
Astrocytic GPCR-Induced Ca2+ Signaling Is Not Causally Related to Local Cerebral Blood Flow Changes
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
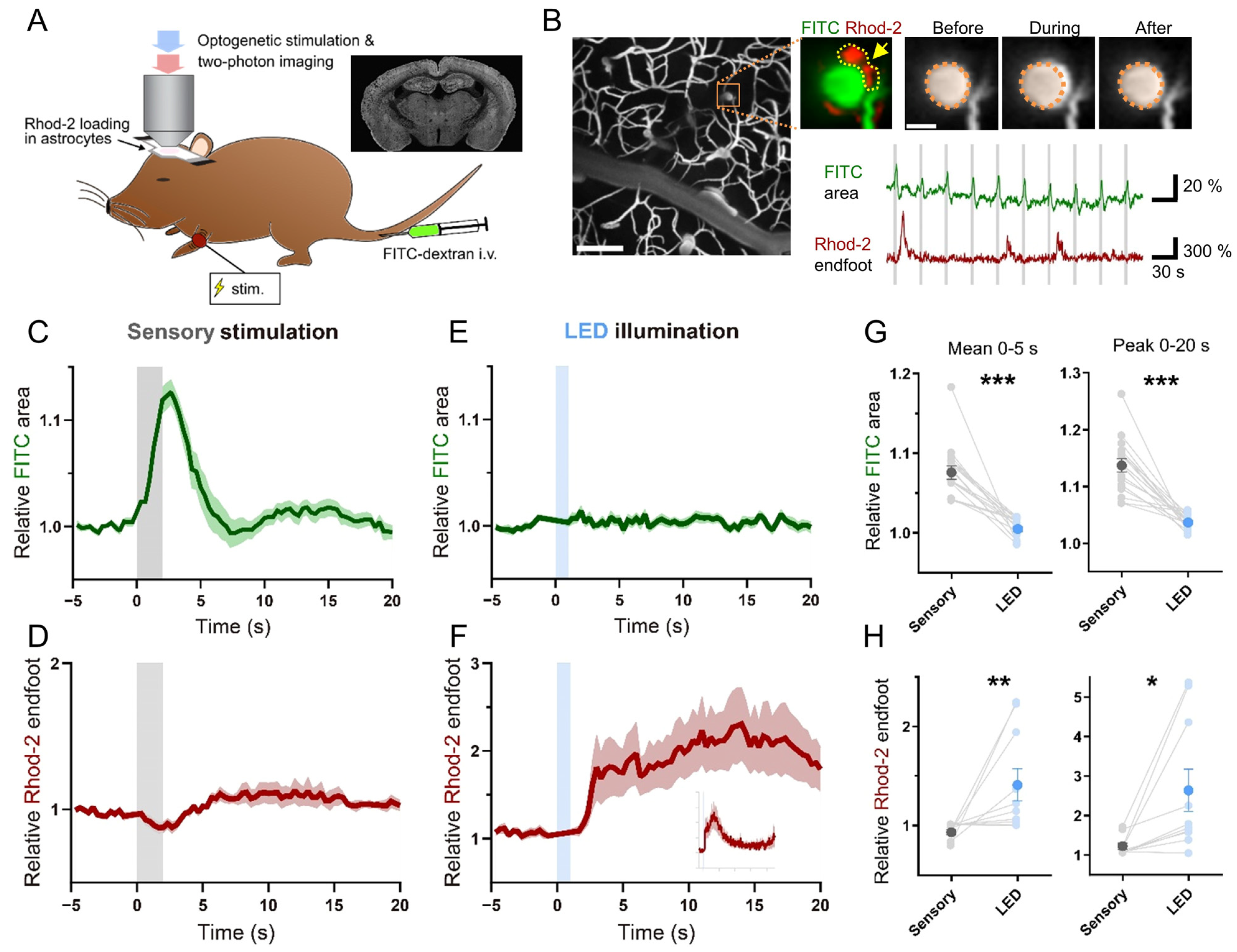
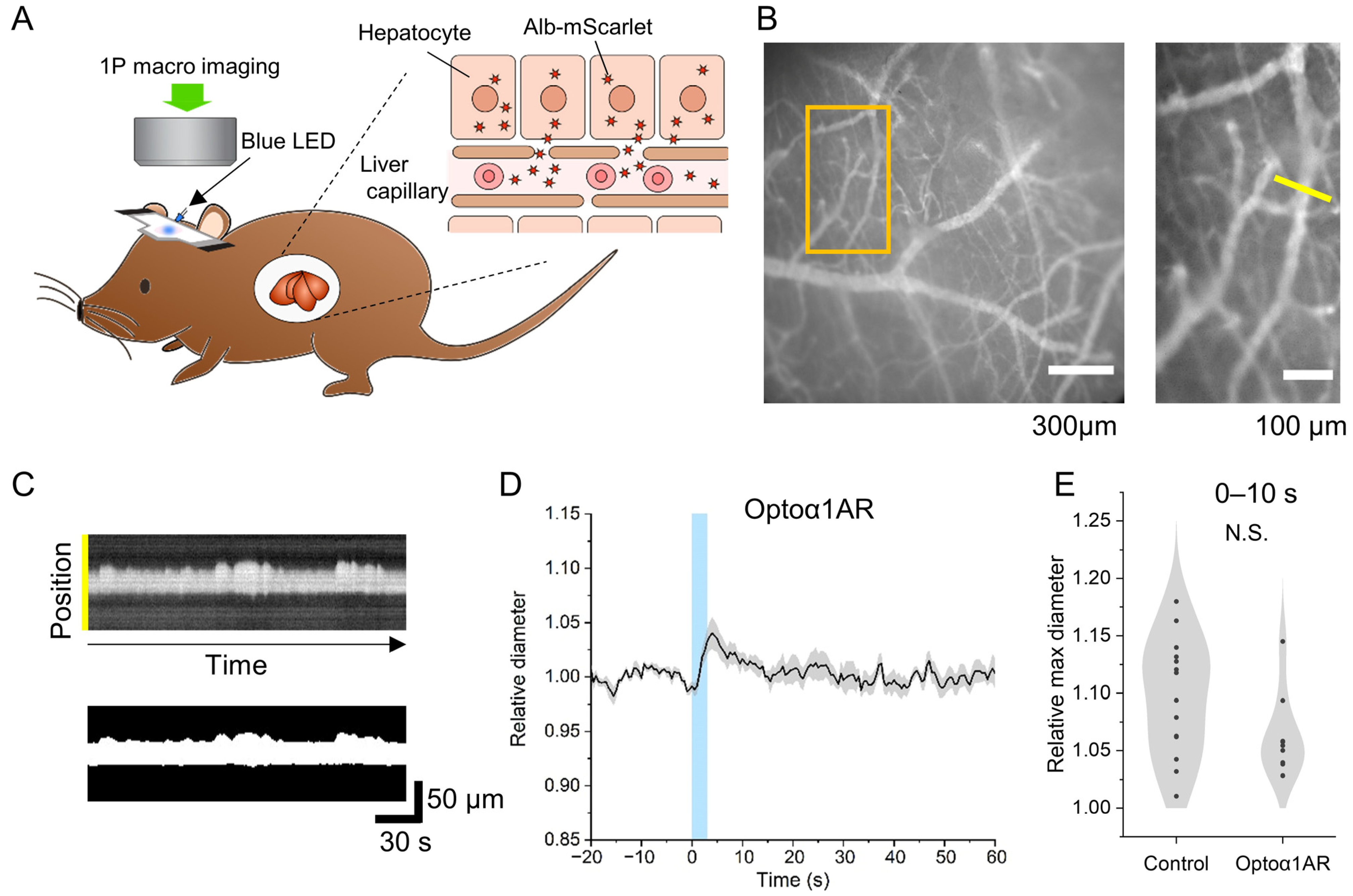
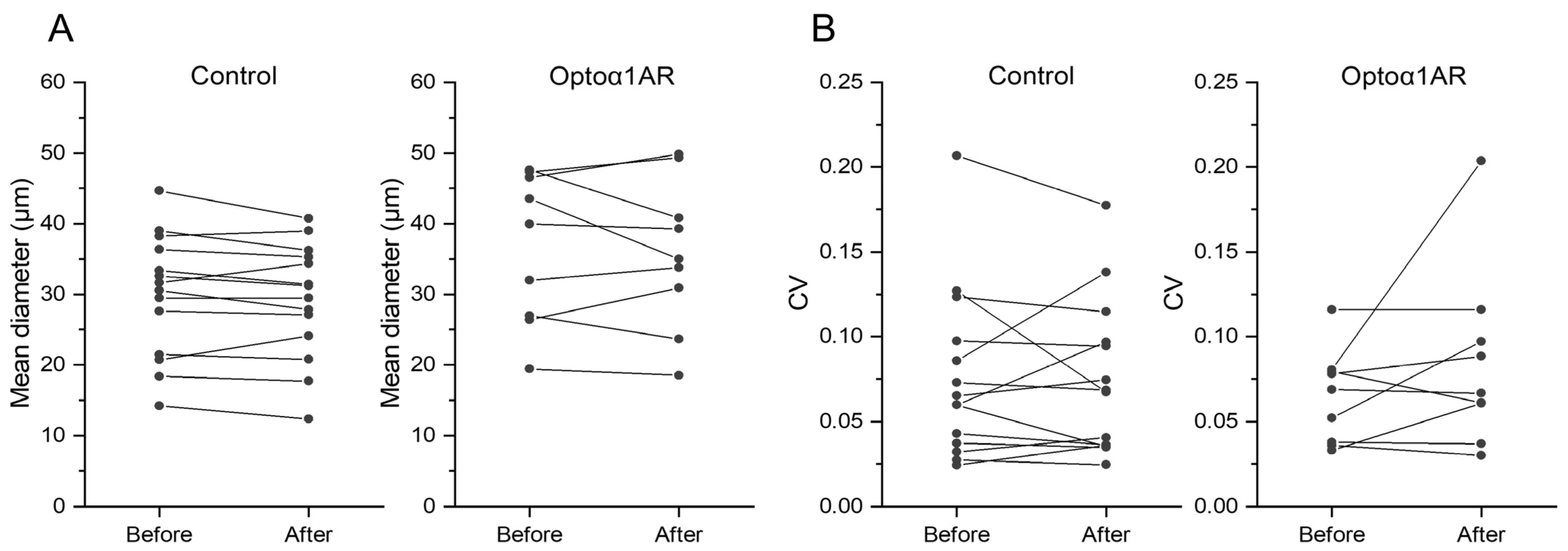
2. Results
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Surgery for Acute In Vivo Two-Photon Imaging
4.3. In Vivo Two-Photon Imaging
4.4. Imaging of Awake Mice
4.5. Data Analysis and Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roy, C.; Sherrington, C. On the regulation of the blood supply of the brain. J. Physiol. 1890, 11, 85–108. [Google Scholar] [CrossRef] [PubMed]
- Attwell, D.; Buchan, A.M.; Charpak, S.; Lauritzen, M.; MacVicar, B.A.; Newman, E.A. Glial and Neuronal Control of Brain Blood Flow. Nature 2010, 468, 232–243. Available online: https://pubmed.ncbi.nlm.nih.gov/21068832/ (accessed on 31 January 2023). [CrossRef] [PubMed]
- Attwell, D.; Iadecola, C. The Neural Basis of Functional Brain Imaging Signals. Trends Neurosci. 2002, 25, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Drew, P.J. Vascular and Neural Basis of the BOLD Signal. Curr. Opin. Neurobiol. 2019, 58, 61–69. Available online: https://pubmed.ncbi.nlm.nih.gov/31336326/ (accessed on 31 January 2023). [CrossRef]
- Hillman, E.M.C. Coupling Mechanism and Significance of the BOLD Signal: A Status Report. Annu. Rev. Neurosci. 2014, 37, 161–181. Available online: https://pubmed.ncbi.nlm.nih.gov/25032494/ (accessed on 31 January 2023). [CrossRef]
- Herculano-Houzel, S. The Glia/Neuron Ratio: How It Varies Uniformly across Brain Structures and Species and What That Means for Brain Physi-Ology and Evolution. Glia 2014, 62, 1377–1391. Available online: https://pubmed.ncbi.nlm.nih.gov/24807023/ (accessed on 28 February 2023). [CrossRef]
- Keller, D.; Erö, C.; Markram, H. Cell Densities in the Mouse Brain: A Systematic Review. Front. Neuroanat. 2018, 12, 83. Available online: https://www.frontiersin.org/articles/10.3389/fnana.2018.00083/full (accessed on 28 February 2023). [CrossRef]
- Mishima, T.; Sakatani, S.; Hirase, H. Intracellular Labeling of Single Cortical Astrocytes In Vivo. J. Neurosci. Methods 2007, 166, 32–40. Available online: https://pubmed.ncbi.nlm.nih.gov/17686526/ (accessed on 31 January 2023). [CrossRef]
- von Bartheld, C.S.; Bahney, J.; Herculano-Houzel, S. The Search for True Numbers of Neurons and Glial Cells in the Human Brain: A Review of 150 Years of Cell Counting. J. Comp. Neurol. 2016, 524, 3865–3895. Available online: https://pubmed.ncbi.nlm.nih.gov/27187682/ (accessed on 28 February 2023). [CrossRef]
- Kofuji, P.; Araque, A. G-Protein-Coupled Receptors in Astrocyte–Neuron Communication. Neuroscience 2021, 456, 71–84. Available online: https://pubmed.ncbi.nlm.nih.gov/32224231/ (accessed on 4 March 2021). [CrossRef]
- Mulligan, S.J.; MacVicar, B.A. Calcium Transients in Astrocyte Endfeet Cause Cerebrovascular Constrictions. Nature 2004, 431, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Petzold, G.C.; Albeanu, D.F.; Sato, T.F.; Murthy, V.N. Coupling of Neural Activity to Blood Flow in Olfactory Glomeruli Is Mediated by Astrocytic Pathways. Neuron 2008, 58, 897–910. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Tian, G.-F.F.; Peng, W.; Lou, N.; Libionka, W.; Han, X.; Nedergaard, M. Astrocyte-Mediated Control of Cerebral Blood Flow. Nat. Neurosci. 2006, 9, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Zonta, M.; Sebelin, A.; Gobbo, S.; Fellin, T.; Pozzan, T.; Carmignoto, G. Glutamate-Mediated Cytosolic Calcium Oscillations Regulate a Pulsatile Prostaglandin Release from Cultured Rat Astrocytes. J. Physiol. 2003, 553, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Bonder, D.E.; McCarthy, K.D. Astrocytic Gq-GPCR-Linked IP3R-Dependent Ca2+ Signaling Does Not Mediate Neurovascular Coupling in Mouse Visual Cortex In Vivo. J. Neurosci. 2014, 34, 13139–13150. [Google Scholar] [CrossRef] [PubMed]
- Nizar, K.; Uhlirova, H.; Tian, P.; Saisan, P.A.; Cheng, Q.; Reznichenko, L.; Weldy, K.L.; Steed, T.C.; Sridhar, V.B.; MacDonald, C.L.; et al. In Vivo Stimulus-Induced Vasodilation Occurs without IP3 Receptor Activation and May Precede Astrocytic Calcium Increase. J. Neurosci. 2013, 33, 8411–8422. [Google Scholar] [CrossRef]
- Takata, N.; Nagai, T.; Ozawa, K.; Oe, Y.; Mikoshiba, K.; Hirase, H. Cerebral Blood Flow Modulation by Basal Forebrain or Whisker Stimulation Can Occur Independently of Large Cytosolic Ca2+ Signaling in Astrocytes. PLoS ONE 2013, 8, e66525. Available online: https://pubmed.ncbi.nlm.nih.gov/23785506/ (accessed on 9 August 2021). [CrossRef]
- Srinivasan, R.; Huang, B.S.; Venugopal, S.; Johnston, A.D.; Chai, H.; Zeng, H.; Golshani, P.; Khakh, B.S. Ca2+ signaling in astrocytes from Ip3r2-/- mice in brain slices and during startle responses in vivo. Nat. Neurosci. 2015, 18, 708–717. [Google Scholar] [CrossRef]
- Stobart, J.L.; Ferrari, K.D.; Barrett, M.J.P.; Glück, C.; Stobart, M.J.; Zuend, M.; Weber, B. Cortical Circuit Activity Evokes Rapid Astrocyte Calcium Signals on a Similar Timescale to Neurons. Neuron 2018, 98, 726–735.e4. [Google Scholar] [CrossRef]
- Del Franco, A.P.; Chiang, P.P.; Newman, E.A. Dilation of Cortical Capillaries Is Not Related to Astrocyte Calcium Signaling. Glia 2022, 70, 508–521. Available online: https://pubmed.ncbi.nlm.nih.gov/34767261/ (accessed on 10 March 2023). [CrossRef]
- Lind, B.L.; Brazhe, A.R.; Jessen, S.B.; Tan, F.C.C.; Lauritzen, M.J. Rapid stimulus-evoked astrocyte Ca2+ elevations and hemodynamic responses in mouse somatosensory cortex in vivo. Proc. Natl. Acad. Sci. USA 2013, 110, E4678–E4687. [Google Scholar] [CrossRef]
- Otsu, Y.; Couchman, K.; Lyons, D.G.; Collot, M.; Agarwal, A.; Mallet, J.-M.; Pfrieger, F.W.; Bergles, D.E.; Charpak, S. Calcium Dynamics in Astrocyte Processes during Neurovascular Coupling. Nat. Neurosci. 2015, 18, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Reynolds, J.P.; Chen, Y.; Gourine, A.V.; Rusakov, D.A.; Attwell, D. Astrocytes mediate neurovascular signaling to capillary pericytes but not to arterioles. Nat. Neurosci. 2016, 19, 1619–1627. [Google Scholar] [CrossRef] [PubMed]
- Institoris, Á.; Rosenegger, D.G.; Gordon, G.R. Arteriole Dilation to Synaptic Activation That Is Sub-Threshold to Astrocyte Endfoot Ca2+ Transients. J. Cereb. Blood Flow Metab. 2015, 35, 1411–1415. Available online: http://0-journals-sagepub-com.brum.beds.ac.uk/doi/10.1038/jcbfm.2015.141 (accessed on 4 August 2019). [CrossRef]
- Tran, C.H.T.; Peringod, G.; Gordon, G.R. Astrocytes Integrate Behavioral State and Vascular Signals during Functional Hyperemia. Neuron 2018, 100, 1133–1148.e3. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/30482689 (accessed on 4 August 2019). [CrossRef]
- Lind, B.L.; Jessen, S.B.; Lønstrup, M.; Joséphine, C.; Bonvento, G.; Lauritzen, M. Fast Ca2+ Responses in Astrocyte End-Feet and Neurovascular Coupling in Mice. Glia 2018, 66, 348–358. Available online: https://pubmed.ncbi.nlm.nih.gov/29058353/ (accessed on 10 March 2023). [CrossRef]
- Vaidyanathan, T.V.; Collard, M.; Yokoyama, S.; Reitman, M.E.; Poskanzer, K.E. Cortical Astrocytes Independently Regulate Sleep Depth and Duration via Separate GPCR Pathways. eLife 2021, 10, e63329. [Google Scholar] [CrossRef]
- Iwai, Y.; Ozawa, K.; Yahagi, K.; Mishima, T.; Akther, S.; Vo, C.T.; Lee, A.B.; Tanaka, M.; Itohara, S.; Hirase, H. Transient Astrocytic Gq Signaling Underlies Remote Memory Enhancement. Front. Neural Circuits 2021, 15, 658343. Available online: https://pubmed.ncbi.nlm.nih.gov/33828463/ (accessed on 13 April 2021). [CrossRef]
- Iwai, Y.; Ozawa, K.; Yahagi, K.; Tanaka, M.; Itohara, S.; Hirase, H. Transient astrocytic Gq signaling underlies remote memory enhancement. bioRxiv 2019. bioRxiv:753095. [Google Scholar]
- Wang, X.; Delle, C.; Asiminas, A.; Akther, S.; Vittani, M.; Brøgger, P.; Kusk, P.; Vo, C.T.; Radovanovic, T.; Konno, A.; et al. Liver-secreted fluorescent blood plasma markers enable chronic imaging of the microcirculation. Cell Rep. Methods 2022, 2, 100302. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Hata, A.; Miyata, T.; Hirase, H. Programmable Wireless Light-Emitting Diode Stimulator for Chronic Stimulation of Optogenetic Molecules in Freely Moving Mice. Neurophotonics 2014, 1, 011002. Available online: https://pubmed.ncbi.nlm.nih.gov/26157963/ (accessed on 4 March 2021). [CrossRef]
- Cauli, B.; Hamel, E. Brain Perfusion and Astrocytes. Trends Neurosci. 2018, 41, 409–413. Available online: https://pubmed.ncbi.nlm.nih.gov/29933772/ (accessed on 13 March 2023). [CrossRef] [PubMed]
- Filosa, J.A.; Iddings, J.A. Astrocyte Regulation of Cerebral Vascular Tone. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H609. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/PMC3761330/ (accessed on 7 March 2023). [CrossRef]
- Howarth, C. The Contribution of Astrocytes to the Regulation of Cerebral Blood Flow. Front. Neurosci. 2014, 8, 103. Available online: https://pubmed.ncbi.nlm.nih.gov/24847203/ (accessed on 7 March 2023). [CrossRef] [PubMed]
- Lia, A.; Spiezio, A.; Di Speggiorin, M.; Zonta, M. Two Decades of Astrocytes in Neurovascular Coupling. Front. Netw. Physiol. 2023, 3, 1162757. Available online: https://www.frontiersin.org/articles/10.3389/fnetp.2023.1162757/full (accessed on 4 May 2023). [CrossRef]
- Macvicar, B.A.; Newman, E.A. Astrocyte Regulation of Blood Flow in the Brain. Cold Spring Harb. Perspect. Biol. 2015, 7, a020388. Available online: https://cshperspectives.cshlp.org/content/7/5/a020388.short (accessed on 7 March 2023). [CrossRef]
- Mishra, A. Binaural Blood Flow Control by Astrocytes: Listening to Synapses and the Vasculature. J. Physiol. 2017, 595, 1885–1902. Available online: https://pubmed.ncbi.nlm.nih.gov/27619153/ (accessed on 7 March 2023). [CrossRef]
- Nuriya, M.; Hirase, H. Involvement of Astrocytes in Neurovascular Communication. Prog. Brain Res. 2016, 225, 41–62. Available online: https://pubmed.ncbi.nlm.nih.gov/27130410/ (accessed on 7 March 2023).
- Jego, P.; Pacheco-Torres, J.; Araque, A.; Canals, S. Functional MRI in Mice Lacking IP3-Dependent Calcium Signaling in Astrocytes. J. Cereb. Blood Flow Metab. 2014, 34, 1599–1603. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Wu, P.-H.; Hughes, E.G.; Fukaya, M.; Tischfield, M.A.; Langseth, A.J.; Wirtz, D.; Bergles, D.E. Transient Opening of the Mitochondrial Permeability Transition Pore Induces Microdomain Calcium Transients in Astrocyte Processes. Neuron 2017, 93, 587–605.e7. Available online: http://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/S0896627316310078 (accessed on 26 August 2023). [CrossRef] [PubMed]
- Stobart, J.L.; Ferrari, K.D.; Barrett, M.J.P.; Stobart, M.J.; Looser, Z.J.; Saab, A.S.; Weber, B. Long-Term In Vivo Calcium Imaging of Astrocytes Reveals Distinct Cellular Compartment Responses to Sensory Stimulation. Cereb. Cortex 2018, 28, 184–198. Available online: https://pubmed.ncbi.nlm.nih.gov/28968832/ (accessed on 9 August 2021). [CrossRef]
- Masamoto, K.; Unekawa, M.; Watanabe, T.; Toriumi, H.; Takuwa, H.; Kawaguchi, H.; Kanno, I.; Matsui, K.; Tanaka, K.F.; Tomita, Y.; et al. Unveiling Astrocytic Control of Cerebral Blood Flow with Optogenetics. Sci. Rep. 2015, 5, 11455. Available online: https://pubmed.ncbi.nlm.nih.gov/26076820/ (accessed on 10 March 2023). [CrossRef]
- Takata, N.; Sugiura, Y.; Yoshida, K.; Koizumi, M.; Hiroshi, N.; Honda, K.; Yano, R.; Komaki, Y.; Matsui, K.; Suematsu, M.; et al. Optogenetic Astrocyte Activation Evokes BOLD fMRI Response with Oxygen Consumption without Neuronal Activity Modulation. Glia 2018, 66, 2013–2023. Available online: https://pubmed.ncbi.nlm.nih.gov/29845643/ (accessed on 13 March 2023). [CrossRef] [PubMed]
- Octeau, J.C.; Gangwani, M.R.; Allam, S.L.; Tran, D.; Huang, S.; Hoang-Trong, T.M.; Golshani, P.; Rumbell, T.H.; Kozloski, J.R.; Khakh, B.S. Transient, Consequential Increases in Extracellular Potassium Ions Accompany Channelrhodopsin2 Excitation. Cell Rep. 2019, 27, 2249–2261.e7. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/31116972 (accessed on 15 August 2019). [CrossRef]
- Wang, F.; Smith, N.A.; Xu, Q.; Goldman, S.; Peng, W.; Huang, J.H.; Takano, T.; Nedergaard, M. Photolysis of caged Ca2+ but not receptor-mediated Ca2+ signaling triggers astrocytic glutamate release. J. Neurosci. 2013, 33, 17404–17412. [Google Scholar] [CrossRef] [PubMed]
- Rungta, R.L.; Bernier, L.P.; Dissing-Olesen, L.; Groten, C.J.; LeDue, J.M.; Ko, R.; Drissler, S.; MacVicar, B.A. Ca2+ Transients in Astrocyte Fine Processes Occur via Ca2+ Influx in the Adult Mouse Hippocampus. Glia 2016, 64, 2093–2103. Available online: https://pubmed.ncbi.nlm.nih.gov/27479868/ (accessed on 12 May 2023). [CrossRef]
- Gu, X.; Chen, W.; Volkow, N.D.; Koretsky, A.P.; Du, C.; Pan, Y. Synchronized Astrocytic Ca2+ Responses in Neurovascular Coupling during Somatosensory Stimulation and for the Resting State. Cell Rep. 2018, 23, 3878–3890. Available online: https://pubmed.ncbi.nlm.nih.gov/29949771/ (accessed on 10 March 2023). [CrossRef] [PubMed]
- Wang, X.; Lou, N.; Xu, Q.; Tian, G.F.; Peng, W.G.; Han, X.; Kang, J.; Takano, T.; Nedergaard, M. Astrocytic Ca2+ Signaling Evoked by Sensory Stimulation In Vivo. Nat. Neurosci. 2006, 9, 816–823. [Google Scholar] [CrossRef]
- Girouard, H.; Bonev, A.D.; Hannah, R.M.; Meredith, A.; Aldrich, R.W.; Nelson, M.T. Astrocytic Endfoot Ca2+ and BK Channels Determine both Arteriolar Dilation and Constriction. Proc. Natl. Acad. Sci. USA 2010, 107, 3811–3816. [Google Scholar] [CrossRef]
- Hill, R.A.; Tong, L.; Yuan, P.; Murikinati, S.; Gupta, S.; Grutzendler, J. Regional Blood Flow in the Normal and Ischemic Brain Is Controlled by Arteriolar Smooth Muscle Cell Contractility and Not by Capillary Pericytes. Neuron 2015, 87, 95–110. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/26119027 (accessed on 6 August 2019). [CrossRef]
- Hall, C.N.; Reynell, C.; Gesslein, B.; Hamilton, N.B.; Mishra, A.; Sutherland, B.A.; O’Farrell, F.M.; Buchan, A.M.; Lauritzen, M.; Attwell, D. Capillary pericytes regulate cerebral blood flow in health and disease. Nature 2014, 508, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Institoris, A.; Vandal, M.; Peringod, G.; Catalano, C.; Tran, C.H.; Yu, X.; Visser, F.; Breiteneder, C.; Molina, L.; Khakh, B.S.; et al. Astrocytes Amplify Neurovascular Coupling to Sustained Activation of Neocortex in Awake Mice. Nat. Commun. 2022, 13, 7872. Available online: https://pubmed.ncbi.nlm.nih.gov/36550102/ (accessed on 10 March 2023). [CrossRef] [PubMed]
- Rungta, R.L.; Osmanski, B.F.; Boido, D.; Tanter, M.; Charpak, S. Light Controls Cerebral Blood Flow in Naive Animals. Nat. Commun. 2017, 8, 14191. Available online: https://pubmed.ncbi.nlm.nih.gov/28139643/ (accessed on 15 May 2023). [CrossRef] [PubMed]
- Hirase, H.; Creso, J.; Buzsaki, G.; Buzsáki, G. Capillary Level Imaging of Local Cerebral Blood Flow in Bicuculline-Induced Epileptic Foci. Neuroscience 2004, 128, 209–216. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozawa, K.; Nagao, M.; Konno, A.; Iwai, Y.; Vittani, M.; Kusk, P.; Mishima, T.; Hirai, H.; Nedergaard, M.; Hirase, H. Astrocytic GPCR-Induced Ca2+ Signaling Is Not Causally Related to Local Cerebral Blood Flow Changes. Int. J. Mol. Sci. 2023, 24, 13590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241713590
Ozawa K, Nagao M, Konno A, Iwai Y, Vittani M, Kusk P, Mishima T, Hirai H, Nedergaard M, Hirase H. Astrocytic GPCR-Induced Ca2+ Signaling Is Not Causally Related to Local Cerebral Blood Flow Changes. International Journal of Molecular Sciences. 2023; 24(17):13590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241713590
Chicago/Turabian StyleOzawa, Katsuya, Masaki Nagao, Ayumu Konno, Youichi Iwai, Marta Vittani, Peter Kusk, Tsuneko Mishima, Hirokazu Hirai, Maiken Nedergaard, and Hajime Hirase. 2023. "Astrocytic GPCR-Induced Ca2+ Signaling Is Not Causally Related to Local Cerebral Blood Flow Changes" International Journal of Molecular Sciences 24, no. 17: 13590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241713590