, 17, 269–273. [Google Scholar] [CrossRef]
- Granato, M.; Montani, M.S.G.; Zompetta, C.; Santarelli, R.; Gonnella, R.; Romeo, M.A.; D’Orazi, G.; Faggioni, A.; Cirone, M. Quercetin Interrupts the Positive Feedback Loop Between STAT3 and IL-6, Promotes Autophagy, and Reduces ROS, Preventing EBV-Driven B Cell Immortalization. Biomolecules 2019, 9, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Syed, D.N.; Ahmad, N.; Mukhtar, H. Fisetin: A Dietary Antioxidant for Health Promotion. Antioxid. Redox Signal. 2013, 19, 151–162. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Naureen, H.; Zahid, R.; Youssef, L.; Attar, R.; Xu, B. Cancer chemopreventive role of fisetin: Regulation of cell signaling pathways in different cancers. Pharmacol. Res. 2021, 172, 105784. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhao, Y.; Chen, J.; Shao, S.; Zhang, X. Fisetin inhibits migration, invasion and epithelial-mesenchymal transition of LMP1-positive nasopharyngeal carcinoma cells. Mol. Med. Rep. 2014, 9, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Liang, H.-Y.; Li, M.-Y.; Lin, C.-Y.; Shi, M.-J.; Zhang, X.-J. Interference of Fisetin with Targets of the Nuclear Factor-κB Signal Transduction Pathway Activated by Epstein-Barr Virus Encoded Latent Membrane Protein 1. Asian Pac. J. Cancer Prev. 2014, 15, 9835–9839. [Google Scholar] [CrossRef] [Green Version]
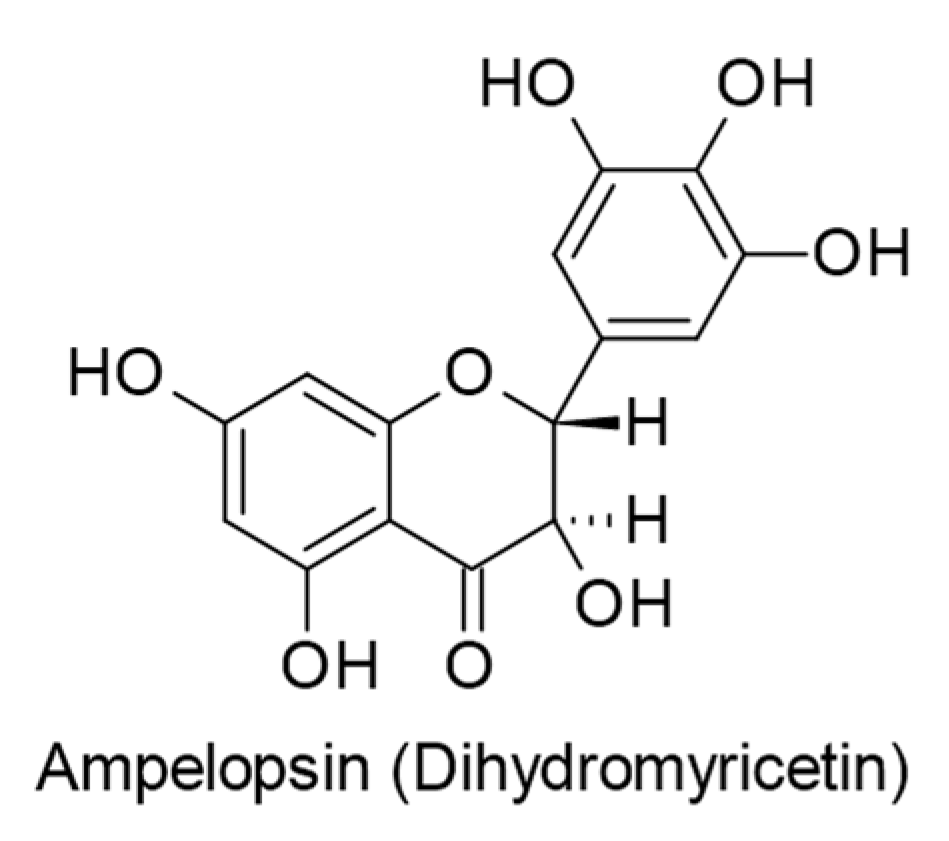
- Yun, S.-M.; Kim, Y.S.; Kim, K.H.; Hur, D.Y. Ampelopsin Induces DR5-Mediated Apoptotic Cell Death in EBV-Infected Cells through the p38 Pathway. Nutr. Cancer 2020, 72, 489–494. [Google Scholar] [CrossRef]
- Wang, K.-L.; Yu, Y.-C.; Hsia, S.-M. Perspectives on the Role of Isoliquiritigenin in Cancer. Cancers 2021, 13, 115. [Google Scholar] [CrossRef]
- Zhao, T.-T.; Xu, Y.-Q.; Hu, H.-M.; Gong, H.-B.; Zhu, H.-L. Isoliquiritigenin (ISL) and its Formulations: Potential Antitumor Agents. Curr. Med. Chem. 2019, 26, 6786–6796. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-E.; Hong, E.-J.; Nam, H.-Y.; Hwang, M.; Kim, J.-H.; Han, B.-G.; Jeon, J.-P. Molecular signatures in response to Isoliquiritigenin in lymphoblastoid cell lines. Biochem. Biophys. Res. Commun. 2012, 427, 392–397. [Google Scholar] [CrossRef] [PubMed]
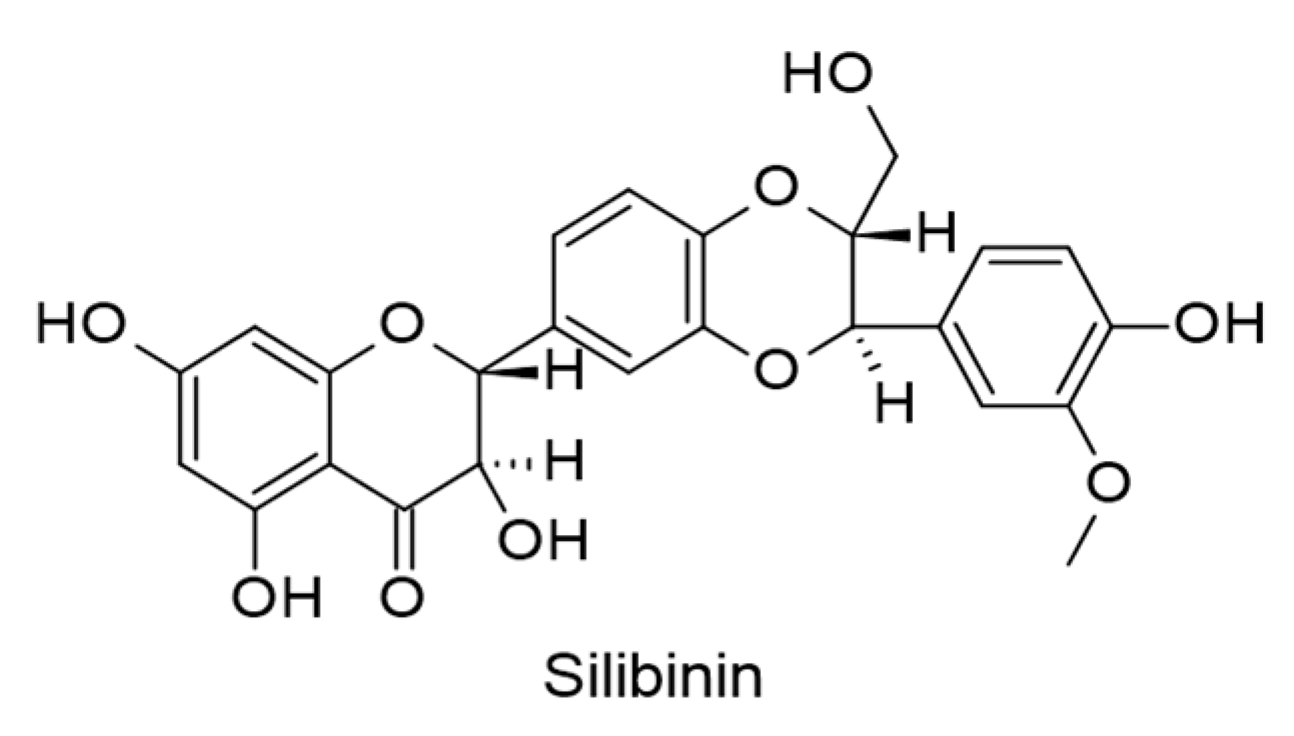
- Sellam, L.S.; Zappasodi, R.; Chettibi, F.; Djennaoui, D.; Mesbah, N.Y.-A.; Amir-Tidadini, Z.-C.; Touil-Boukoffa, C.; Ouahioune, W.; Merghoub, T.; Bourouba, M. Silibinin down-regulates PD-L1 expression in nasopharyngeal carcinoma by interfering with tumor cell glycolytic metabolism. Arch. Biochem. Biophys. 2020, 690, 108479. [Google Scholar] [CrossRef]
- Gaglia, M.M. Kaposi’s sarcoma-associated herpesvirus at 27. Tumour Virus Res. 2021, 12, 200223. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, P.H.; Ziegelbauer, J.; Uldrick, T.S.; Yarchoan, R. Kaposi sarcoma herpesvirus-associated cancers and related diseases. Curr. Opin. HIV AIDS 2017, 12, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Bai, L.; Dong, J.; Sun, R.; Lan, K. Kaposi’s Sarcoma-Associated Herpesvirus: Epidemiology and Molecular Biology. In Infectious Agents Associated Cancers: Epidemiology and Molecular Biology; Cai, Q., Yuan, Z., Lan, K., Eds.; Advances in Experimental Medicine and Biology; Springer Singapore: Singapore, 2017; Volume 1018, pp. 91–127. ISBN 978-981-10-5764-9. [Google Scholar]
- Aneja, K.K.; Yuan, Y. Reactivation and Lytic Replication of Kaposi’s Sarcoma-Associated Herpesvirus: An Update. Front. Microbiol. 2017, 8, 613. [Google Scholar] [CrossRef] [Green Version]
- Cesarman, E.; Damania, B.; Krown, S.E.; Martin, J.; Bower, M.; Whitby, D. Kaposi sarcoma. Nat. Rev. Dis. Prim. 2019, 5, 9. [Google Scholar] [CrossRef]
- Mesri, E.A.; Cesarman, E.; Boshoff, C. Kaposi’s sarcoma and its associated herpesvirus. Nat. Rev. Cancer 2010, 10, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Giffin, L.; Damania, B. KSHV: Pathways to Tumorigenesis and Persistent Infection. Adv. Virus Res. 2014, 88, 111–159. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Majerciak, V.; Zheng, Z.-M.; Lan, K. Towards Better Understanding of KSHV Life Cycle: From Transcription and Posttranscriptional Regulations to Pathogenesis. Virol. Sin. 2019, 34, 135–161. [Google Scholar] [CrossRef]
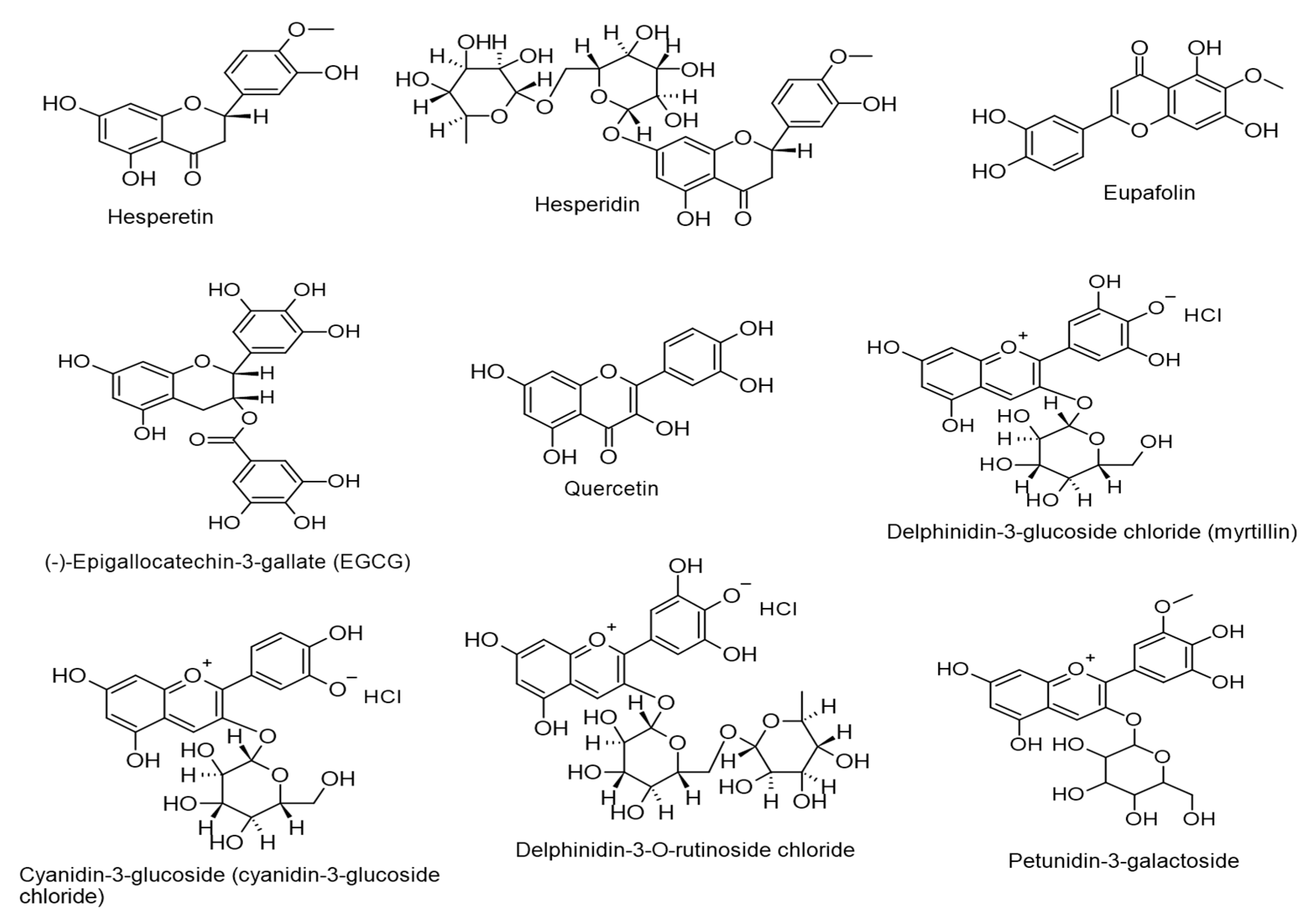
- Long, W.-Y.; Zhao, G.-H.; Wu, Y. Hesperetin inhibits KSHV reactivation and is reversed by HIF1α overexpression. J. Gen. Virol. 2021, 102, 001686. [Google Scholar] [CrossRef]
- Mekni-Toujani, M.; Mousavizadeh, L.; Gallo, A.; Ghram, A. Thymus capitatus flavonoids inhibit infection of Kaposi’s sarcoma-associated herpesvirus. FEBS Open Bio 2022, 12, 1166–1177. [Google Scholar] [CrossRef]
- Tsai, C.-Y.; Chen, C.-Y.; Chiou, Y.-H.; Shyu, H.-W.; Lin, K.-H.; Chou, M.-C.; Huang, M.-H.; Wang, Y.-F. Epigallocatechin-3-Gallate Suppresses Human Herpesvirus 8 Replication and Induces ROS Leading to Apoptosis and Autophagy in Primary Effusion Lymphoma Cells. Int. J. Mol. Sci. 2017, 19, 16. [Google Scholar] [CrossRef] [Green Version]
- Granato, M.; Montani, M.S.G.; Angiolillo, C.; D’Orazi, G.; Faggioni, A.; Cirone, M. Cytotoxic Drugs Activate KSHV Lytic Cycle in Latently Infected PEL Cells by Inducing a Moderate ROS Increase Controlled by HSF1, NRF2 and p62/SQSTM1. Viruses 2018, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- Sivarajan, R.; Oberwinkler, H.; Roll, V.; König, E.-M.; Steinke, M.; Bodem, J. A defined anthocyanin mixture sourced from bilberry and black currant inhibits Measles virus and various herpesviruses. BMC Complement. Med. Ther. 2022, 22, 181. [Google Scholar] [CrossRef]
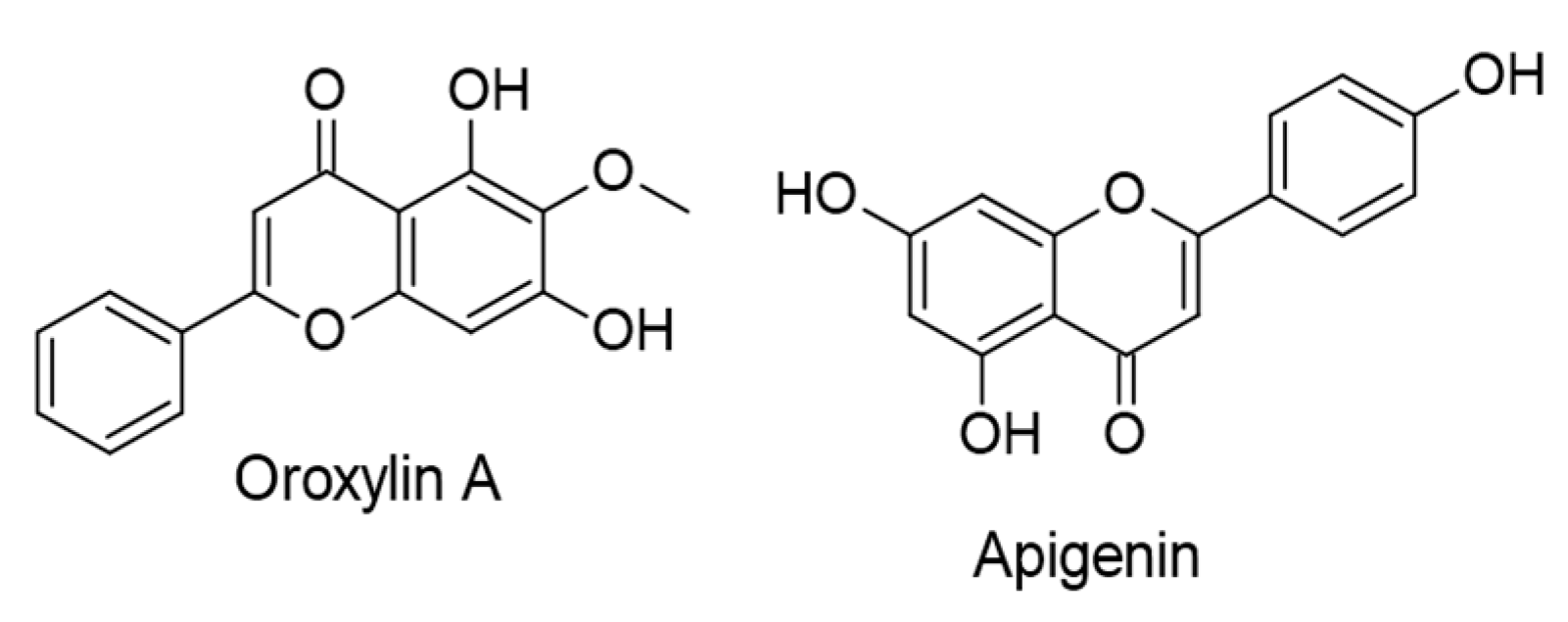
- Zhu, X.; Chen, Y.; Zhu, W.; Ji, M.; Xu, J.; Guo, Y.; Gao, F.; Gu, W.; Yang, X.; Zhang, C. Oroxylin A inhibits Kaposi’s sarcoma-associated herpes virus (KSHV) vIL-6-mediated lymphatic reprogramming of vascular endothelial cells through modulating PPARγ/Prox1 axis. J. Med Virol. 2019, 91, 463–472. [Google Scholar] [CrossRef]
- Granato, M.; Gilardini Montani, M.S.; Santarelli, R.; D’Orazi, G.; Faggioni, A.; Cirone, M. Apigenin, by activating p53 and inhibiting STAT3, modulates the balance between pro-apoptotic and pro-survival pathways to induce PEL cell death. J. Exp. Clin. Cancer Res. 2017, 36, 167. [Google Scholar] [CrossRef] [Green Version]
- Granato, M.; Rizzello, C.; Gilardini Montani, M.S.; Cuomo, L.; Vitillo, M.; Santarelli, R.; Gonnella, R.; D’Orazi, G.; Faggioni, A.; Cirone, M. Quercetin induces apoptosis and autophagy in primary effusion lymphoma cells by inhibiting PI3K/AKT/mTOR and STAT3 signaling pathways. J. Nutr. Biochem. 2017, 41, 124–136. [Google Scholar] [CrossRef]
- Gonnella, R.; Yadav, S.; Montani, M.S.G.; Granato, M.; Santarelli, R.; Garufi, A.; D’Orazi, G.; Faggioni, A.; Cirone, M. Oxidant species are involved in T/B-mediated ERK1/2 phosphorylation that activates p53-p21 axis to promote KSHV lytic cycle in PEL cells. Free Radic. Biol. Med. 2017, 112, 327–335. [Google Scholar] [CrossRef]
- Yeh, L.-C.; Shyu, H.-W.; Jin, Y.-R.; Chiou, Y.-H.; Lin, K.-H.; Chou, M.-C.; Huang, M.-H.; Wang, Y.-F. Epigallocatechin-3-gallate downregulates PDHA1 interfering the metabolic pathways in human herpesvirus 8 harboring primary effusion lymphoma cells. Toxicol. Vitr. 2020, 65, 104753. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound, Concentration, and Source | Chemical Class | Molecular Mechanisms (Inhibition/Downregulation) | Reference |
|---|---|---|---|
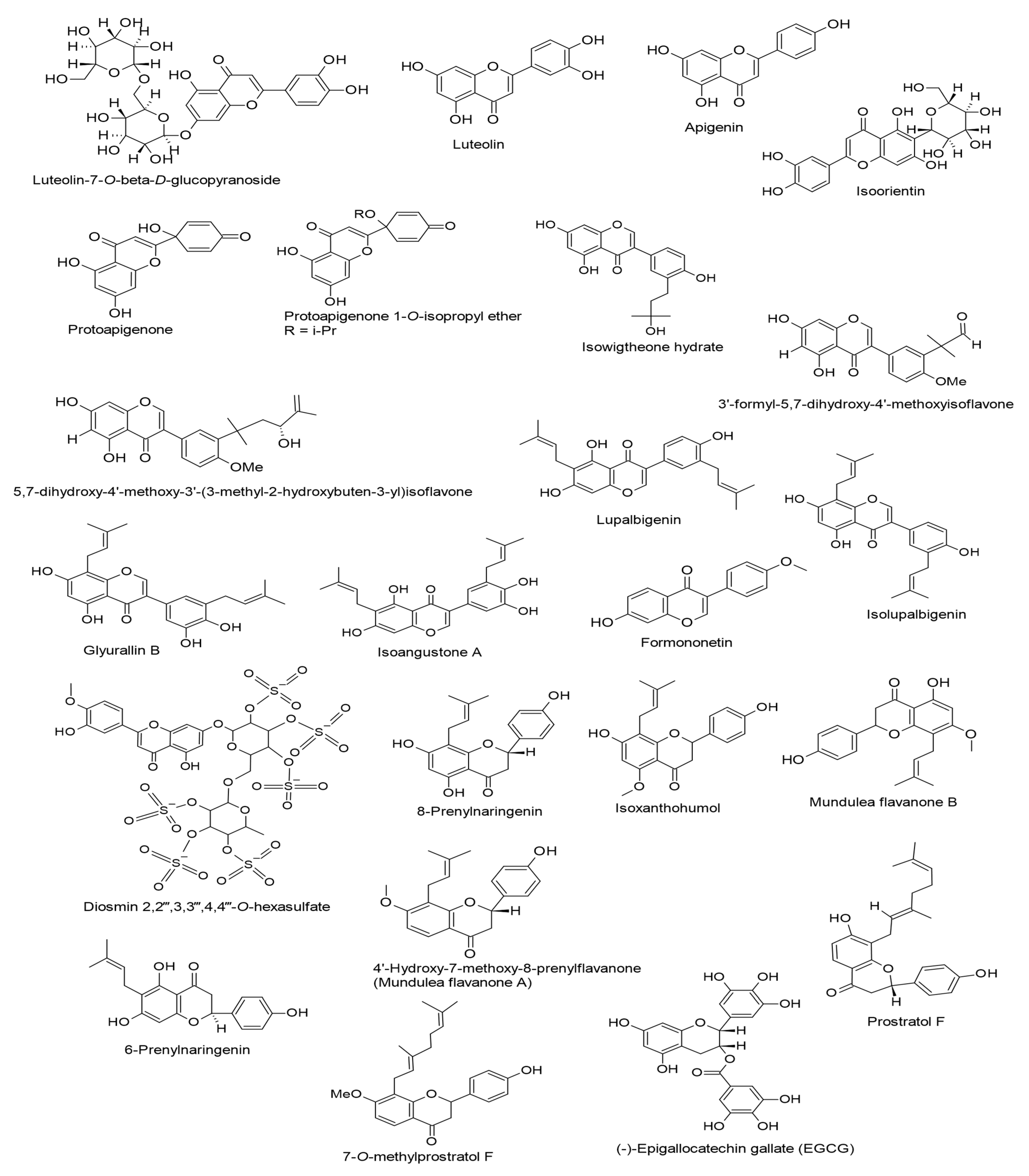
| Luteolin-7-O-β-D-glucopyranoside. (20 µg/mL). Lindernia Crustacea. | Flavones | Lytic replication. Rta expression. | [55] |
| Luteolin. (10, 20, and 50 µM). Various medicinal herbs, fruits, and vegetables. | Flavones | Lytic replication. Rta and Zta expressions. Sp1-luc activity. | [56] |
| Apigenin. (50 µM). Distributed in various fruits and vegetables. | Flavones | Lytic replication. Virion production. Rta and Zta expressions. | [57] |
| Isoorientin (An IC50 value of 393 mol ratio/32 pmol TPA). Passiflora edulis. | Flavones | Lytic cycle. EBV-EA. | [58] |
| Protoapigenone. (0.31 mM). Thelypteris torresiana. | Protoflavones | Lytic cycle. Rta, Zta, EA-D, and VCA expressions. | [59] |
| Protoapigenone. (IC50 = 0.127 µM and 0.50 µM). Thelypteris torresiana | Protoflavones | Lytic replication. Rta expression. | [60] |
| Protoapigenone 1′-O-isopropyl ether. (IC50 = 0.467 µM and 0.25 µM). Synthatically derived from apigenin. | Protoflavones | Lytic replication. Rta expression. | [60] |
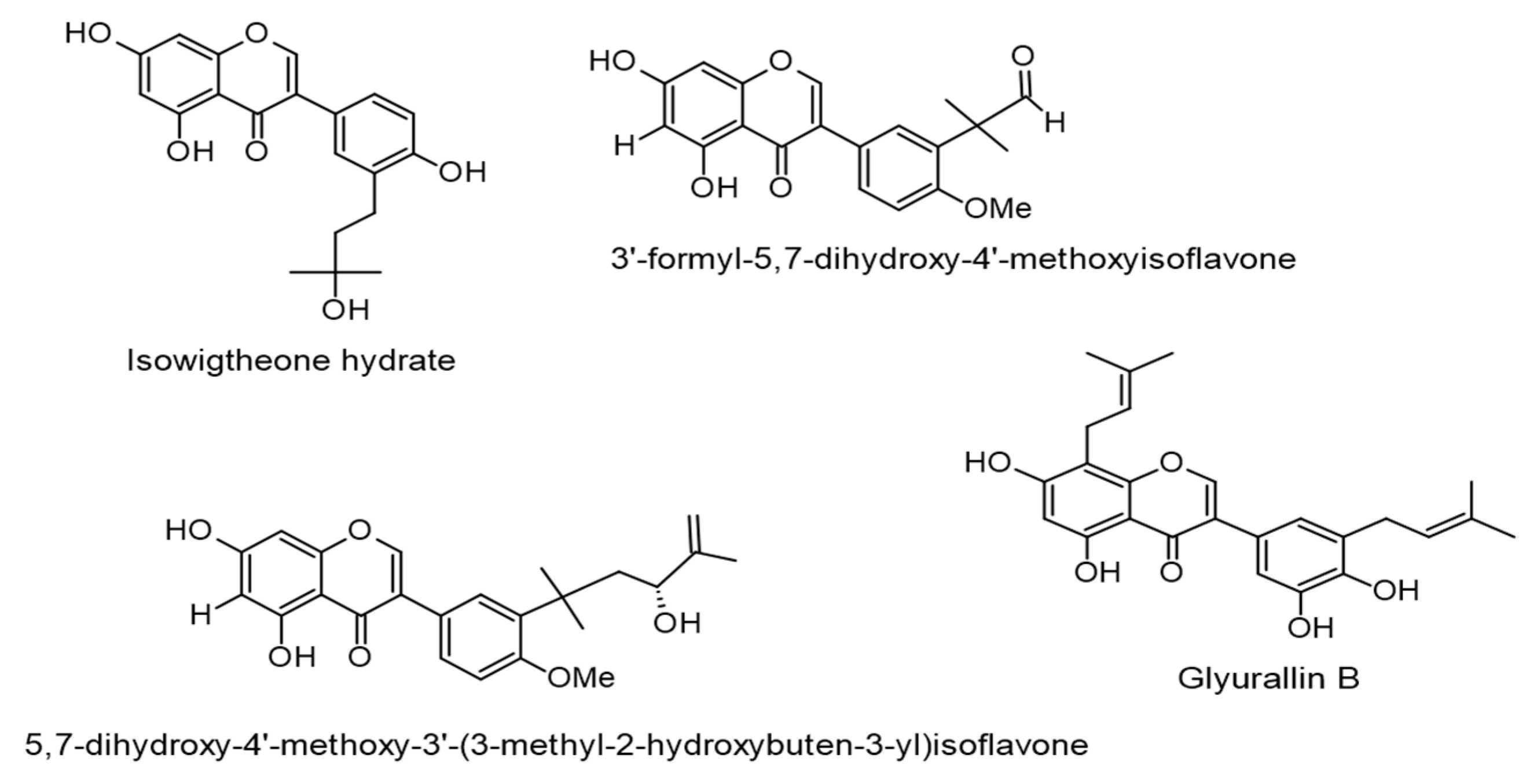
| Isowigtheone hydrate, 3′-formyl-5,7-dihydroxy-4′-methoxyisoflavone, and 5,7-dihydroxy-4′-methoxy-3′-(3-methyl-2-hydroxybuten-3-yl)isoflavone. (IC50 values of 271, 358, and 285 mol ratio/32 pmol TPA, respectively). Ficus hispida L.f. | Isoflavones | Lytic cycle. EBV-EA. | [61] |
| Lupalbigenin, isolupalbigenin, glyurallin B, and isoangustone A. (IC50 values of 290, 285, 278, and 282 mol ratio/32 pmol TPA, respectively). Derris Scandens (Roxb.) Benth. | Isoflavones | Lytic cycle. EBV-EA. | [62] |
| Formononetin. Binding affinity (−6.6 kcal/mol). PubChem CID: 5280378. | Isoflavones | EBV life cycle (entry). EBV gH (in silico). | [63] |
| Diosmin 2″,2‴,3″,3‴,4″,4‴-O-hexasulfate. (20 µM). Binding affinity with Zta (−8.7 kcal/mol). Synthesized. | Flavones (Sulfated) | Lytic replication. EBV DNA load. LMP1. Zta (in silico). | [64] |
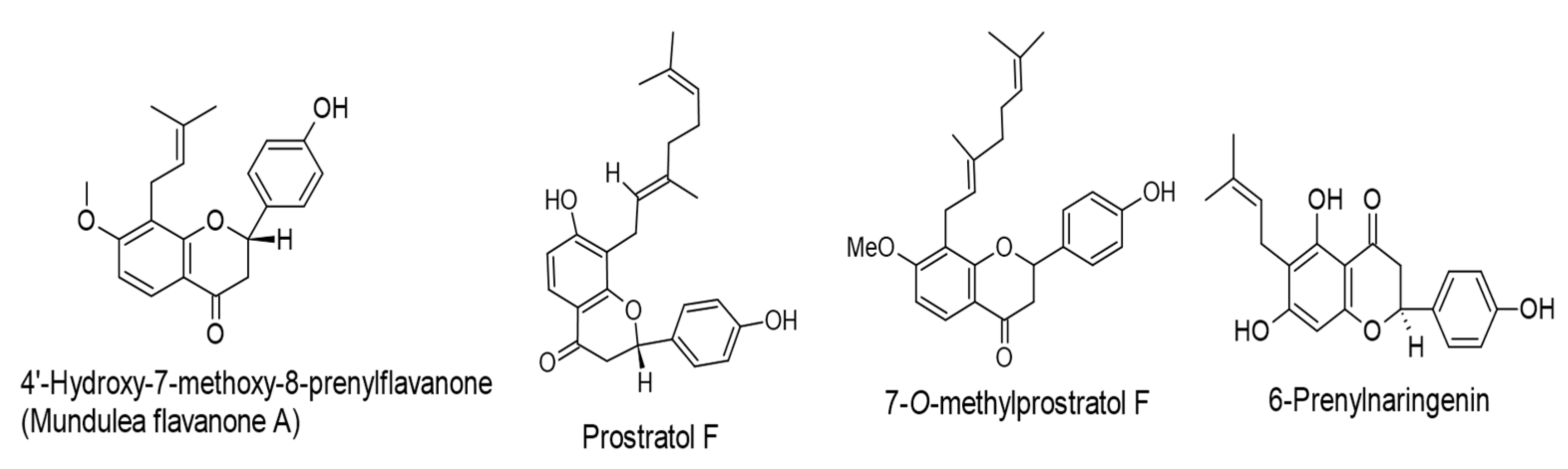
| 8-Prenylnaringenin, isoxanthohumol, mundulea flavanone B, and 6-Prenylnaringenin. (IC50 values of 263, 293, 281, and 263 mol ratio/32 pmol TPA, respectively). Humulus lupulus L. | Flavanones | Lytic cycle. EBV-EA. | [65] |
| 4′-Hydroxy-7-methoxy-8-prenylflavanone (Mundulea flavanone A), prostratol F, and 7-O-methylprostratol F. (IC50 values of 230, 270, and 348 mol ratio/32 pmol TPA, respectively). Biotransformed products of prenylated chalcones by Aspergillus saitoi. | Flavanones | Lytic cycle. EBV-EA. | [66] |
| Lytic replication. MEK/ERK1/2. PI3K/AKT. | [67] | ||
| Lytic replication. LMP1. | [68] | ||
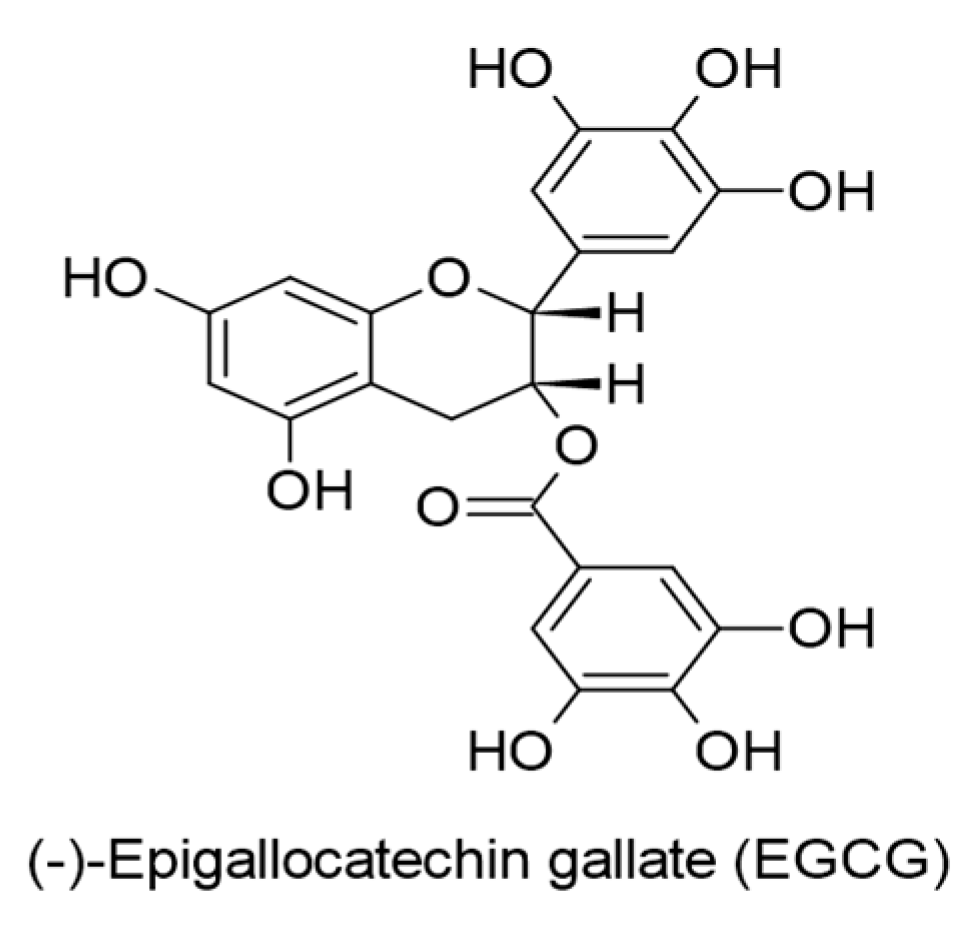
| (-)-Epigallocatechin gallate (EGCG). (0.5-50 µM). Binding affinity with EBNA1 (−6.8 kcal/mol). Binding affinity with gH (−7.8 kcal/mol). Camellia sinensis. | Flavanols (Catechins) | Lytic replication. LMP1. MAPKs/wt-p53. JNKs/c-Jun. | [69] |
| Lytic replication. EBNA1. oriP-DNA. | [70] | ||
| Lytic replication. EBNA1 (in silico). | [71] | ||
| EBV life cycle (entry). EBV gH (in silico). | [63] | ||
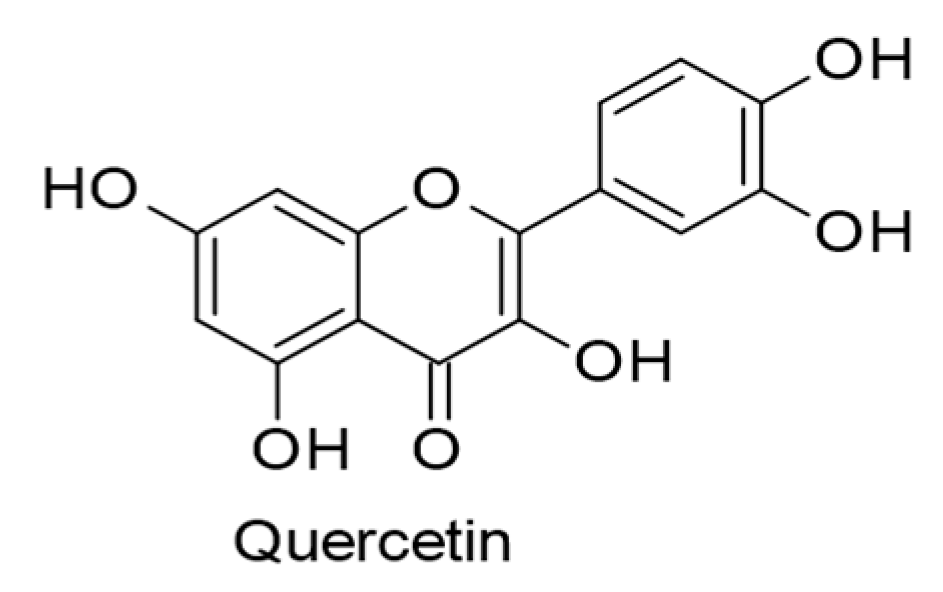
| Quercetin. (62 µM). Glycyrrhiza uralensis. | Flavonols | EBV life cycle (entry and latency). EBNA1. | [72] |
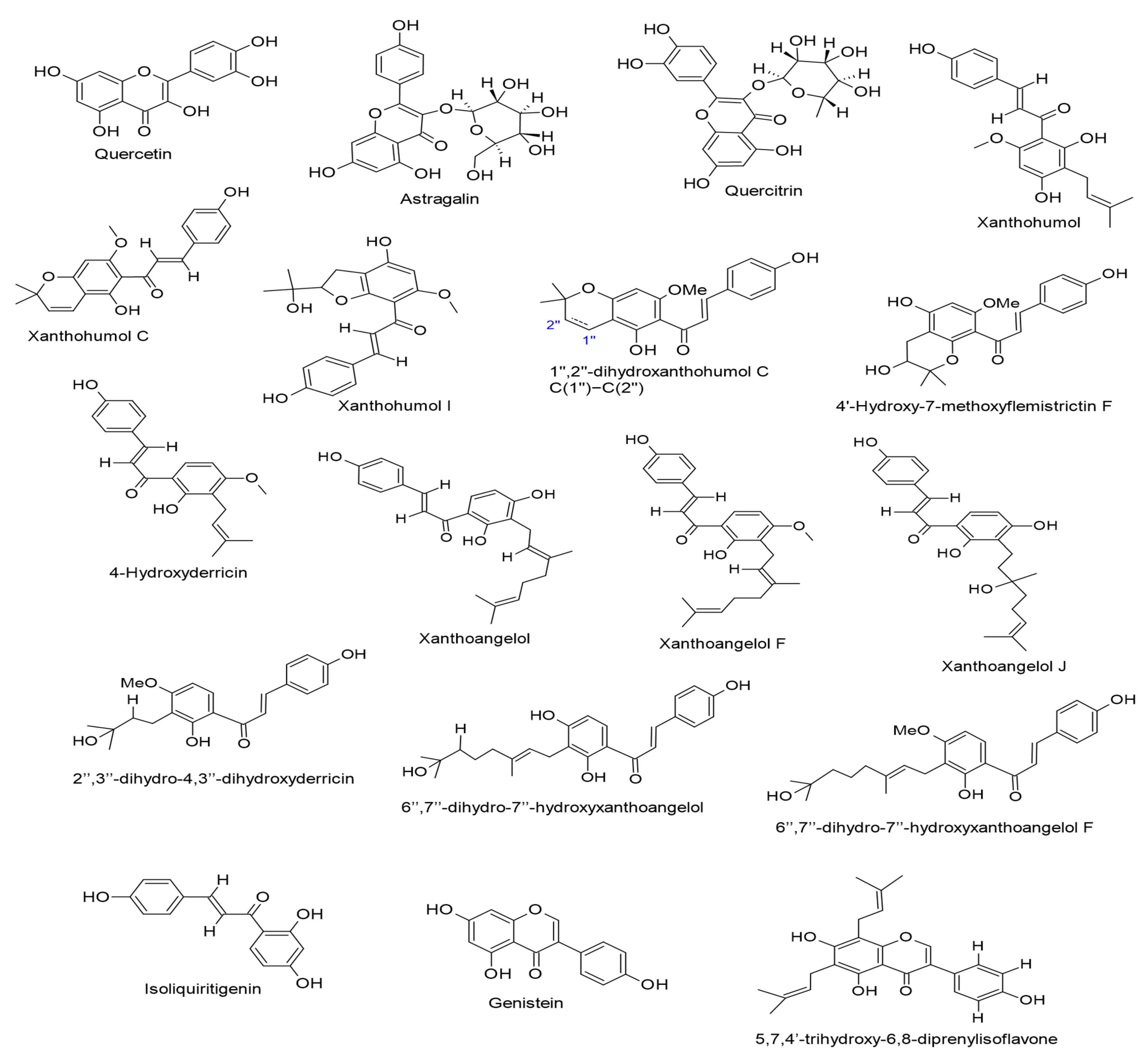
| Astragalin and quercitrin. (IC50 values of 543 and 532 mol ratio/32 pmol TPA, respectively). Humulus lupulus L. | Flavonols (Flavonol glycosides) | Lytic cycle. EBV-EA. | [65] |
| Xanthohumol, Xanthohumol C, 1″,2″-dihydroxanthohumol C, Xanthohumol I, and 4′-Hydroxy-7-methoxyflemistrictin F. (IC50 values of 485, 520, 526, 470, and 501 mol ratio/32 pmol TPA, respectively). Humulus lupulus L. | Chalcones | Lytic cycle. EBV-EA. | [65] |
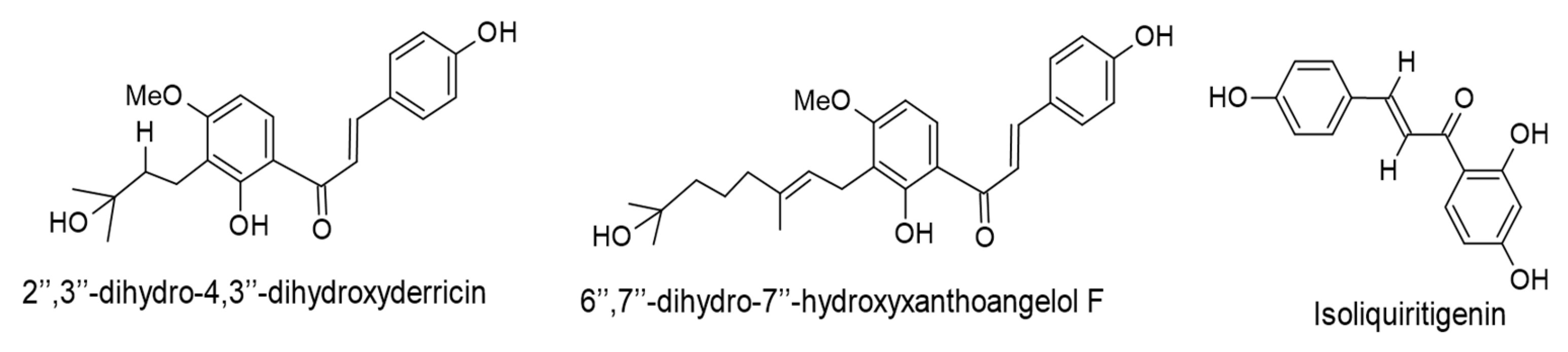
| 4-Hydroxyderricin, xanthoangelol, xanthoangelol F, 2″,3″-dihydro-4,3″-dihydroxyderricin, 6″,7″-dihydro-7″-hydroxyxanthoangelol, xanthoangelol J, and 6″,7″-dihydro-7″-hydroxyxanthoangelol F. (IC50 values of 213, 269, 262, 210, 211, 219, and 215 mol ratio/32 pmol TPA, respectively). The first three compounds were isolated from Angelica keiskei, while the rest are biotransformed products of prenylated chalcones by Aspergillus saitoi. | Chalcones | Lytic cycle. EBV-EA. | [66] |
| Isoliquiritigenin. (45 µM). Glycyrrhiza uralensis. | Chalcones | EBV entry. | [72] |
| Compound, Concentration, and Source | Chemical Class | Molecular Mechanisms (Inhibition) | Reference |
|---|---|---|---|
| Hesperetin. (0.5–50 µM). Distributed in the genus Citrus. | Flavanones | Lytic DNA replication. Virus progeny production. HIF1α expression. | [112] |
| Hesperidin. (EC50 = 0.2399 µM and 18 µM). Thymus capitatus. | Flavanones | Lytic DNA replication. ORF45 and K8.1 expressions. | [113] |
| Eupafolin. (EC50 = 1.396 µM and 105 µM). Thymus capitatus. | Flavones | Lytic DNA replication. ORF45 and K8.1 expressions. | [113] |
| (-)-Epigallocatechin gallate (EGCG). (50 µg/mL). Camellia sinensis. | Flavanols (Catechins) | Lytic DNA replication. Virus progeny production. Rta expression. | [114] |
| Quercetin. (15 µM). Fruits and vegetables. | Flavonols | Lytic DNA replication. ROS. | [115] |
| Delphinidin-3-glucoside chloride (myrtillin), cyanidin-3-glucoside, delphinidin-3-O-rutinoside chloride, and petunidin-3-galactoside. (75 and 150 µg/mL). Ribes nigrum L. Vaccinium myrtillus L. | Anthocyanins | Lytic DNA replication. | [116] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, S.T.S.; Šudomová, M. Molecular Mechanisms of Flavonoids against Tumor Gamma-Herpesviruses and Their Correlated Cancers—A Focus on EBV and KSHV Life Cycles and Carcinogenesis. Int. J. Mol. Sci. 2023, 24, 247. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010247
Hassan STS, Šudomová M. Molecular Mechanisms of Flavonoids against Tumor Gamma-Herpesviruses and Their Correlated Cancers—A Focus on EBV and KSHV Life Cycles and Carcinogenesis. International Journal of Molecular Sciences. 2023; 24(1):247. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010247
Chicago/Turabian StyleHassan, Sherif T. S., and Miroslava Šudomová. 2023. "Molecular Mechanisms of Flavonoids against Tumor Gamma-Herpesviruses and Their Correlated Cancers—A Focus on EBV and KSHV Life Cycles and Carcinogenesis" International Journal of Molecular Sciences 24, no. 1: 247. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010247