

The Direct Anti-Virulence but Not Bactericidal Activity of Human Neutrophil Elastase against Moraxella catarrhalis


Abstract
:1. Introduction
2. Results
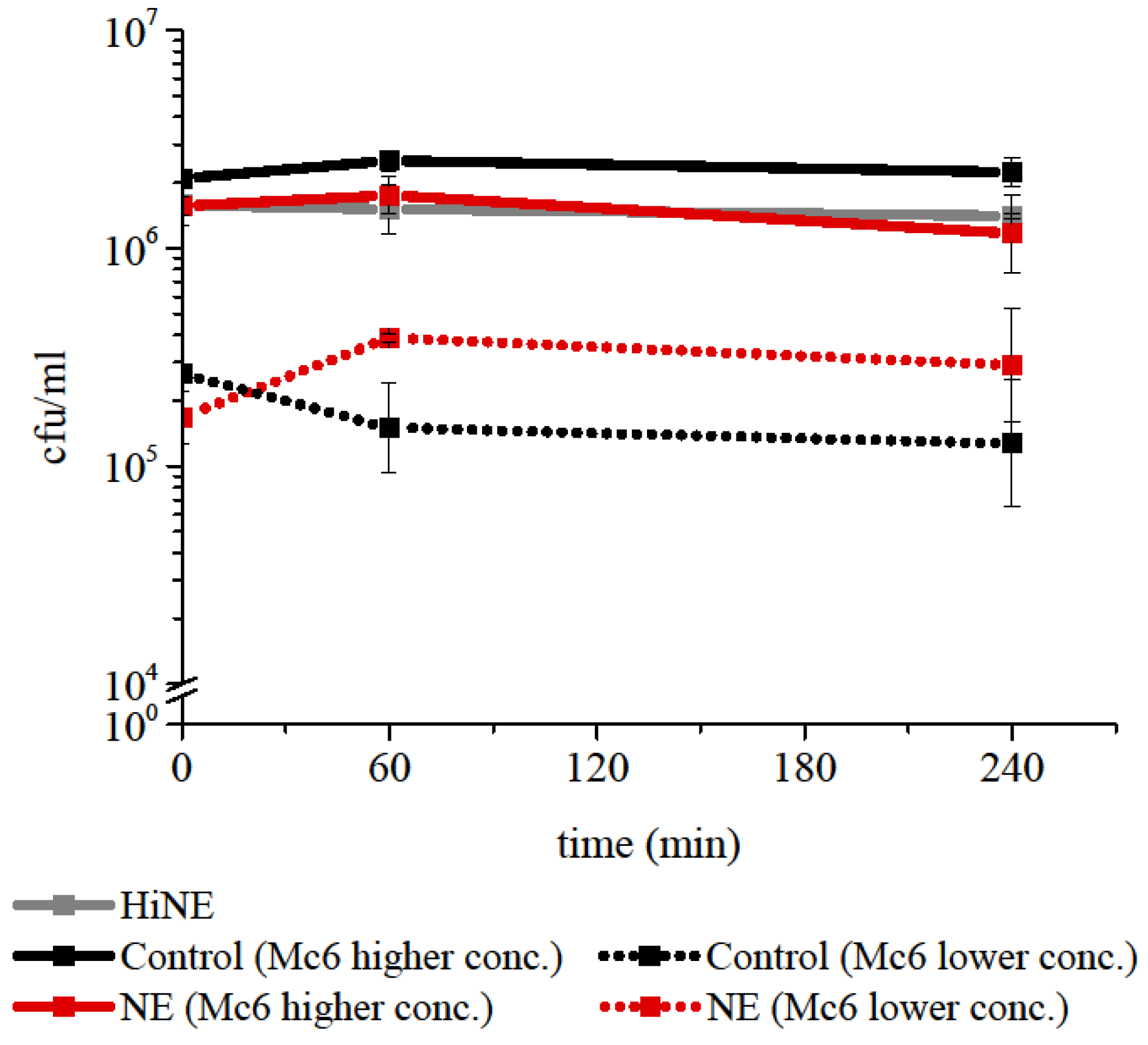
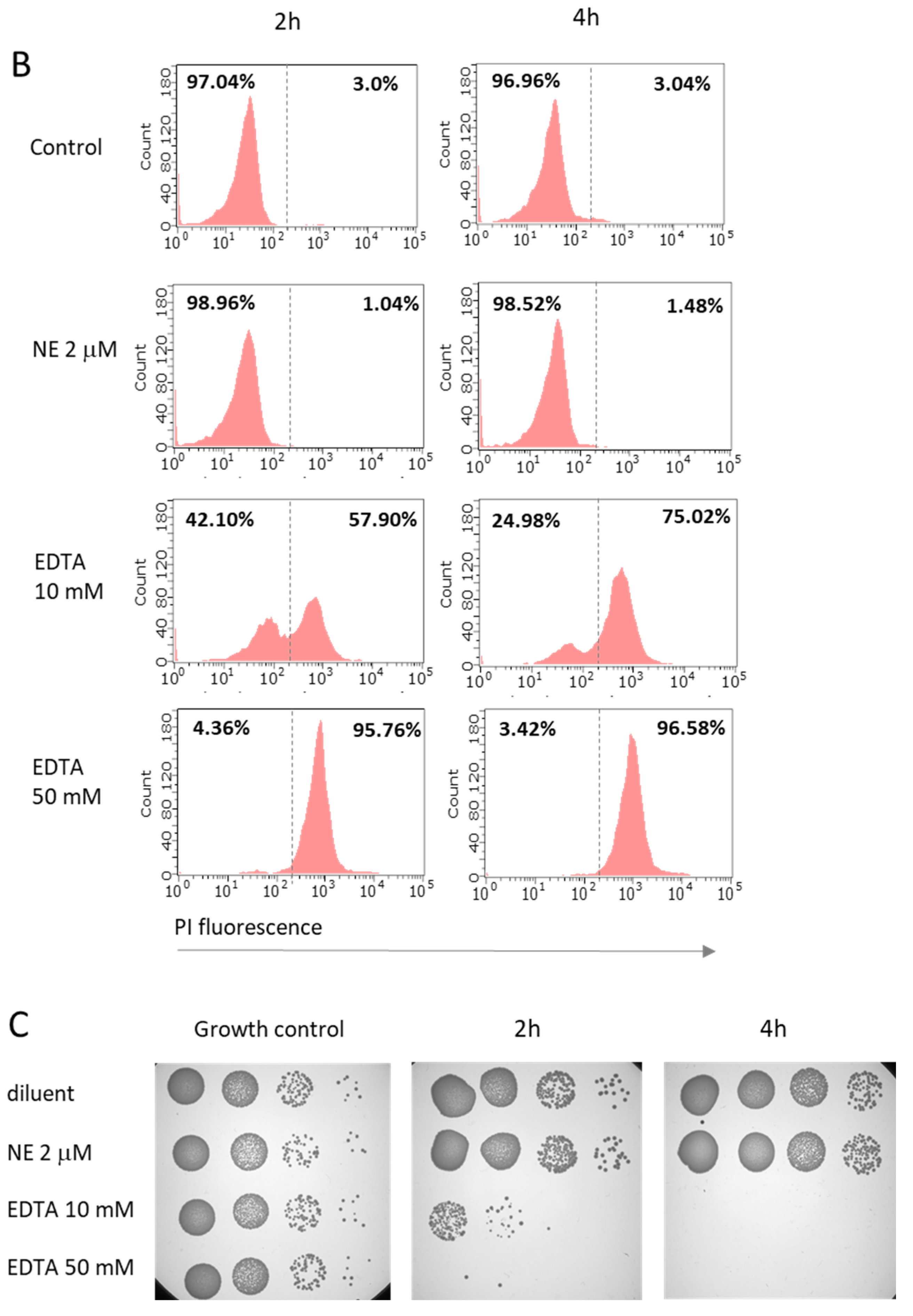
2.1. Neutrophil elastase at Concentration Representative for Pathological Conditions of Respiratory Tract Does Not Exert Direct Killing against M. catarrhalis
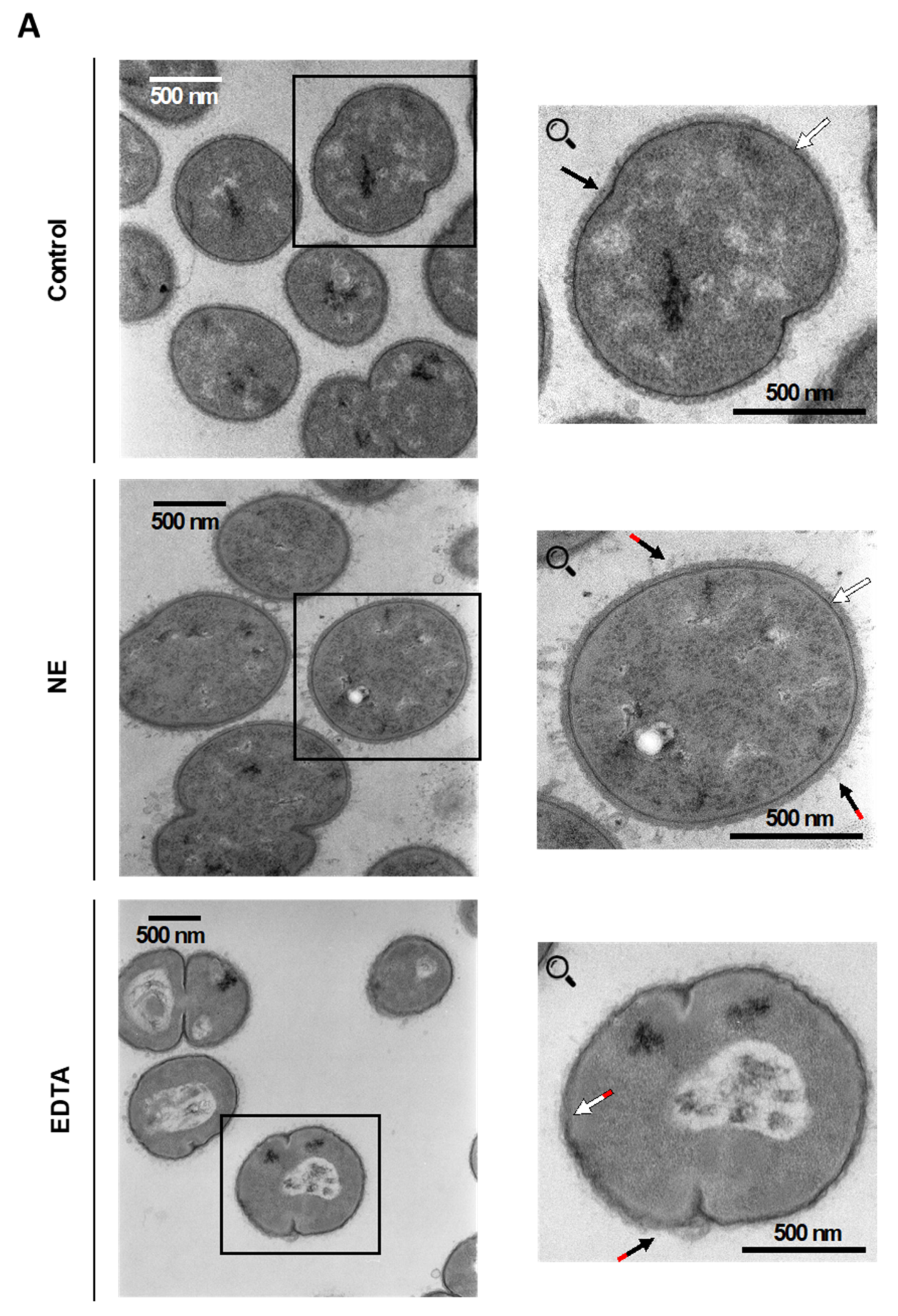
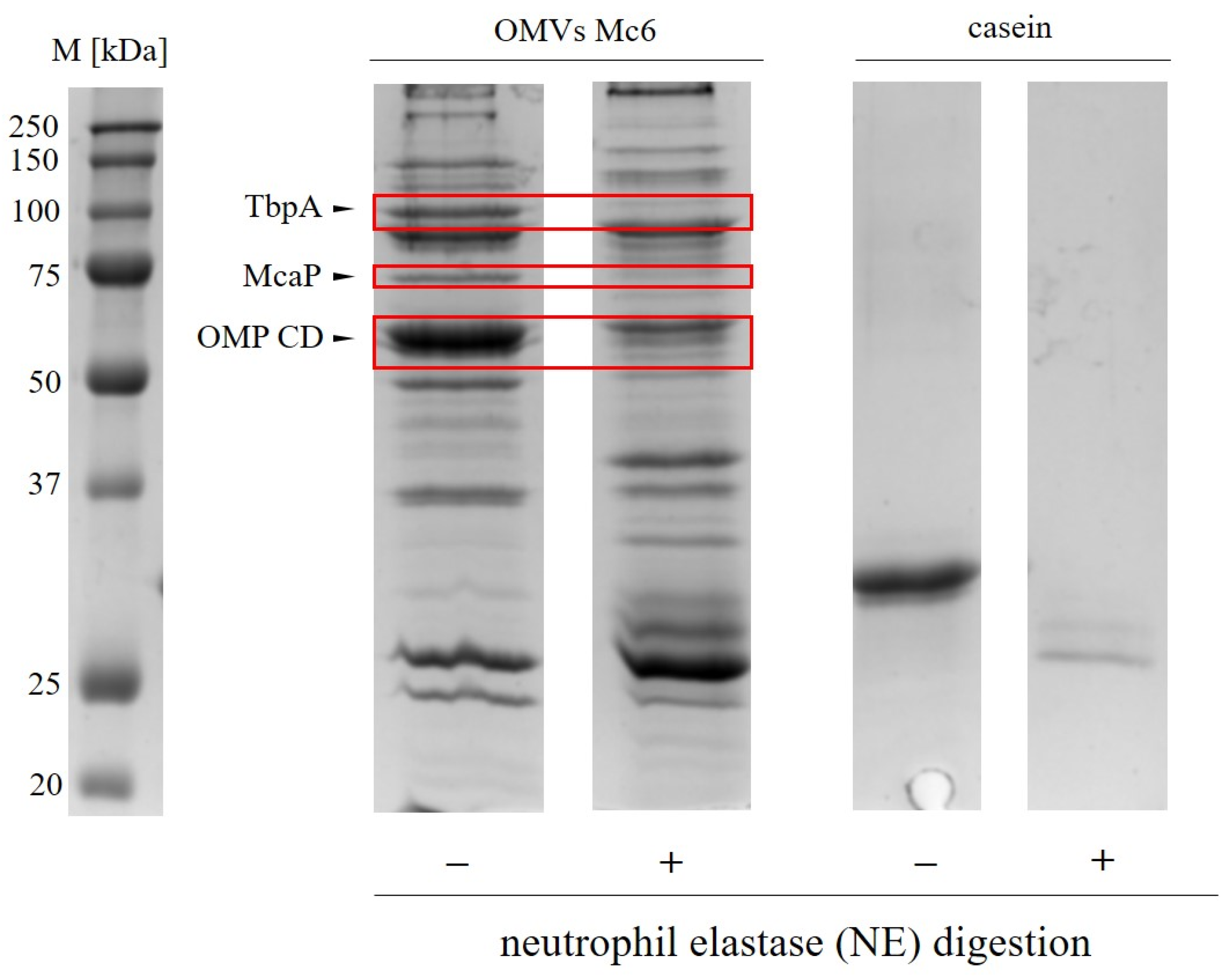
2.2. Neutrophil Elastase Degrades Pivotal Outer Membrane Proteins (Virulence Factors) of M. catarrhalis
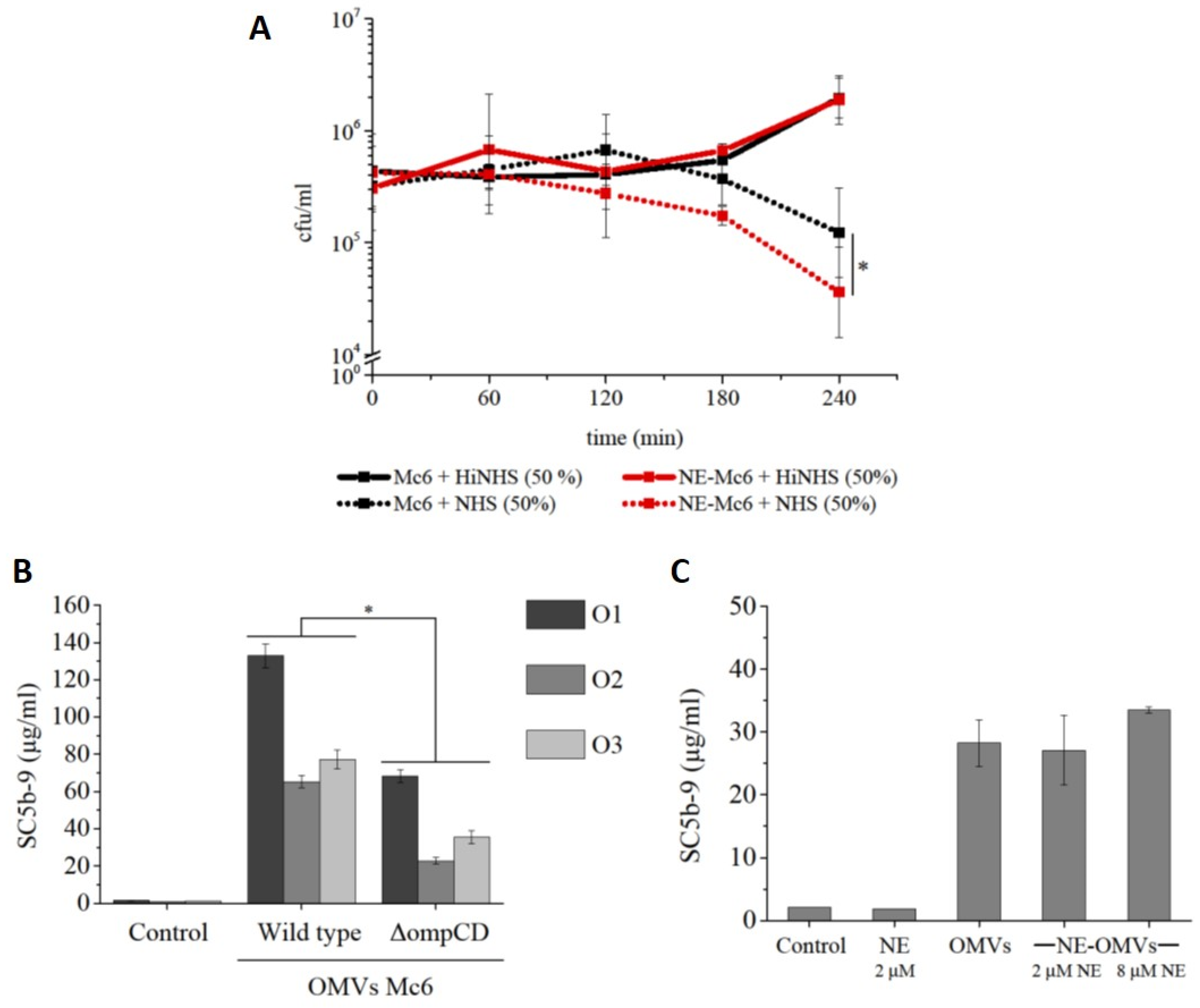
2.3. Degradation of OMP CD by Neutrophil Elastase Sensitizes M. catarrhalis to Complement Action
2.3.1. Complement-Dependent Bactericidal Activity
2.3.2. Activation of Terminal Complement SC5b-9 Component
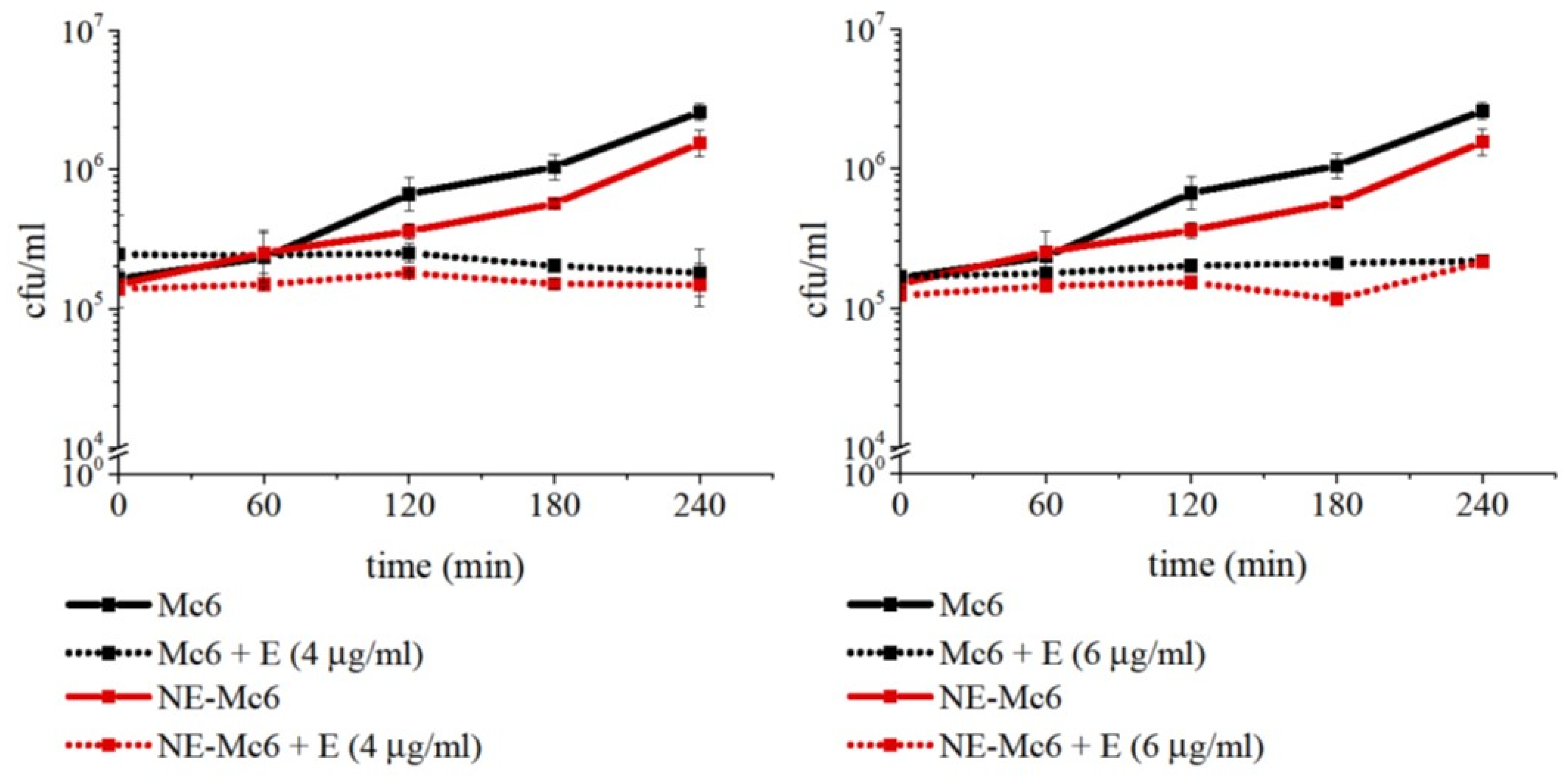
2.4. Degradation of McaP by Neutrophil Elastase Does Not Sensitize M. catarrhalis to Erythromycin Action
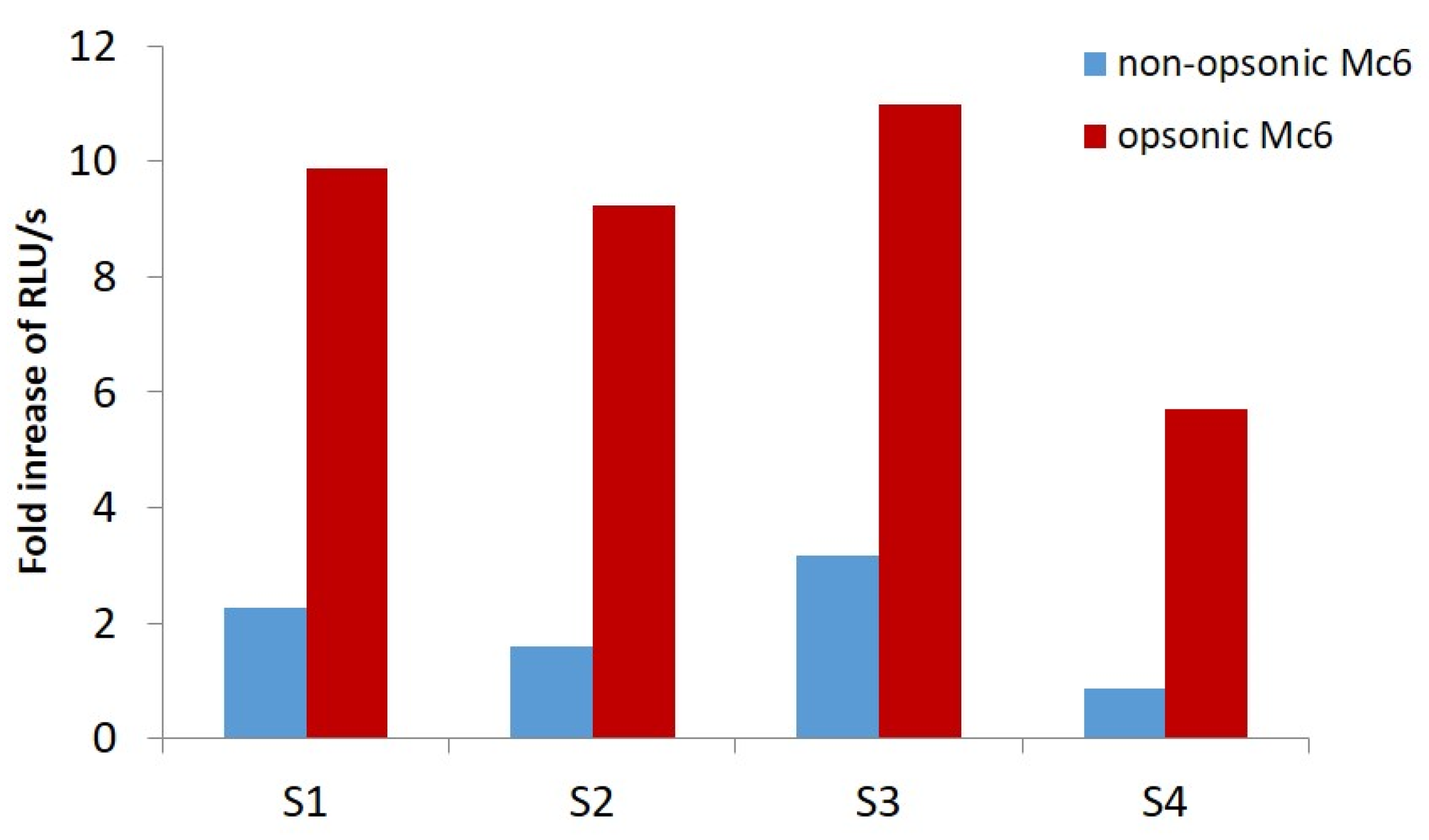
2.5. M. catarrhalis Is a Potent Inducer of Neutrophil Elastase Release
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Microbial Strains and Growth Condition
4.3. PMNs Isolation
4.4. PMNs Degranulation Assay
4.5. Neutrophil Elastase Activity Measurement
4.6. Flow Cytometry Analysis
4.7. In Gel Trypsin Digestion and Peptide Identification by LC MS/MS Analysis
4.8. Time-Kill Assay for WT and Isogenic Mutants
4.9. Bactericidal Activity of Serum Complement or Erythromycin against NE-Treated Bacteria
4.10. Complement Complex SC5b-9 Activation
4.11. Outer Membrane Vesicles Isolation
4.12. Outer Membrane Protein Isolation
4.13. TEM
4.14. Construction of Mc6 Isogenic Mutants
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Belaaouaj, A.; McCarthy, R.; Baumann, M.; Gao, Z.; Ley, T.J.; Abraham, S.N.; Shapiro, S.D. Mice lacking neutrophil elastase reveal impaired host defense against gram negative bacterial sepsis. Nat. Med. 1998, 4, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Belaaouaj, A.; Kim, K.S.; Shapiro, S.D. Degradation of outer membrane protein a in Escherichia coli killing by neutrophil elastase. Science 2000, 289, 1185–1187. [Google Scholar] [CrossRef]
- Hirche, T.O.; Benabid, R.; Deslee, G.; Gangloff, S.; Achilefu, S.; Guenounou, M.; Lebargy, F.; Hancock, R.E.; Belaaouaj, A. Neutrophil elastase mediates innate host protection against Pseudomonas aeruginosa. J. Immunol. 2008, 181, 4945–4954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benabid, R.; Wartelle, J.; Malleret, L.; Guyot, N.; Gangloff, S.; Lebargy, F.; Belaaouaj, A. Neutrophil elastase modulates cytokine expression: Contribution to host defense against Pseudomonas aeruginosa-induced pneumonia. J. Biol. Chem. 2012, 287, 34883–34894. [Google Scholar] [CrossRef] [Green Version]
- Scocchi, M.; Skerlavaj, B.; Romeo, D.; Gennaro, R. Proteolytic cleavage by neutrophil elastase converts inactive storage proforms to antibacterial bactenecins. Eur. J. Biochem. 1992, 209, 589–595. [Google Scholar] [CrossRef]
- Degasperi, M.; Sgarra, R.; Mardirossian, M.; Pacor, S.; Maschio, M.; Scocchi, M. Elastase-activated antimicrobial peptide for a safer pulmonary treatment of cystic fibrosis infections. Antibiotics 2022, 11, 319. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Wenzel, S.E.; Wu, Q.; Bowler, R.P.; Schnell, C.; Chu, H.W. Human neutrophil elastase degrades SPLUNC1 and impairs airway epithelial defense against bacteria. PLoS ONE 2013, 8, e64689. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, M.; Del Barrio, L.; Miller, M.A.; Re, F.; Weiss, D. Neutrophil elastase causes tissue damage that decreases host tolerance to lung infection with Burkholderia species. PLoS Pathog. 2014, 10, e1004327. [Google Scholar] [CrossRef] [Green Version]
- Le-Barillec, K.; Si-Tahar, M.; Balloy, V.; Chignard, M. Proteolysis of monocyte CD14 by human leukocyte elastase inhibits lipopolysaccharide-mediated cell activation. J. Clin. Investig. 1999, 103, 1039–1046. [Google Scholar] [CrossRef] [Green Version]
- William Vandivier, R.; Fadok, V.A.; Hoffmann, P.R.; Bratton, D.L.; Penvari, C.; Brown, K.K.; Brain, J.D.; Accurso, F.J.; Henson, P.M. Elastase-mediated phosphatidylserine receptor cleavage impairs apoptotic cell clearance in cystic fibrosis and bronchiectasis. J. Clin. Investig. 2002, 109, 661–670. [Google Scholar] [CrossRef]
- Chua, F.; Laurent, G.J. Neutrophil elastase: Mediator of extracellular matrix destruction and accumulation. Proc. Am. Thorac. Soc. 2006, 3, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Polańska, B.; Niemczuk, M.; Augustyniak, D.; Jankowski, A. Plasma neutrophil elastase in children with recurrent aphthous stomatitis. Cent. Eur. J. Immunol. 2006, 31, 15–17. [Google Scholar]
- Polańska, B.; Augustyniak, D.; Makulska, I.; Niemczuk, M.; Zwolińska, D.; Jankowski, A. Elastase, α1-proteinase inhibitor, and interleukin-8 in pre-dialyzed and hemodialyzed patients with chronic kidney disease. Pediatr. Int. 2010, 52, 735–743. [Google Scholar] [CrossRef]
- Jasper, A.E.; Mciver, W.J.; Sapey, E.; Walton, G.M. Understanding the role of neutrophils in chronic inflammatory airway disease. F1000Research 2019, 8, 557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero-Cervera, A.; Soehnlein, O.; Kenne, E. Neutrophils in chronic inflammatory diseases. Cell. Mol. Immunol. 2022, 19, 177–191. [Google Scholar] [CrossRef]
- Nadel, J.A. Role of neutrophil elastase in hypersecretion during copd exacerbations, and proposed therapies. Chest 2000, 117, 386S–389S. [Google Scholar] [CrossRef] [Green Version]
- Koga, H.; Miyahara, N.; Fuchimoto, Y.; Ikeda, G.; Waseda, K.; Ono, K.; Tanimoto, Y.; Kataoka, M.; Gelfand, E.W.; Tanimoto, M.; et al. Inhibition of neutrophil elastase attenuates airway hyperresponsiveness and inflammation in a mouse model of secondary allergen challenge: Neutrophil elastase inhibition attenuates allergic airway responses. Respir. Res. 2013, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Crisford, H.; Sapey, E.; Rogers, G.B.; Taylor, S.; Nagakumar, P.; Lokwani, R.; Simpson, J.L. Neutrophils in asthma: The good, the bad and the bacteria. Thorax 2021, 76, 835–844. [Google Scholar] [CrossRef]
- Tuipulotu, D.E.; Feng, S.; Pandey, A.; Zhao, A.; Ngo, C.; Mathur, A.; Lee, J.; Shen, C.; Fox, D.; Xue, Y.; et al. Immunity against Moraxella catarrhalis requires guanylate-binding proteins and caspase-11-NLRP3 inflammasomes. EMBO J. 2023, 42, 1–24. [Google Scholar] [CrossRef]
- Sandhaus, R.A.; Turino, G. Neutrophil elastase-mediated lung disease. COPD J. Chronic Obstr. Pulm. Dis. 2013, 10, 60–63. [Google Scholar] [CrossRef]
- Green, B.J.; Wiriyachaiporn, S.; Grainge, C.; Rogers, G.B.; Kehagia, V.; Lau, L.; Carroll, M.P.; Bruce, K.D.; Howarth, P.H. Potentially pathogenic airway bacteria and neutrophilic inflammation in treatment resistant severe asthma. PLoS ONE 2014, 9, e100645. [Google Scholar] [CrossRef]
- Augustyniak, D.; Majkowska-Skrobek, G.; Kraus-Filarska, M.; Jankowski, A. Effects of M. catarrhalis antigens on IFN-γ, IL-4 and IL-13 production by PBMC from children with atopic asthma or recurrent respiratory tract infections. Cent. Eur. J. Immunol. 2003, 28, 6–13. [Google Scholar]
- Bisgaard, H.; Hermansen, N.; Buchvald, F.; Loland, L.; Halkjaer, B.; Bønnelykke, K.; Brasholt, M.; Heltberg, A.; Vissing, H.; Thorsen, V.; et al. Childhood asthma after bacterial colonization of the airway in neonates. N. Engl. J. Med. 2007, 357, 1487–1495. [Google Scholar] [CrossRef]
- Manolov, T.; Thuan, T.T.; Forsgren, A.; Riesbeck, K. Moraxella-dependent α1-antichymotrypsin neutralization: A unique virulence mechanism. Am. J. Respir. Cell Mol. Biol. 2008, 38, 609–617. [Google Scholar] [CrossRef]
- Alnahas, S.; Hagner, S.; Raifer, H.; Kilic, A.; Gasteiger, G.; Mutters, R.; Hellhund, A.; Prinz, I.; Pinkenburg, O.; Visekruna, A.; et al. IL-17 and tnf-α are key mediators of Moraxella catarrhalis triggered exacerbation of allergic airway inflammation. Front. Immunol. 2017, 8, 1562. [Google Scholar] [CrossRef] [Green Version]
- Angrill, J.; Agustí, C.; De Celis, R.; Filella, X.; Rañó, A.; Elena, M.; De La Bellacasa, J.P.; Xaubet, A.; Torres, A. Bronchial inflammation and colonization in patients with clinically stable bronchiectasis. Am. J. Respir. Crit. Care Med. 2001, 164, 1628–1632. [Google Scholar] [CrossRef] [PubMed]
- Thulborn, S.J.; Mistry, V.; Brightling, C.E.; Moffitt, K.L.; Ribeiro, D.; Bafadhel, M. Neutrophil elastase as a biomarker for bacterial infection in copd. Respir. Res. 2019, 20, 170. [Google Scholar] [CrossRef] [Green Version]
- Augustyniak, D.; Roszkowiak, J.; Wiśniewska, I.; Skała, J.; Gorczyca, D.; Drulis-Kawa, Z. Neuropeptides SP and CGRP diminish the Moraxella catarrhalis outer membrane vesicle- (OMV-) triggered inflammatory response of human A549 epithelial cells and neutrophils. Mediators Inflamm. 2018, 2018, 4847205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luke, N.R.; Campagnari, A.A. Construction and characterization of Moraxella catarrhalis mutants defective in expression of transferrin receptors. Infect. Immun. 1999, 67, 5815–5819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holm, M.M.; Vanlerberg, S.L.; Foley, I.M.; Sledjeski, D.D.; Lafontaine, E.R. The Moraxella catarrhalis porin-like outer membrane protein CD is an adhesin for human lung cells. Infect. Immun. 2004, 72, 1906–1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipski, S.L.; Akimana, C.; Timpe, J.M.; Wooten, R.M.; Lafontaine, E.R. The Moraxella catarrhalis autotransporter McaP is a conserved surface protein that mediates adherence to human epithelial cells through its n-terminal passenger domain. Infect. Immun. 2007, 75, 314–324. [Google Scholar] [CrossRef] [Green Version]
- Timpe, J.M.; Holm, M.M.; Vanlerberg, S.L.; Basrur, V.; Lafontaine, E.R. Identification of a Moraxella catarrhalis outer membrane protein exhibiting both adhesin and lipolytic activities. Infect. Immun. 2003, 71, 4341–4350. [Google Scholar] [CrossRef] [Green Version]
- Golkar, T.; Zielinski, M.; Berghuis, A.M. Look and outlook on enzyme-mediated macrolide resistance. Front. Microbiol. 2018, 9, 1942. [Google Scholar] [CrossRef] [Green Version]
- Perez, A.C.; Murphy, T.F. A Moraxella catarrhalis vaccine to protect against otitis media and exacerbations of COPD: An update on current progress and challenges. Hum. Vaccines Immunother. 2017, 13, 2322–2331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockley, J.A.; Walton, G.M.; Lord, J.M.; Sapey, E. Aberrant neutrophil functions in stable chronic obstructive pulmonary disease: The neutrophil as an immunotherapeutic target. Int. Immunopharmacol. 2013, 17, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Dicker, A.J.; Crichton, M.L.; Pumphrey, E.G.; Cassidy, A.J.; Suarez-Cuartin, G.; Sibila, O.; Furrie, E.; Fong, C.J.; Ibrahim, W.; Brady, G.; et al. Neutrophil extracellular traps are associated with disease severity and microbiota diversity in patients with chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2018, 141, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Block, H.; Zarbock, A. A fragile balance: Does neutrophil extracellular trap formation drive pulmonary disease progression? Cells 2021, 10, 1932. [Google Scholar] [CrossRef] [PubMed]
- Mosca, T.; Menezes, M.C.; Silva, A.V.; Stirbulov, R.; Forte, W.C. Chemotactic and phagocytic activity of blood neutrophils in allergic asthma. Immunol. Investig. 2015, 44, 509–520. [Google Scholar] [CrossRef]
- Nicchi, S.; Giusti, F.; Carello, S.; Utrio Lanfaloni, S.; Tavarini, S.; Frigimelica, E.; Ferlenghi, I.; Rossi Paccani, S.; Merola, M.; Delany, I.; et al. Moraxella catarrhalis evades neutrophil oxidative stress responses providing a safer niche for nontypeable Haemophilus influenzae. iScience 2022, 25, 103931. [Google Scholar] [CrossRef]
- Murphy, T.F.; Kirkham, C.; Liu, D.F.; Sethi, S. Human immune response to outer membrane protein cd of Moraxella catarrhalis in adults with chronic obstructive pulmonary disease. Infect. Immun. 2003, 71, 1288–1294. [Google Scholar] [CrossRef] [Green Version]
- Murphy, T.F.; Brauer, A.L.; Aebi, C.; Sethi, S. Identification of surface antigens of Moraxella catarrhalis as targets of human serum antibody responses in chronic obstructive pulmonary disease. Infect. Immun. 2005, 73, 3471–3478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustyniak, D.; Mleczko, J.; Gutowicz, J. The immunogenicity of the liposome-associated outer membrane proteins (OMPs) of Moraxella catarrhalis. Cell. Mol. Biol. Lett. 2010, 15, 70–89. [Google Scholar] [CrossRef] [Green Version]
- Augustyniak, D.; Seredyński, R.; McClean, S.; Roszkowiak, J.; Roszniowski, B.; Smith, D.L.; Drulis-Kawa, Z.; MacKiewicz, P. Virulence factors of Moraxella catarrhalis outer membrane vesicles are major targets for cross-reactive antibodies and have adapted during evolution. Sci. Rep. 2018, 8, 4955. [Google Scholar] [CrossRef] [Green Version]
- Hare, K.M.; Seib, K.L.; Chang, A.B.; Harris, T.M.; Spargo, J.C.; Smith-Vaughan, H.C. Antimicrobial susceptibility and impact of macrolide antibiotics on Moraxella catarrhalis in the upper and lower airways of children with chronic endobronchial suppuration. J. Med. Microbiol. 2019, 68, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-l.; Ding, R.; Jia, X.-m.; Huang, J.-j.; Yu, S.; Chan, H.T.; Li, W.; Mao, L.-l.; Zhang, L.; Zhang, X.-y.; et al. Correlation of Moraxella catarrhalis macrolide susceptibility with the ability to adhere and invade human respiratory epithelial cells. Emerg. Microbes Infect. 2022, 11, 2055–2068. [Google Scholar] [CrossRef]
- Ozuna, H.; Uriarte, S.M.; Demuth, D.R. The hunger games: Aggregatibacter actinomycetemcomitans exploits human neutrophils as an epinephrine source for survival. Front. Immunol. 2021, 12, 3156. [Google Scholar] [CrossRef]
- Eichelberger, K.R.; Goldman, W.E. Manipulating neutrophil degranulation as a bacterial virulence strategy. PLoS Pathog. 2020, 16, e1009054. [Google Scholar] [CrossRef]
- Eilers, B.; Mayer-Scholl, A.; Walker, T.; Tang, C.; Weinrauch, Y.; Zychlinsky, A. Neutrophil antimicrobial proteins enhance Shigella flexneri adhesion and invasion. Cell. Microbiol. 2010, 12, 1134–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- du Teil Espina, M.; Fu, Y.; van der Horst, D.; Hirschfeld, C.; López-Álvarez, M.; Mulder, L.M.; Gscheider, C.; Haider Rubio, A.; Huitema, M.; Becher, D.; et al. Coating and corruption of human neutrophils by bacterial outer membrane vesicles. Microbiol. Spectr. 2022, 10, e00753-22. [Google Scholar] [CrossRef]
- Pandey, K.C.; De, S.; Mishra, P.K. Role of proteases in chronic obstructive pulmonary disease. Front. Pharmacol. 2017, 8, 512. [Google Scholar] [CrossRef]
- Standish, A.J.; Weiser, J.N. Human neutrophils kill Streptococcus pneumoniae via serine proteases. J. Immunol. 2009, 183, 2602–2609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.J.; Kang, H.K.; Park, E.; Kim, M.K.; Park, Y. Bactericidal activities and action mechanism of the novel antimicrobial peptide Hylin a1 and its analog peptides against Acinetobacter baumannii infection. Eur. J. Pharm. Sci. 2022, 175, 106205. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programsitle. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, R.; Matsuoka, S.; Fujinami, Y.; Nonaka, S.; Ichinose, S.; Kubota, T.; Okamura, N. Role of Moraxella catarrhalis outer membrane protein CD in bacterial cell morphology and autoaggregation. Res. Microbiol. 2013, 164, 236–243. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name and Description | MW 1 [kDa] | Sequence Coverage (%) | Score 2 | No. of Unique Peptides |
|---|---|---|---|---|
| TbpA (Transferrin binding protein A) | 119.3 | 61.45 | 163.57 | 53 |
| McaP (Moraxella catarrhalis adherence protein) | 71.3 | 70.15 | 71.45 | 28 |
| OMP CD (Outer membrane protein CD) | 48.3 | 56.95 | 236.58 | 29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roszkowiak, J.; McClean, S.; Mirończuk, A.M.; Augustyniak, D. The Direct Anti-Virulence but Not Bactericidal Activity of Human Neutrophil Elastase against Moraxella catarrhalis. Int. J. Mol. Sci. 2023, 24, 6607. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24076607
Roszkowiak J, McClean S, Mirończuk AM, Augustyniak D. The Direct Anti-Virulence but Not Bactericidal Activity of Human Neutrophil Elastase against Moraxella catarrhalis. International Journal of Molecular Sciences. 2023; 24(7):6607. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24076607
Chicago/Turabian StyleRoszkowiak, Justyna, Siobhán McClean, Aleksandra M. Mirończuk, and Daria Augustyniak. 2023. "The Direct Anti-Virulence but Not Bactericidal Activity of Human Neutrophil Elastase against Moraxella catarrhalis" International Journal of Molecular Sciences 24, no. 7: 6607. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24076607