
Prevention of Chemotherapy-Induced Peripheral Neuropathy by Inhibiting C-X-C Motif Chemokine Receptor 2
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
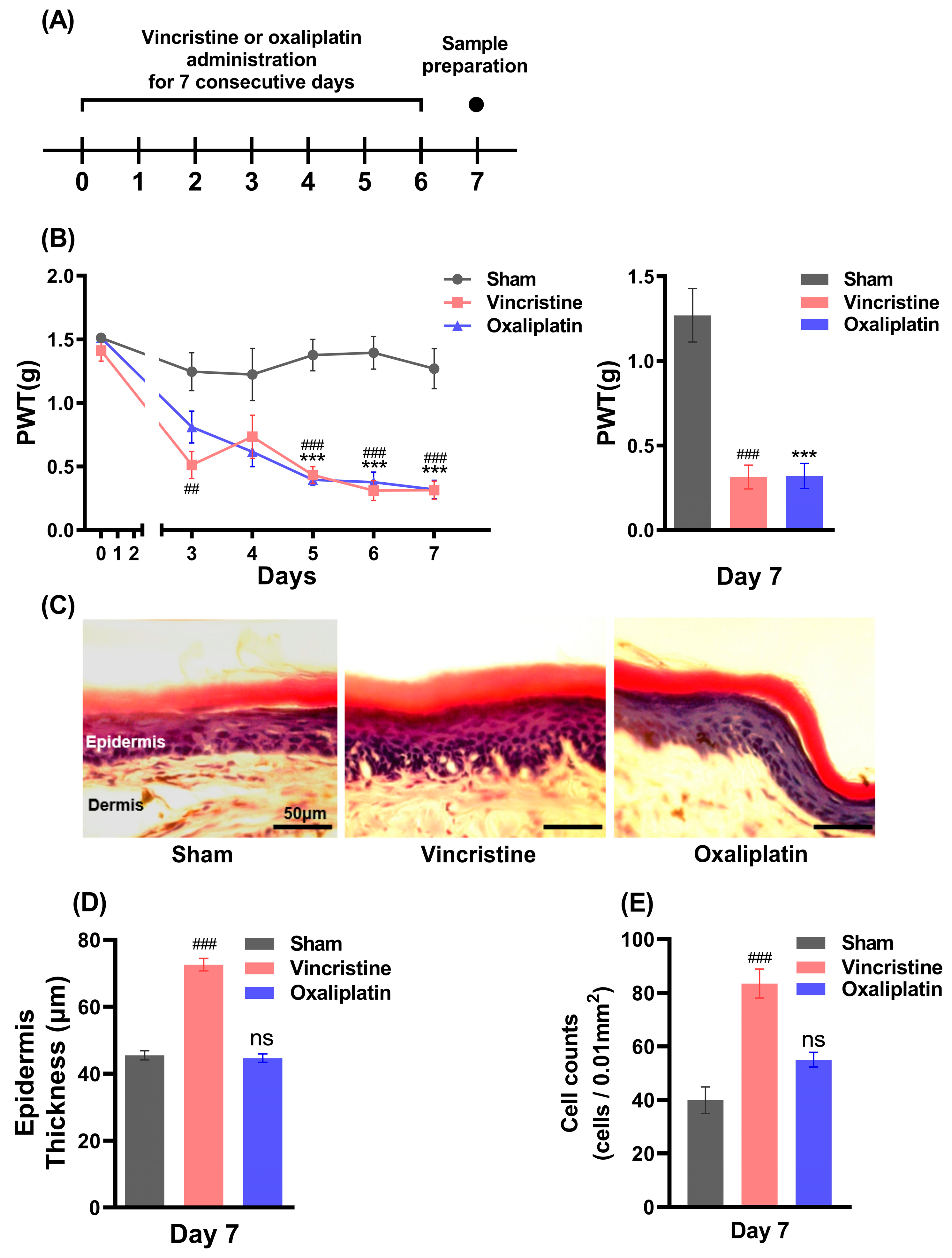
2.1. Vincristine Induces Peripheral Inflammation along with CIPN
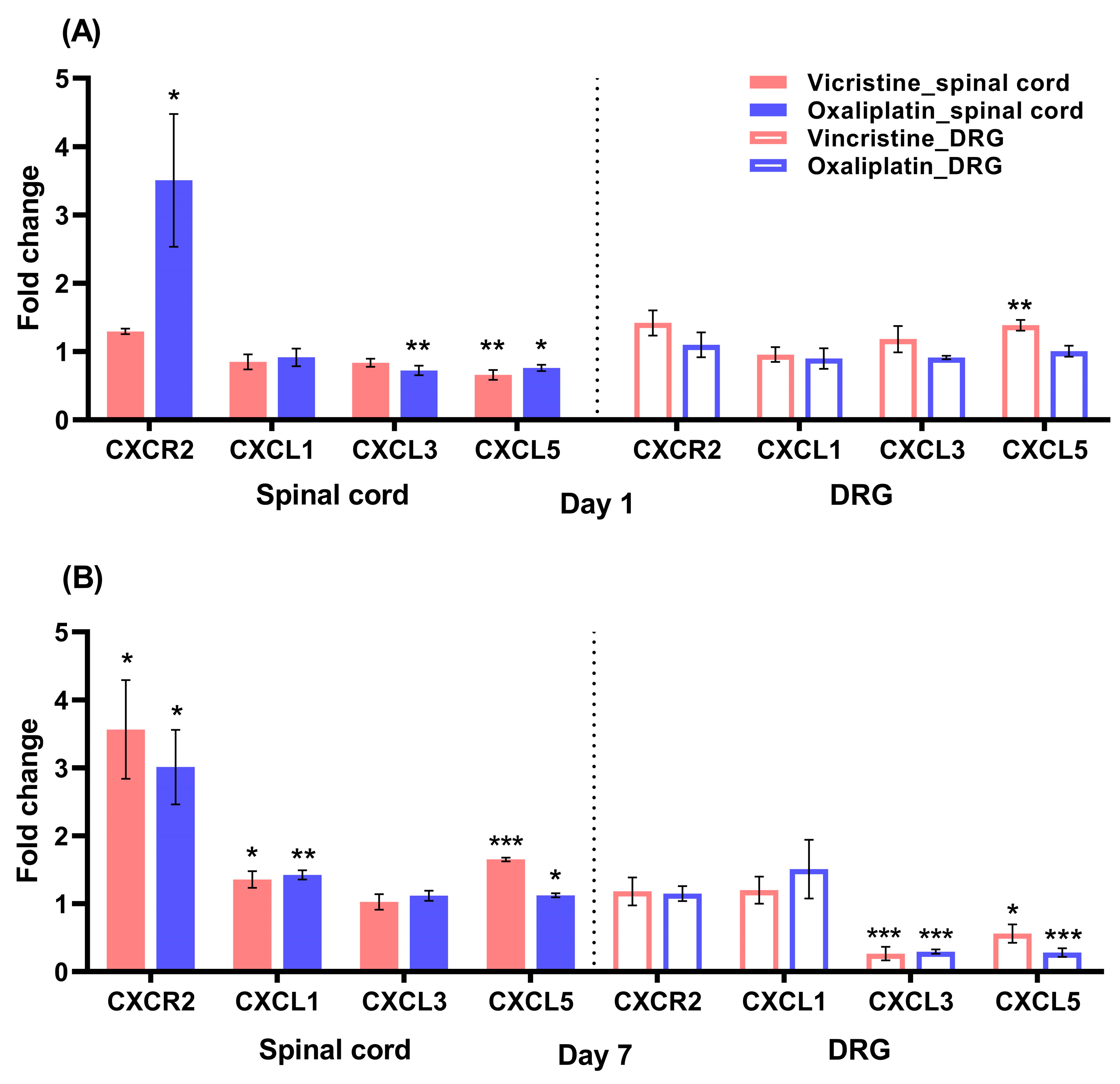
2.2. Increases in CXCR2 mRNA in the Lumbar Spinal Cord in the Early Stage of Oxaliplatin-Induced Neuropathy
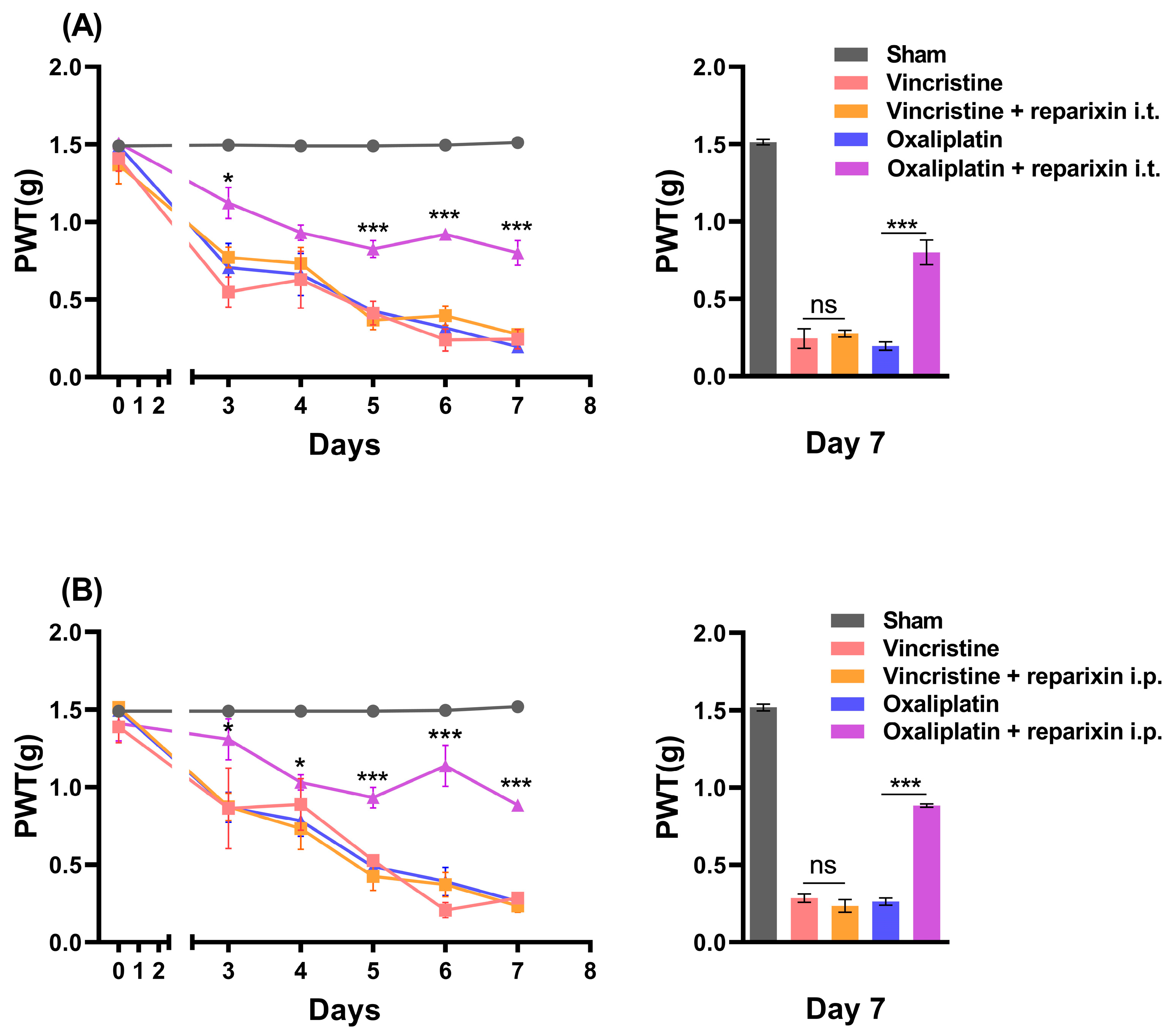
2.3. Inhibition of CXCR2 with Reparixin Prevents Oxaliplatin-Induced CIPN
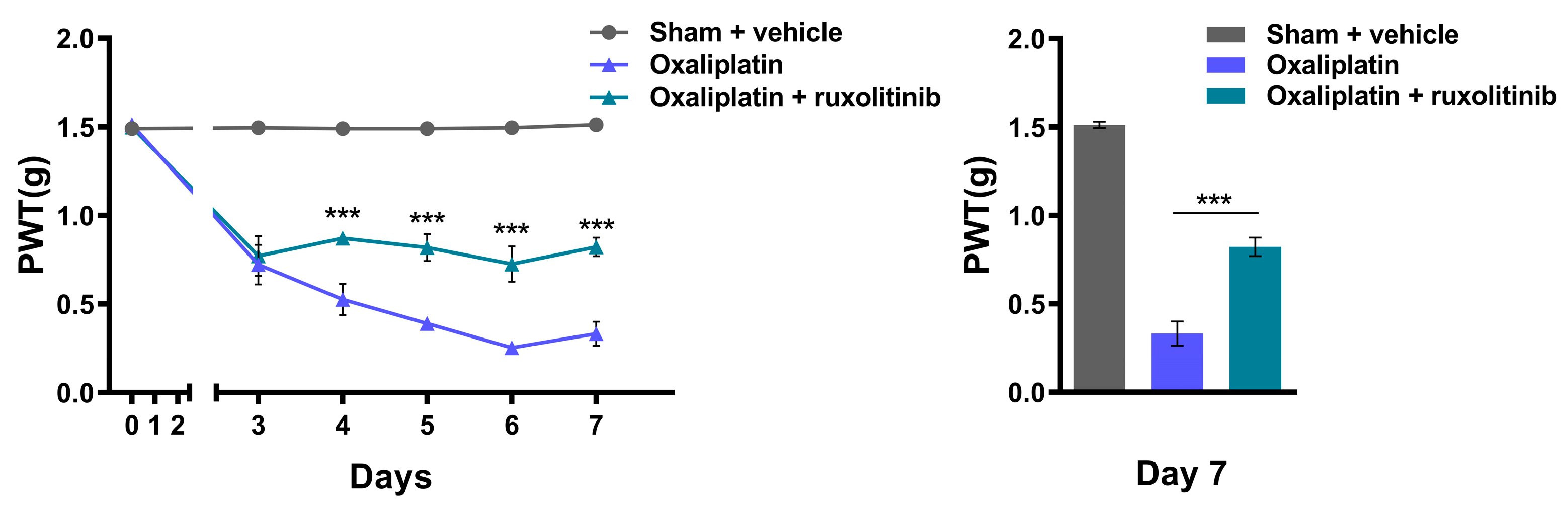
2.4. Inhibition of JAK2 with Ruxolitinib Prevents Oxaliplatin-Induced CIPN
3. Discussion
3.1. The Peripheral Neuropathy and Epidermal Change
3.2. The Changes in CXCR2 mRNA Expression in the Spinal Cord
3.3. The Role of Spinal CXCR2 in the Development of Oxaliplatin-Induced Neuropathy
3.4. The Clinical Implication for the Prevention of Oxaliplatin-Induced CIPN
4. Materials and Methods
4.1. Animals
4.2. Reagents
4.3. Drug Treatment
4.4. Von Frey Test
4.5. Real-Time PCR
- GAPDH forward, 5′-AGGTCGGTGTGAACGGATTTG-3′;
- GAPDH reverse, 5′-TGTAGACCATGTAGTTGAGGTCA-3′;
- CXCR2 forward, 5′-TGAGGGTCGTACTGCGTATC-3′;
- CXCR2 reverse, 5′-AGTGTGAACCCGTAGCAGAA-3′;
- CXCL1 forward, 5′-TCCAGAGCTTGAAGGTGTTGCC-3′;
- CXCL1 reverse, 5′-AACCAAGGGAGCTTCAGGGTCA-3′;
- CXCL3 forward, 5′-TGAGACCATCCAGAGCTTGACG-3′;
- CXCL3 reverse, 5′-CCTTGGGGGTTGAGGCAAACTT-3′;
- CXCL5 forward, 5′-CCGCTGGCATTTCTGTTGCTGT-3′;
- CXCL5 reverse, 5′-CAGGGATCACCTCCAAATTAGCG-3′.
4.6. Staining
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maihöfner, C.; Diel, I.; Tesch, H.; Quandel, T.; Baron, R. Chemotherapy-induced peripheral neuropathy (CIPN): Current therapies and topical treatment option with high-concentration capsaicin. Support. Care Cancer 2021, 29, 4223–4238. [Google Scholar] [CrossRef] [PubMed]
- Boyette-Davis, J.A.; Hou, S.; Abdi, S.; Dougherty, P.M. An updated understanding of the mechanisms involved in chemotherapy-induced neuropathy. Pain Manag. 2018, 8, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Park, S.B.; Goldstein, D.; Krishnan, A.V.; Lin, C.S.Y.; Friedlander, M.L.; Cassidy, J.; Koltzenburg, M.; Kiernan, M.C. Chemotherapy-induced peripheral neurotoxicity: A critical analysis. CA Cancer J. Clin. 2013, 63, 419–437. [Google Scholar] [CrossRef] [PubMed]
- Boyette-Davis, J.A.; Cata, J.P.; Driver, L.C.; Novy, D.M.; Bruel, B.M.; Mooring, D.L.; Wendelschafer-Crabb, G.; Kennedy, W.R.; Dougherty, P.M. Persistent chemoneuropathy in patients receiving the plant alkaloids paclitaxel and vincristine. Cancer Chemother. Pharmacol. 2013, 71, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Windebank, A.J.; Grisold, W. Chemotherapy-induced neuropathy. J. Peripher. Nerv. Syst. 2008, 13, 27–46. [Google Scholar] [CrossRef]
- Sałat, K. Chemotherapy-induced peripheral neuropathy: Part 1—Current state of knowledge and perspectives for pharmacotherapy. Pharmacol. Rep. 2020, 72, 486–507. [Google Scholar] [CrossRef]
- Starobova, H.; Mueller, A.; Deuis, J.R.; Carter, D.A.; Vetter, I. Inflammatory and neuropathic gene expression signatures of chemotherapy-induced neuropathy induced by vincristine, cisplatin, and oxaliplatin in C57BL/6J mice. J. Pain 2020, 21, 182–194. [Google Scholar] [CrossRef]
- Cascella, M.; Muzio, M.R. Pharmacological Approaches and Natural Products for Prevention of Chemotherapy-Induced Peripheral Neuropathy-A Review. Assessment 2017, 12, 13. [Google Scholar] [CrossRef] [Green Version]
- Oun, R.; Moussa, Y.E.; Wheate, N.J. The side effects of platinum-based chemotherapy drugs: A review for chemists. Dalton Trans. 2018, 47, 6645–6653. [Google Scholar] [CrossRef]
- André, T.; Boni, C.; Mounedji-Boudiaf, L.; Navarro, M.; Tabernero, J.; Hickish, T.; Topham, C.; Zaninelli, M.; Clingan, P.; Bridgewater, J. Oxaliplatin, fluorouracil, and leucovorin as adjuvant treatment for colon cancer. N. Engl. J. Med. 2004, 350, 2343–2351. [Google Scholar] [CrossRef]
- Tesniere, A.; Schlemmer, F.; Boige, V.; Kepp, O.; Martins, I.; Ghiringhelli, F.; Aymeric, L.; Michaud, M.; Apetoh, L.; Barault, L. Immunogenic death of colon cancer cells treated with oxaliplatin. Oncogene 2010, 29, 482–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khushalani, N.I.; Leichman, C.G.; Proulx, G.; Nava, H.; Bodnar, L.; Klippenstein, D.; Litwin, A.; Smith, J.; Nava, E.; Pendyala, L. Oxaliplatin in combination with protracted-infusion fluorouracil and radiation: Report of a clinical trial for patients with esophageal cancer. J. Clin. Oncol. 2002, 20, 2844–2850. [Google Scholar] [CrossRef] [PubMed]
- Rödel, C.; Grabenbauer, G.G.; Papadopoulos, T.; Hohenberger, W.; Schmoll, H.-J.; Sauer, R. Phase I/II trial of capecitabine, oxaliplatin, and radiation for rectal cancer. J. Clin. Oncol. 2003, 21, 3098–3104. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.V.; Goldstein, D.; Friedlander, M.; Kiernan, M.C. Oxaliplatin-induced neurotoxicity and the development of neuropathy. Muscle Nerve Off. J. Am. Assoc. Electrodiagn. Med. 2005, 32, 51–60. [Google Scholar] [CrossRef]
- Carbone, P.P.; Bono, V.; Frei, E., III; Brindley, C.O. Clinical studies with vincristine. Blood 1963, 21, 640–647. [Google Scholar] [CrossRef]
- Rosenthal, S.; Kaufman, S. Vincristine neurotoxicity. Ann. Intern. Med. 1974, 80, 733–737. [Google Scholar] [CrossRef]
- Weiss, H.D.; Walker, M.D.; Wiernik, P.H. Neurotoxicity of commonly used antineoplastic agents. N. Engl. J. Med. 1974, 291, 127–133. [Google Scholar] [CrossRef]
- Bohannon, R.A.; Miller, D.G.; Diamond, H.D. Vincristine in the treatment of lymphomas and leukemias. Cancer Res. 1963, 23, 613–621. [Google Scholar]
- Brandolini, L.; d’Angelo, M.; Antonosante, A.; Cimini, A.; Allegretti, M. Chemokine signaling in chemotherapy-induced neuropathic pain. Int. J. Mol. Sci. 2019, 20, 2904. [Google Scholar] [CrossRef] [Green Version]
- Brzeziński, K. Chemotherapy-induced polyneuropathy. Part I. Pathophysiology [Polish version: Polineuropatia wywołana chemioterapią. Część I. Patofizjologia p. 79]. Contemp. Oncol. Współczesna Onkol. 2012, 16, 72–85. [Google Scholar] [CrossRef] [Green Version]
- White, F.A.; Jung, H.; Miller, R.J. Chemokines and the pathophysiology of neuropathic pain. Proc. Natl. Acad. Sci. USA 2007, 104, 20151–20158. [Google Scholar] [CrossRef] [PubMed]
- Watkins, L.R.; Maier, S.F. Beyond neurons: Evidence that immune and glial cells contribute to pathological pain states. Physiol. Rev. 2002, 82, 981–1011. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.-J.; Ji, R.-R. Chemokines, neuronal–glial interactions, and central processing of neuropathic pain. Pharmacol. Ther. 2010, 126, 56–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Hu, Y.; Li, C.; Yan, Y.; Ao, L.; Yu, B.; Fang, W.; Liu, J.; Li, Y. Levo-corydalmine alleviates vincristine-induced neuropathic pain in mice by inhibiting an NF-kappa B-dependent CXCL1/CXCR2 signaling pathway. Neuropharmacology 2018, 135, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Montague, K.; Simeoli, R.; Valente, J.; Malcangio, M. A novel interaction between CX3CR1 and CCR2 signalling in monocytes constitutes an underlying mechanism for persistent vincristine-induced pain. J. Neuroinflammation 2018, 15, 101. [Google Scholar] [CrossRef] [Green Version]
- Illias, A.M.; Gist, A.C.; Zhang, H.; Kosturakis, A.K.; Dougherty, P.M. Chemokine CCL2 and its receptor CCR2 in the dorsal root ganglion contribute to oxaliplatin-induced mechanical hypersensitivity. Pain 2018, 159, 1308. [Google Scholar] [CrossRef]
- Brandolini, L.; Castelli, V.; Aramini, A.; Giorgio, C.; Bianchini, G.; Russo, R.; De Caro, C.; d’Angelo, M.; Catanesi, M.; Benedetti, E. DF2726A, a new IL-8 signalling inhibitor, is able to counteract chemotherapy-induced neuropathic pain. Sci. Rep. 2019, 9, 11729. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Ye, J.; Yan, H. Investigation of key genes and pathways in inhibition of oxycodone on vincristine-induced microglia activation by using bioinformatics analysis. Dis. Markers 2019, 2019, 3521746. [Google Scholar] [CrossRef]
- Chen, X.-J.; Wang, L.; Song, X.-Y. Mitoquinone alleviates vincristine-induced neuropathic pain through inhibiting oxidative stress and apoptosis via the improvement of mitochondrial dysfunction. Biomed. Pharmacother. 2020, 125, 110003. [Google Scholar] [CrossRef]
- Old, E.A.; Nadkarni, S.; Grist, J.; Gentry, C.; Bevan, S.; Kim, K.-W.; Mogg, A.J.; Perretti, M.; Malcangio, M. Monocytes expressing CX3CR1 orchestrate the development of vincristine-induced pain. J. Clin. Investig. 2014, 124, 2023–2036. [Google Scholar] [CrossRef] [Green Version]
- Makker, P.G.; Duffy, S.S.; Lees, J.G.; Perera, C.J.; Tonkin, R.S.; Butovsky, O.; Park, S.B.; Goldstein, D.; Moalem-Taylor, G. Characterisation of immune and neuroinflammatory changes associated with chemotherapy-induced peripheral neuropathy. PLoS ONE 2017, 12, e0170814. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Yu, Z.; Song, Y.; Zhang, Y.; Du, J.; Su, Y.; Ma, X. Hair follicle bulge-derived stem cells promote tissue regeneration during skin expansion. Biomed. Pharmacother. 2020, 132, 110805. [Google Scholar] [CrossRef]
- Boman, N.L.; Tron, V.A.; Bally, M.B.; Cullis, P.R. Vincristine-induced dermal toxicity is significantly reduced when the drug is given in liposomes. Cancer Chemother. Pharmacol. 1996, 37, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Brandolini, L.; Cristiano, L.; Fidoamore, A.; De Pizzol, M.; Di Giacomo, E.; Florio, T.M.; Confalone, G.; Galante, A.; Cinque, B.; Benedetti, E. Targeting CXCR1 on breast cancer stem cells: Signaling pathways and clinical application modelling. Oncotarget 2015, 6, 43375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branca, J.J.V.; Maresca, M.; Morucci, G.; Becatti, M.; Paternostro, F.; Gulisano, M.; Ghelardini, C.; Salvemini, D.; Mannelli, L.D.C.; Pacini, A. Oxaliplatin-induced blood brain barrier loosening: A new point of view on chemotherapy-induced neurotoxicity. Oncotarget 2018, 9, 23426–23438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Ma, X.-L.; Wei, Y.-Q.; Wei, X.-W. Potential roles and targeted therapy of the CXCLs/CXCR2 axis in cancer and inflammatory diseases. Biochim. Et Biophys. Acta (BBA)-Rev. Cancer 2019, 1871, 289–312. [Google Scholar] [CrossRef]
- Haile, W.B.; Gavegnano, C.; Tao, S.; Jiang, Y.; Schinazi, R.F.; Tyor, W.R. The Janus kinase inhibitor ruxolitinib reduces HIV replication in human macrophages and ameliorates HIV encephalitis in a murine model. Neurobiol. Dis. 2016, 92, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Boyette-Davis, J.; Xin, W.; Zhang, H.; Dougherty, P. Intraepidermal nerve fiber loss corresponds to the development of taxol-induced hyperalgesia and can be prevented by treatment with minocycline. PAIN® 2011, 152, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Agbaje, M.; Rutland, C.S.; Maboni, G.; Blanchard, A.; Bexon, M.; Stewart, C.; Jones, M.A.; Totemeyer, S. Novel inflammatory cell infiltration scoring system to investigate healthy and footrot affected ovine interdigital skin. PeerJ 2018, 6, e5097. [Google Scholar] [CrossRef] [Green Version]
- Kingery, W.S. Role of neuropeptide, cytokine, and growth factor signaling in complex regional pain syndrome. Pain Med. 2010, 11, 1239–1250. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Zhu, M.; Yuan, S.; Yu, W. Spinal CXCL5 contributes to nerve injury-induced neuropathic pain via modulating GSK-3β phosphorylation and activity in rats. Neurosci. Lett. 2016, 634, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-J.; Cao, D.-L.; Zhang, X.; Ji, R.-R.; Gao, Y.-J. Chemokine contribution to neuropathic pain: Respective induction of CXCL1 and CXCR2 in spinal cord astrocytes and neurons. PAIN® 2013, 154, 2185–2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.-B.; Wang, Y.-J.; Wan, W.-J.; Wu, J.; Wang, B.-J.; Zhu, H.-L.; Xie, M.; Liu, L. Resveratrol ameliorates oxaliplatin-induced neuropathic pain via anti-inflammatory effects in rats. Exp. Ther. Med. 2022, 24, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Starobova, H.; Monteleone, M.; Adolphe, C.; Batoon, L.; Sandrock, C.J.; Tay, B.; Deuis, J.R.; Smith, A.V.; Mueller, A.; Nadar, E.I. Vincristine-induced peripheral neuropathy is driven by canonical NLRP3 activation and IL-1β release. J. Exp. Med. 2021, 218, e20201452. [Google Scholar] [CrossRef] [PubMed]
- Larsson, B.; Skarby, T.; Edvinsson, L.; Hardebo, J.; Owman, C. Vincristine reduces damage of the blood-brain barrier induced by high intravascular pressure. Neurosci. Lett. 1980, 17, 155–159. [Google Scholar] [CrossRef]
- Burger, M.; Hartmann, T.; Burger, J.A.; Schraufstatter, I. KSHV-GPCR and CXCR2 transforming capacity and angiogenic responses are mediated through a JAK2-STAT3-dependent pathway. Oncogene 2005, 24, 2067–2075. [Google Scholar] [CrossRef] [Green Version]
- Bonin, R.P.; Bories, C.; De Koninck, Y. A simplified up-down method (SUDO) for measuring mechanical nociception in rodents using von Frey filaments. Mol. Pain 2014, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.; Chung, J.; Yaksh, T. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, H.S.; Choi, Y.I.; Park, S.U.; Han, Y.S.; Kwon, J.; Jung, S.J. Prevention of Chemotherapy-Induced Peripheral Neuropathy by Inhibiting C-X-C Motif Chemokine Receptor 2. Int. J. Mol. Sci. 2023, 24, 1855. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24031855
Cho HS, Choi YI, Park SU, Han YS, Kwon J, Jung SJ. Prevention of Chemotherapy-Induced Peripheral Neuropathy by Inhibiting C-X-C Motif Chemokine Receptor 2. International Journal of Molecular Sciences. 2023; 24(3):1855. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24031855
Chicago/Turabian StyleCho, Hee Seong, Young In Choi, Seon Uk Park, Yi Seul Han, Jean Kwon, and Sung Jun Jung. 2023. "Prevention of Chemotherapy-Induced Peripheral Neuropathy by Inhibiting C-X-C Motif Chemokine Receptor 2" International Journal of Molecular Sciences 24, no. 3: 1855. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24031855