
Inhaled Nitric Oxide Promotes Angiogenesis in the Rodent Developing Brain
 , and
, and 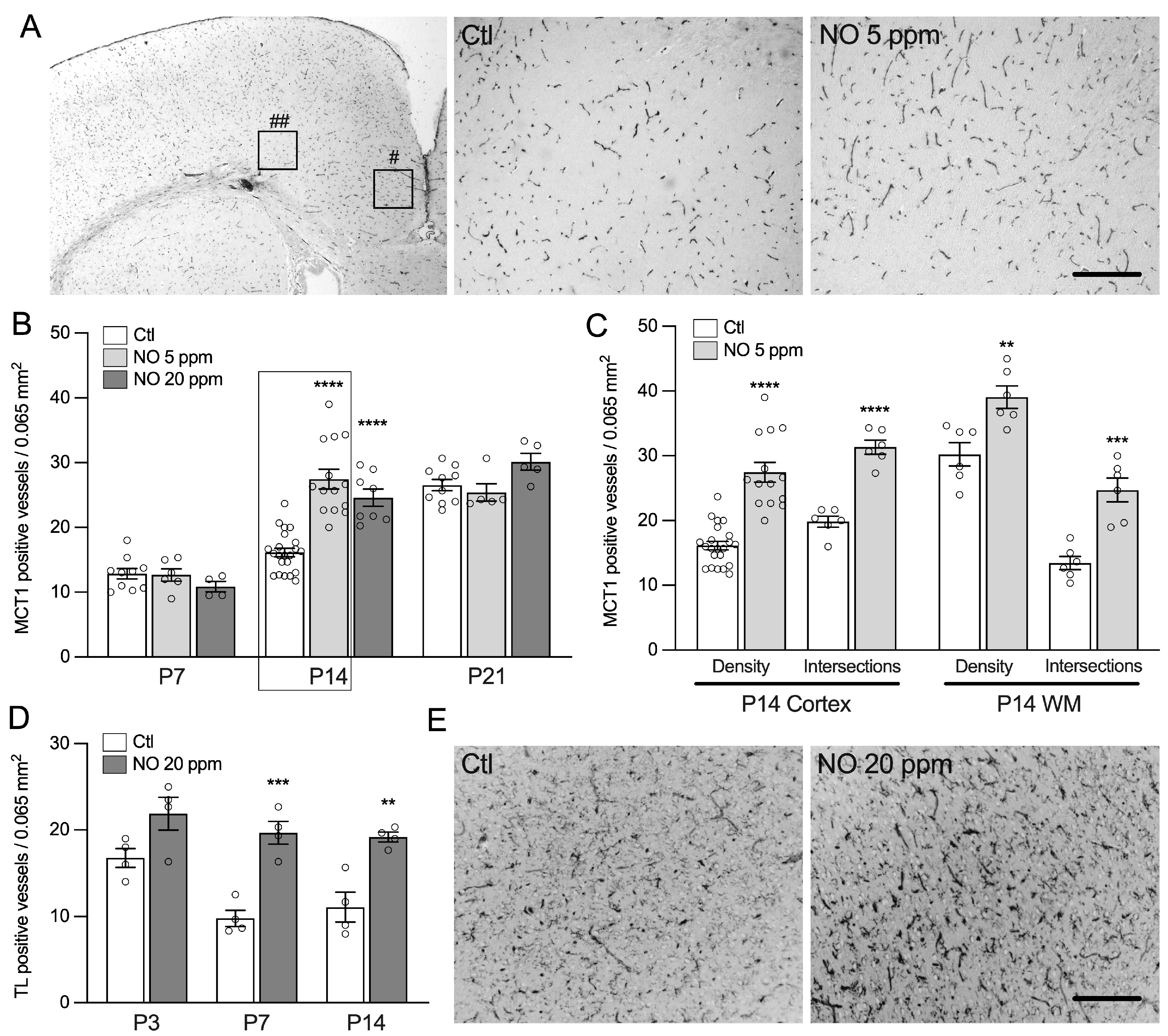{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
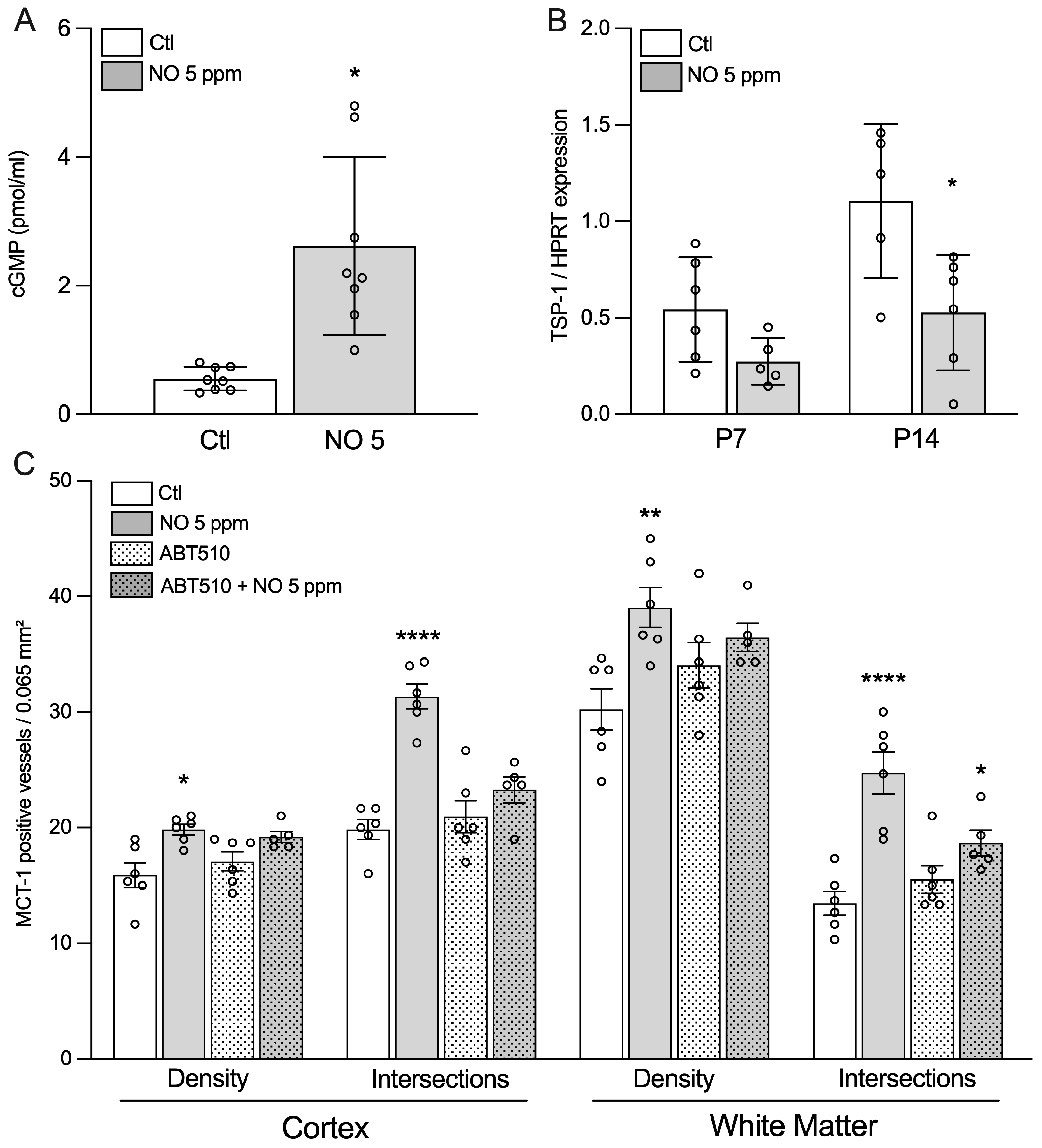
2. Results
2.1. Inhaled NO Promotes Angiogenesis in the Developing Brain during a Critical Window
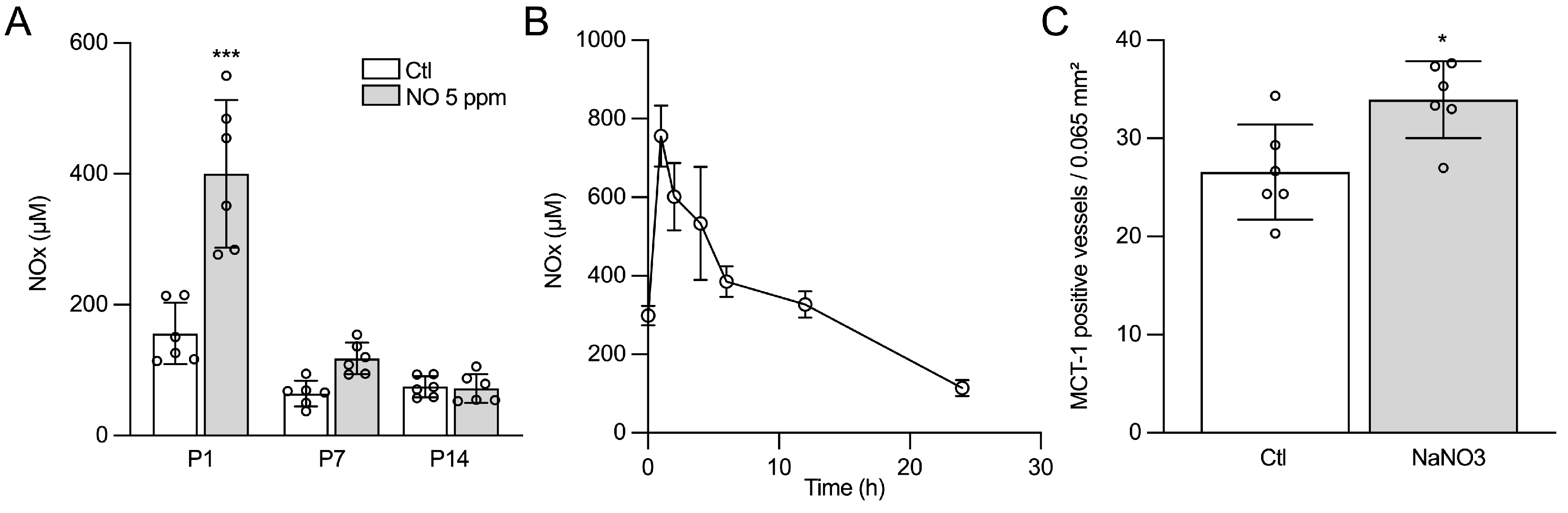
2.2. The Effects of iNO on Angiogenesis Are Mimicked by Nitrate/Nitrite
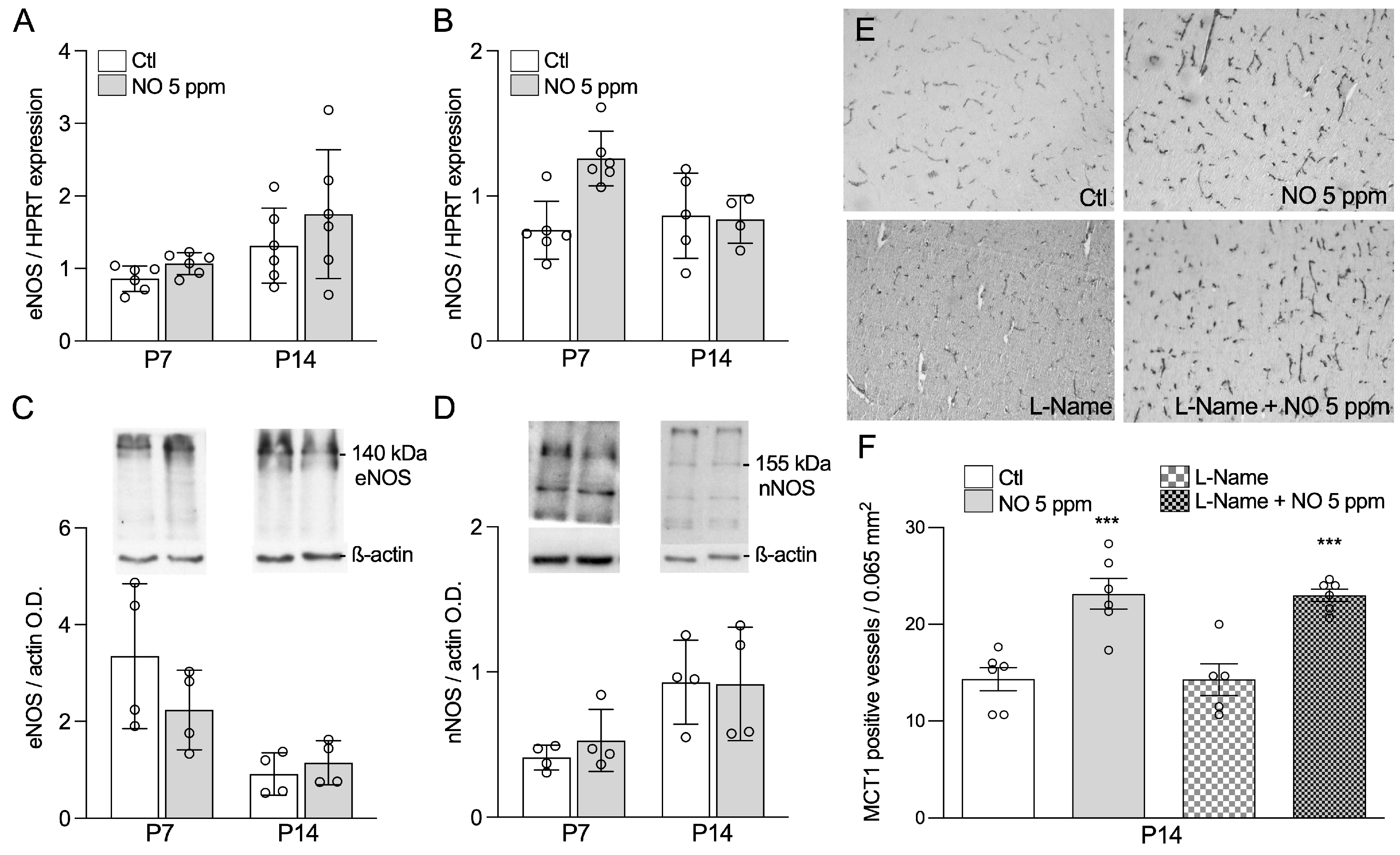
2.3. Inhaled NO Effects in the Developing Brain Do Not Engage Changes in Endogenous NO Synthesis
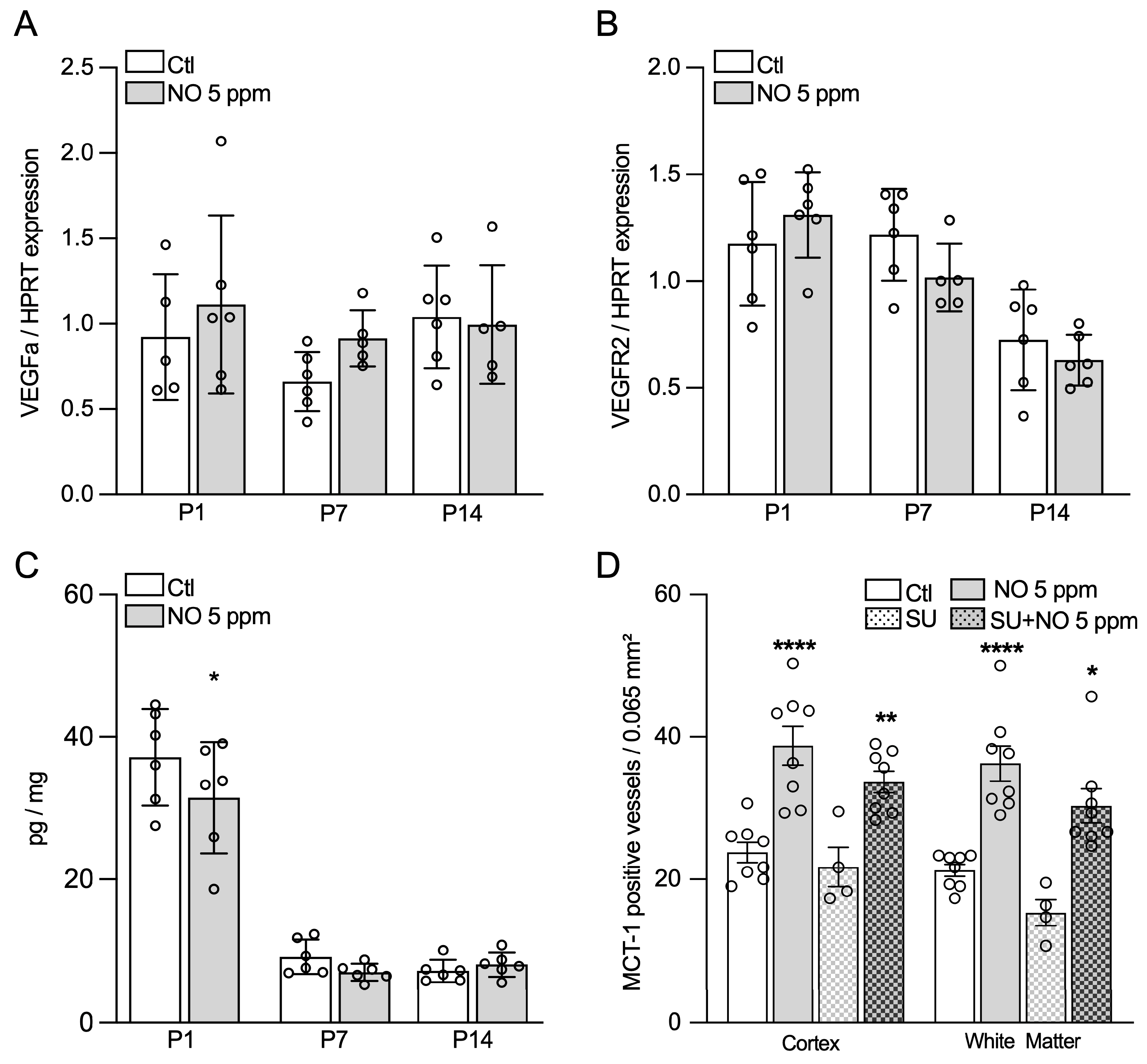
2.4. Inhaled NO Effects in the Developing Brain Are Not Mediated by NO-Related Angiogenic Factors
2.5. Evidence for the Role of Soluble Guanylate Cyclase/cGMP Pathway
3. Discussion
3.1. Signaling Pathways Involved in iNO Angiogenic Effects in the Developing Brain
3.2. Remote Effect of iNO on Peripheral Organs
3.3. iNO and Angiogenesis: A Role in the Neuroprotective Effect of the NO Pathway in Preclinical Models of Brain Injury?
3.4. NO Pathway: A New Neuroprotective Strategy in Human Neonates?
3.5. Conclusions
4. Materials and Methods
4.1. Animals, NO Exposure, and Brain Sampling
4.2. Drugs
4.3. Immunohistochemistry and Vessel Wall Density Quantification
4.4. Physiological Parameters
4.5. ELISA Assays
4.6. Western Blot
4.7. Quantitative Real-Time Polymerase Chain Reaction
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chachlaki, K.; Prevot, V. Nitric Oxide Signalling in the Brain and Its Control of Bodily Functions. Br. J. Pharmacol. 2020, 177, 5437–5458. [Google Scholar] [CrossRef] [PubMed]
- Zapol, W.M. Nitric Oxide Story. Anesthesiology 2019, 130, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.D.; Polaner, D.M.; Zapol, W.M.; Lang, P. Inhaled Nitric Oxide in Persistent Pulmonary Hypertension of the Newborn. Lancet 1992, 340, 818–819. [Google Scholar] [CrossRef] [PubMed]
- Barrington, K.J.; Finer, N.; Pennaforte, T.; Altit, G. Nitric Oxide for Respiratory Failure in Infants Born at or near Term. Cochrane Database Syst. Rev. 2017, 1, CD000399. [Google Scholar] [CrossRef]
- Thébaud, B.; Goss, K.N.; Laughon, M.; Whitsett, J.A.; Abman, S.H.; Steinhorn, R.H.; Aschner, J.L.; Davis, P.G.; McGrath-Morrow, S.A.; Soll, R.F.; et al. Bronchopulmonary Dysplasia. Nat. Rev. Dis. Prim. 2019, 5, 78. [Google Scholar] [CrossRef]
- Barrington, K.J.; Finer, N.; Pennaforte, T. Inhaled Nitric Oxide for Respiratory Failure in Preterm Infants. Cochrane Database Syst. Rev. 2017, 1, CD000509. [Google Scholar] [CrossRef]
- Terpolilli, N.A.; Kim, S.-W.; Thal, S.C.; Kataoka, H.; Zeisig, V.; Nitzsche, B.; Klaesner, B.; Zhu, C.; Schwarzmaier, S.; Meissner, L.; et al. Inhalation of Nitric Oxide Prevents Ischemic Brain Damage in Experimental Stroke by Selective Dilatation of Collateral Arterioles. Circ. Res. 2012, 110, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Pansiot, J.; Loron, G.; Olivier, P.; Fontaine, R.; Charriaut-Marlangue, C.; Mercier, J.-C.; Gressens, P.; Baud, O. Neuroprotective Effect of Inhaled Nitric Oxide on Excitotoxic-Induced Brain Damage in Neonatal Rat. PLoS ONE 2010, 5, e10916. [Google Scholar] [CrossRef] [Green Version]
- Charriaut-Marlangue, C.; Bonnin, P.; Gharib, A.; Leger, P.-L.; Villapol, S.; Pocard, M.; Gressens, P.; Renolleau, S.; Baud, O. Inhaled Nitric Oxide Reduces Brain Damage by Collateral Recruitment in a Neonatal Stroke Model. Stroke 2012, 43, 3078–3084. [Google Scholar] [CrossRef] [Green Version]
- Premont, R.T.; Reynolds, J.D.; Zhang, R.; Stamler, J.S. Role of Nitric Oxide Carried by Hemoglobin in Cardiovascular Physiology: Developments on a Three-Gas Respiratory Cycle. Circ. Res. 2020, 126, 129–158. [Google Scholar] [CrossRef]
- Olivier, P.; Loron, G.; Fontaine, R.H.; Pansiot, J.; Dalous, J.; Thi, H.P.; Charriaut-Marlangue, C.; Thomas, J.-L.; Mercier, J.-C.; Gressens, P.; et al. Nitric Oxide Plays a Key Role in Myelination in the Developing Brain. J. Neuropathol. Exp. Neurol. 2010, 69, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Yuen, T.J.; Silbereis, J.C.; Griveau, A.; Chang, S.M.; Daneman, R.; Fancy, S.P.J.; Zahed, H.; Maltepe, E.; Rowitch, D.H. Oligodendrocyte-Encoded HIF Function Couples Postnatal Myelination and White Matter Angiogenesis. Cell 2014, 158, 383–396. [Google Scholar] [CrossRef] [Green Version]
- Duy, A.P.; Pham, H.; Pansiot, J.; Gressens, P.; Charriaut-Marlangue, C.; Baud, O. Nitric Oxide Pathway and Proliferation of Neural Progenitors in the Neonatal Rat. DNE 2015, 37, 417–427. [Google Scholar] [CrossRef]
- McQuade, L.E.; Lippard, S.J. Fluorescent Probes to Investigate Nitric Oxide and Other Reactive Nitrogen Species in Biology (Truncated Form: Fluorescent Probes of Reactive Nitrogen Species). Curr. Opin. Chem. Biol. 2010, 14, 43–49. [Google Scholar] [CrossRef]
- Eichmann, A.; Simons, M. VEGF Signaling inside Vascular Endothelial Cells and Beyond. Curr. Opin. Chem. Biol. 2012, 24, 188–193. [Google Scholar] [CrossRef] [Green Version]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [Green Version]
- Isenberg, J.S.; Frazier, W.A.; Roberts, D.D. Thrombospondin-1: A Physiological Regulator of Nitric Oxide Signaling. Cell. Mol. Life Sci. 2008, 65, 728–742. [Google Scholar] [CrossRef] [Green Version]
- Isenberg, J.S.; Ridnour, L.A.; Perruccio, E.M.; Espey, M.G.; Wink, D.A.; Roberts, D.D. Thrombospondin-1 Inhibits Endothelial Cell Responses to Nitric Oxide in a CGMP-Dependent Manner. Proc. Natl. Acad. Sci. USA 2005, 102, 13141–13146. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.D.; Isenberg, J.S.; Ridnour, L.A.; Wink, D.A. Nitric Oxide and Its Gatekeeper Thrombospondin-1 in Tumor Angiogenesis. Clin. Cancer Res. 2007, 13, 795–798. [Google Scholar] [CrossRef] [Green Version]
- Ebbinghaus, S.; Hussain, M.; Tannir, N.; Gordon, M.; Desai, A.A.; Knight, R.A.; Humerickhouse, R.A.; Qian, J.; Gordon, G.B.; Figlin, R. Phase 2 Study of ABT-510 in Patients with Previously Untreated Advanced Renal Cell Carcinoma. Clin. Cancer Res. 2007, 13, 6689–6695. [Google Scholar] [CrossRef] [Green Version]
- Baker, L.H.; Rowinsky, E.K.; Mendelson, D.; Humerickhouse, R.A.; Knight, R.A.; Qian, J.; Carr, R.A.; Gordon, G.B.; Demetri, G.D. Randomized, Phase II Study of the Thrombospondin-1-Mimetic Angiogenesis Inhibitor ABT-510 in Patients with Advanced Soft Tissue Sarcoma. J. Clin. Oncol. 2008, 26, 5583–5588. [Google Scholar] [CrossRef] [PubMed]
- Kuebler, W.M.; Kisch-Wedel, H.; Kemming, G.I.; Meisner, F.; Bruhn, S.; Koehler, C.; Flondor, M.; Messmer, K.; Zwissler, B. Inhaled Nitric Oxide Induces Cerebrovascular Effects in Anesthetized Pigs. Neurosci. Lett. 2003, 348, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Troncy, E.; Francoeur, M.; Salazkin, I.; Yang, F.; Charbonneau, M.; Leclerc, G.; Vinay, P.; Blaise, G. Extra-Pulmonary Effects of Inhaled Nitric Oxide in Swine with and without Phenylephrine. Br. J. Anaesth. 1997, 79, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Cannon, R.O.; Schechter, A.N.; Panza, J.A.; Ognibene, F.P.; Pease-Fye, M.E.; Waclawiw, M.A.; Shelhamer, J.H.; Gladwin, M.T. Effects of Inhaled Nitric Oxide on Regional Blood Flow Are Consistent with Intravascular Nitric Oxide Delivery. J. Clin. Investig. 2001, 108, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Fox-Robichaud, A.; Payne, D.; Hasan, S.U.; Ostrovsky, L.; Fairhead, T.; Reinhardt, P.; Kubes, P. Inhaled NO as a Viable Antiadhesive Therapy for Ischemia/Reperfusion Injury of Distal Microvascular Beds. J. Clin. Investig. 1998, 101, 2497–2505. [Google Scholar] [CrossRef] [Green Version]
- Cirino, G.; Vellecco, V.; Bucci, M. Nitric Oxide and Hydrogen Sulfide: The Gasotransmitter Paradigm of the Vascular System. Br. J. Pharmacol. 2017, 174, 4021–4031. [Google Scholar] [CrossRef] [Green Version]
- Kazerounian, S.; Lawler, J. Integration of Pro- and Anti-Angiogenic Signals by Endothelial Cells. J. Cell Commun. Signal. 2018, 12, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Isenberg, J.S.; Martin-Manso, G.; Maxhimer, J.B.; Roberts, D.D. Regulation of Nitric Oxide Signalling by Thrombospondin 1: Implications for Anti-Angiogenic Therapies. Nat. Rev. Cancer 2009, 9, 182–194. [Google Scholar] [CrossRef] [Green Version]
- Hess, D.T.; Stamler, J.S. Regulation by S-Nitrosylation of Protein Post-Translational Modification. J. Biol. Chem. 2012, 287, 4411–4418. [Google Scholar] [CrossRef] [Green Version]
- Anand, P.; Stamler, J.S. Enzymatic Mechanisms Regulating Protein S-Nitrosylation: Implications in Health and Disease. J. Mol. Med. 2012, 90, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Benhar, M.; Forrester, M.T.; Stamler, J.S. Protein Denitrosylation: Enzymatic Mechanisms and Cellular Functions. Nat. Rev. Mol. Cell Biol. 2009, 10, 721–732. [Google Scholar] [CrossRef]
- Seth, D.; Hess, D.T.; Hausladen, A.; Wang, L.; Wang, Y.; Stamler, J.S. A Multiplex Enzymatic Machinery for Cellular Protein S-Nitrosylation. Mol. Cell 2018, 69, 451–464.e6. [Google Scholar] [CrossRef] [Green Version]
- Gupta, K.J.; Kolbert, Z.; Durner, J.; Lindermayr, C.; Corpas, F.J.; Brouquisse, R.; Barroso, J.B.; Umbreen, S.; Palma, J.M.; Hancock, J.T.; et al. Regulating the Regulator: Nitric Oxide Control of Post-Translational Modifications. New Phytol. 2020, 227, 1319–1325. [Google Scholar] [CrossRef]
- Ballou, D.P.; Zhao, Y.; Brandish, P.E.; Marletta, M.A. Revisiting the Kinetics of Nitric Oxide (NO) Binding to Soluble Guanylate Cyclase: The Simple NO-Binding Model Is Incorrect. Proc. Natl. Acad. Sci. USA 2002, 99, 12097–12101. [Google Scholar] [CrossRef] [Green Version]
- Wanstall, J.C.; Homer, K.L.; Doggrell, S.A. Evidence for, and Importance of, CGMP-Independent Mechanisms with NO and NO Donors on Blood Vessels and Platelets. Curr. Vasc. Pharmacol. 2005, 3, 41–53. [Google Scholar] [CrossRef]
- Conahey, G.R.; Power, G.G.; Hopper, A.O.; Terry, M.H.; Kirby, L.S.; Blood, A.B. Effect of Inhaled Nitric Oxide on Cerebrospinal Fluid and Blood Nitrite Concentrations in Newborn Lambs. Pediatr. Res. 2008, 64, 375–380. [Google Scholar] [CrossRef]
- Pham, H.; Vottier, G.; Pansiot, J.; Duong-Quy, S.; Bollen, B.; Dalous, J.; Gallego, J.; Mercier, J.-C.; Dinh-Xuan, A.T.; Bonnin, P.; et al. Inhaled NO Prevents Hyperoxia-Induced White Matter Damage in Neonatal Rats. Exp. Neurol. 2014, 252, 114–123. [Google Scholar] [CrossRef]
- Pham, H.; Vottier, G.; Pansiot, J.; Dalous, J.; Gallego, J.; Gressens, P.; Duong-Quy, S.; Dinh-Xuan, A.-T.; Mercier, J.-C.; Biran, V.; et al. Inhaled NO Protects Cerebral White Matter in Neonatal Rats with Combined Brain and Lung Injury. Am. J. Respir. Crit. Care Med. 2012, 185, 897–899. [Google Scholar] [CrossRef]
- Pham, H.; Duy, A.P.; Pansiot, J.; Bollen, B.; Gallego, J.; Charriaut-Marlangue, C.; Baud, O. Impact of Inhaled Nitric Oxide on White Matter Damage in Growth-Restricted Neonatal Rats. Pediatr. Res. 2015, 77, 563–569. [Google Scholar] [CrossRef]
- Elahi, M.M.; Worner, M.; Khan, J.S.; Matata, B.M. Inspired Nitric Oxide and Modulation of Oxidative Stress during Cardiac Surgery. Curr. Drug Saf. 2009, 4, 188–198. [Google Scholar] [CrossRef]
- Brücken, A.; Derwall, M.; Bleilevens, C.; Stoppe, C.; Götzenich, A.; Gaisa, N.T.; Weis, J.; Nolte, K.W.; Rossaint, R.; Ichinose, F.; et al. Brief Inhalation of Nitric Oxide Increases Resuscitation Success and Improves 7-Day-Survival after Cardiac Arrest in Rats: A Randomized Controlled Animal Study. Crit. Care 2015, 19, 408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdani, A.; Khoja, Z.; Johnstone, A.; Dale, L.; Rampakakis, E.; Wintermark, P. Sildenafil Improves Brain Injury Recovery Following Term Neonatal Hypoxia-Ischemia in Male Rat Pups. Dev. Neurosci. 2016, 38, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Charriaut-Marlangue, C.; Nguyen, T.; Bonnin, P.; Duy, A.P.; Leger, P.-L.; Csaba, Z.; Pansiot, J.; Bourgeois, T.; Renolleau, S.; Baud, O. Sildenafil Mediates Blood-Flow Redistribution and Neuroprotection after Neonatal Hypoxia-Ischemia. Stroke 2014, 45, 850–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdani, A.; Howidi, B.; Shi, M.Z.; Tugarinov, N.; Khoja, Z.; Wintermark, P. Sildenafil Improves Hippocampal Brain Injuries and Restores Neuronal Development after Neonatal Hypoxia-Ischemia in Male Rat Pups. Sci. Rep. 2021, 11, 22046. [Google Scholar] [CrossRef]
- Moretti, R.; Leger, P.-L.; Besson, V.C.; Csaba, Z.; Pansiot, J.; Di Criscio, L.; Gentili, A.; Titomanlio, L.; Bonnin, P.; Baud, O.; et al. Sildenafil, a Cyclic GMP Phosphodiesterase Inhibitor, Induces Microglial Modulation after Focal Ischemia in the Neonatal Mouse Brain. J. Neuroinflamm. 2016, 13, 95. [Google Scholar] [CrossRef]
- Zinni, M.; Pansiot, J.; Léger, P.-L.; El Kamouh, M.; Baud, O. Sildenafil-Mediated Neuroprotection from Adult to Neonatal Brain Injury: Evidence, Mechanisms, and Future Translation. Cells 2021, 10, 2766. [Google Scholar] [CrossRef]
- Terpolilli, N.A.; Kim, S.-W.; Thal, S.C.; Kuebler, W.M.; Plesnila, N. Inhaled Nitric Oxide Reduces Secondary Brain Damage after Traumatic Brain Injury in Mice. J. Cereb. Blood Flow Metab. 2013, 33, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Armstead, W.M.; Vavilala, M.S. Translational Approach towards Determining the Role of Cerebral Autoregulation in Outcome after Traumatic Brain Injury. Exp. Neurol. 2019, 317, 291–297. [Google Scholar] [CrossRef]
- Liu, P.; Li, Y.-S.; Quartermain, D.; Boutajangout, A.; Ji, Y. Inhaled Nitric Oxide Improves Short Term Memory and Reduces the Inflammatory Reaction in a Mouse Model of Mild Traumatic Brain Injury. Brain Res. 2013, 1522, 67–75. [Google Scholar] [CrossRef]
- Degos, V.; Loron, G.; Mantz, J.; Gressens, P. Neuroprotective Strategies for the Neonatal Brain. Anesth. Analg. 2008, 106, 1670–1680. [Google Scholar] [CrossRef] [Green Version]
- Volpe, J.J. Dysmaturation of Premature Brain: Importance, Cellular Mechanisms, and Potential Interventions. Pediatr. Neurol. 2019, 95, 42–66. [Google Scholar] [CrossRef]
- Greenough, A.; Decobert, F.; Field, D.; Hallman, M.; Hummler, H.D.; Jonsson, B.; Sánchez Luna, M.; Van Overmeire, B.; Carnielli, V.P.; Potenziano, J.L.; et al. Inhaled Nitric Oxide (INO) for Preventing Prematurity-Related Bronchopulmonary Dysplasia (BPD): 7-Year Follow-up of the European Union Nitric Oxide (EUNO) Trial. J. Perinat. Med. 2020, 49, 104–110. [Google Scholar] [CrossRef]
- Charriaut-Marlangue, C.; Baud, O. A Model of Perinatal Ischemic Stroke in the Rat: 20 Years Already and What Lessons? Front. Neurol. 2018, 9, 650. [Google Scholar] [CrossRef]
- Angelis, D.; Savani, R.; Chalak, L. Nitric Oxide and the Brain. Part 2: Effects Following Neonatal Brain Injury-Friend or Foe? Pediatr. Res. 2021, 89, 746–752. [Google Scholar] [CrossRef]
- Ginsberg, M.D. The Cerebral Collateral Circulation: Relevance to Pathophysiology and Treatment of Stroke. Neuropharmacology 2018, 134, 280–292. [Google Scholar] [CrossRef]
- Baud, O.; Fayol, L.; Gressens, P.; Pellerin, L.; Magistretti, P.; Evrard, P.; Verney, C. Perinatal and Early Postnatal Changes in the Expression of Monocarboxylate Transporters MCT1 and MCT2 in the Rat Forebrain. J. Comp. Neurol. 2003, 465, 445–454. [Google Scholar] [CrossRef]
- Fayol, L.; Baud, O.; Monier, A.; Pellerin, L.; Magistretti, P.; Evrard, P.; Verney, C. Immunocytochemical Expression of Monocarboxylate Transporters in the Human Visual Cortex at Midgestation. Brain Res. Dev. Brain Res. 2004, 148, 69–76. [Google Scholar] [CrossRef]
- Fontaine, R.H.; Olivier, P.; Massonneau, V.; Leroux, P.; Degos, V.; Lebon, S.; El Ghouzzi, V.; Lelièvre, V.; Gressens, P.; Baud, O. Vulnerability of White Matter towards Antenatal Hypoxia Is Linked to a Species-Dependent Regulation of Glutamate Receptor Subunits. Proc. Natl. Acad. Sci. USA 2008, 105, 16779–16784. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loron, G.; Pansiot, J.; Olivier, P.; Charriaut-Marlangue, C.; Baud, O. Inhaled Nitric Oxide Promotes Angiogenesis in the Rodent Developing Brain. Int. J. Mol. Sci. 2023, 24, 5871. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065871
Loron G, Pansiot J, Olivier P, Charriaut-Marlangue C, Baud O. Inhaled Nitric Oxide Promotes Angiogenesis in the Rodent Developing Brain. International Journal of Molecular Sciences. 2023; 24(6):5871. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065871
Chicago/Turabian StyleLoron, Gauthier, Julien Pansiot, Paul Olivier, Christiane Charriaut-Marlangue, and Olivier Baud. 2023. "Inhaled Nitric Oxide Promotes Angiogenesis in the Rodent Developing Brain" International Journal of Molecular Sciences 24, no. 6: 5871. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065871