

A Culturomics-Based Bacterial Synthetic Community for Improving Resilience towards Arsenic and Heavy Metals in the Nutraceutical Plant Mesembryanthemum crystallinum

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Determination of the Minimal Inhibitory Concentrations (MICs) for Arsenic and Metals and Selection of Bacterial Consortiums
2.2. Identification of Arsenic and Metal Resistance Traits in the Core Genome of SynComs
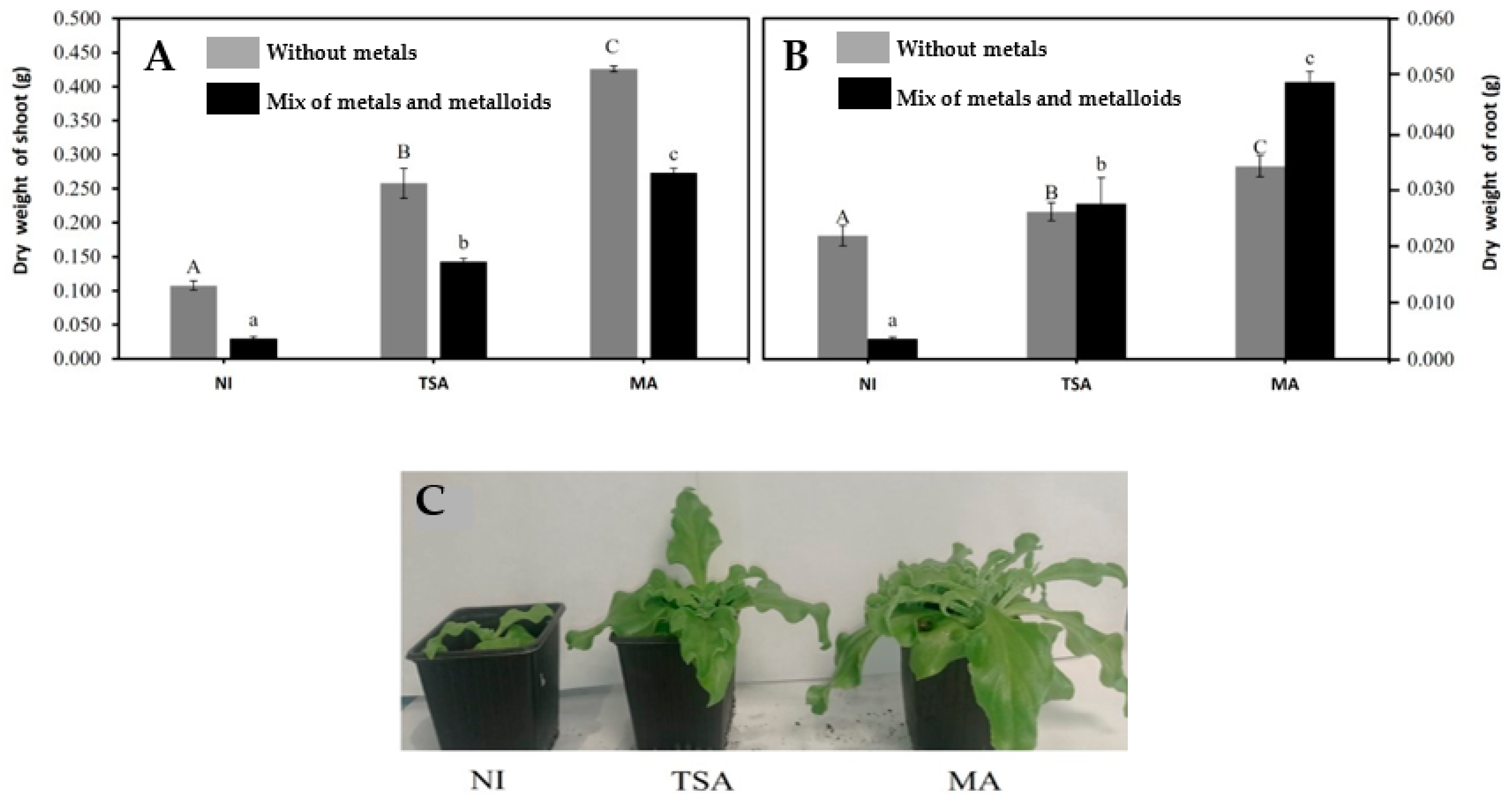
2.3. Effect of Inoculation on Plant Biomass
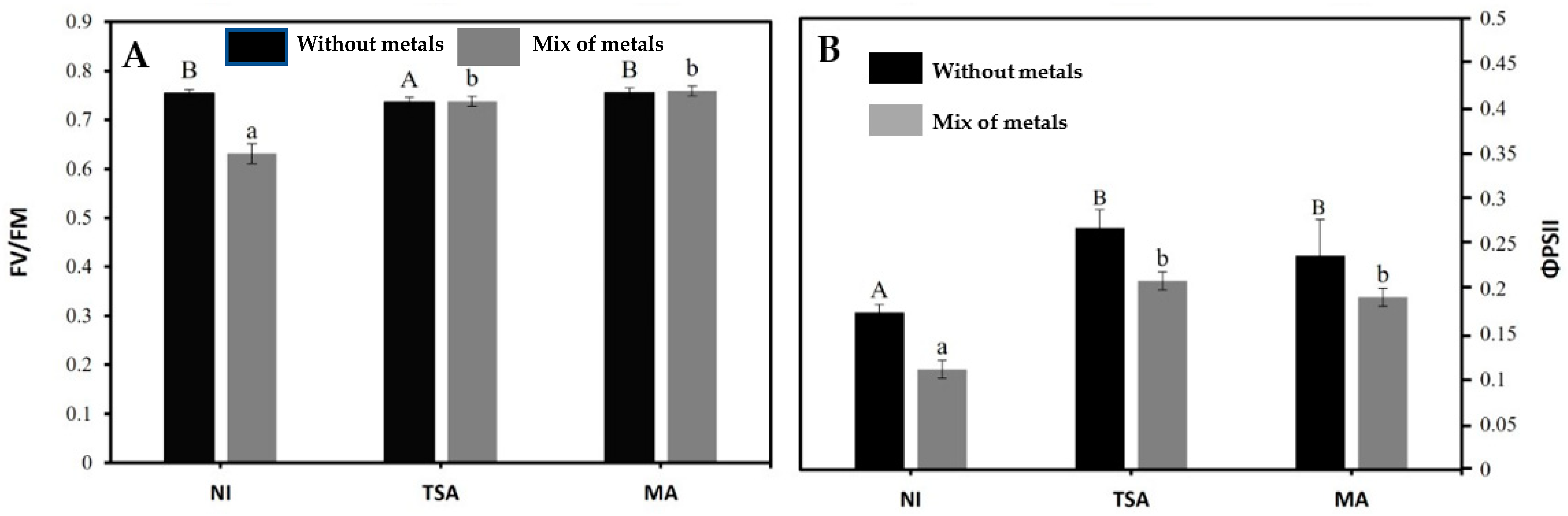
2.4. Effect of Inoculation on Plant Nutritional and Physiological Parameters
2.5. Accumulation of Metals in Plant Tissues
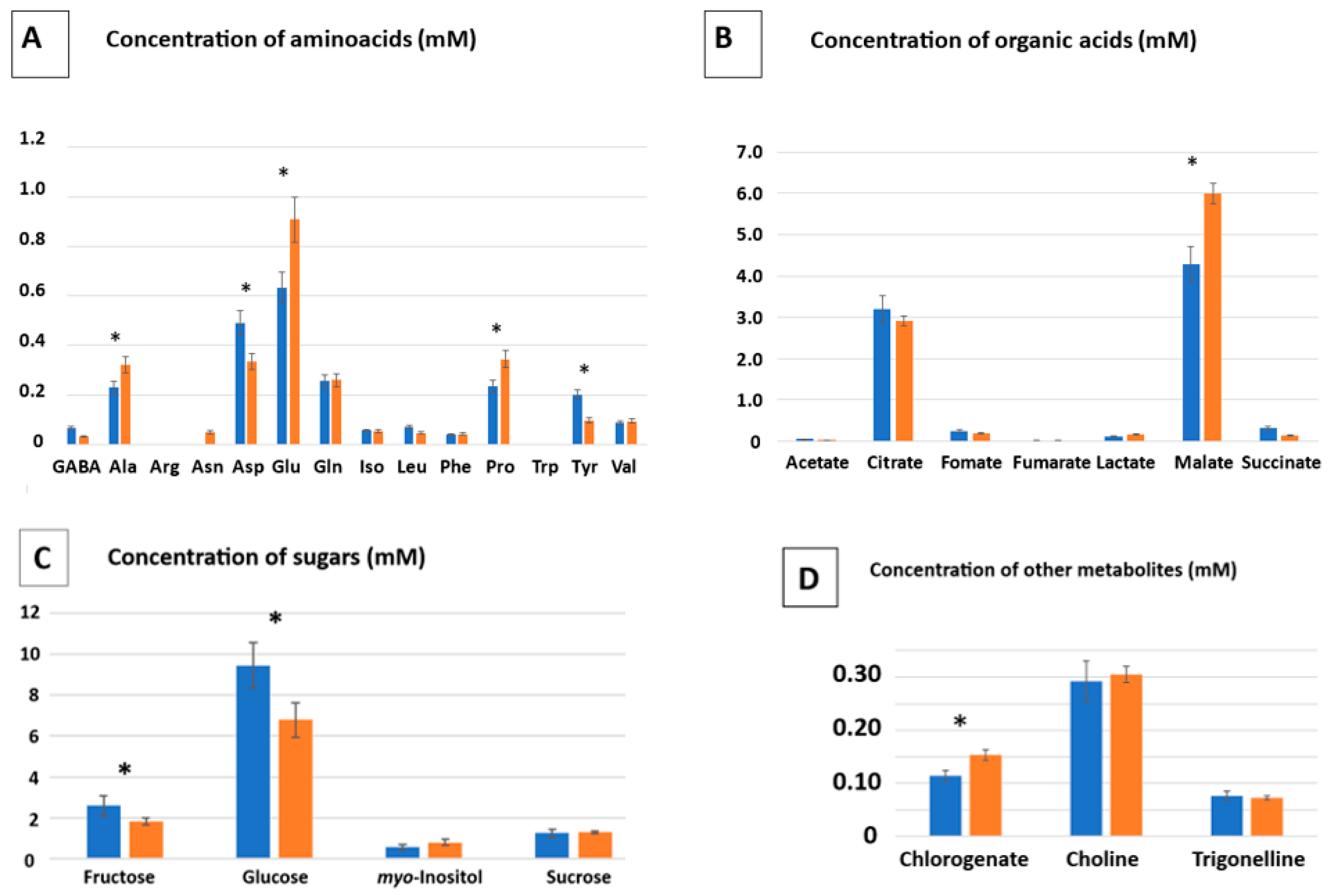
2.6. Metabolomics Analysis
2.7. Testing the Usefulness of the SynComs in a Crop Model Plant
3. Discussion
3.1. Deleterious Effects of Environmental Stresses on Mesembryanthemum crystallinum
3.2. Role of Metal-Resistant Biostumulants in Alleviating Plant Stress
3.3. Metal Resistance in the Selected Strains
3.4. Effect of Inoculation with SynComs on Growth, Physiology and Metal Accumulation by M. crystallinum
3.5. Improving the Accumulation of Desired Metabolites by Synergy Application of Metal Stress and Inoculation
3.6. Usefulness of the Selected Strains in a Crop Plant
4. Materials and Methods
4.1. Determination of Metal/Loid Resistance in Bacteria
4.2. Identification of Selected Bacteria by 16SrRNA Sequencing
4.3. Whole-Genome Sequencing of the Selected Strains and Identification of Metal Resistance Traits
4.4. Preparation of the Bacterial Consortium for Plant Inoculation
4.5. Cultivation of Plants in the Greenhouse
4.6. Determination of the Physiological Status of the Plants
4.7. Determination of Plant Growth Parameters
4.8. Determination of Macronutrients and Metal/Metaloid Accumulation in Plants
4.9. Metabolomic Analyses
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant Associated Rhizobacteria for Biocontrol and Plant Growth Enhancement. Front. Plant Sci. 2021, 12, 634796. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, R.; Akashi, M.; Taniguchi, T.; Kinos, Y.; Yaprak, A.E.; Turgay, O.C. Metabolomics Analyses Reveal Metabolites Affected by Plant Growth-Promoting Endophytic Bacteria in Roots of the Halophyte Mesembryanthemum crystallinum. Int. J. Mol. Sci. 2021, 22, 11813. [Google Scholar] [CrossRef]
- Bhupenchandra, I.; Chongtham, S.K.; Devi, E.L.; Ramesh, R.; Choudhary, A.K.; Salam, M.D.; Sahoo, M.R.; Bhutia, T.L.; Devi, S.H.; Thounaojam, A.S.; et al. Role of biostimulants in mitigating the effects of climate change on crop performance. Front. Plant Sci. 2022, 13, 967665. [Google Scholar] [CrossRef] [PubMed]
- Toju, H.; Peay, K.G.; Yamamichi, M.; Narisawa, K.; Hiruma, K.; Naito, K.; Fukuda, S.; Ushio, M.; Nakaoka, S.; Onoda, Y.; et al. Core microbiomes for sustainable agroecosystems. Nat. Plants 2018, 4, 247–257. [Google Scholar] [CrossRef]
- Pajuelo, E.; Carrasco, J.A.; Flores-Duarte, N.J.; Rodríguez-Llorente, I.D.; Mesa-Marín, J.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Navarro-Torre, S. Designing Tailored Bioinoculants for Sustainable Agrobiology in Multi-Stressed Environments. In Sustainable Agrobiology: Design and Development of Microbial Consortiums; Maheshwari, D.K., Dheeman, S., Eds.; Springer: Berlin/Heidelberg, Germany; Series Microorganisms for Sustainability; 2023; Volume 23, pp. 359–397. [Google Scholar] [CrossRef]
- Marín, O.; González, B.; Poupin, M.J. From Microbial Dynamics to Functionality in the Rhizosphere: A Systematic Review of the Opportunities with Synthetic Microbial Communities. Front. Plant Sci. 2021, 12, 650609. [Google Scholar] [CrossRef] [PubMed]
- Sarhan, M.S.; Hamza, M.A.; Youssef, H.H.; Patz, S.; Becker, M.; ElSawey, H.; Nemr, A.; Daana, H.-S.A.; Mourad, E.F.; Morsi, A.T.; et al. Culturomics of the plant prokaryotic microbiome and the dawn of plant-based culture media—A review. J. Adv. Res. 2019, 19, 15–27. [Google Scholar] [CrossRef]
- Lagier, J.C.; Khelaifia, S.; Alou, M.; Ndongo, S.; Niokhor, D.; Perrine, H.; Caputo, A.; Cadoret, F.; Traore, S.I.; Seck, E.H.; et al. Culture of previously uncultured members of the human gut microbiota by culturomics. Nat. Microbiol. 2016, 1, 16203. [Google Scholar] [CrossRef]
- Guan, Q.; Tan, B.; Kelley, T.M.; Tian, J.; Chen, S. Physiological Changes in Mesembryanthemum crystallinum During the C3 to CAM Transition Induced by Salt Stress. Front. Plant Sci. 2020, 11, 283. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.H.; Barkla, B.; Vera-Estrella, R.; Pantoja, O.; Lee, S.-Y.; Bohnert, H.J.; Dassanayake, M. Cell type-specific responses to salinity—The epidermal bladder cell transcriptome of Mesembryanthemum crystallinum. New Phytol. 2015, 207, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kim, H.M.; Kim, H.M.; Lee, H.R.; Jeong, B.R.; Lee, H.J.; Hwang, S.J. Growth and phytochemicals of ice plant (Mesembryanthemum crystallinum L.) as affected by various combined ratios of red and blue LEDs in a closed-type plant production system. J. Appl. Res. Med. Aromat. Plants 2021, 20, 100267. [Google Scholar] [CrossRef]
- Calvo, M.M.; Martín-Diana, A.B.; Rico, D.; López-Caballero, M.E.; Martínez-Álvarez, O. Antioxidant, Antihypertensive, Hypoglycaemic and Nootropic Activity of a Polyphenolic Extract from the Halophyte Ice Plant (Mesembryanthemum crystallinum). Foods 2022, 11, 1581. [Google Scholar] [CrossRef] [PubMed]
- Śliwa-Cebula, M.; Kaszycki, P.; Kaczmarczyk, A.; Nosek, M.; Lis-Krzyścin, A.; Miszalski, Z. The Common Ice Plant (Mesembryanthemum crystallinum L.)–Phytoremediation Potential for Cadmium and Chromate-Contaminated Soils. Plants 2020, 9, 1230. [Google Scholar] [CrossRef]
- Supel, P.; Śliwa-Cebula, M.; Miszalski, Z.; Kaszycki, P. Cadmium-Tolerant Rhizospheric Bacteria of the C3/CAM Intermediate Semi-Halophytic Common Ice Plant (Mesembryanthemum crystallinum L.) Grown in Contaminated Soils. Front. Plant Sci. 2022, 13, 820097. [Google Scholar] [CrossRef]
- Mahmood, A.; Amaya, R.; Turgay, O.C.; Yaprak, A.E.; Taniguchi, T.; Kataoka, R. High salt tolerant plant growth promoting rhizobacteria from the common ice-plant Mesembryanthemum crystallinum L. Rhizosphere 2019, 9, 10–17. [Google Scholar] [CrossRef]
- Pajuelo, E.; Flores-Duarte, N.F.; Navarro-Torre, S.; Mateos Naranjo, E.; Redondo-Gómez, S.; Rodríguez-Llorente, I.D. Culturomics and circular agronomy: Two sides of the same coin for the design of a tailored biofertilizer for the pharmaceuticals-producing halophyte Mesembryanthemum crystallinum. Submitted.
- Mendez, M.O.; Maier, R.M. Phytostabilization of Mine Tailings in Arid and Semiarid Environments—An Emerging Remediation Technology. Environ. Health Perspect. 2008, 116, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Kholodova, V.; Volkov, K.; Abdeyeva, A.; Kuznetsov, V. Water status in Mesembryanthemum crystallinum under heavy metal stress. Environ. Exp. Bot. 2011, 71, 382–389. [Google Scholar] [CrossRef]
- Bertani, I.; Zampieri, E.; Bez, C.; Volante, A.; Venturi, V.; Monaco, S. Isolation and Characterization of Pseudomonas chlororaphis Strain ST9; Rhizomicrobiota and in Planta Studies. Plants 2021, 10, 1466. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, W.; Jiang, Q.; Fei, Z.; Xiao, M. Genome analysis of plant growth-promoting rhizobacterium Pseudomonas chlororaphis subsp. Aurantiaca JD37 and insights from comparison of genomics with three Pseudomonas strains. Microbiol. Res. 2020, 237, 126483. [Google Scholar] [CrossRef]
- Fan, B.; Wang, C.; Song, X.; Ding, X.; Wu, L.; Wu, H.; Gao, X.; Borriss, R. Bacillus velezensis FZB42 in 2018: The Gram-Positive Model Strain for Plant Growth Promotion and Biocontrol. Front. Microbiol. 2018, 9, 2491. [Google Scholar] [CrossRef] [Green Version]
- Rabbee, M.F.; Ali, M.S.; Choi, J.; Hwang, B.S.; Jeong, S.C.; Baek, K.H. Bacillus velezensis: A Valuable Member of Bioactive Molecules within Plant Microbiomes. Molecules 2019, 24, 1046. [Google Scholar] [CrossRef] [Green Version]
- Pajuelo, E.; Arjona, S.; Rodríguez-Llorente, I.D.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Merchán, F.; Navarro-Torre, S. Coastal Ecosystems as Sources of Biofertilizers in Agriculture: From Genomics to Application in an Urban Orchard. Front. Mar. Sci. 2021, 8, 685076. [Google Scholar] [CrossRef]
- Silini-Chérif, H.; Silini, A.; Ghoul, M.; Yadav, S. Isolation and characterization of plant growth promoting traits of a rhizobacteria: Pantoea agglomerans lma2. Pak. J. Biol. Sci. 2012, 15, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Shariati, J.V.; Malboobi, M.A.; Tabrizi, Z.; Tavakol, E.; Owlia, P.; Safari, M. Comprehensive genomic analysis of a plant growth-promoting rhizobacterium Pantoea agglomerans strain P5. Sci. Rep. 2017, 7, 15610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luziatelli, F.; Gatti, L.; Ficca, A.G.; Medori, G.; Silvestri, C.; Melini, F.; Muleo, R.; Ruzzi, M. Metabolites Secreted by a Plant-Growth-Promoting Pantoea agglomerans Strain Improved Rooting of Pyrus communis L. cv Dar Gazi Cuttings. Front. Microbiol. 2020, 11, 539359. [Google Scholar] [CrossRef]
- Rezzonico, F.; Smits, T.H.; Montesinos, E.; Frey, J.E.; Duffy, B. Genotypic comparison of Pantoea agglomerans plant and clinical strains. BMC Microbiol. 2009, 9, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, A.; Tabassum, B.; Allah, E.F.A. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Blake, C.; Christensen, M.N.; Kovács, A. Molecular Aspects of Plant Growth Promotion and Protection by Bacillus subtilis. MPMI 2021, 34, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Altaf, A.R.; Teng, H.; Saleem, M.; Ahmad, H.R.; Adil, M.; Shahzad, K. Associative interplay of Pseudomonas gessardii BLP141 and pressmud ameliorated growth, physiology, yield, and Pb-toxicity in sunflower. Bioremed. J. 2021, 25, 178–188. [Google Scholar] [CrossRef]
- Biedendieck, R.; Knuuti, T.; Moore, S.J.; Dieter, J. The “beauty in the beast”—The multiple uses of Priestia megaterium in biotechnology. Appl. Microbiol. Biotechnol. 2021, 105, 5719–5737. [Google Scholar] [CrossRef] [PubMed]
- Eisenstein, M. Living factories of the future. Nature 2016, 531, 401–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartz, P.; Mattes, C.; Schad, M.; Bernhardt, R.; Hannemann, F. Expanding the promoter toolbox of Bacillus megaterium. J. Biotechnol. 2019, 294, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, U.; Chakraborty, B.; Basnet, M. Plant growth promotion and induction of resistance in Camellia sinensis by Bacillus megaterium. J. Basic Microbiol. 2006, 46, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Ge, J.; Li, Y.; He, S.; Zhong, J.; Liu, X.; Yu, X. Enhanced degradation of chlorpyrifos in rice (Oryza sativa L.) by five strains of endophytic bacteria and their plant growth promotional ability. Chemosphere 2017, 184, 505–513. [Google Scholar] [CrossRef]
- Hu, Z.Y.; Hua, W.; Zhang, L.; Deng, L.B.; Wang, X.F.; Liu, G.H.; Hao, W.-J.; Wang, H.-Z. Seed structure characteristics to form ultrahigh oil content in rapeseed. PLoS ONE 2013, 8, e62099. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Moreno, J.; García-Ortega, L.F.; Torres-Saucedo, L.; Rivas-Noriega, P.; Ramírez-Santoyo, R.M.; Sánchez-Calderón, L.; Quiroz-Serrano, I.N.; Vidales-Rodríguez, L.E. Bacillus megaterium HgT21: A Promising Metal Multiresistant Plant Growth-Promoting Bacteria for Soil Biorestoration. Microbiol. Spectrum 2022, 10, e00656-22. [Google Scholar] [CrossRef]
- Huang, H.; Wu, K.; Khan, A.; Jiang, Y.; Ling, Z.; Liu, P.; Chen, Y.; Tao, X.; Li, X. A novel Pseudomonas gessardii strain LZ-E simultaneously degrades naphthalene and reduces hexavalent chromium. Biores. Technol. 2016, 207, 370–378. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Li, X.; Liu, D.; Xu, C.; Ai, Y.; Sun, X.; Zhang, M.; Gao, Y.; Zhang, Y.; Yang, T.; et al. A Novel Pb-Resistant Bacillus subtilis Bacterium Isolate for Co-Biosorption of Hazardous Sb(III) and Pb(II): Thermodynamics and Application Strategy. Int. J. Environ. Res. Public Health 2018, 15, 702. [Google Scholar] [CrossRef] [Green Version]
- Nosek, M.; Kaczmarczyk, A.; Śliwa, M.; Jędrzejczyk, R.; Kornaś, A.; Supel, P.; Kaszycki, P.; Miszalski, Z. The response of a model C3/CAM intermediate semi-halophyte Mesembryanthemum crystallinum L. to elevated cadmium concentrations. J. Plant Physiol. 2019, 240, 153005. [Google Scholar] [CrossRef]
- Nosek, M.; Kaczmarczyk, A.; Edrzejczyk, R.J.J.; Supel, P.; Kaszycki, P.; Miszalski, Z. Expression of Genes Involved in Heavy Metal Trafficking in Plants Exposed to Salinity Stress and Elevated Cd Concentrations. Plants 2020, 9, 475. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Koh, D.J.Q.; Qin, L. LED spectral quality and NaCl salinity interact to affect growth, photosynthesis and phytochemical production of Mesembryanthemum crystallinum. Func. Plant Biol. 2022, 49, 483–495. [Google Scholar] [CrossRef]
- Lešková, A.; Giehl, R.F.H.; Hartmann, A.; Fargašová, A.; von Wirén, N. Heavy Metals Induce Iron Deficiency Responses at Different Hierarchic and Regulatory Levels. Plant Physiol. 2017, 174, 1648–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caracciolo, A.B.; Terenzi, V. Rhizosphere Microbial Communities and Heavy Metals. Microorganisms 2021, 9, 1462. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Dubey, R.S. Heavy Metal Uptake and Detoxification Mechanisms in Plants. Int. J. Agric. Res. 2006, 1, 122–141. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.K. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. South African Journal of Botany 2010, 76, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Berni, R.; Luyckx, M.; Xu, X.; Legay, S.; Sergeant, K.; Hausman, J.F.; Lutts, S.; Cai, G.; Guerriero, G. Reactive oxygen species and heavy metal stress in plants: Impact on the cell wall and secondary metabolism. Environ. Exp. Bot. 2019, 161, 98–106. [Google Scholar] [CrossRef]
- Paredes-Páliz, K.I.; Mateos-Naranjo, E.; Doukkali, B.; Caviedes, M.A.; Redondo-Gómez, S.; Rodríguez-Llorente, I.D.; Pajuelo, E. Modulation of Spartina densiflora plant growth and metal accumulation upon selective inoculation treatments: A comparison of gram negative and gram positive rhizobacteria. Mar. Pollut. Bull. 2017, 125, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Páliz, K.I.; Rodríguez-Vázquez, R.; Duarte, B.; Caviedes, M.A.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Caçador, M.I.; Rodríguez-Llorente, I.D.; Pajuelo, E. Investigating the mechanisms underlying phytoprotection by plant growth-promoting rhizobacteria in Spartina densiflora under metal stress. Plant Biol. 2018, 20, 497–506. [Google Scholar] [CrossRef]
- Mahmood, A.; Kataoka, R. Metabolite profiling reveals a complex response of plants to application of plant growth-promoting endophytic bacteria. Microbiol. Res. 2020, 234, 126421. [Google Scholar] [CrossRef]
- Said, A.A.E.; Ali, T.F.S.; Attia, E.Z.; Ahmed, A.S.F.; Shehata, A.H.; Abdelmohsen, U.R.; Fouad, M.A. Antidepressant potential of Mesembryanthemum cordifolium roots assisted by metabolomic analysis and virtual screening. Natural Product Research 2021, 35, 5493–5497. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yang, Y.; Jia, L.; Chen, H.; Wei, X. Zinc-induced oxidative damage, antioxidant enzyme response and proline metabolism in roots and leaves of wheat plants. Ecotoxicol. Environ. Saf. 2013, 89, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Hoque., M.D.A.; Okuma, E.; Banu, M.N.A.; Shimoishi, Y. Exogenous proline and glycine betaine increase antioxidant enzyme activities and confer tolerance to cadmium stress in cultured tobacco cells. J. Plant Physiol. 2009, 166, 1587–1597. [Google Scholar] [CrossRef]
- He, J.; Qin, L. Impacts of Reduced Nitrate Supply on Nitrogen Metabolism, Photosynthetic Light-Use Efficiency, and Nutritional Values of Edible Mesembryanthemum crystallinum. Front. Plant Sci. 2021, 12, 686910. [Google Scholar] [CrossRef] [PubMed]
- Doumett, S.; Fibbi, D.; Azzarello, E.; Mancuso, S.; Mugnai, S.; Petruzzelli, G.; Del Bubba, M. Influence of the Application Renewal of Glutamate and Tartrate on Cd, Cu, Pb and Zn Distribution Between Contaminated Soil and Paulownia Tomentosa in a Pilot-Scale Assisted Phytoremediation Study. Int. J. Phytorem. 2010, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Farid, M.; Farid, S.; Zubair, M.; Ghani, M.A.; Rizwan, M.; Ishaq, H.K.; Alkahtani, S.; Abdel-Daim, M.M.; Ali, S. Glutamic Acid-Assisted Phytomanagement of Chromium Contaminated Soil by Sunflower (Helianthus annuus L.): Morphophysiological and Biochemical Alterations. Front. Plant Sci. 2020, 11, 1297. [Google Scholar] [CrossRef]
- Flis, P.; Ouerdane, L.; Grillet, L.; Curie, C.; Mari, S.; Lobinski, R. Inventory of metal complexes circulating in plant fluids: A reliable method based on HPLC coupled with dual elemental and high-resolution molecular mass spectrometric detection. New Phytol. 2016, 211, 1129–1141. [Google Scholar] [CrossRef] [Green Version]
- Chia, J.-C. Phytochelatin Synthase in Heavy Metal Detoxification and Xenobiotic Metabolism. In Biodegradation Technology of Organic and Inorganic Pollutants; Ferreira Mendes, K., Nogueira de Sousa, R., Cabral Mielke, K., Eds.; IntechOpen: Sao Paulo, Brazil, 2021. [Google Scholar] [CrossRef]
- Rellán-Álvarez, R.; Giner-Martínez-Sierra, J.; Orduna, J.; Orera, I.; Rodríguez-Castrillón, J.Á.; García-Alonso, J.I.; Abadía, J.; Alvarez-Fernández, A. Iden-tification of a tri-iron (III), tri-citrate complex in the xylem sap of iron-deficient tomato resupplied with iron: New insights into plant iron long-distance transport. Plant Cell Physiol. 2010, 51, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Kısa, D.; Elmastaş, M.; Öztürk, L.; Kayır, Ö. Responses of the phenolic compounds of Zea mays under heavy metal stress. Appl. Biol. Chem. 2016, 59, 813–820. [Google Scholar] [CrossRef]
- Osmolovskaya, N.; Dung, V. V.; Kuchaeva, L. The role of organic acids in heavy metal tolerance in plants. Biol. Commun. 2018, 63, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. Shikimate and phenylalanine biosynthesis in the green lineage. Front. Plant Sci. 2013, 4, 62. [Google Scholar] [CrossRef] [Green Version]
- Al-Mushhin, A.A.M.; Qari, S.H.; Fakhr, M.A.; Alnusairi, G.S.H.; Alnusaire, T.S.; ALra-shidi, A.A.; Latef, A.A.H.A.; Ali, O.M.; Khan, A.A.; Soliman, M.-H. Exogenous Myo-Inositol Alleviates Salt Stress by Enhancing Antioxidants and Membrane Stability via the Upregulation of Stress Responsive Genes in Chenopodium quinoa L. Plants 2021, 10, 2416. [Google Scholar] [CrossRef]
- Chen, C.; Chen, K.; Su, T.; Zhang, B.; Li, G.; Pan, J.; Si, M. Myo-inositol-1-phosphate synthase (Ino-1) functions as a protection mechanism in Corynebacterium glutamicum under oxidative stress. MicrobiologyOpen 2019, 8, e721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Li, M.; Zhang, X.; Yang, Q.; Huang, B. Up-regulation of lipid metabolism and glycine betaine synthesis are associated with choline-induced salt tolerance in halophytic seashore paspalum. Plant Cell Environ. 2020, 43, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Summers, P.S.; Weretilnyk, E.A. Choline Synthesis in Spinach in Relation to Salt Stress. Plant Physiol. 1993, 103, 1269–1276. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Abbas, Z.; Seleiman, M.F.; Rizwan, M.; Yavaș, İ.; Alhammad, B.A.; Shami, A.; Hasanuzzaman, M.; Kalderis, D. Glycine Betaine Accumulation, Significance and Interests for Heavy Metal Tolerance in Plants. Plants 2020, 9, 896. [Google Scholar] [CrossRef]
- Wallace, T.C.; Blusztajn, J.K.; Caudill, M.A.; Klatt, K.C.; Natker, E.M.S.; Zeisel, S.H.; Zelman, K.M. Choline: The Underconsumed and Underappreciated Essential Nutrient. Nutrition Today 2018, 53, 240–253. [Google Scholar] [CrossRef]
- Moretti, A.; Paoletta, M.; Liguori, S.; Bertone, M.; Toro, G.; Iolascon, G. Choline: An Essential Nutrient for Skeletal Muscle. Nutrients 2020, 12, 2144. [Google Scholar] [CrossRef]
- Biswas, S.; Giri, S. Importance of choline as essential nutrient and its role in prevention of various toxicities. Prague Med. Rep. 2015, 116, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Kalinowska, M.; Sienkiewicz-Gromiuk, J.; Świderski, G.; Pietryczuk, A.; Cudowski, A.; Lewandowski, W. Zn(II) Complex of Plant Phenolic Chlorogenic Acid: Antioxidant, Antimicrobial and Structural Studies. Materials 2020, 13, 3745. [Google Scholar] [CrossRef]
- Stennert, A.; Maier, H.G. Trigonelline in coffee. II. Content of green, roasted and instant coffee. Z Lebensm Unters Forsch. 1994, 199, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chan, L.; Zhou, S. Trigonelline: A Plant Alkaloid with Therapeutic Potential for Diabetes and Central Nervous System Disease. Curr. Med. Chem. 2012, 19, 3523–3531. [Google Scholar] [CrossRef] [PubMed]
- Nandety, R.S.; Wen, J.; Mysore, K.S. Medicago truncatula resources to study legume biology and symbiotic nitrogen fixation. Fundam. Res. 2023, 3, 219–224. [Google Scholar] [CrossRef]
- Pajuelo, E.; Carrasco, J.A.; Romero, L.C.; Chamber, M.A.; Gotor, C. Evaluation of the Metal Phytoextraction Potential of Crop Legumes. Regulation of the Expression of O-Acetylserine (Thiol)Lyase under Metal Stress. Plant Biol. 2007, 9, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Raklami, A.; Oufdou, K.; Tahiri, A.I.; Mateos-Naranjo, E.; Navarro-Torre, S.; Rodríguez-Llorente, I.D.; Meddich, A.; Redondo-Gómez, S.; Pajuelo, E. Safe Cultivation of Medicago sativa in Metal-Polluted Soils from Semi-Arid Regions Assisted by Heat- and Metallo-Resistant PGPR. Microorganisms 2019, 7, 212. [Google Scholar] [CrossRef] [Green Version]
- Redondo-Gómez, S.; Mesa-Marín, J.; Pérez-Romero, J.A.; López-Jurado, J.; García-López, J.V.; Mariscal, V.; Molina-Heredia, F.P.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Flowers, T.J.; et al. Consortia of Plant-Growth-Promoting Rhizobacteria Isolated from Halophytes Improve Response of Eight Crops to Soil Salinization and Climate Change Conditions. Agronomy 2021, 11, 1609. [Google Scholar] [CrossRef]
- CLSI (Clinical Laboratory Standard Institute). M07 Methods for Dilution Antimicrobial Susceptibility Test for Bacteria That Grow Aerobically, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Srinivasan, R.; Karaoz, U.; Volegova, M.; MacKichan, J.; Kato-Maeda, M.; Miller, S.; Nadarajan, R.; Brodie, E.L.; Lynch, S.V. Use of 16S rRNA Gene for Identification of a Broad Range of Clinically Relevant Bacterial Pathogens. PLoS ONE 2015, 10, e0117617. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Chun, J.; Lee, J.H.; Jung, Y.; Kim, M.; Kim, S.; Kim, B.K.; Lim, Y.W. EzTaxon: A web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences. Int. J. Syst. Evol. Microbiol. 2007, 57(Pt10)(Pt10), 2259–2261. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997v2. [Google Scholar]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.J.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Libik-Konieczny, M.; Kozieradzka-Kiszkurno, M.; Michalec-Warzecha, Ż.; Miszalski, Z.; Bizan, J.; Konieczny, R. Influence of anti- and prooxidants on rhizogenesis from hypocotyls of Mesembryanthemum crystallinum L. cultured in vitro. Acta Physiol. Plant 39 2017, 39, 166. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Drakopoulou, S.; Orfanakis, E.; Karagiannaki, I.; Gaitis, F.; Skoulika, S.; Papaioannou, A.; Boukouvalas, G.; Petropoulos, G.; Katsoudas, V.; Kontzedaki, R.; et al. Comparative Evaluation of Different Targeted and Untargeted Analytical Approaches to Assess Greek Extra Virgin Olive Oil Quality and Authentication. Molecules 2022, 27, 1350. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | PGP Properties and Enzymatic Activities (1) | MIC (As) mg∙L−1 | MIC (Cd) mg∙L−1 | MIC (Cu) mg∙L−1 | MIC (Zn) mg∙L−1 |
|---|---|---|---|---|---|
| TSA medium | |||||
| S3 | Bio, N2-fix, Aux, Cel | 128 | 32 | 256 | 128 |
| R5 | N2-fix, Aux, Pect, Cel, Amy, DNAase, Prot | 128 | 2 | 128 | 64 |
| H4 | Bio, N2-fix, Amy | 16 | 16 | 256 | 64 |
| MA medium | |||||
| MS2 | Sid, Aux, Prot, DNAase, Amy, Pect | 512 | 2 | 128 | 32 |
| MR4 | Bio, N2-fix, Prot, Cel, Amy, Pect, DNAase | 128 | 0.5 | 256 | 128 |
| MH8 | KS, PS, Sid, Bio, Pect, Amy | 32 | 32 | 256 | 64 |
| Strain | Metal Resistance Traits | Resistance |
|---|---|---|
| S3 | Uncharacterized protein Yfg | As |
| Exported zinc metalloprotease YfgC precursor | Zn | |
| Zinc transport protein ZntB | Zn | |
| Zinc transporter ZitB | Zn | |
| Copper resistance proteins CopCD | Co | |
| Lead, cadmium, zinc, and mercury-transporting ATPase (2 copies) | Pb, Cd, Zn, Hg | |
| R5 | Cobalt/zinc/cadmium-resistance protein CzcD | Co, Zn |
| Putative metallopeptidase (Zinc) SprT family | Zn | |
| Cadmium, zinc, and cobalt-transporting P-type ATPase | Cd, Zn, Co | |
| Lead, cadmium, zinc, and mercury-transporting ATPase (4 copies) | Pb, Cd, Zn, Hg | |
| Copper resistance transcriptional regulator CueR (MerR family) | Cu | |
| Arsenite/antimonite:H+ antiporter ArsB | As, Sb | |
| Arsenical-resistance operon repressor (2 copies) | As | |
| H4 | Cadmium-translocating P-type ATPase | Pb, Cd, Zn, Hg |
| Arsenical-resistance operon repressor | As | |
| Arsenical-resistance protein Acr3 | As | |
| Arsenic-resistance protein ArsH (2 copies) | As | |
| Copper(I) chaperone CopZ | Cu | |
| Lead, cadmium, zinc, and mercury-transporting ATPase (2 copies) | Pb, Cd, Zn, Hg | |
| Copper-sensing two-component system response regulator CpxRCusSR | Cu | |
| Cobalt-zinc-cadmium-resistance protein CzcD (2 copies) | Co, Zn, Cd | |
| Copper-resistance operon CopABCD, 1 extra copy of CopA | Cu | |
| Flavin-dependent monooxygenase ArsO | As | |
| MS2 | Arsenical-resistance operon repressor (5 copies) | As |
| Arsenical-resistance protein Acr3 | As | |
| Zinc ABC transporter operon ZnuBC | Zn | |
| Zinc uptake regulation protein Zur | Zn | |
| Lead, cadmium, zinc, and mercury-transporting ATPase (4 copies) | Pb, Cd, Zn, Hg | |
| Zinc transporter, ZIP family | Zn | |
| Cobalt-zinc-cadmium-resistance protein CzcD (3 copies) | Co, Zn, Cd | |
| Cytoplasmic copper homeostasis protein CutC (2 copies) | Cu | |
| Copper-resistance operon CopACDZ (1 extra copy of copZ) | Cu | |
| Response regulator of zinc sigma-54-dependent two-component system | Zn | |
| Magnesium and cobalt transport protein CorA | Mg, Co | |
| MR4 | Arsenical-resistance protein Acr3 | As |
| Arsenical-resistance operon repressor | As | |
| Cobalt/zinc/cadmium-resistance protein CzcD (4 copies) | Co, Zn, Cd | |
| Cadmium, zinc, and cobalt-transporting P-type ATPase | Cd, Zn, Co | |
| Lead, cadmium, zinc, and mercury-transporting ATPase (2 copies) | Pb, Cd, Zn, Hg | |
| Copper-resistance transcriptional regulator CueR (MerR family) | Cu | |
| Copper-resistance proteins CopABCD | Cu | |
| MH8 | Copper tolerance protein | Cu |
| Copper-sensing two-component system response regulator CusSR | Cu | |
| Cadmium-translocating P-type ATPase | Cd | |
| Arsenical-resistance operon repressor | As | |
| Arsenical-resistance protein Acr3 | As | |
| Arsenic-resistance protein ArsH (2 copies) | As | |
| Copper-resistance operon copABCD | Cu | |
| Cobalt/zinc/cadmium-resistance protein CzcD (2 copies) | Co, Zn, Cd | |
| Lead, cadmium, zinc, and mercury-transporting ATPase (2 copies) | Pb, Cd, Zn, Hg | |
| Flavin-dependent monooxygenase ArsO | As |
| Culture Conditions | P Content (%) | N Content (%) | Ca Content (%) | Mg Content (%) | K Content (%) | Na Content (%) | Fe Content (mg∙Kg−1) |
|---|---|---|---|---|---|---|---|
| Without metals (control) | 1.47 ± 0.03 | 0.88 ± 0.30 | 1.61 ± 0.02 | 0.73 ± 0.01 | 7.40 ± 0.20 | 2.30 ± 0.07 | 75.09 ± 2.74 |
| In the presence of metal(loid)s | 1.68 ± 0.01 ** | 0.91 ± 0.00 | 1.67 ± 0.00 * | 0.81 ± 0.01 ** | 9.00 ± 0.10 * | 2.60 ± 0.05 * | 66.60 ± 0.97 ** |
| Plant Tissue | Inoculation Treatment | As (mg∙Kg −1) | Cd (mg∙Kg −1) | Cu (mg∙Kg −1) | Zn (mg∙Kg −1) |
|---|---|---|---|---|---|
| SHOOTS | Non inoculated | ND | ND | 2.08 ± 0.05 | 3.07 ± 0.04 |
| TSA | ND | ND | 1.23 ± 0.07 * | 5.15 ± 0.24 * | |
| MA | ND | ND | 3.12 ± 0.16 | 10.92 ± 0.12 * | |
| ROOTS | Non inoculated | ND | 0.71 ± 0.00 | 6.89 ± 0.10 | 134.07 ± 0 |
| TSA | 4.09 ± 0.27 * | 1.08 ± 0.12 * | 8.91 ± 0.48 | 98.36 ± 3.70 * | |
| MA | 2.70 ± 0.14 * | 1.17 ± 0.02 * | 8.35 ± 0.38 | 104.99 ± 1.30 * | |
| (1) Thresholds for plant leaf tissue toxicity limits (mg∙Kg−1) | 5–20 | 5–30 | 2–20 | 100–400 | |
| TSA SynCOM | MA SynCom | |
|---|---|---|
| Aminoacids | ||
| GABA | -- | 84 |
| Alanine | 87 | 89 |
| Asparagine | 104 | --- |
| Aspartate | 100 | 102 |
| Glutamate | 80 | 75 |
| Glutamine | 123 | 100 |
| Isoleucine | 82 | 88 |
| Leucine | 43 (*) | 56 (*) |
| Phenylalanine | 56 (*) | 54 (*) |
| Proline | 48 (*) | 92 |
| Tyrosine | 71 | 82 |
| Valine | 80 | 90 |
| Organic acids | ||
| Acetate | 161 (*) | 105 |
| Citrate | 163 (*) | 251 (*) |
| Formate | 165 (*) | 152 (*) |
| Fumarate | 187 (*) | 155 (*) |
| Lactate | 72 | 100 |
| Malate | 141 (*) | 138 (*) |
| Succinate | 260 (*) | 153 (*) |
| Sugars | ||
| Fructose | 203 (*) | 141 |
| Glucose | 175 (*) | 138 |
| myo-inostol | 35 (*) | 106 |
| Sucrose | 112 | 328 (*) |
| Other metabolites | ||
| Choline | 136 | 155 (*) |
| Trigonelline | 153 (*) | 125 |
| Metals | Parameter | Non-Inoculated | TSA | MA |
|---|---|---|---|---|
| Absence of metals | Shoot dry matter (g) | 0.105 ± 0.10 (a) | 0.125 ± 0.03 (b) | 0.130 ± 0.09 (c) |
| Root dry matter (g) | 0.06 ± 0.006 (a) | 0.07± 0.006 (a,b) | 0.085± 0.006 (b) | |
| Size of leaves (cm) | 0.67 ± 0 (a) | 0.82 ± 0.11 (b) | 0.79 ± 0.02 (b) | |
| Number of leaves | 12 ± 0 (a) | 15 ± 0.5 (b) | 18 ± 0 (c) | |
| Maximum quantum efficiency of PSII | 0.75 ± 0.01 (a) | 0.76± 0.02 (a) | 0.72± 0.02 (a) | |
| Quantum efficiency of PSII | 0.34 ± 0.03 (a) | 0.37 ± 0.04 (a) | 0.35 ± 0.04 (a) | |
| Net photosynthesis rate (AN) (µM m−2 s−1) | 12.32 ± 1.10 (a) | 13.77 ± 0.80 (a) | 13.18 ± 1.90 (a) | |
| Electron transport rate | 74 ± 1 (a) | 115 ± 14 (b) | 102 ± 18 (b) | |
| Stomatal conductance (gs)(mM m−2 s−1) | 390 ± 37 (a) | 320 ± 30 (b) | 315 ± 20 (b) | |
| Presence of a mix of metals: 10 µM As + 1 µM Cd + 20 µM Cu + 50 µM Zn | Shoot dry matter (g) | 0.016 ± 0.004 (a) | 0.033 ± 0.003 (b) | 0.029 ± 0.002 (b) |
| Root dry matter (g) | 0.007 ± 0.005 (a) | 0.030 ± 0.004 (b) | 0.019 ± 0.004 (c) | |
| Size of leaves (cm) | 0.48 ± 0.02 (a) | 0.62 ± 0.02 (b) | 0.70 ± 0.04 (b) | |
| Number of leaves | 5.0 ± 0.5 (a) | 5.0 ± 0.5 (a) | 6.4 ± 1.0 (b) | |
| Maximum quantum efficiency of PSII | 0.70 ± 0.01 (a) | 0.76 ± 0.01 (c) | 0.73 ± 0.02 (b) | |
| Quantum efficiency of PSII | 0.29 ± 0.03 (a) | 0.32 ± 0.03 (a) | 0.34 ± 0.03 (a) | |
| Net photosynthesis rate (AN) (µM m−2 s−1) | 42 ± 9 (a) | 81 ± 12 (b) | 80 ± 10 (b) | |
| Electron transport rate | 4.8 ± 0.4 (a) | 9.3 ± 0.15 (b) | 10.9 ± 0.4 (c) | |
| Stomatal conductance (gs)(mM m−2 s−1) | 125 ± 35 (a) | 120 ± 34 (a) | 212 ± 25 (b) | |
| Content of metals in SHOOTS (mg∙Kg−1) | ||||
| As | 15.4 ± 0.17 (a) | 18.2 ± 0.15 (b) | 31.1 ± 0.17 (c) | |
| Cd | 0.3 ± 0.17 (a) | 1.0 ± 0.02 (b) | 0.8 ± 0.01 (a) | |
| Cu | 5.2 ± 0.23 (a) | 5.9 ± 0.21 (a) | 11.0 ± 0.32 (b) | |
| Zn | 28.2 ± 1.1 (a) | 75.2 ± 0.8 (b) | 77.7 ± 1.7 (b) | |
| Content of metals in ROOTS (mg∙Kg−1) | ||||
| As | 31.0 ± 1.4 (a) | 87.7 ± 2.2 (c) | 61.7 ± 2.2 (b) | |
| Cd | 2.0 ± 0.06 (a) | 3.8 ± 0.03 (c) | 1.4 ± 0.01 (a) | |
| Cu | 23.9 ± 0.67 (a) | 31.2 ± 1.11 (c) | 25.0 ± 0.76 (b) | |
| Zn | 149.5 ± 5.0 (a) | 197.1 ± 7.2 (c) | 94.8 ± 2.4 (a) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores-Duarte, N.J.; Pajuelo, E.; Mateos-Naranjo, E.; Navarro-Torre, S.; Rodríguez-Llorente, I.D.; Redondo-Gómez, S.; Carrasco López, J.A. A Culturomics-Based Bacterial Synthetic Community for Improving Resilience towards Arsenic and Heavy Metals in the Nutraceutical Plant Mesembryanthemum crystallinum. Int. J. Mol. Sci. 2023, 24, 7003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24087003
Flores-Duarte NJ, Pajuelo E, Mateos-Naranjo E, Navarro-Torre S, Rodríguez-Llorente ID, Redondo-Gómez S, Carrasco López JA. A Culturomics-Based Bacterial Synthetic Community for Improving Resilience towards Arsenic and Heavy Metals in the Nutraceutical Plant Mesembryanthemum crystallinum. International Journal of Molecular Sciences. 2023; 24(8):7003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24087003
Chicago/Turabian StyleFlores-Duarte, Noris J., Eloísa Pajuelo, Enrique Mateos-Naranjo, Salvadora Navarro-Torre, Ignacio D. Rodríguez-Llorente, Susana Redondo-Gómez, and José A. Carrasco López. 2023. "A Culturomics-Based Bacterial Synthetic Community for Improving Resilience towards Arsenic and Heavy Metals in the Nutraceutical Plant Mesembryanthemum crystallinum" International Journal of Molecular Sciences 24, no. 8: 7003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24087003