
Relationship between Urinary Metabolomic Profiles and Depressive Episode in Antarctica
 , and
, and
Abstract
:1. Introduction
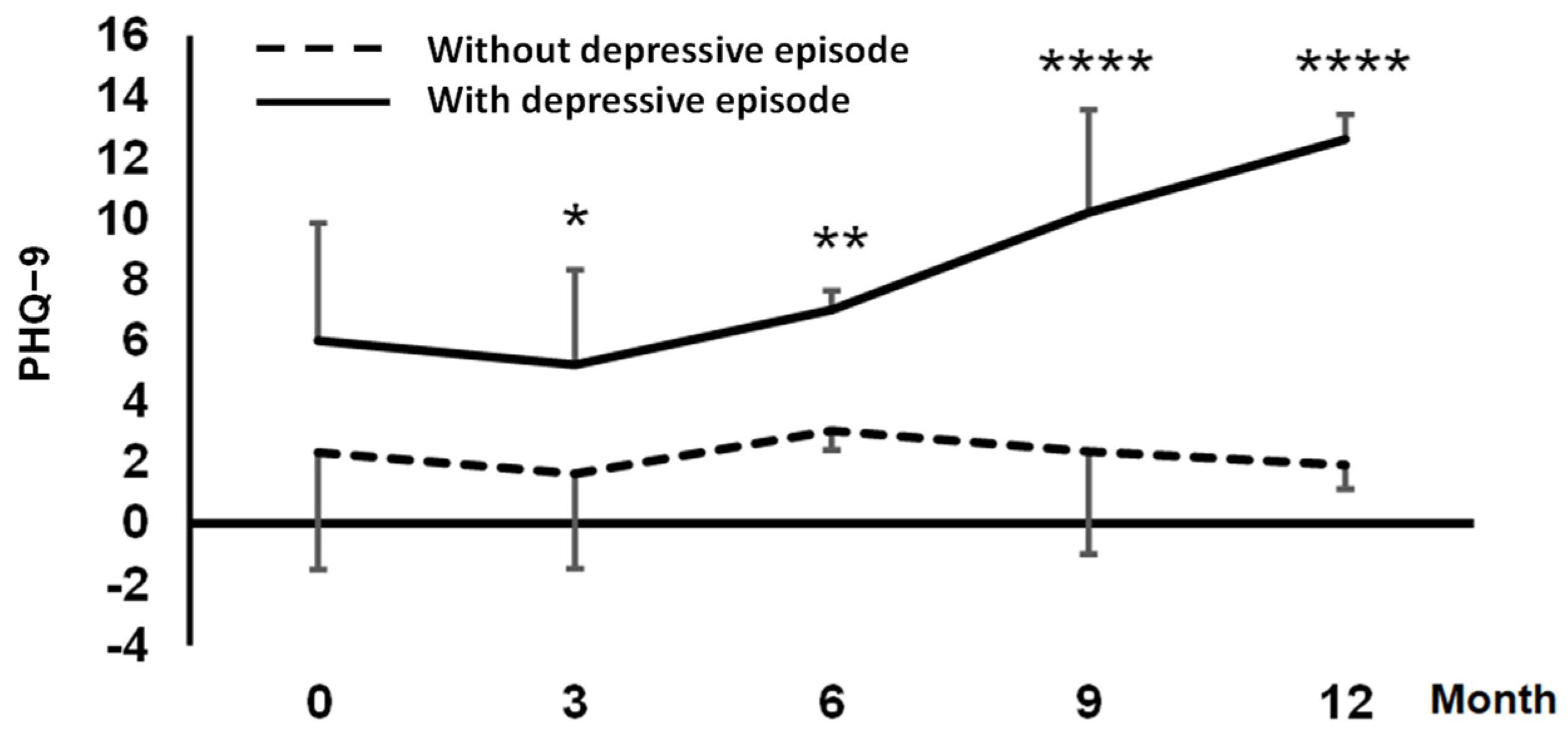
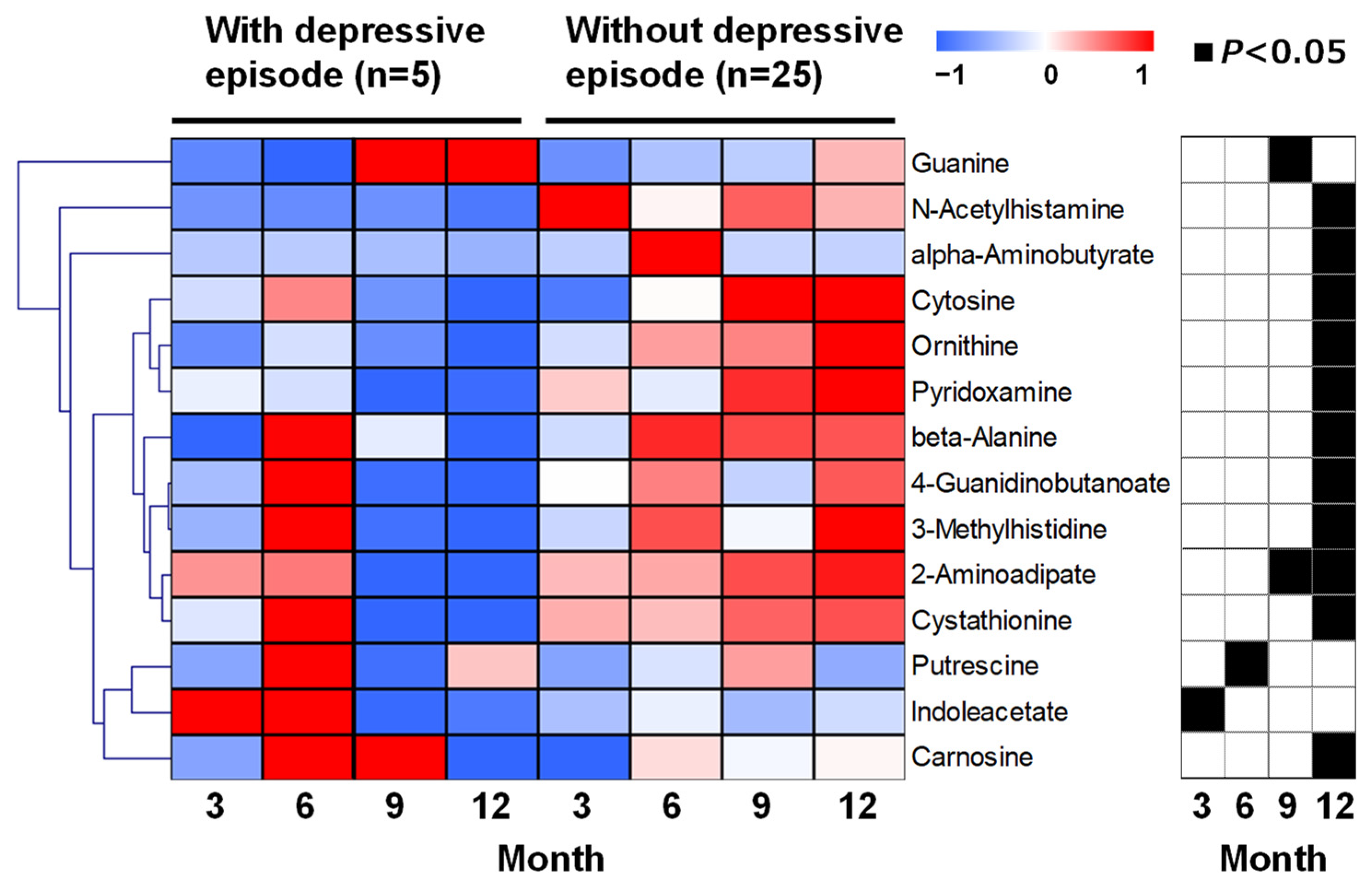
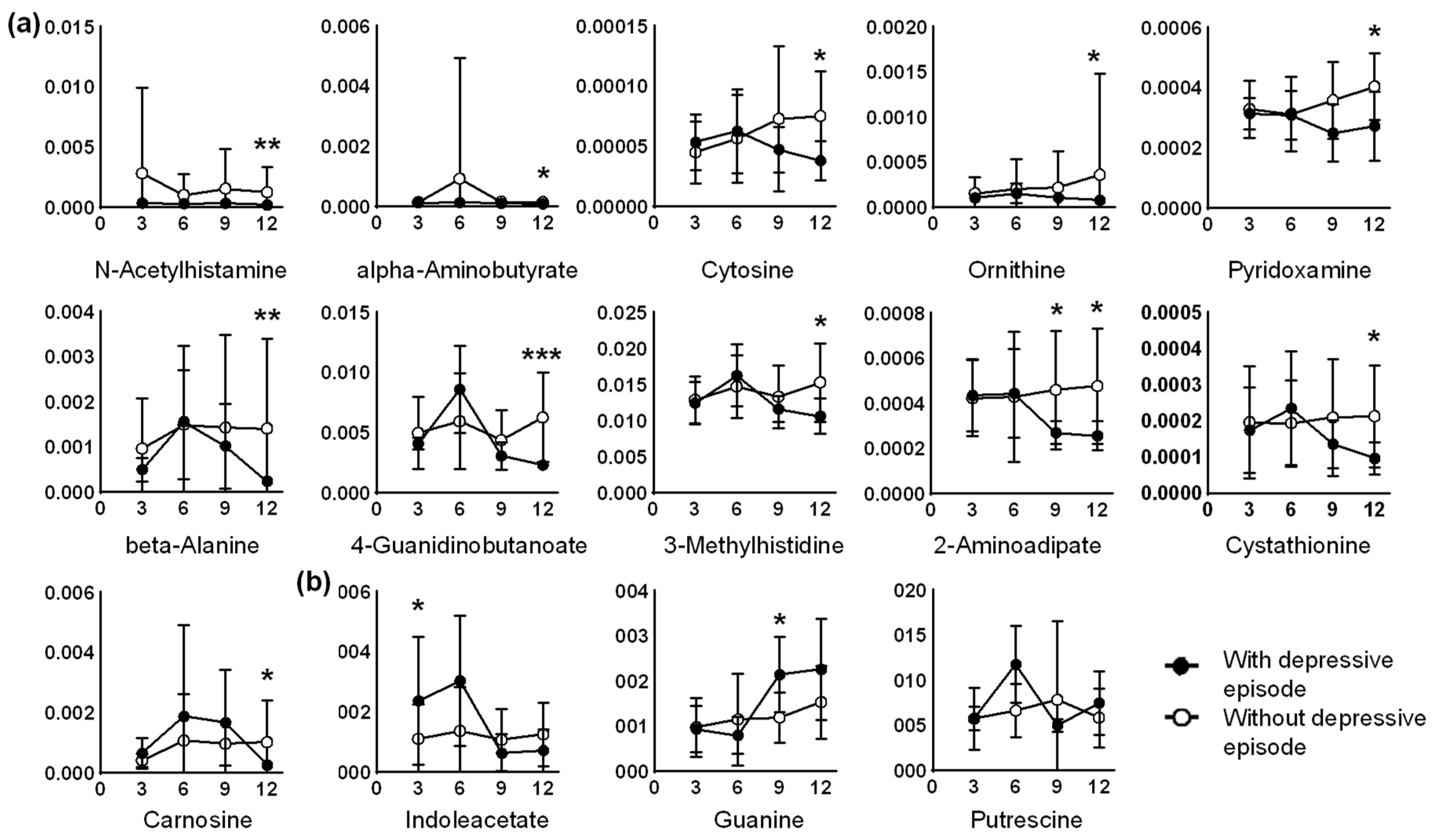
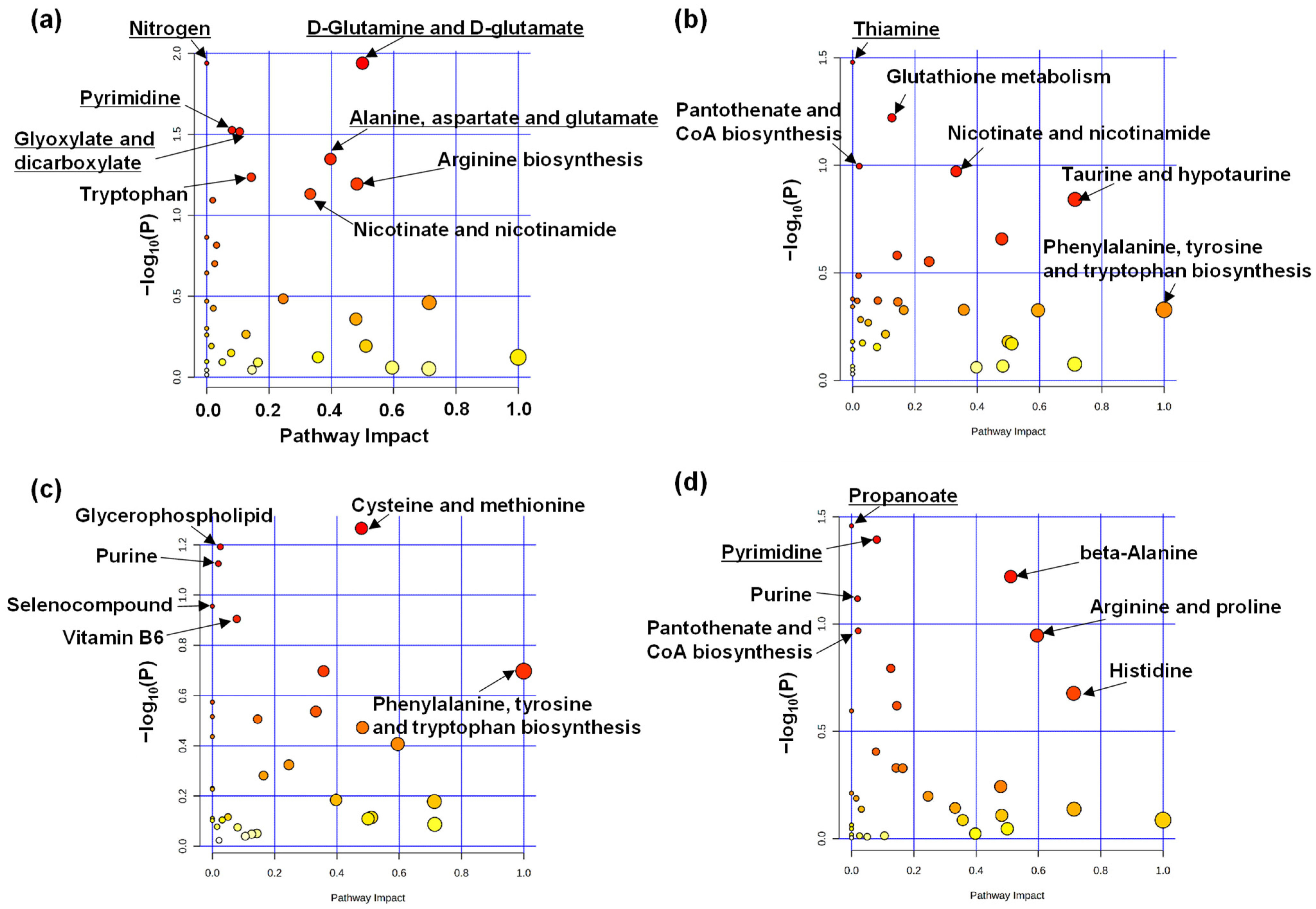
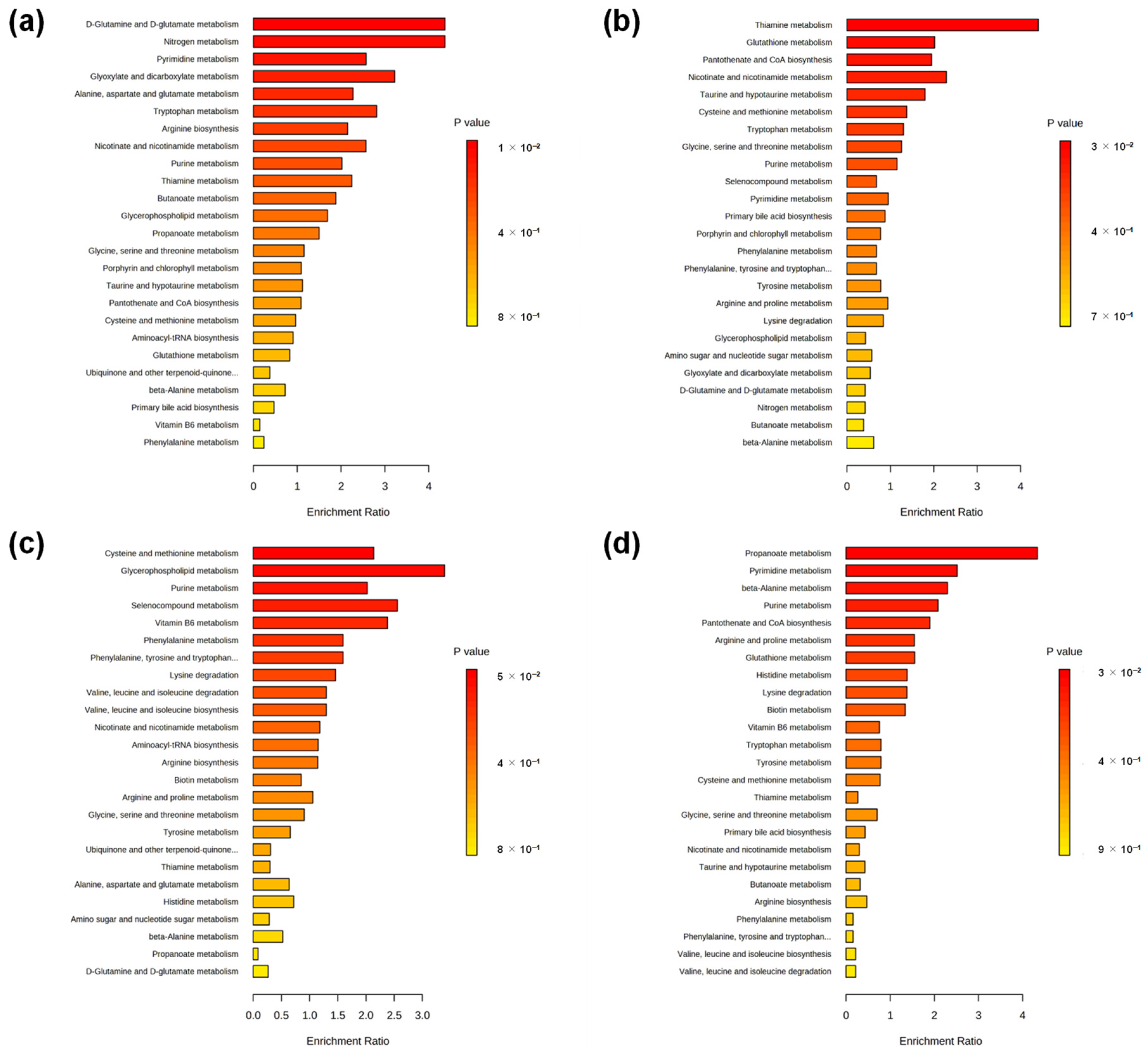
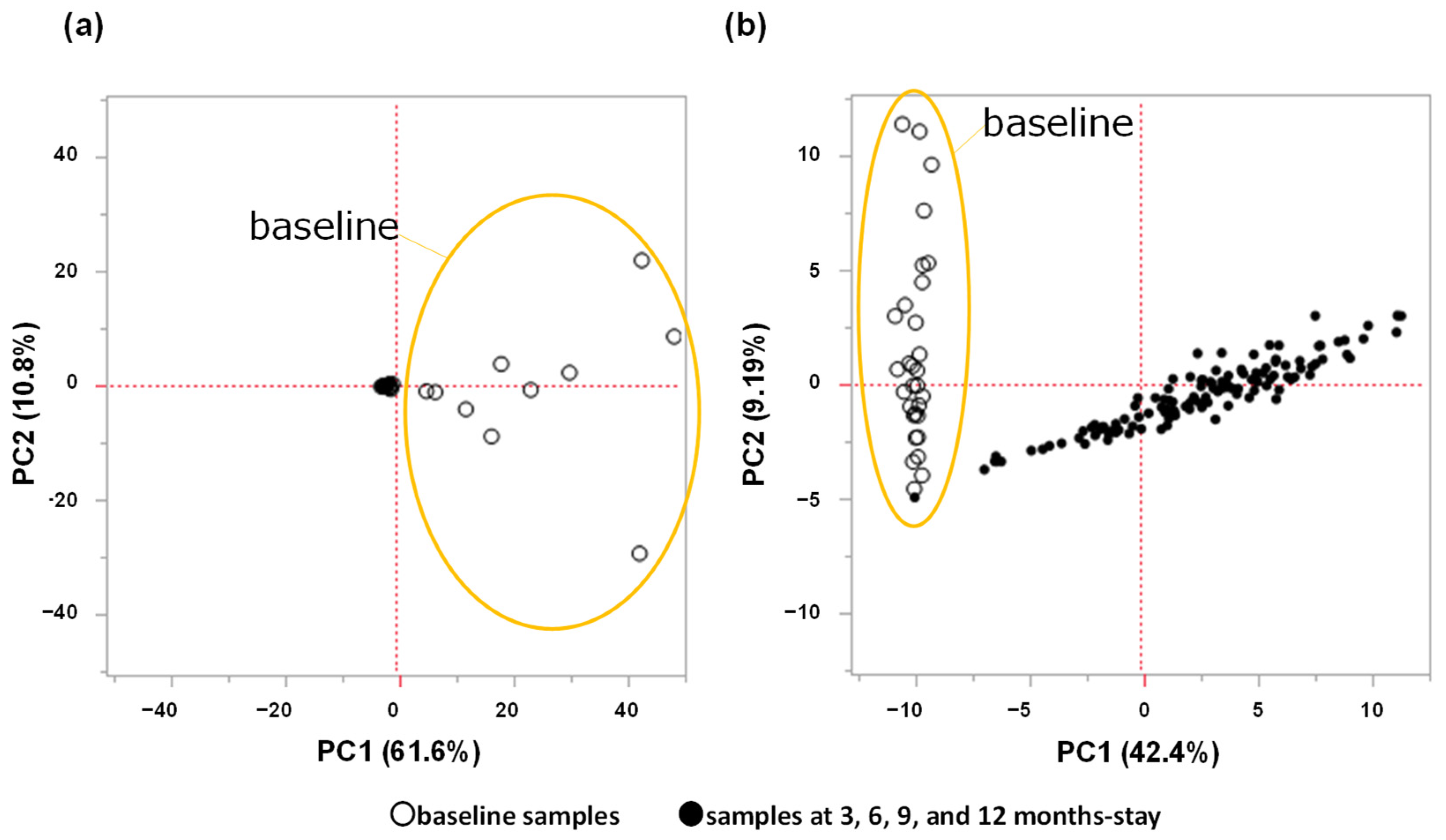
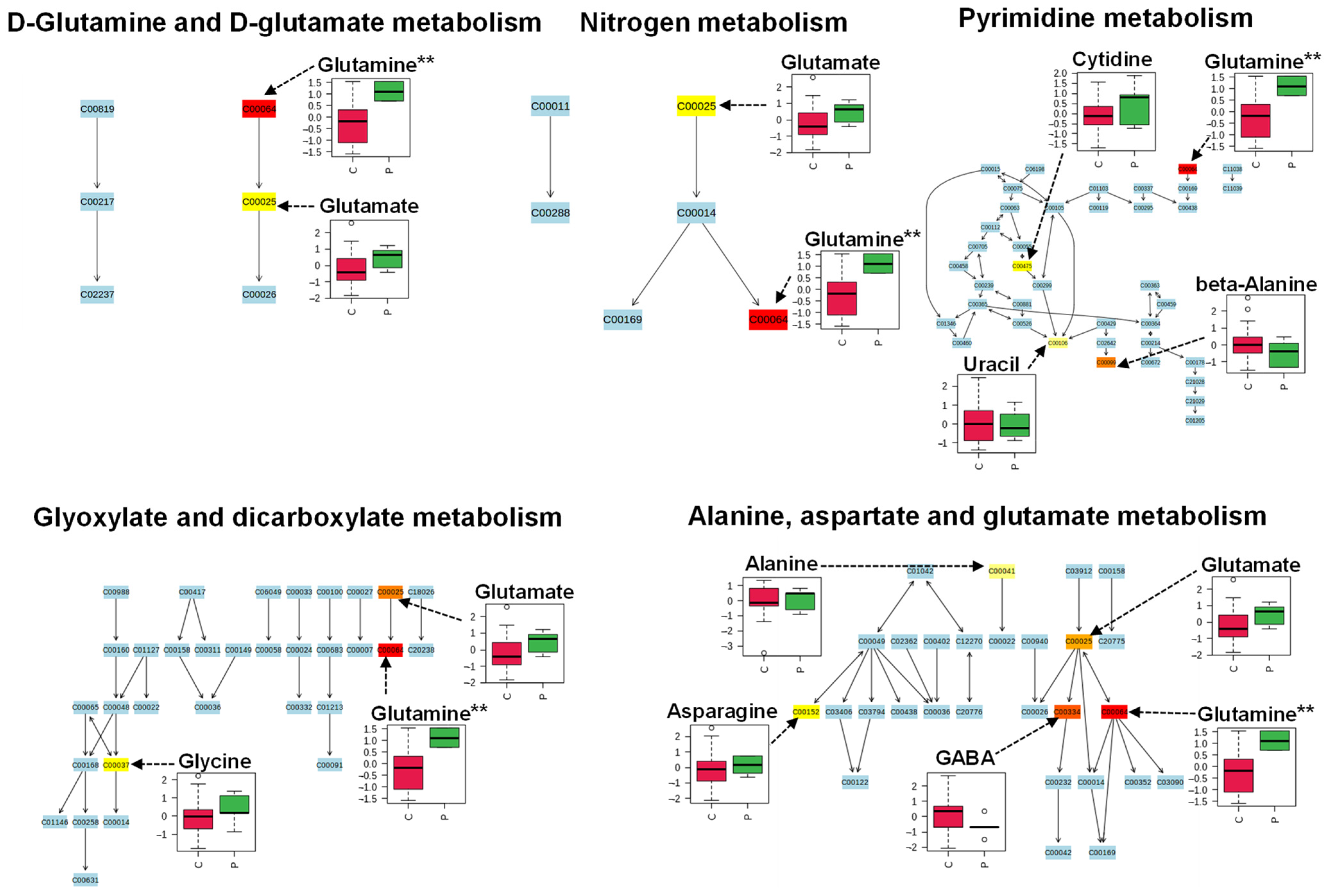
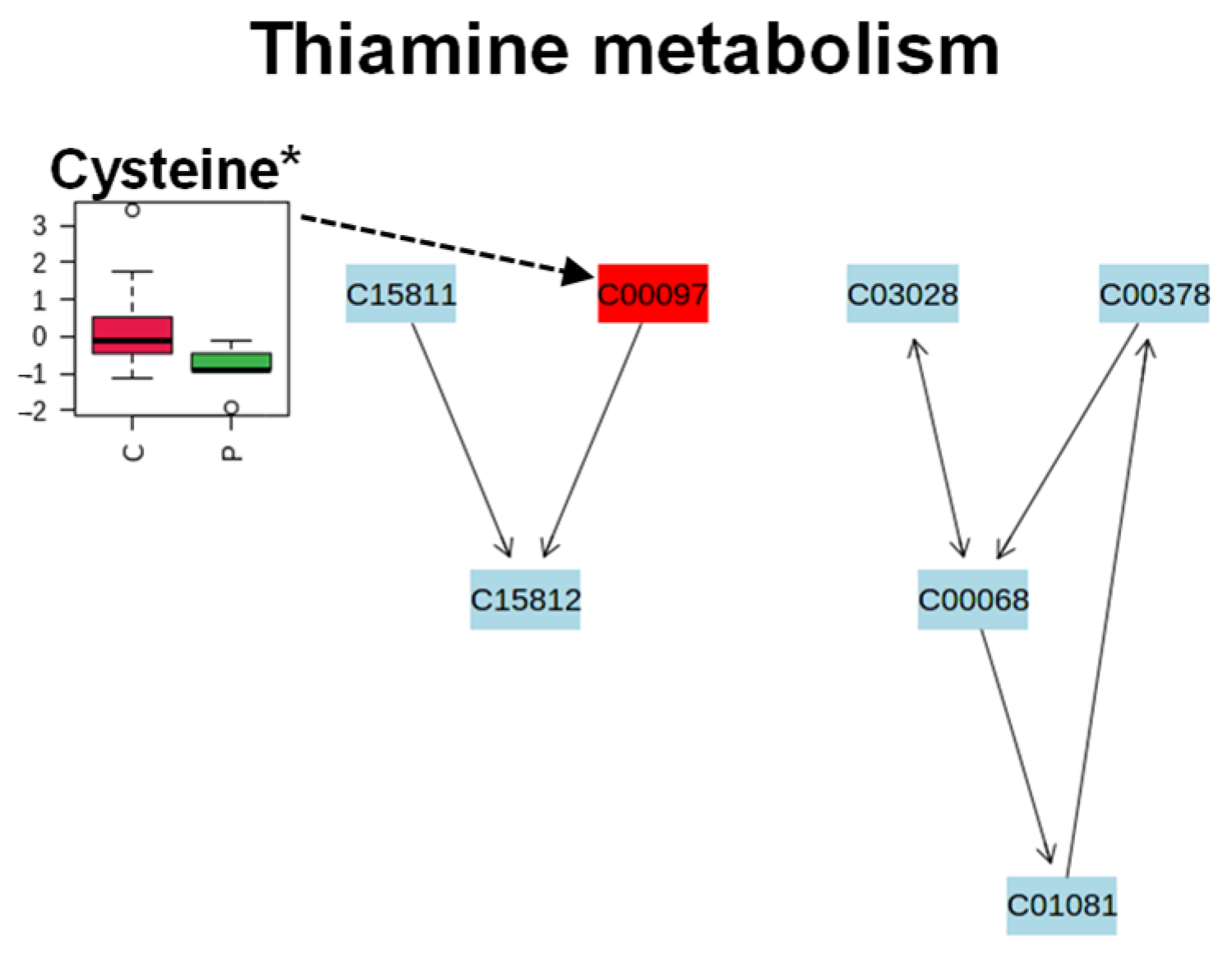
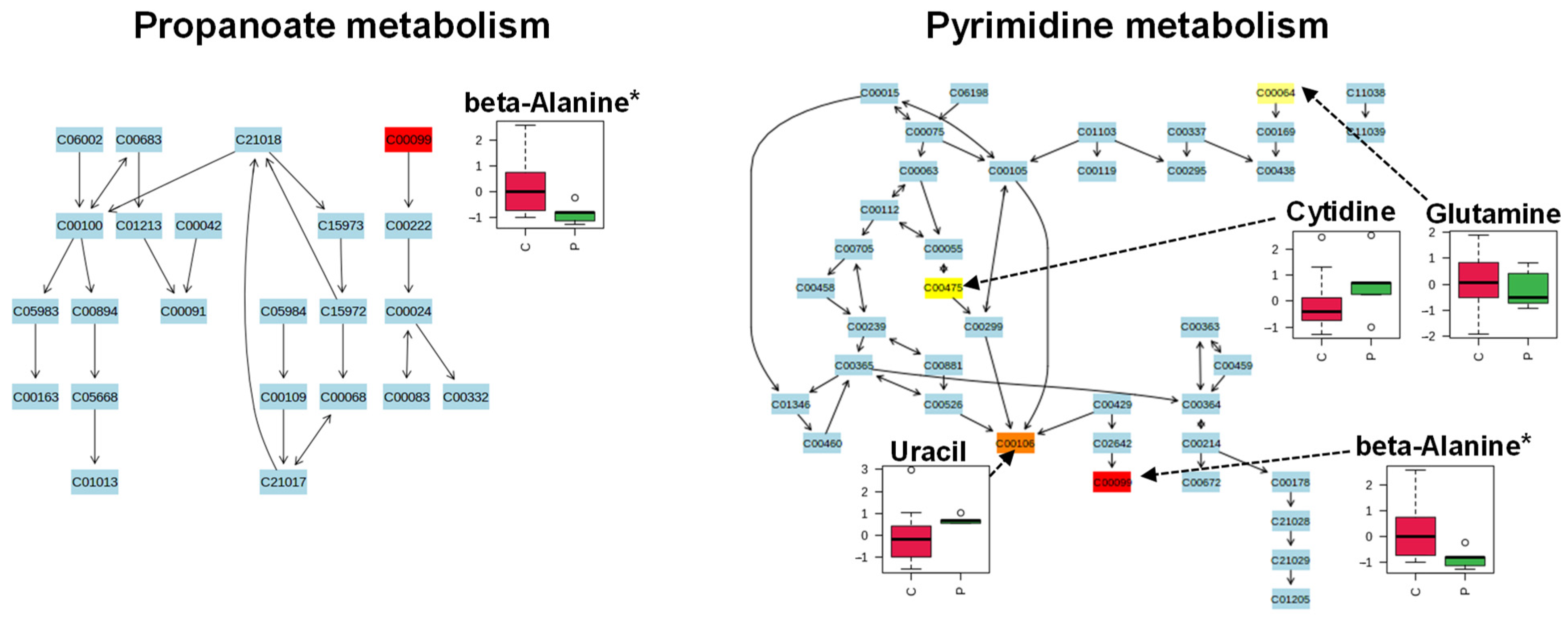
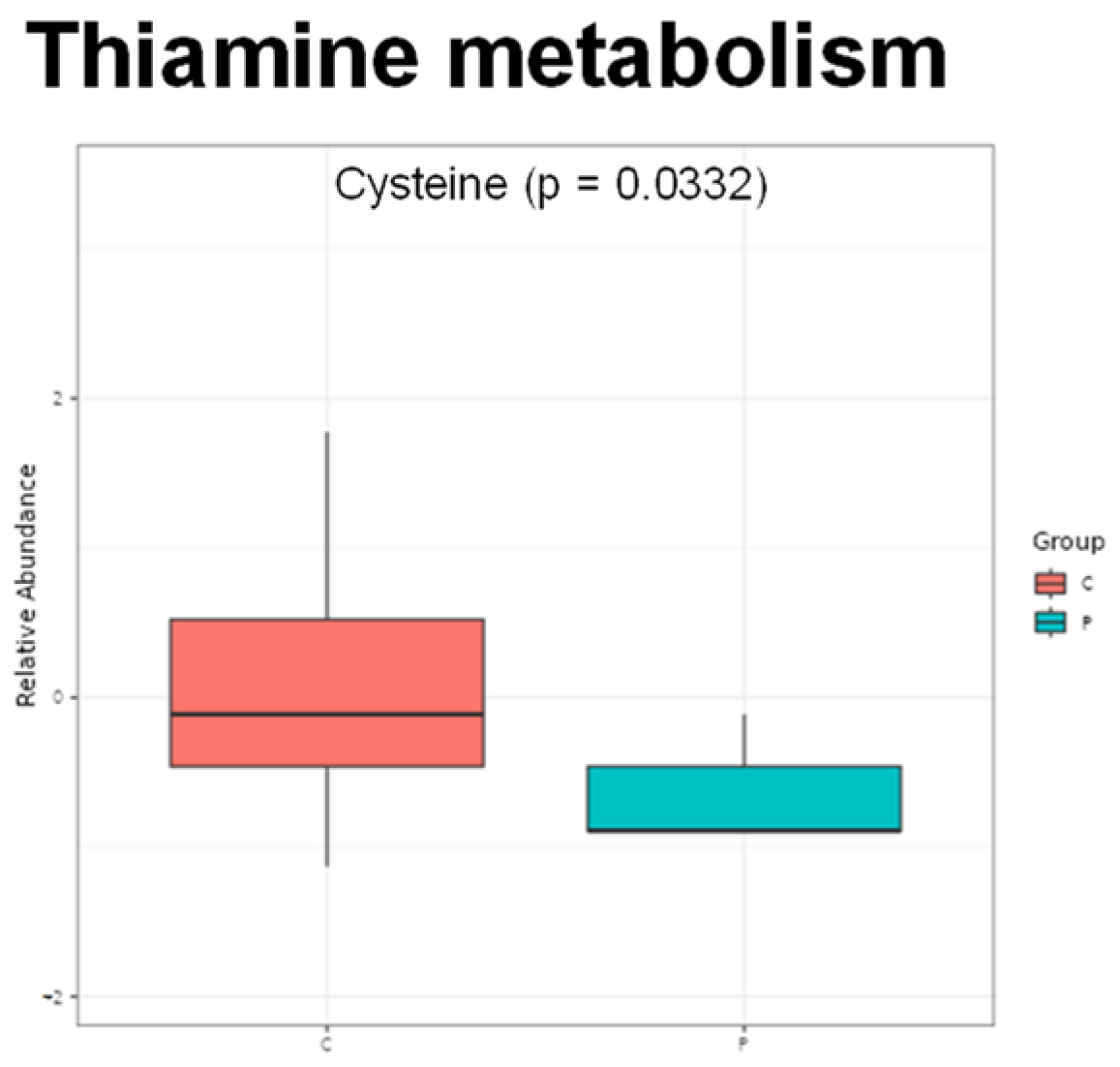
2. Results
3. Discussion
4. Materials and Methods
4.1. Subjects and Sample Collections
4.2. Sample Preparation for Metabolomic Analysis
4.3. Instrument Parameters for Metabolomic Analysis
4.4. Data Processing
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A








References
- Rothblum, E.D. Psychological factors in the antarctic. J. Psychol. 1990, 124, 253–273. [Google Scholar] [CrossRef] [PubMed]
- Mullin, C.S., Jr. Some psychological aspects of isolated Antarctic living. Am. J. Psychiatry 1960, 117, 323–325. [Google Scholar] [CrossRef] [PubMed]
- Palinkas, L.A.; Suedfeld, P. Psychological effects of polar expeditions. Lancet 2008, 371, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Palinkas, L.A.; Glogower, F.; Dembert, M.; Hansen, K.; Smullen, R. Incidence of psychiatric disorders after extended residence in Antarctica. Int. J. Circumpolar. Health 2004, 63, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norman, J.N. A comparison of the patterns of illness and injury occurring on offshore structures in the northern North Sea and the stations of the British Antarctic Survey. Arctic Med. Res. 1991, 719–721. [Google Scholar]
- Zimmer, M.; Cabral, J.C.C.R.; Borges, F.C.; Côco, K.G.; Hameister, B.d.R. Psychological changes arising from an Antarctic stay: Systematic overview. Estud. Psicol. (Camp.) 2013, 30, 415–423. [Google Scholar] [CrossRef]
- Palinkas, L.A.; Cravalho, M.; Browner, D. Seasonal variation of depressive symptoms in Antarctica. Acta Psychiatr. Scand. 1995, 91, 423–429. [Google Scholar] [CrossRef]
- Palinkas, L.A. Effects of physical and social environments on the health and well-being of Antarctic winter-over personnel. Environ. Behav. 1991, 23, 782–799. [Google Scholar] [CrossRef]
- Reed, H.L.; Reedy, K.R.; Palinkas, L.A.; Van Do, N.; Finney, N.S.; Case, H.S.; LeMar, H.J.; Wright, J.; Thomas, J. Impairment in cognitive and exercise performance during prolonged antarctic residence: Effect of thyroxine supplementation in the polar triiodothyronine syndrome. J. Clin. Endocrinol. Metab. 2001, 86, 110–116. [Google Scholar] [CrossRef]
- Weiss, K.; Suedfeld, P.; Steel, G.D.; Tanaka, M. Psychological adjustment during three Japanese Antarctic research expeditions. Environ. Behav. 2000, 32, 142–156. [Google Scholar] [CrossRef]
- Ikegawa, M.; Kimura, M.; Makita, K.; Itokawa, Y. Psychological studies of a Japanese winter-over group at Asuka Station, Antarctica. Aviat Space Environ. Med. 1998, 69, 452–460. [Google Scholar] [PubMed]
- Alfano, C.A.; Bower, J.L.; Connaboy, C.; Agha, N.H.; Baker, F.L.; Smith, K.A.; So, C.J.; Simpson, R.J. Mental health, physical symptoms and biomarkers of stress during prolonged exposure to Antarctica’s extreme environment. Acta Astronaut. 2021, 181, 405–413. [Google Scholar] [CrossRef]
- Moraes, M.M.; Bruzzi, R.S.; Martins, Y.A.T.; Mendes, T.T.; Maluf, C.B.; Ladeira, R.V.P.; Núñez-Espinosa, C.; Soares, D.D.; Wanner, S.P.; Arantes, R.M.E. Hormonal, autonomic cardiac and mood states changes during an Antarctic expedition: From ship travel to camping in Snow Island. Physiol. Behav. 2020, 224, 113069. [Google Scholar] [CrossRef] [PubMed]
- Tingate, T.R.; Lugg, D.J.; Muller, H.K.; Stowe, R.P.; Pierson, D.L. Antarctic isolation: Immune and viral studies. Immunol. Cell Biol. 1997, 75, 275–283. [Google Scholar] [CrossRef]
- Shearer, W.T.; Lee, B.N.; Cron, S.G.; Rosenblatt, H.M.; Smith, E.O.; Lugg, D.J.; Nickolls, P.M.; Sharp, R.M.; Rollings, K.; Reuben, J.M. Suppression of human anti-inflammatory plasma cytokines IL-10 and IL-1RA with elevation of proinflammatory cytokine IFN-gamma during the isolation of the Antarctic winter. J. Allergy Clin. Immunol. 2002, 109, 854–857. [Google Scholar] [CrossRef]
- Shirai, T.; Magara, K.K.; Motohashi, S.; Yamashita, M.; Kimura, M.; Suwazomo, Y.; Nogawa, K.; Kuriyama, T.; Taniguchi, M.; Nakayama, T. TH1-biased immunity induced by exposure to Antarctic winter. J. Allergy Clin. Immunol. 2003, 111, 1353–1360. [Google Scholar] [CrossRef]
- Xu, C.; Zhu, G.; Xue, Q.; Zhang, S.; Du, G.; Xi, Y.; Palinkas, L.A. Effect of the Antarctic environment on hormone levels and mood of Chinese expeditioners. Int. J. Circumpolar. Health 2003, 62, 255–267. [Google Scholar] [CrossRef]
- Shih, P.B. Metabolomics Biomarkers for Precision Psychiatry. Adv. Exp. Med. Biol. 2019, 1161, 101–113. [Google Scholar] [CrossRef]
- Lin, L.; Chen, X.M.; Liu, R.H. Novel urinary metabolite signature for diagnosing postpartum depression. Neuropsychiatr. Dis. Treat. 2017, 13, 1263–1270. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Han, Y.; Hong, Y.; Li, W.W.; Pei, Q.; Zhou, X.; Zhang, B.; Wang, Y. Identification of Potential Metabolite Markers for Middle-Aged Patients with Post-Stroke Depression Using Urine Metabolomics. Neuropsychiatr. Dis. Treat. 2020, 16, 2017–2024. [Google Scholar] [CrossRef]
- Chen, J.J.; Xie, J.; Li, W.W.; Bai, S.J.; Wang, W.; Zheng, P.; Xie, P. Age-specific urinary metabolite signatures and functions in patients with major depressive disorder. Aging (Albany N. Y.) 2019, 11, 6626–6637. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Zhou, C.J.; Liu, Z.; Fu, Y.Y.; Zheng, P.; Yang, D.Y.; Li, Q.; Mu, J.; Wei, Y.D.; Zhou, J.J.; et al. Divergent Urinary Metabolic Phenotypes between Major Depressive Disorder and Bipolar Disorder Identified by a Combined GC-MS and NMR Spectroscopic Metabonomic Approach. J. Proteome Res. 2015, 14, 3382–3389. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Liu, Z.; Fan, S.H.; Yang, D.Y.; Zheng, P.; Shao, W.H.; Qi, Z.G.; Xu, X.J.; Li, Q.; Mu, J.; et al. Combined application of NMR- and GC-MS-based metabonomics yields a superior urinary biomarker panel for bipolar disorder. Sci. Rep. 2014, 4, 5855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamura, N.; Shinoda, K.; Sato, H.; Sasaki, K.; Suzuki, M.; Yamaki, K.; Fujimori, T.; Yamamoto, H.; Osei-Hyiaman, D.; Ohashi, Y. Plasma metabolome analysis of patients with major depressive disorder. Psychiatry Clin. Neurosci. 2018, 72, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Homorogan, C.; Nitusca, D.; Enatescu, V.; Schubart, P.; Moraru, C.; Socaciu, C.; Marian, C. Untargeted Plasma Metabolomic Profiling in Patients with Major Depressive Disorder Using Ultra-High Performance Liquid Chromatography Coupled with Mass Spectrometry. Metabolites 2021, 11, 466. [Google Scholar] [CrossRef]
- Marescau, B.; De Deyn, P.P.; Holvoet, J.; Possemiers, I.; Nagels, G.; Saxena, V.; Mahler, C. Guanidino compounds in serum and urine of cirrhotic patients. Metabolism 1995, 44, 584–588. [Google Scholar] [CrossRef]
- Heath, A.; Lindberg, D.R.; Makowiecki, K.; Gray, A.; Asp, A.J.; Rodger, J.; Choi, D.-S.; Croarkin, P.E. Medium-and high-intensity rTMS reduces psychomotor agitation with distinct neurobiologic mechanisms. Transl Psychiatry 2018, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Mo, Y.Y.; Feng, S.S.; Meng, M.W.; Chen, S.Y.; Huang, H.M.; Ling, X.; Song, H.; Liang, Y.H.; Ou, S.F.; et al. Urinary metabonomics study of anti-depressive mechanisms of Millettia speciosa Champ on rats with chronic unpredictable mild stress-induced depression. J. Pharm. Biomed. Anal. 2021, 205, 114338. [Google Scholar] [CrossRef]
- Manabe, S.; Kurroda, I.; Okada, K.; Morishima, M.; Okamoto, M.; Harada, N.; Takahashi, A.; Sakai, K.; Nakaya, Y. Decreased blood levels of lactic acid and urinary excretion of 3-methylhistidine after exercise by chronic taurine treatment in rats. J. Nutr. Sci. Vitaminol. (Tokyo) 2003, 49, 375–380. [Google Scholar] [CrossRef] [Green Version]
- Cao, B.; Deng, R.; Wang, D.; Li, L.; Ren, Z.; Xu, L.; Gao, X. Association between arginine catabolism and major depressive disorder: A protocol for the systematic review and meta-analysis of metabolic pathway. Medicine (Baltimore) 2020, 99, e21068. [Google Scholar] [CrossRef]
- MacDonald, K.; Krishnan, A.; Cervenka, E.; Hu, G.; Guadagno, E.; Trakadis, Y. Biomarkers for major depressive and bipolar disorders using metabolomics: A systematic review. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2019, 180, 122–137. [Google Scholar] [CrossRef]
- Zarate, C.A., Jr.; Singh, J.; Manji, H.K. Cellular plasticity cascades: Targets for the development of novel therapeutics for bipolar disorder. Biol. Psychiatry 2006, 59, 1006–1020. [Google Scholar] [CrossRef] [PubMed]
- Auer, D.P.; Pütz, B.; Kraft, E.; Lipinski, B.; Schill, J.; Holsboer, F. Reduced glutamate in the anterior cingulate cortex in depression: An in vivo proton magnetic resonance spectroscopy study. Biol. Psychiatry 2000, 47, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Nirwan, M. Human psychophysiology in Antarctica. Sri. Ramachandra J. Health Sci. 2022, 2, 12–18. [Google Scholar] [CrossRef]
- Spinelli, E.; Werner, J.J. Human adaptative behavior to Antarctic conditions: A review of physiological aspects. WIREs Mech. Dis. 2022, 14, e1556. [Google Scholar] [CrossRef]
- Iuliano-Burns, S.; Wang, X.F.; Ayton, J.; Jones, G.; Seeman, E. Skeletal and hormonal responses to sunlight deprivation in Antarctic expeditioners. Osteoporos. Int. 2009, 20, 1523–1528. [Google Scholar] [CrossRef]
- Smith, S.M.; Gardner, K.K.; Locke, J.; Zwart, S.R. Vitamin D supplementation during Antarctic winter. Am. J. Clin. Nutr. 2009, 89, 1092–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, A.P.; Mishra, K.P.; Ganju, L.; Singh, S.B. Wintering in Antarctica: Impact on immune response of Indian expeditioners. Neuroimmunomodulation 2012, 19, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, A.S.; Pal, K.; Prasad, R.; Bajaj, A.C.; Kumar, R.; Sawhney, R.C. Plasma insulin and growth hormone during antarctic residence. Jpn. J. Physiol. 1998, 48, 167–169. [Google Scholar] [CrossRef] [Green Version]
- Homandberg, L.K.; Fuller-Rowell, T.E. Experiences of discrimination and urinary catecholamine concentrations: Longitudinal associations in a college student sample. Ann. Behav. Med. 2020, 54, 843–852. [Google Scholar] [CrossRef]
- Kokka, I.; Chrousos, G.P.; Darviri, C.; Bacopoulou, F. Measuring Adolescent Chronic Stress: A Review of Established Biomarkers and Psychometric Instruments. Horm. Res. Paediatr. 2022. [Google Scholar] [CrossRef]
- Gelenberg, A.J.; Gibson, C.J. Tyrosine for the treatment of depression. Nutr. Health 1984, 3, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Tomita, A.; Mori, M.; Hiwatari, K.; Yamaguchi, E.; Itoi, T.; Sunamura, M.; Soga, T.; Tomita, M.; Sugimoto, M. Effect of storage conditions on salivary polyamines quantified via liquid chromatography-mass spectrometry. Sci. Rep. 2018, 8, 12075. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Tanaka, T.; Nakagawa, S.; Nakato, Y.; Kameyama, R.; Boku, S.; Toda, H.; Kurita, T.; Koyama, T. Utility and limitations of PHQ-9 in a clinic specializing in psychiatric care. BMC Psychiatry 2012, 12, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Average/n/h | Standard Deviation |
|---|---|---|
| Age | 40.2 | 9.2 |
| Sex | M 29, F 1 | |
| Education Years | 15.3 | 2.2 |
| Subjective Social Status 1 | 5.5 | 1.2 |
| Marital Status | Married 18, Unmarried 11 | |
| Presence of Physical Disease | Y 1, N 29 | |
| Presence of Psychiatric Disease | Y 0, N 30 | |
| History of Psychiatric Disease | Y 2, N 28 | |
| Family history of Psychiatric Disease | Y 2, N 28 | |
| Alcohol Drinking | Y 24, N 6 | |
| Smoking | Y 6, N 24 | |
| Overtime work (h/month) | ||
| ≤20 | 19 | |
| ≤30 | 4 | |
| ≤40 | 4 | |
| ≤50 | 2 | |
| Parental Bonding Instrument | ||
| Paternal Care | 21.7 | 6.7 |
| Paternal Overprotection | 10.8 | 5.7 |
| Maternal Care | 26.4 | 5.4 |
| Maternal Overprotection | 11.5 | 6.2 |
| PHQ-9 total scores 2 | 3 | 3.5 |
| State anxiety of STAI-Y | 41.7 | 9.2 |
| Trait anxiety of STAI-Y | 40.1 | 9.6 |
| Neuroticism (0−12) | 3.2 | 3 |
| Pittsburgh Sleep Quality Index Global Scores | 4.5 | 1.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasuya, K.; Imura, S.; Ishikawa, T.; Sugimoto, M.; Inoue, T. Relationship between Urinary Metabolomic Profiles and Depressive Episode in Antarctica. Int. J. Mol. Sci. 2023, 24, 943. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24020943
Kasuya K, Imura S, Ishikawa T, Sugimoto M, Inoue T. Relationship between Urinary Metabolomic Profiles and Depressive Episode in Antarctica. International Journal of Molecular Sciences. 2023; 24(2):943. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24020943
Chicago/Turabian StyleKasuya, Kazuhiko, Satoshi Imura, Takashi Ishikawa, Masahiro Sugimoto, and Takeshi Inoue. 2023. "Relationship between Urinary Metabolomic Profiles and Depressive Episode in Antarctica" International Journal of Molecular Sciences 24, no. 2: 943. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24020943