

Circulating Histones to Detect and Monitor the Progression of Cancer
 ,
,  , ,
, ,
Abstract
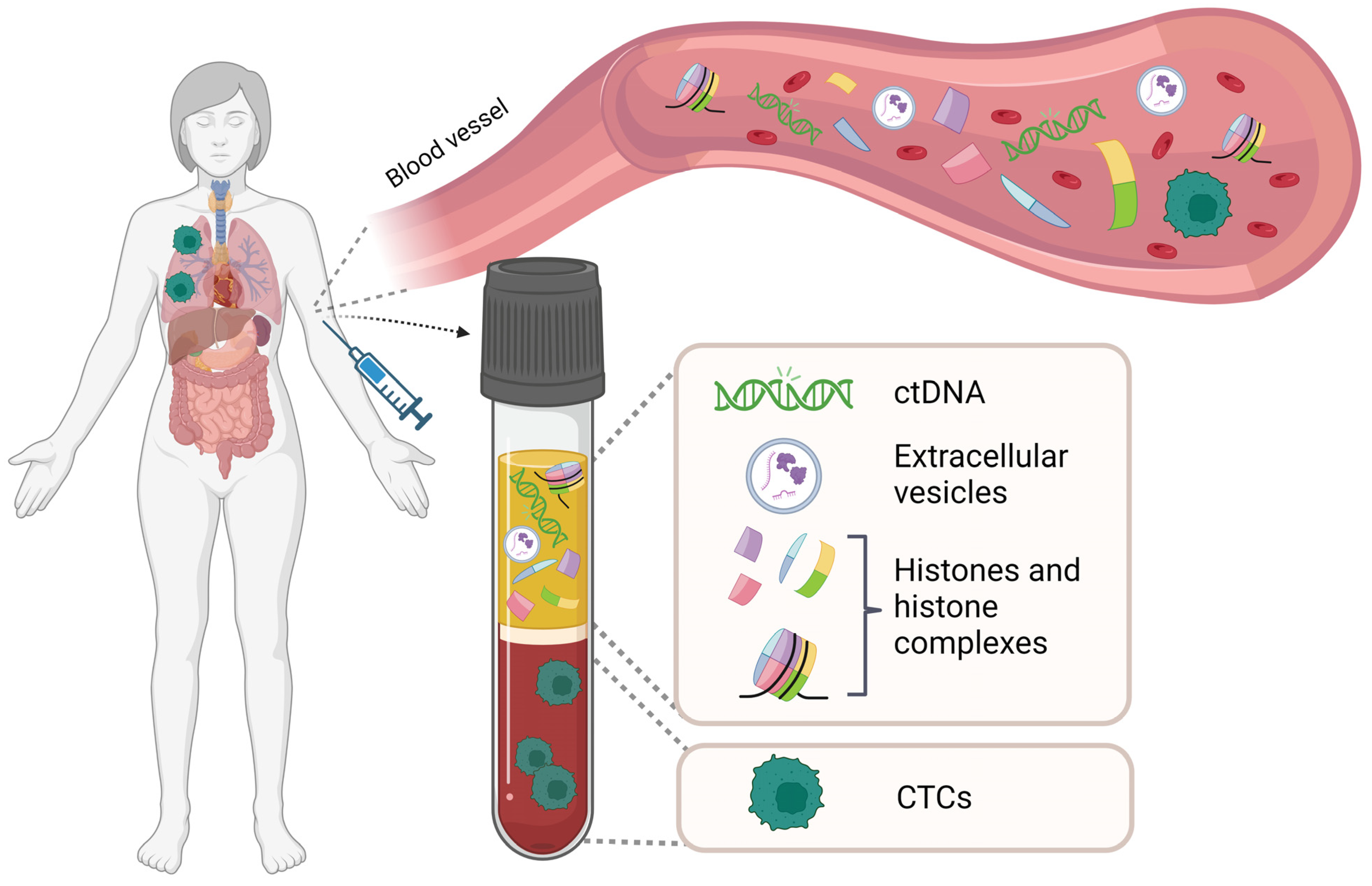
:1. Introduction
1.1. Liquid Biopsy
1.2. Circulating Histones
Methods of Detection
2. Tracing the Tissue of Origin of Circulating Histones
3. Circulating Histones in Hematological Malignancies: Markers, Predictors, and Therapeutic Potential
4. Circulating Histones in Solid Cancers: Detection, Monitoring, and Tumorigenesis
4.1. Cancer Detection

{kind=link}
| Malignancy | Target for Detection | Detection Method | Level/Value | Suggested Function | Reference |
|---|---|---|---|---|---|
| Breast | Anti-histones + anti-DNA antibodies | ELISA | High | Detection | [124] |
| Lung, colorectal, and other gastrointestinal cancers;breast, ovarian, and other gynecological cancers;lymphoma, renal, prostate, and other non-defined cancers | Anti-histones + anti-DNA antibodies | ELISA | High | Detection | [118] |
| Small-cell lung cancer Head and neck cancer | Anti-histones + anti-DNA antibodies | ELISA | High + consistent decrease | Chemotherapy response | [118] |
| Pancreatic cancer | Anti-histones + anti-DNA antibodies | ELISA | Fluctuation (high-low-high) | Disease progression following chemotherapy | [118] |
| CRC | Anti-histones + anti-DNA antibodies | ELISA | High + consistent decrease | Radiochemotherapy response | [125] |
| Breast | Anti-histones + anti-DNA antibodies | ELISA | High/low | Response prediction to neoadjuvant chemotherapy | [126] |
| Breast | Anti-histones + anti-DNA antibodies | ELISA | High | Disease progression | [127] |
| Advanced malignancies | Anti-Citrullinated histone H3 | Flow-cytometry | High/low | Prognostic marker | [88] |
| CRC | H2AK119Ub, H3K9Ac, H3K27Ac, and the global level of nucleosomes | ELISA | High Score-the 4 marks combined | Detection | [84] |
| Pancreatic stage II | 5MC, H2AZ, H3K4Me2, H2AK119Ub and mH2A1.1 | ELISA | High Score-the 5 marks combined | Detection | [85] |
| Colorectal, pancreatic, breast, and lung cancer patients | H3K9me3 and H4K20me3 | Chromatin- immunoprecipitation | High total levels | Detection H4K20me3-Cancer vs. healthy H4K20me3-CRC vs. benign gastrointestinal diseases | [130] |
| CRC | H3K9me3 and H4K20me3 | Chromatin- immunoprecipitation | Low, when normalized to total nucleosome content | Detection | [130] |
| Breast | H3K9me3 and H4K20me3 | Chromatin- immunoprecipitation | High, when normalized to total nucleosome content | Detection | [130] |
| CRC | H4K20me3 | ELISA | Low | Detection | [82] |
| CRC | H3K27me3 | ELISA | Low | Detection | [82] |
| Ovarian | Nucleosomal DNA fragmentation pattern | WGS sequencing and bioinformatics | NA | Detection, specifically cancers with low chromosomal instability | [131] |
| Breast, colorectal, lung, ovarian, pancreatic, gastric, bile duct | Nucleosomal DNA fragmentation pattern | Genome-wide cell-free DNA fragmentation | NA | Detection | [132] |
| Hepatocellular carcinoma (HCC) | H3, the canonical H3.1 variant, H3K27me3 and H3K36me3 | ELISA | H3K27me3/H3K36me3 ratio-High in disease progression H3K27me3, H3K36me3-low at best therapy response | Responde to sorafenib; Monitoring and disease progression | [133] |
| Prostate | H3K27me3 | ELISA | Very low in metastatic disease | Stage differentiation | [134] |
| Lung, colorectal, and other gastrointestinal cancers; breast, ovarian, and other gynecological cancers; lymphoma, renal, prostate, and other non-defined, cervical, and pancreatic cancers | Anti-histone and anti-DNA | ELISA | High and further increase | Disease progression; therapy response | [118,125,135,136,137,138,139,140,141] |
| HCC | Anti-histones + anti-DNA antibodies | ELISA | High; Increased | Radiofrequency ablation (RFA) therapy response | [142] |
| Non-small-cell lung cancer | Nucleosomal DNA fragmentation pattern | WGS sequencing and bioinformatics | Changes in epigenetic profile | anti-EGFR, anti-ERBB2 response monitoring | [132] |
| Breast | Nucleosomal DNA fragmentation pattern | WGS sequencing and bioinformatics | Changes in epigenetic profile | Estrogen receptor subtyping | [143] |
| Breast | Anti-histone and anti-DNA | ELISA | High | Chemotherapy response; Disease progression | [126] |
| Not specified | 5mC and H3K9Me3 | ELISA | Low | Detection | [144] |
| CRC | Nucleosome antibody and antibody against 5mC | ELISA | Low | Detection | [79] |
| CRC Pancreas, lung, and breast | H3K27me3, H3K9me3, H3K9ac, H3K4me3, H3K36me3, H3K4me1, and H3K27me3 | Single-molecule imaging | High; Decrease in some combinatorial patterns (e.g., H3K9me3- and H3K36me3 | Detection | [92] |
| CRC | Anti-histone H3.1 antibody; H3K27Me1, -Me2, -Me3; H3K36Me1, -Me2, -Me3; H3K56Me2 H3K27Ac, H4K20Me1, Me2; H4K4; 17_2Ac, -3Ac, -4Ac; and H2A1R3Cit; H3, H4, H2A1 | Nucleosome immunoprecipitation; LC-MS/MS | High | Detection | [91] |
| Variety of advanced malignancies | Citrullinated histone H3 (H3Cit); Anti-histone H3 and anti-H3Cit antibodies | ELISA | High | Detection | [88] |
| Cervical | Anti-histone and anti-DNA | ELISA | High + decrease | Chemotherapy response | [136] |
4.2. Treatment Guidance, Disease, and Therapy Response Monitoring
4.3. Role in Disease Progression
5. Market Size and Private Investments
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cescon, D.W.; Bratman, S.V.; Chan, S.M.; Siu, L.L. Circulating tumor DNA and liquid biopsy in oncology. Nat. Cancer 2020, 1, 276–290. [Google Scholar] [CrossRef] [PubMed]
- Dang, D.K.; Park, B.H. Circulating tumor DNA: Current challenges for clinical utility. J. Clin. Investig. 2022, 132, e154941. [Google Scholar] [CrossRef] [PubMed]
- Parikh, A.R.; Leshchiner, I.; Elagina, L.; Goyal, L.; Levovitz, C.; Siravegna, G.; Livitz, D.; Rhrissorrakrai, K.; Martin, E.E.; Van Seventer, E.E.; et al. Liquid versus tissue biopsy for detecting acquired resistance and tumor heterogeneity in gastrointestinal cancers. Nat. Med. 2019, 25, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Alix-Panabières, C.; Pantel, K. Liquid Biopsy: From Discovery to Clinical Application. Cancer Discov. 2021, 11, 858–873. [Google Scholar] [CrossRef] [PubMed]
- Cisneros-Villanueva, M.; Hidalgo-Pérez, L.; Rios-Romero, M.; Cedro-Tanda, A.; Ruiz-Villavicencio, C.A.; Page, K.; Hastings, R.; Fernandez-Garcia, D.; Allsopp, R.; Fonseca-Montaño, M.A.; et al. Cell-free DNA analysis in current cancer clinical trials: A review. Br. J. Cancer. 2022, 126, 391–400. [Google Scholar] [CrossRef]
- Barault, L.; Amatu, A.; Siravegna, G.; Ponzetti, A.; Moran, S.; Cassingena, A.; Mussolin, B.; Falcomatà, C.; Binder, A.M.; Cristiano, C.; et al. Discovery of methylated circulating DNA biomarkers for comprehensive non-invasive monitoring of treatment response in metastatic colorectal cancer. Gut 2018, 67, 1995–2005. [Google Scholar] [CrossRef]
- Tan, L.; Sandhu, S.; Lee, R.; Li, J.; Callahan, J.; Ftouni, S.; Dhomen, N.; Middlehurst, P.; Wallace, A.; Raleigh, J.; et al. Prediction and monitoring of relapse in stage III melanoma using circulating tumor DNA. Ann. Oncol. 2019, 30, 804–814. [Google Scholar] [CrossRef]
- Tie, J.; Cohen, J.D.; Wang, Y.; Li, L.; Christie, M.; Simons, K.; Elsaleh, H.; Kosmider, S.; Wong, R.; Yip, D.; et al. Serial circulating tumour DNA analysis during multimodality treatment of locally advanced rectal cancer: A prospective biomarker study. Gut 2019, 68, 663–671. [Google Scholar] [CrossRef]
- Wan, J.C.M.; Mughal, T.I.; Razavi, P.; Dawson, S.-J.; Moss, E.L.; Govindan, R.; Tan, I.B.; Yap, Y.-S.; Robinson, W.A.; Morris, C.D.; et al. Liquid biopsies for residual disease and recurrence. Med 2021, 2, 1292–1313. [Google Scholar] [CrossRef]
- Tie, J.; Cohen, J.D.; Lahouel, K.; Lo, S.N.; Wang, Y.; Kosmider, S.; Wong, R.; Shapiro, J.; Lee, M.; Harris, S.; et al. Circulating Tumor DNA Analysis Guiding Adjuvant Therapy in Stage II Colon Cancer. N. Engl. J. Med. 2022, 386, 2261–2272. [Google Scholar] [CrossRef]
- Sartore-Bianchi, A.; Pietrantonio, F.; Lonardi, S.; Mussolin, B.; Rua, F.; Crisafulli, G.; Bartolini, A.; Fenocchio, E.; Amatu, A.; Manca, P.; et al. Circulating tumor DNA to guide rechallenge with panitumumab in metastatic colorectal cancer: The phase 2 CHRONOS trial. Nat. Med. 2022, 28, 1612–1618. [Google Scholar] [CrossRef]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging Biological Principles of Metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [Green Version]
- Visal, T.H.; Hollander, P.D.; Cristofanilli, M.; Mani, S.A. Circulating tumour cells in the -omics era: How far are we from achieving the ‘singularity’? Br. J. Cancer. 2022, 127, 173–184. [Google Scholar] [CrossRef]
- Hofman, V.; Ilie, M.I.; Long, E.; Selva, E.; Bonnetaud, C.; Molina, T.; Venissac, N.; Mouroux, J.; Vielh, P.; Hofman, P. Detection of circulating tumor cells as a prognostic factor in patients undergoing radical surgery for non-small-cell lung carcinoma: Comparison of the efficacy of the CellSearch Assay™ and the isolation by size of epithelial tumor cell method. Int. J. Cancer 2011, 129, 1651–1660. [Google Scholar] [CrossRef]
- Andreopoulou, E.; Yang, L.-Y.; Rangel, K.M.; Reuben, J.M.; Hsu, L.; Krishnamurthy, S.; Valero, V.; Fritsche, H.A.; Cristofanilli, M. Comparison of assay methods for detection of circulating tumor cells in metastatic breast cancer: AdnaGenAdnaTestBreastCancer Select/Detect™ versus VeridexCellSearch™ system. Int. J. Cancer. 2012, 130, 1590–1597. [Google Scholar] [CrossRef]
- Zavridou, M.; Mastoraki, S.; Strati, A.; Koutsodontis, G.; Klinakis, A.; Psyrri, A.; Lianidou, E. Direct comparison of size-dependent versus EpCAM-dependent CTC enrichment at the gene expression and DNA methylation level in head and neck squamous cell carcinoma. Sci. Rep. 2020, 10, 6551. [Google Scholar] [CrossRef] [Green Version]
- Went, P.T.; Lugli, A.; Meier, S.; Bundi, M.; Mirlacher, M.; Sauter, G.; Dirnhofer, S. Frequent EpCam protein expression in human carcinomas. Hum. Pathol. 2004, 35, 122–128. [Google Scholar] [CrossRef]
- Dalerba, P.; Dylla, S.J.; Park, I.-K.; Liu, R.; Wang, X.; Cho, R.W.; Hoey, T.; Gurney, A.; Huang, E.H.; Simeone, D.M.; et al. Phenotypic characterization of human colorectal cancer stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 10158–10163. [Google Scholar] [CrossRef] [Green Version]
- Sieuwerts, A.M.; Kraan, J.; Bolt, J.; Van Der Spoel, P.; Elstrodt, F.; Schutte, M.; Martens, J.W.M.; Gratama, J.-W.; Sleijfer, S.; Foekens, J.A. Anti-Epithelial Cell Adhesion Molecule Antibodies and the Detection of Circulating Normal-Like Breast Tumor Cells. J. Natl. Cancer Inst. 2009, 101, 61–66. [Google Scholar] [CrossRef]
- Gradilone, A.; Raimondi, C.; Nicolazzo, C.; Petracca, A.; Gandini, O.; Vincenzi, B.; Naso, G.; Aglianò, A.M.; Cortesi, E.; Gazzaniga, P. Circulating tumour cells lacking cytokeratin in breast cancer: The importance of being mesenchymal. J. Cell Mol. Med. 2011, 15, 1066–1070. [Google Scholar] [CrossRef]
- MacArthur, K.M.; Kao, G.D.; Chandrasekaran, S.; Alonso-Basanta, M.; Chapman, C.; Lustig, R.A.; Wileyto, E.P.; Hahn, S.M.; Dorsey, J.F. Detection of Brain Tumor Cells in the Peripheral Blood by a Telomerase Promoter-Based Assay. Cancer Res. 2014, 74, 2152–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanssen, A.; Wagner, J.; Gorges, T.M.; Taenzer, A.; Uzunoglu, F.G.; Driemel, C.; Stoecklein, N.H.; Knoefel, W.T.; Angenendt, S.; Hauch, S.; et al. Characterization of different CTC subpopulations in non-small cell lung cancer. Sci. Rep. 2016, 6, 28010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horimoto, Y.; Tokuda, E.; Murakami, F.; Uomori, T.; Himuro, T.; Nakai, K.; Orihata, G.; Iijima, K.; Togo, S.; Shimizu, H.; et al. Analysis of circulating tumour cell and the epithelial mesenchymal transition (EMT) status during eribulin-based treatment in 22 patients with metastatic breast cancer: A pilot study. J. Transl. Med. 2018, 16, 287. [Google Scholar] [CrossRef] [PubMed]
- Cuatrecasas, M.; Gorostiaga, I.; Riera, C.; Saperas, E.; Llort, G.; Costa, I.; Matias-Guiu, X.; Carrato, C.; Navarro, M.; Pineda, M.; et al. Complete loss of EPCAM immunoexpression identifies EP-CAM deletion carriers in MSH2-negative colorectal neoplasia. Cancers 2020, 12, 2803. [Google Scholar] [CrossRef]
- Gorges, T.M.; Tinhofer, I.; Drosch, M.; Röse, L.; Zollner, T.M.; Krahn, T.; Von Ahsen, O. Circulating tumour cells escape from EpCAM-based detection due to epithelial-to-mesenchymal transition. BMC Cancer 2012, 12, 178. [Google Scholar] [CrossRef] [Green Version]
- Denève, E.; Riethdorf, S.; Ramos, J.; Nocca, D.; Coffy, A.; Daurès, J.-P.; Maudelonde, T.; Fabre, J.-M.; Pantel, K.; Alix-Panabières, C. Capture of viable circulating tumor cells in the liver of colo-rectal cancer patients. Clin. Chem. 2013, 59, 1384–1392. [Google Scholar] [CrossRef] [Green Version]
- Hyun, K.-A.; Koo, G.-B.; Han, H.; Sohn, J.; Choi, W.; Kim, S.-I.; Jung, H.-I.; Kim, Y.-S. Epithelial-to-mesenchymal transition leads to loss of EpCAM and different physical properties in circulating tumor cells from metastatic breast cancer. Oncotarget 2016, 7, 24677–24687. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Le, W.; Wang, Y.; Li, Z.; Wang, D.; Ren, L.; Lin, L.; Cui, S.; Hu, J.J.; Hu, Y.; et al. Targeting Negative Surface Charges of Cancer Cells by Multifunctional Nanoprobes. Theranostics 2016, 6, 1887–1898. [Google Scholar] [CrossRef]
- Deng, Z.; Wu, S.; Wang, Y.; Shi, D. Circulating tumor cell isolation for cancer diagnosis and prognosis. eBioMedicine 2022, 83, 104237. [Google Scholar] [CrossRef]
- Denis, J.A.; Patroni, A.; Guillerm, E.; Pépin, D.; Benali-Furet, N.; Wechsler, J.; Manceau, G.; Bernard, M.; Coulet, F.; Larsen, A.K.; et al. Droplet digital PCR of circulating tumor cells from colorectal cancer patients can predict KRAS mutations before surgery. Mol. Oncol. 2016, 10, 1221–1231. [Google Scholar] [CrossRef]
- An, T.; Qin, S.; Xu, Y.; Tang, Y.; Huang, Y.; Situ, B.; Inal, J.M.; Zheng, L. Exosomes serve as tumour markers for personalized diagnostics owing to their important role in cancer metastasis. J. Extracell. Vesicles 2015, 4, 27522. [Google Scholar] [CrossRef]
- Lane, R.E.; Korbie, D.; Hill, M.M.; Trau, M. Extracellular vesicles as circulating cancer biomarkers: Opportunities and challenges. Clin. Transl. Med. 2018, 7, 14. [Google Scholar] [CrossRef]
- König, L.; Kasimir-Bauer, S.; Bittner, A.-K.; Hoffmann, O.; Wagner, B.; Manvailer, L.F.S.; Kimmig, R.; Horn, P.A.; Rebmann, V. Elevated levels of extracellular vesicles are associated with therapy failure and disease progression in breast cancer patients undergoing neoadjuvant chemotherapy. Oncoimmunology 2017, 7, e1376153. [Google Scholar] [CrossRef] [Green Version]
- Shukuya, T.; Ghai, V.; Amann, J.M.; Okimoto, T.; Shilo, K.; Kim, T.-K.; Wang, K.; Carbone, D.P. Circulating MicroRNAs and Extracellular Vesicle–Containing MicroRNAs as Response Biomarkers of Anti–programmed Cell Death Protein 1 or Programmed Death-Ligand 1 Therapy in NSCLC. J. Thorac. Oncol. 2020, 15, 1773–1781. [Google Scholar] [CrossRef]
- Zhou, B.; Xu, K.; Zheng, X.; Chen, T.; Wang, J.; Song, Y.; Shao, Y.; Zheng, S. Application of exosomes as liquid biopsy in clinical diagnosis. Signal Transduct. Target. Ther. 2020, 5, 144. [Google Scholar] [CrossRef]
- Sun, Z.; Shi, K.; Yang, S.; Liu, J.; Zhou, Q.; Wang, G.; Song, J.; Li, Z.; Zhang, Z.; Yuan, W. Effect of exosomal miRNA on cancer biology and clinical applications. Mol. Cancer 2018, 17, 147. [Google Scholar] [CrossRef] [Green Version]
- Thind, A.; Wilson, C. Exosomal miRNAs as cancer biomarkers and therapeutic targets. J. Extracell. Vesicles 2016, 5, 31292. [Google Scholar] [CrossRef]
- Zhou, E.; Li, Y.; Wu, F.; Guo, M.; Xu, J.; Wang, S.; Tan, Q.; Ma, P.; Song, S.; Jin, Y. Circulating extracellular vesicles are effective biomarkers for predicting response to cancer therapy. Ebiomedicine 2021, 67, 103365. [Google Scholar] [CrossRef]
- Eguchi, A.; Kostallari, E.; Feldstein, A.E.; Shah, V.H. Extracellular vesicles, the liquid biopsy of the future. J. Hepatol. 2019, 70, 1292–1294. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Akbar, A.; Malekian, F.; Baghban, N.; Kodam, S.P.; Ullah, M. Methodologies to Isolate and Purify Clinical Grade Extracellular Vesicles for Medical Applications. Cells 2022, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Danesi, R.; Lo, Y.; Oellerich, M.; Beck, J.; Galbiati, S.; Del Re, M.; Lianidou, E.; Neumaier, M.; van Schaik, R. What do we need to obtain high quality circulating tumor DNA (ctDNA) for routine diagnostic test in oncology?—Considerations on pre-analytical aspects by the IFCC workgroup cfDNA. Clin. Chim. Acta 2021, 520, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, Â.; Ferreira, G.; Seixas, D.; Guimarães-Teixeira, C.; Henrique, R.; Monteiro, F.; Jerónimo, C. Emerging Lab-on-a-Chip Approaches for Liquid Biopsy in Lung Cancer: Status in CTCs and ctDNA Research and Clinical Validation. Cancers 2021, 13, 2101. [Google Scholar] [CrossRef] [PubMed]
- Rupp, B.; Ball, H.; Wuchu, F.; Nagrath, D.; Nagrath, S. Circulating tumor cells in precision medicine: Challenges and opportunities. Trends Pharmacol. Sci. 2022, 43, 378–391. [Google Scholar] [CrossRef]
- Willms, E.; Cabañas, C.; Mäger, I.; Wood, M.J.A.; Vader, P. Extracellular Vesicle Heterogeneity: Subpopulations, Isolation Techniques, and Diverse Functions in Cancer Progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Pertierra, E.; Oliveira-Rodríguez, M.; Matos, M.; Gutiérrez, G.; Moyano, A.; Salvador, M.; Rivas, M.; Blanco-López, M.C. Extracellular Vesicles: Current Analytical Techniques for Detection and Quantification. Biomolecules 2020, 10, 824. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Song, Z.; Hu, X.-Y.; Wang, H.-S. Circulating tumor DNA analysis for tumor diagnosis. Talanta 2021, 228, 122220. [Google Scholar] [CrossRef]
- Kumar, J.; Basak, S.; Kalkal, A.; Packirisamy, G. Recent advances in nanotechnology and microfluidic-based approaches for isolation and detection of circulating tumor cells (CTCs). Nano-Struct. Nano-Objects 2022, 31, 100886. [Google Scholar] [CrossRef]
- Yörüker, E.E.; Holdenrieder, S.; Gezer, U. Potential of circulating nucleosome-associated histone modifications in cancer. Transl. Cancer Res. 2018, 7, S185–S191. [Google Scholar] [CrossRef]
- Gutiérrez-Adrianzén, O.A.; Koutouzov, S.; Mota, R.M.S.; das Chagas Medeiros, M.M.; Bach, J.-F.; de Holanda Campos, H. Diagnostic value of anti-nucleosome antibodies in the assessment of disease activity of systemic lupus erythematosus: A prospective study comparing anti-nucleosome with anti-dsDNA antibodies. J. Rheumatol. 2006, 33, 1538–1544. [Google Scholar]
- Holdenrieder, S.; Stieber, P.; Bodenmüller, H.; Busch, M.; Von Pawel, J.; Schalhorn, A.; Nagel, D.; Seidel, D. Circulating Nucleosomes in Serum. Ann. N. Y. Acad. Sci. 2006, 945, 93–102. [Google Scholar] [CrossRef]
- Widom, J. Toward a Unified Model of Chromatin Folding. Annu. Rev. Biophys. Biophys. Chem. 1989, 18, 365–395. [Google Scholar] [CrossRef]
- Luger, K.; Mäder, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 Å resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef]
- Karantza, V.; Freire, E.; Moudrianakis, E.N. Thermodynamic Studies of the Core Histones: Stability of the Octamer Subunits Is Not Altered by Removal of Their Terminal Domains. Biochemistry 2001, 40, 13114–13123. [Google Scholar] [CrossRef]
- Zhou, K.; Gaullier, G.; Luger, K. Nucleosome structure and dynamics are coming of age. Nat. Struct. Mol. Biol. 2018, 26, 3–13. [Google Scholar] [CrossRef]
- Arrieta, A.; Vondriska, T.M. Nucleosome proteostasis and histone turnover. Front. Mol. Biosci. 2022, 9, 990006. [Google Scholar] [CrossRef]
- Savas, J.N.; Toyama, B.H.; Xu, T.; Yates, J.R., III; Hetzer, M.W. Extremely Long-Lived Nuclear Pore Proteins in the Rat Brain. Science 2012, 335, 942. [Google Scholar] [CrossRef] [Green Version]
- Toyama, B.H.; Savas, J.N.; Park, S.K.; Harris, M.S.; Ingolia, N.T.; Yates, J.R., 3rd; Hetzer, M.W. Identification of Long-Lived Proteins Reveals Exceptional Stability of Essential Cellular Structures. Cell 2013, 154, 971–982. [Google Scholar] [CrossRef] [Green Version]
- Mathieson, T.; Franken, H.; Kosinski, J.; Kurzawa, N.; Zinn, N.; Sweetman, G.; Poeckel, D.; Ratnu, V.S.; Schramm, M.; Becher, I.; et al. Systematic analysis of protein turnover in primary cells. Nat. Commun. 2018, 9, 689. [Google Scholar] [CrossRef] [Green Version]
- Escobar, T.M.; Oksuz, O.; Saldaña-Meyer, R.; Descostes, N.; Bonasio, R.; Reinberg, D. Active and Repressed Chromatin Domains Exhibit Distinct Nucleosome Segregation during DNA Replication. Cell 2019, 179, 953–963.e11. [Google Scholar] [CrossRef]
- Wu, D.; Ingram, A.; Lahti, J.H.; Mazza, B.; Grenet, J.; Kapoor, A.; Liu, L.; Kidd, V.J.; Tang, D. Apoptotic Release of Histones from Nucleosomes. J. Biol. Chem. 2002, 277, 12001–12008. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, X.; Monestier, M.; Esmon, N.L.; Esmon, C.T. Extracellular Histones Are Mediators of Death through TLR2 and TLR4 in Mouse Fatal Liver Injury. J. Immunol. 2011, 187, 2626–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Evankovich, J.; Yan, W.; Nace, G.; Zhang, L.; Ross, M.; Liao, X.; Billiar, T.; Xu, J.; Esmon, C.T.; et al. Endogenous histones function as alarmins in sterile inflammatory liver injury through Toll-like receptor 9 in mice. Hepatology 2011, 54, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Allam, R.; Scherbaum, C.R.; Darisipudi, M.N.; Mulay, S.R.; Hägele, H.; Lichtnekert, J.; Hagemann, J.H.; Rupanagudi, K.V.; Ryu, M.; Schwarzenberger, C.; et al. Histones from Dying Renal Cells Aggravate Kidney InjuryviaTLR2 and TLR4. J. Am. Soc. Nephrol. 2012, 23, 1375–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillrie, M.R.; Lee, K.; Gowda, D.C.; Davis, S.P.; Monestier, M.; Cui, L.; Hien, T.T.; Day, N.P.; Ho, M. Plasmodium falciparum Histones Induce Endothelial Proinflammatory Response and Barrier Dysfunction. Am. J. Pathol. 2012, 180, 1028–1039. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Parseghian, M.H.; Luhrs, K.A. Beyond the walls of the nucleus: The role of histones in cellular signaling and innate immunity. Biochem. Cell Biol. 2006, 84, 589–595. [Google Scholar] [CrossRef]
- Von KöCkritz-Blickwede, M.; Goldmann, O.; Thulin, P.; Heinemann, K.; Norrby-Teglund, A.; Rohde, M.; Medina, E. Phagocytosis-independent antimicrobial activity of mast cells by means of extracellular trap formation. Blood 2008, 111, 3070–3080. [Google Scholar] [CrossRef]
- Schorn, C.; Janko, C.; Latzko, M.; Chaurio, R.; Schett, G.; Herrmann, M. Monosodium urate crystals induce extracellular DNA traps in neutrophils, eosinophils, and basophils but not in mononuclear cells. Front. Immunol. 2012, 3, 277. [Google Scholar] [CrossRef] [Green Version]
- Mohanan, S.; Horibata, S.; McElwee, J.L.; Dannenberg, A.J.; Coonrod, S.A. Identification of Macrophage Extracellular Trap-Like Structures in Mammary Gland Adipose Tissue: A Preliminary Study. Front. Immunol. 2013, 4, 67. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Le Pham, D.; Lee, D.-H.; Lee, S.-H.; Kim, S.-H.; Park, H.-S. Biological function of eosinophil extracellular traps in patients with severe eosinophilic asthma. Exp. Mol. Med. 2018, 50, 1–8. [Google Scholar] [CrossRef]
- Pertiwi, K.R.; De Boer, O.J.; Mackaaij, C.; Pabittei, D.R.; De Winter, R.J.; Li, X.; van der Wal, A. Extracellular traps derived from macrophages, mast cells, eosinophils and neutrophils are generated in a time-dependent manner during atherothrombosis: Etosis in coronary athero-thrombosis. J. Pathol. 2019, 247, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, K.H.; Hawkins, C.L. Role of macrophage extracellular traps in innate immunity and inflammatory disease. Biochem. Soc. Trans. 2022, 50, 21–32. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef]
- Barrero, C.A.; Perez-Leal, O.; Aksoy, M.; Moncada, C.; Ji, R.; Lopez, Y.; Mallilankaraman, K.; Madesh, M.; Criner, G.J.; Kelsen, S.G.; et al. Histone 3.3 Participates in a Self-Sustaining Cascade of Apoptosis that Contributes to the Progression of Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2013, 188, 673–683. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhang, X.; Pelayo, R.; Monestier, M.; Ammollo, C.T.; Semeraro, F.; Taylor, F.B.; Esmon, N.L.; Lupu, F.; Esmon, C.T. Extracellular histones are major mediators of death in sepsis. Nat. Med. 2009, 15, 1318–1321. [Google Scholar] [CrossRef] [Green Version]
- Saffarzadeh, M.; Juenemann, C.; Queisser, M.A.; Lochnit, G.; Barreto, G.; Galuska, S.P.; Lohmeyer, J.; Preissner, K.T. Neutrophil Extracellular Traps Directly Induce Epithelial and Endothelial Cell Death: A Predominant Role of Histones. PLoS ONE 2012, 7, e32366. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Holdenrieder, S.; Dharuman, Y.; Standop, J.; Trimpop, N.; Herzog, M.; Hettwer, K.; Simon, K.; Uhlig, S.; Micallef, J. Novel serum nucleosomics biomarkers for the detection of colorectal cancer. Anticancer Res. 2014, 34, 2357–2362. [Google Scholar]
- Dai, B.; Rasmussen, T.P. Global Epiproteomic Signatures Distinguish Embryonic Stem Cells from Differentiated Cells. Stem Cells 2007, 25, 2567–2574. [Google Scholar] [CrossRef]
- Knutson, S.K.; Warholic, N.M.; Wigle, T.J.; Klaus, C.R.; Allain, C.J.; Raimondi, A.; Scott, M.P.; Chesworth, R.; Moyer, M.P.; Copeland, R.A.; et al. Durable tumor regression in genetically altered malignant rhabdoid tumors by inhibition of methyltransferase EZH2. Proc. Natl. Acad. Sci. USA 2013, 110, 7922–7927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gezer, U.; Yörüker, E.E.; Keskin, M.; Kulle, C.B.; Dharuman, Y.; Holdenrieder, S. Histone Methylation Marks on Circulating Nucleosomes as Novel Blood-Based Biomarker in Colorectal Cancer. Int. J. Mol. Sci. 2015, 16, 29654–29662. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumari, N.; Sharma, U.; Ram, S.; Singh, S.K.; Kakkar, N.; Kaushal, K.; Prasad, R. Reduction in H3K4me patterns due to aberrant expression of methyltransferases and demethylases in renal cell carcinoma: Prognostic and therapeutic implications. Sci. Rep. 2019, 9, 8189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahier, J.-F.; Druez, A.; Faugeras, L.; Martinet, J.-P.; Géhénot, M.; Josseaux, E.; Herzog, M.; Micallef, J.; George, F.; Delos, M.; et al. Circulating nucleosomes as new blood-based biomarkers for detection of colorectal cancer. Clin. Epigenet. 2017, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Bauden, M.P.; Pamart, D.; Ansari, D.; Herzog, M.; Eccleston, M.E.; Micallef, J.; Andersson, B.; Andersson, R. Circulating nucleosomes as epigenetic biomarkers in pancreatic cancer. Clin. Epigenet. 2015, 7, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thålin, C.; Daleskog, M.; Göransson, S.P.; Schatzberg, D.; Lasselin, J.; Laska, A.-C.; Kallner, A.; Helleday, T.; Wallén, H.; Demers, M. Validation of an enzyme-linked immunosorbent assay for the quantification of citrullinated histone H3 as a marker for neutrophil extracellular traps in human plasma. Immunol. Res. 2017, 65, 706–712. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Lin, C.; Leso, A.; Nefedova, Y. Quantification of Citrullinated Histone H3 Bound DNA for Detection of Neutrophil Extracellular Traps. Cancers 2020, 12, 3424. [Google Scholar] [CrossRef]
- Thålin, C.; Lundström, S.; Seignez, C.; Daleskog, M.; Lundström, A.; Henriksson, P.; Helleday, T.; Phillipson, M.; Wallén, H.; Demers, M. Citrullinated histone H3 as a novel prognostic blood marker in patients with advanced cancer. PLoS ONE 2018, 13, e0191231. [Google Scholar] [CrossRef]
- García-Giménez, J.L.; Seco-Cervera, M.; Tollefsbol, T.O.; Romá-Mateo, C.; Peiró-Chova, L.; Lapunzina, P.; Pallardó, F.V. Epigenetic biomarkers: Current strategies and future challenges for their use in the clinical laboratory. Crit. Rev. Clin. Lab. Sci. 2017, 54, 529–550. [Google Scholar] [CrossRef]
- García-Giménez, J.L.; Romá-Mateo, C.; Carbonell, N.; Palacios, L.; Peiró-Chova, L.; García-López, E.; García-Simón, M.; Lahuerta, R.; Gimenez-Garzó, C.; Berenguer-Pascual, E.; et al. A new mass spectrometry-based method for the quantification of histones in plasma from septic shock patients. Sci. Rep. 2017, 7, 10643. [Google Scholar] [CrossRef]
- Ackerveken, P.V.D.; Lobbens, A.; Turatsinze, J.-V.; Solis-Mezarino, V.; Völker-Albert, M.; Imhof, A.; Herzog, M. A novel proteomics approach to epigenetic profiling of circulating nucleosomes. Sci. Rep. 2021, 11, 7256. [Google Scholar] [CrossRef]
- Fedyuk, V.; Erez, N.; Furth, N.; Beresh, O.; Andreishcheva, E.; Shinde, A.; Jones, D.; Bar Zakai, B.; Mavor, Y.; Peretz, T.; et al. Multiplexed, single-molecule, epigenetic analysis of plasma-isolated nucleosomes for cancer diagnostics. Nat. Biotechnol. 2022, 1–10. [Google Scholar] [CrossRef]
- Headland, S.E.; Jones, H.R.; D’Sa, A.S.V.; Perretti, M.; Norling, L. Cutting-Edge Analysis of Extracellular Microparticles using ImageStreamX Imaging Flow Cytometry. Sci. Rep. 2014, 4, 5237. [Google Scholar] [CrossRef] [Green Version]
- Buzova, D.; Maugeri, A.; Liguori, A.; Napodano, C.; Re, O.L.; Oben, J.; Alisi, A.; Gasbarrini, A.; Grieco, A.; Cerveny, J.; et al. Circulating histone signature of human lean metabolic-associated fatty liver disease (MAFLD). Clin. Epigenet. 2020, 12, 126. [Google Scholar] [CrossRef]
- Buzova, D.; Braghini, M.R.; Bianco, S.D.; Re, O.L.; Raffaele, M.; Frohlich, J.; Kisheva, A.; Crudele, A.; Mosca, A.; Sartorelli, M.R.; et al. Profiling of cell-free DNA methylation and histone signatures in pediatric NAFLD: A pilot study. Hepatol. Commun. 2022, 6, 3311–3323. [Google Scholar] [CrossRef]
- Ogle, L.F.; Orr, J.G.; Willoughby, C.E.; Hutton, C.; McPherson, S.; Plummer, R.; Boddy, A.V.; Curtin, N.J.; Jamieson, D.; Reeves, H.L. Imagestream detection and characterisation of circulating tumour cells—A liquid biopsy for hepatocellular carcinoma? J. Hepatol. 2016, 65, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Diamant, A.; Chatterjee, A.; Vallières, M.; Shenouda, G.; Seuntjens, J. Deep learning in head & neck cancer outcome prediction. Sci. Rep. 2019, 9, 2764. [Google Scholar]
- Park, S.; Ock, C.-Y.; Kim, H.; Pereira, S.; Park, S.; Ma, M.; Choi, S.; Kim, S.; Shin, S.; Aum, B.J.; et al. Artificial Intelligence–Powered Spatial Analysis of Tumor-Infiltrating Lymphocytes as Complementary Biomarker for Immune Checkpoint Inhibition in Non–Small-Cell Lung Cancer. J. Clin. Oncol. 2022, 40, 1916–1928. [Google Scholar] [CrossRef]
- Huang, T.Y.; Piunti, A.; Lulla, R.R.; Qi, J.; Horbinski, C.M.; Tomita, T.; James, C.D.; Shilatifard, A.; Saratsis, A.M. Detection of Histone H3 mutations in cerebrospinal fluid-derived tumor DNA from children with diffuse midline glioma. Acta Neuropathol. Commun. 2017, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Bonner, E.R.; Wierzbicki, K.; Panditharatna, E.; Huang, T.; Lulla, R.; Mueller, S.; Koschmann, C.; Nazarian, J.; Saratsis, A.M. Standardization of the liquid biopsy for pediatric diffuse midline glioma using ddPCR. Sci. Rep. 2021, 11, 5098. [Google Scholar] [CrossRef]
- Wang, Y.; Li, M.; Stadler, S.C.; Correll, S.; Li, P.; Wang, D.; Hayama, R.; Leonelli, L.; Han, H.; Grigoryev, S.A.; et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J. Cell Biol. 2009, 184, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, D.; Zhang, Y.; Wang, S. Histone citrullination: A new target for tumors. Mol. Cancer 2021, 20, 90. [Google Scholar] [CrossRef] [PubMed]
- Hewish, D.R.; Burgoyne, L.A. Chromatin sub-structure. The digestion of chromatin DNA at regularly spaced sites by a nuclear deoxyribonuclease. Biochem. Biophys. Res. Commun. 1973, 52, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.C.; Blumenfeld, Y.J.; Chitkara, U.; Hudgins, L.; Quake, S.R. Noninvasive diagnosis of fetal aneuploidy by shotgun sequencing DNA from maternal blood. Proc. Natl. Acad. Sci. USA 2008, 105, 16266–16271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Shollenberger, L.; Conwell, C.C.; Yuan, X.; Huang, L. Mechanism of naked DNA clearance after intravenous injection. J. Gene Med. 2007, 9, 613–619. [Google Scholar] [CrossRef]
- Chan, R.W.; Serpas, L.; Ni, M.; Volpi, S.; Hiraki, L.T.; Tam, L.-S.; Rashidfarrokhi, A.; Wong, P.C.; Tam, L.H.; Wang, Y.; et al. Plasma DNA Profile Associated with DNASE1L3 Gene Mutations: Clinical Observations, Relationships to Nuclease Substrate Preference, and In Vivo Correction. Am. J. Hum. Genet. 2020, 107, 882–894. [Google Scholar] [CrossRef]
- Chandrasekaran, A.R. Nuclease resistance of DNA nanostructures. Nat. Rev. Chem. 2021, 5, 225–239. [Google Scholar] [CrossRef]
- Lo, Y.M.D.; Chan, K.C.A.; Sun, H.; Chen, E.Z.; Jiang, P.; Lun, F.M.F.; Zheng, Y.W.; Leung, T.Y.; Lau, T.K.; Cantor, C.R.; et al. Maternal Plasma DNA Sequencing Reveals the Genome-Wide Genetic and Mutational Profile of the Fetus. Sci. Transl. Med. 2010, 2, 61ra91. [Google Scholar] [CrossRef]
- Snyder, M.W.; Kircher, M.; Hill, A.J.; Daza, R.M.; Shendure, J. Cell-free DNA Comprises an In Vivo Nucleosome Footprint that Informs Its Tissues-Of-Origin. Cell 2016, 164, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Aranda, P.; Agirre, X.; Ballestar, E.; Andreu, E.J.; Roman-Gomez, J.; Prieto, I.; Martin-Subero, J.I.; Cigudosa, J.C.; Siebert, R.; Esteller, M.; et al. Epigenetic Signatures Associated with Different Levels of Differentiation Potential in Human Stem Cells. PLoS ONE 2009, 4, e7809. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho, D.D.; Sharma, S.; You, J.S.; Su, S.-F.; Taberlay, P.C.; Kelly, T.K.; Yang, X.; Liang, G.; Jones, P.A. DNA Methylation Screening Identifies Driver Epigenetic Events of Cancer Cell Survival. Cancer Cell 2012, 21, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Roy, R.; Ramamoorthy, S.; Shapiro, B.D.; Kaileh, M.; Hernandez, D.; Sarantopoulou, D.; Arepalli, S.; Boller, S.; Singh, A.; Bektas, A.; et al. DNA methylation signatures reveal that distinct combinations of transcription factors specify human immune cell epigenetic identity. Immunity 2021, 54, 2465–2480.e5. [Google Scholar] [CrossRef]
- Wu, S.J.; Furlan, S.N.; Mihalas, A.B.; Kaya-Okur, H.S.; Feroze, A.H.; Emerson, S.N.; Zheng, Y.; Carson, K.; Cimino, P.J.; Keene, C.D.; et al. Single-cell CUT&Tag analysis of chromatin modifications in differentiation and tumor progression. Nat. Biotechnol. 2021, 39, 819–824. [Google Scholar]
- Lui, Y.Y.; Chik, K.-W.; Chiu, R.W.; Ho, C.-Y.; Lam, C.W.; Lo, Y.D. Predominant Hematopoietic Origin of Cell-free DNA in Plasma and Serum after Sex-mismatched Bone Marrow Transplantation. Clin. Chem. 2002, 48, 421–427. [Google Scholar] [CrossRef]
- Kim, H. Circulating Histone Is Elevated in Acute Leukemia and Protects Leukemic Cell Death through Enhancement of Leukemic Cell Adhesion to Endothelium. Blood 2016, 128, 4914. [Google Scholar] [CrossRef]
- Yoo, H.J.; Lee, J.-S.; Kim, J.-E.; Gu, J.; Koh, Y.; Kim, I.; Kim, H.K. Extracellular Histone Released from Leukemic Cells Increases Their Adhesion to Endothelium and Protects them from Spontaneous and Chemotherapy-Induced Leukemic Cell Death. PLoS ONE 2016, 11, e0163982. [Google Scholar] [CrossRef] [Green Version]
- Class, R.; Lindman, S.; Fassbender, C.; Leinenbach, H.-P.; Rawer, S.; Emrich, J.G.; Brady, L.W.; Zeppezauer, M. Histone H1 Suppresses Tumor Growth of Leukemia Cells In Vitro, Ex Vivo and in an Animal Model Suggesting Extracellular Functions of Histones. Am. J. Clin. Oncol. 1996, 19, 522–531. [Google Scholar] [CrossRef]
- Holdenrieder, S.; Stieber, P.; Busch, M.; Fertig, G.; Schalhorn, A.; Schmeller, N.; Untch, M.; Seidel, D. Nucleosomes in serum of patients with benign and malignant diseases. Int. J. Cancer 2001, 95, 114–120. [Google Scholar] [CrossRef]
- Terrell, J.; Josseaux, E.; Latora, H.; Rommelaere, G.; Hardat, N.; Herzog, M.; Micallef, J.; Eccleston, M. Circulating nucleosomes in hematological malignancy. J. Clin. Oncol. 2020, 38 (Suppl. S15), e20078. [Google Scholar] [CrossRef]
- Nie, M.; Yang, L.; Bi, X.; Wang, Y.; Sun, P.; Yang, H.; Liu, P.; Li, Z.; Xia, Y.; Jiang, W. Neutrophil Extracellular Traps Induced by IL8 Promote Diffuse Large B-cell Lymphoma Progression via the TLR9 Signaling. Clin. Cancer Res. 2019, 25, 1867–1879. [Google Scholar] [CrossRef]
- Mueller, S.; Holdenrieder, S.; Stieber, P.; Haferlach, T.; Schalhorn, A.; Braess, J.; Nagel, D.; Seidel, D. Early prediction of therapy response in patients with acute myeloid leukemia by nucleosomal DNA fragments. BMC Cancer 2006, 6, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gezer, U.; Yaman, F.; Erten, N.; Dalay, N. Frequent copresence of methylated DNA and fragmented nucleosomal DNA in plasma of lymphoma patients. Clin. Chim. Acta 2003, 335, 89–94. [Google Scholar]
- Deligezer, U.; Erten, N.; Akisik, E.E.; Dalay, N. Circulating fragmented nucleosomal DNA and caspase-3 mRNA in patients with lymphoma and myeloma. Exp. Mol. Pathol. 2006, 80, 72–76. [Google Scholar] [CrossRef]
- Kuroi, K.; Tanaka, C.; Toi, M. Plasma nucleosome levels in node-negative breast cancer patients. Breast Cancer 1999, 6, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Kremer, A.; Holdenrieder, S.; Stieber, P.; Wilkowski, R.; Nagel, D.; Seidel, D. Nucleosomes in Colorectal Cancer Patients during Radiochemotherapy. Tumor Biol. 2006, 27, 235–242. [Google Scholar] [CrossRef]
- Stoetzer, O.J.; Fersching, D.M.I.; Salat, C.; Steinkohl, O.; Gabka, C.J.; Hamann, U.; Braun, M.; Feller, A.M.; Heinemann, V.; Siegele, B.; et al. Prediction of response to neoadjuvant chemothera-py in breast cancer patients by circulating apoptotic biomarkers nucleosomes, DNAse, cytokeratin-18 fragments and survivin. Cancer Lett. 2013, 336, 140–148. [Google Scholar] [CrossRef]
- Roth, C.; Pantel, K.; Müller, V.; Rack, B.; Kasimir-Bauer, S.; Janni, W.; Schwarzenbach, H. Apoptosis-related deregulation of proteolytic activities and high serum levels of circulating nucleosomes and DNA in blood correlate with breast cancer progression. BMC Cancer 2011, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.; Cario, C.L.; Leong, L.; Lopez, K.; Márquez, C.P.; Chu, C.; Li, P.S.; Oropeza, E.; Tenggara, I.; Cowan, J.; et al. Cell-free DNA concentration and fragment size as a biomarker for prostate cancer. Sci. Rep. 2021, 11, 5040. [Google Scholar] [CrossRef]
- Rasmussen, L.; Christensen, I.J.; Herzog, M.; Micallef, J.; Nielsen, H.J.; For the Danish Collaborative Group on Early Detection of Colorectal Cancer. Circulating cell-free nucleosomes as biomarkers for early detection of colorectal cancer. Oncotarget 2017, 9, 10247–10258. [Google Scholar] [CrossRef] [Green Version]
- Leszinski, G.; Gezer, U.; Siegele, B.; Stoetzer, O.; Holdenrieder, S. Relevance of histone marks H3K9me3 and H4K20me3 in cancer. Anticancer Res. 2012, 32, 2199–2205. [Google Scholar]
- Vanderstichele, A.; Busschaert, P.; Landolfo, C.; Olbrecht, S.; Coosemans, A.; Froyman, W.; Loverix, L.; Concin, N.; Braicu, E.I.; Wimberger, P.; et al. Nucleosome footprinting in plasma cell-free DNA for the pre-surgical diagnosis of ovarian cancer. NPJ Genom. Med. 2022, 7, 30. [Google Scholar] [CrossRef]
- Cristiano, S.; Leal, A.; Phallen, J.; Fiksel, J.; Adleff, V.; Bruhm, D.C.; Jensen, S.Ø.; Medina, J.E.; Hruban, C.; White, J.R.; et al. Genome-wide cell-free DNA fragmentation in patients with cancer. Nature 2019, 570, 385–389. [Google Scholar] [CrossRef]
- Salani, F.; Latarani, M.; Casadei-Gardini, A.; Gangadharannambiar, P.; Fornaro, L.; Vivaldi, C.; Pecora, I.; Massa, V.; Marisi, G.; Canale, M.; et al. Predictive significance of circulating histones in hepatocellular carcinoma patients treated with sorafenib. Epigenomics 2022, 14, 507–517. [Google Scholar] [CrossRef]
- Deligezer, U.; Yaman, F.; Darendeliler, E.; Dizdar, Y.; Holdenrieder, S.; Kovancilar, M.; Dalay, N. Post-treatment circulating plasma BMP6 mRNA and H3K27 methylation levels discriminate metastatic prostate cancer from localized disease. Clin. Chim. Acta 2010, 411, 1452–1456. [Google Scholar] [CrossRef]
- Kuroi, K.; Tanaka, C.; Toi, M. Clinical significance of plasma nucleosome levels in cancer patients. Int. J. Oncol. 2001, 19, 143–148. [Google Scholar] [CrossRef]
- Trejo-Becerril, C.; Pérez-Cárdenas, E.; Treviño-Cuevas, H.; Taja-Chayeb, L.; García-López, P.; Segura-Pacheco, B.; Chávez-Blanco, A.; Lizano-Soberon, M.; González-Fierro, A.; Mariscal, I.; et al. Circulating nucleosomes and response to chemotherapy: An in vitro, in vivo and clinical study on cervical cancer patients: Serum Nucleosomes and Chemotherapy Response. Int. J. Cancer 2003, 104, 663–668. [Google Scholar] [CrossRef]
- Holdenrieder, S.; Stieber, P. Therapy Control in Oncology by Circulating Nucleosomes. Ann. N. Y. Acad. Sci. 2004, 1022, 211–216. [Google Scholar] [CrossRef]
- Holdenrieder, S.; Stieber, P.; von Pawel, J.; Raith, H.; Nagel, D.; Feldmann, K.; Seidel, D. Circulating Nucleosomes Predict the Response to Chemotherapy in Patients with Advanced Non–Small Cell Lung Cancer. Clin. Cancer Res. 2004, 10, 5981–5987. [Google Scholar] [CrossRef] [Green Version]
- Kremer, A.; Wilkowski, R.; Holdenrieder, S.; Nagel, D.; Stieber, P.; Seidel, D. Nucleosomes in Pancreatic Cancer Patients during Radiochemotherapy. Tumor Biol. 2005, 26, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Trejo-Becerril, C.; Oñate-Ocaña, L.F.; Taja-Chayeb, L.; Vanoye-Carlo, A.; Cetina, L.; Duenas-Gonzalez, A. Serum nucleosomes during neoadjuvant chemotherapy in patients with cervical cancer. Predictive and prognostic significance. BMC Cancer 2005, 5, 65. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Guleria, R.; Singh, V.; Bharti, A.C.; Mohan, A.; Das, B.C. Plasma Nucleosome Levels Might Predict Response to Therapy in Patients with Advanced Non–Small-Cell Lung Cancer. Clin. Lung Cancer 2010, 11, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Gu, T.; Ge, Y.; Song, Y.; Fu, Z.; Zhang, Y.; Wang, G.; Shao, S.; Wen, T. Hepatic radiofrequency ablation causes an increase of circulating histones in patients with hepatocellular carcinoma. Scand. J. Clin. Lab. Investig. 2015, 75, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Doebley, A.-L.; Ko, M.; Liao, H.; Cruikshank, A.E.; Santos, K.; Kikawa, C.; Hiatt, J.B.; Patton, R.D.; De Sarkar, N.; Collier, K.A.; et al. A framework for clinical cancer subtyping from nucleosome profiling of cell-free DNA. Nat. Commun. 2022, 13, 7475. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.; Herzog, M.; Rømer, E.; Micallef, J.; Bulut, O.; Wilhelmsen, M.; Christensen, I.J.; Nielsen, H.J. Pre-analytical variables of circulating cell-free nucleosomes containing 5-methylcytosine DNA or histone modification H3K9Me3. Scand. J. Clin. Lab. Investig. 2016, 76, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Hu, P.; Donskov, F.; Wang, G.; Liu, Q.; Du, J. Tumor-Associated Neutrophils as a New Prognostic Factor in Cancer: A Systematic Review and Meta-Analysis. PLoS ONE 2014, 9, e98259. [Google Scholar] [CrossRef] [Green Version]
- Uribe-Querol, E.; Rosales, C. Neutrophils in Cancer: Two Sides of the Same Coin. J. Immunol. Res. 2015, 2015, 983698. [Google Scholar] [CrossRef]
- Coffelt, S.B.; Wellenstein, M.D.; De Visser, K.E. Neutrophils in cancer:Neutral no more. Nat. Rev. Cancer 2016, 16, 431–446. [Google Scholar] [CrossRef] [Green Version]
- Jaillon, S.; Ponzetta, A.; Di Mitri, D.; Santoni, A.; Bonecchi, R.; Mantovani, A. Neutrophil diversity and plasticity in tumour progression and therapy. Nat. Rev. Cancer 2020, 20, 485–503. [Google Scholar] [CrossRef]
- Szczerba, B.M.; Castro-Giner, F.; Vetter, M.; Krol, I.; Gkountela, S.; Landin, J.; Scheidmann, M.C.; Donato, C.; Scherrer, R.; Singer, J.; et al. Neutrophils escort circulating tumour cells to enable cell cycle progression. Nature 2019, 566, 553–557. [Google Scholar] [CrossRef]
- Spicer, J.D.; McDonald, B.; Cools-Lartigue, J.J.; Chow, S.C.; Giannias, B.; Kubes, P.; Ferri, L.E. Neutrophils Promote Liver Metastasis via Mac-1–Mediated Interactions with Circulating Tumor Cells. Cancer Res. 2012, 72, 3919–3927. [Google Scholar] [CrossRef] [Green Version]
- Cools-Lartigue, J.; Spicer, J.; McDonald, B.; Gowing, S.; Chow, S.; Giannias, B.; Bourdeau, F.; Kubes, P.; Ferri, L. Neutrophil extracellular traps sequester circulating tumor cells and promote metastasis. J. Clin. Investig. 2013, 123, 3446–3458. [Google Scholar] [CrossRef]
- Meara, C.H.O.; Coupland, L.A.; Kordbacheh, F.; Quah, B.J.C.; Chang, C.-W.; Davis, D.A.S.; Bezos, A.; Browne, A.M.; Freeman, C.; Hammill, D.J.; et al. Neutralizing the pathological effects of extracellular histones with small polyanions. Nat. Commun. 2020, 11, 6408. [Google Scholar] [CrossRef]
- Wilson, A.S.; Randall, K.L.; Pettitt, J.A.; Ellyard, J.I.; Blumenthal, A.; Enders, A.; Quah, B.J.; Bopp, T.; Parish, C.R.; Brüstle, A. Neutrophil extracellular traps and their histones promote Th17 cell differentiation directly via TLR2. Nat. Commun. 2022, 13, 528. [Google Scholar] [CrossRef]
- Kesselring, R.; Thiel, A.; Pries, R.; Trenkle, T.; Wollenberg, B. Human Th17 cells can be induced through head and neck cancer and have a functional impact on HNSCC development. Br. J. Cancer 2010, 103, 1245–1254. [Google Scholar] [CrossRef] [Green Version]
- Novitskiy, S.V.; Pickup, M.W.; Gorska, A.E.; Owens, P.; Chytil, A.; Aakre, M.; Wu, H.; Shyr, Y.; Moses, H.L. TGF-β Receptor II Loss Promotes Mammary Carcinoma Progression by Th17-Dependent Mechanisms. Cancer Discov. 2011, 1, 430–441. [Google Scholar] [CrossRef] [Green Version]
- Hajimoradi, M.; Rezalotfi, A.; Esmaeilnejad-Ahranjani, P.; Hassan, Z.M.; Ebrahimi, M. STAT3 inactivation suppresses stemness properties in gastric cancer stem cells and promotes Th17 in Treg/Th17 balance. Int. Immunopharmacol. 2022, 111, 109048. [Google Scholar] [CrossRef]
- Ventures, M. Liquid Biopsy Market Growth Drivers & Opportunities. MarketsandMarkets. Available online: https://www.marketsandmarkets.com/Market-Reports/liquid-biopsy-market-13966350.html (accessed on 25 December 2022).
- Epigenetics Market Growth Drivers & Opportunities. MarketsandMarkets. Available online: https://www.marketsandmarkets.com/Market-Reports/epigenetics-technologies-market-896.html (accessed on 25 December 2022).

| Type of Liquid Biopsy | Stability | Detection Method | Specificity | Sensitivity |
|---|---|---|---|---|
| Circulating tumor DNA | * | PCR, sequencing (various types); Nanomaterials-based ctDNA analysis; Electrochemical ctDNA detection; Surface-enhanced Raman scattering (SERS) | ** | **** |
| Circulating tumor cells | **** | Flow-cytometry; nano/micro magnetic particles; Microfluidics; mechanical filtration; Hydrodynamics; electrokinetics; acoustophoresis | *** | ** |
| Tumor extracellular vesicles | ** | Centrifugation, density gradient; chromatography; fractionation immunoaffinity; lipidomics/mass spectrometry; flow cytometry; microfluidics; lateral-flow immunoassay (LFIA); nanoparticle tracking analysis (NTA) | *** | *** |
| Circulating histones | *** | ELISA, proteomics, Flow-cytometry | *** | **** |
| Detection Method | Sensitivity | Multiplex | Bias | Easy to Use | Easy to Access | Expensiveness | Speed |
|---|---|---|---|---|---|---|---|
| ELISA | **** | ** | **** | **** | *** | **/*** | ** |
| Proteomics | **** | **** | * | * | * | * | ** |
| ImageStream | **** | *** | *** | **/*** | * | **/*** | **** |
| Malignancy | Target for Detection | Detection Method | Level | Suggested Function | Reference |
|---|---|---|---|---|---|
| Leukemia | Histone—dsDNA complex | ELISA | High | Disease progression Chemotherapy resistance | [115,116] |
| Leukemia | NA | NA | Injection of 200 µg/mL H1 histone | Cytotoxicity of tumor cells | [117] |
| Burkitt’s lymphoma | NA | NA | Injection of 200 µg/mL H1 histone | Inhibits tumor growth | [117] |
| Lymphoma | Anti-histone, anti-DNA-antibodies | ELISA | High | Detection | [118] |
| Non-Hodgkin lymphoma (NHL) | Nu.Q-H3.1 | ELISA | High | Detection | [119] |
| Acute myeloid leukemia (AML) | Nu.Q-H3.1 | ELISA | High | Detection | [119] |
| Acute lymphocytic leukemia (ALL) | Nu.Q-H3.1 | ELISA | High | Detection | [119] |
| AML | Anti-histone, anti-DNA-antibodies | ELISA | Initial rise + Decrease following treatment | Chemotherapy response | [121] |
| Lymphoma | UV intensities of individual DNA fragments | Detection on 2% agarose gel | High | Detection of disease progression | [122] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsoneva, D.K.; Ivanov, M.N.; Conev, N.V.; Manev, R.; Stoyanov, D.S.; Vinciguerra, M. Circulating Histones to Detect and Monitor the Progression of Cancer. Int. J. Mol. Sci. 2023, 24, 942. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24020942
Tsoneva DK, Ivanov MN, Conev NV, Manev R, Stoyanov DS, Vinciguerra M. Circulating Histones to Detect and Monitor the Progression of Cancer. International Journal of Molecular Sciences. 2023; 24(2):942. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24020942
Chicago/Turabian StyleTsoneva, Desislava K., Martin N. Ivanov, Nikolay Vladimirov Conev, Rostislav Manev, Dragomir Svetozarov Stoyanov, and Manlio Vinciguerra. 2023. "Circulating Histones to Detect and Monitor the Progression of Cancer" International Journal of Molecular Sciences 24, no. 2: 942. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24020942