Music with Different Tones Affects the Development of Brain Nerves in Mice in Early Life through BDNF and Its Downstream Pathways

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of Music in Different Tones on Nissl Bodies in the Hippocampus and Prefrontal Cortex
2.2. Effects of Music in Different Tones on Neuronal Dendritic Development in the Hippocampus and Prefrontal Cortex
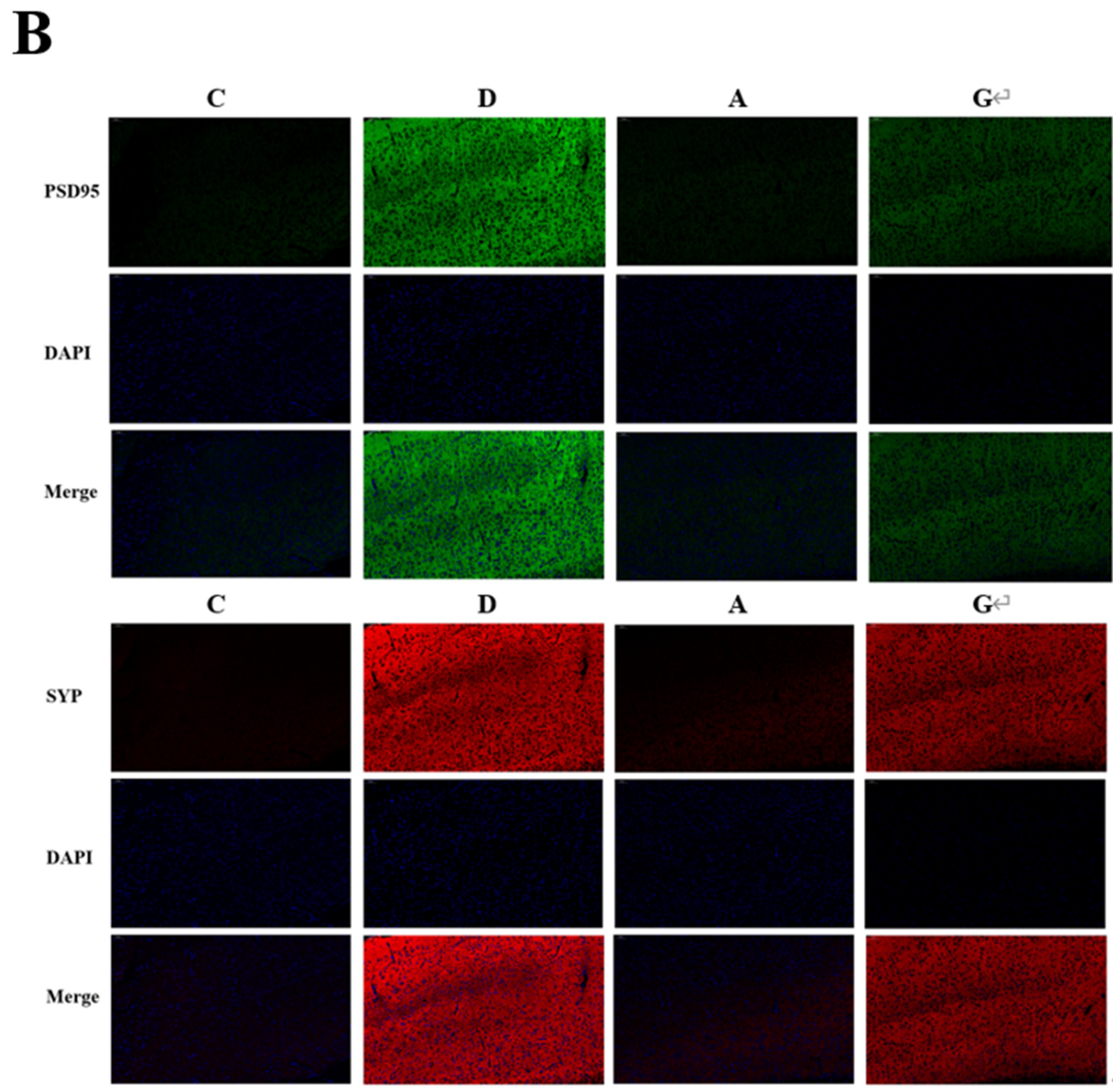
2.3. Effects of Music in Different Tones on the Expression of Synaptic Protein in the Hippocampus and Prefrontal Cortex
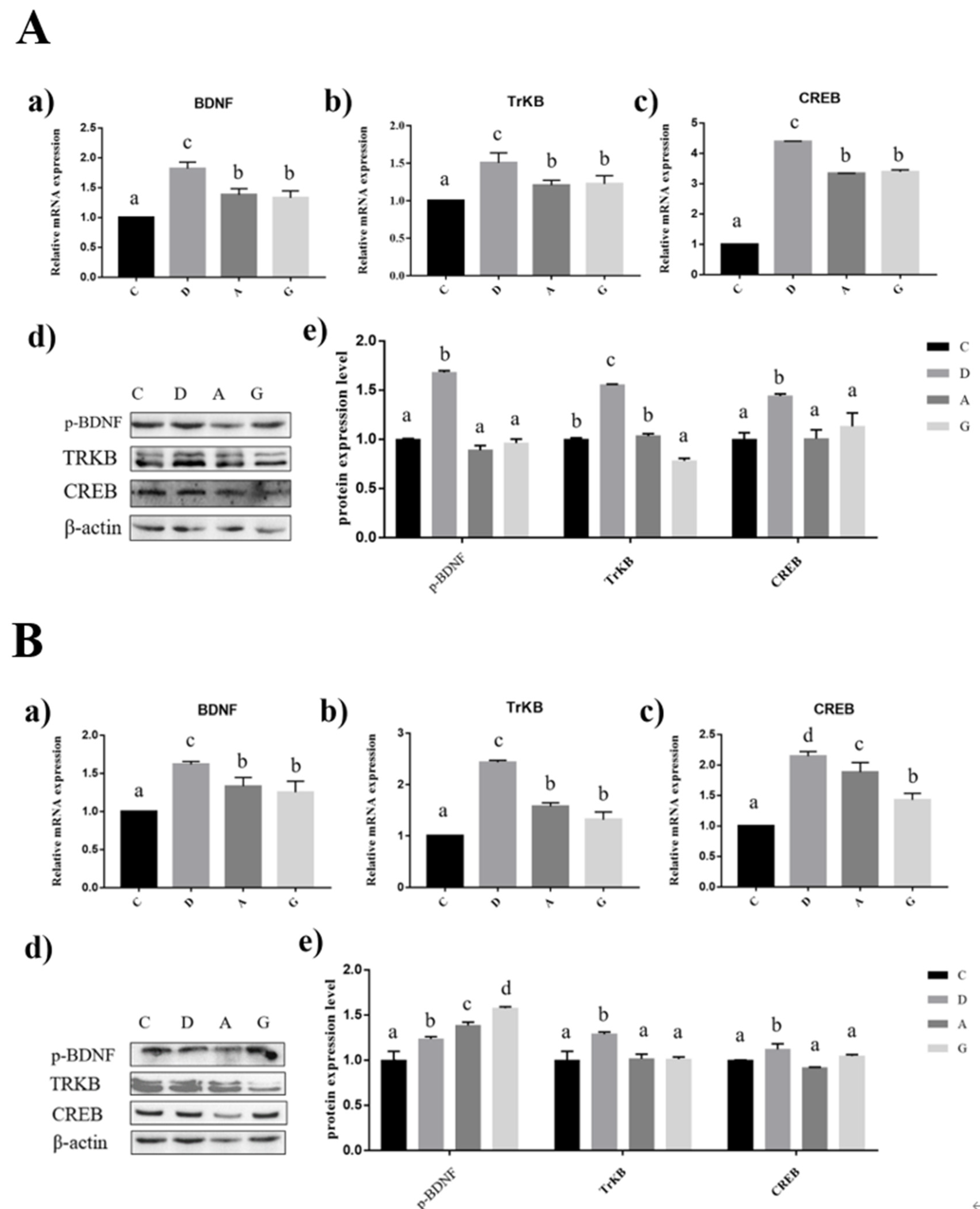
2.4. Effects of Music in Different Tones on mRNA and Protein Expression Levels of BDNF/TRKB/CREB Pathways in the Hippocampus and Prefrontal Cortex
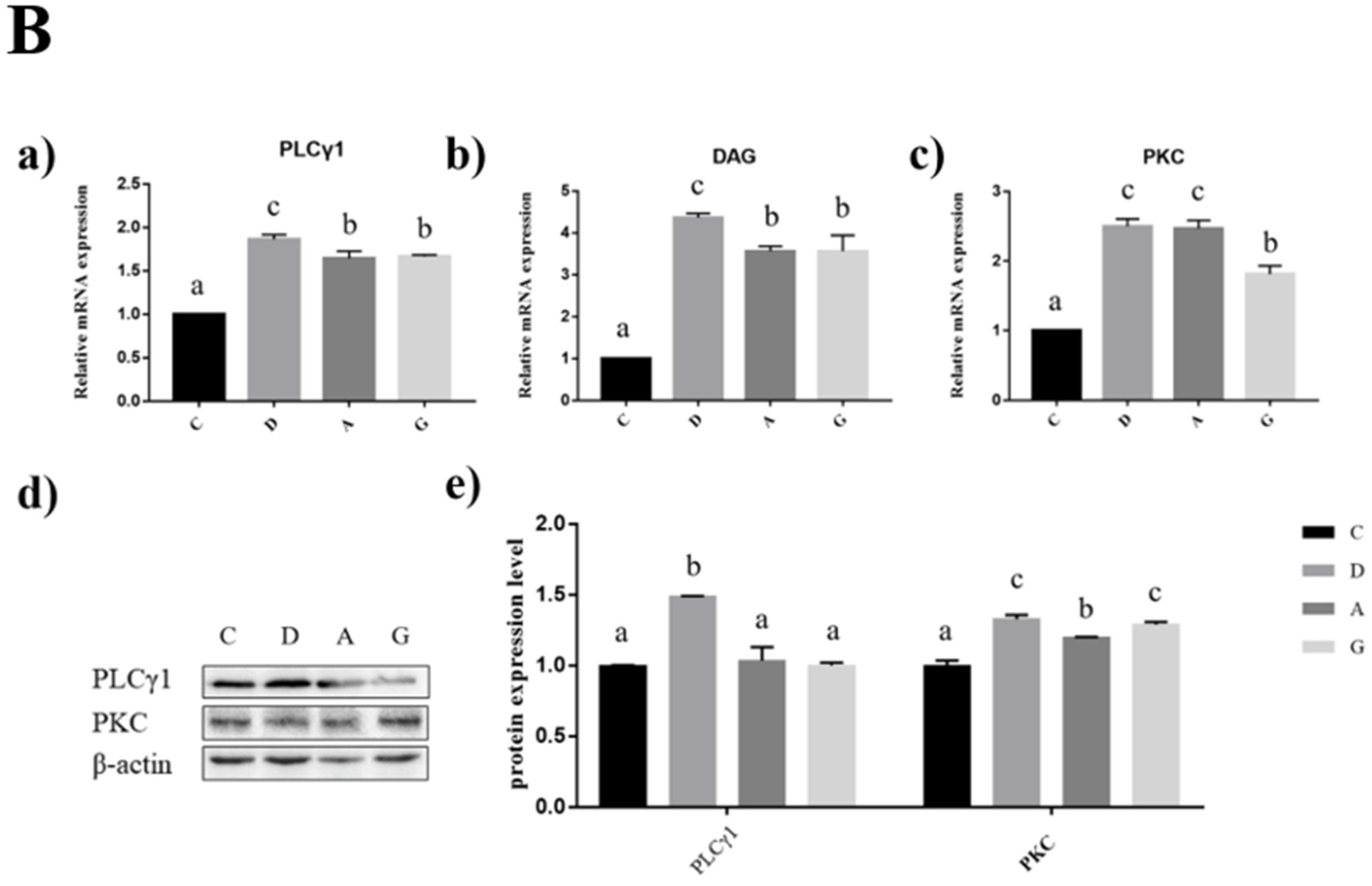
2.5. Effects of Music in Different Tones on mRNA and Protein Expression Levels of PLCγ1/PKC Pathways in the Hippocampus and Frontal Cortex
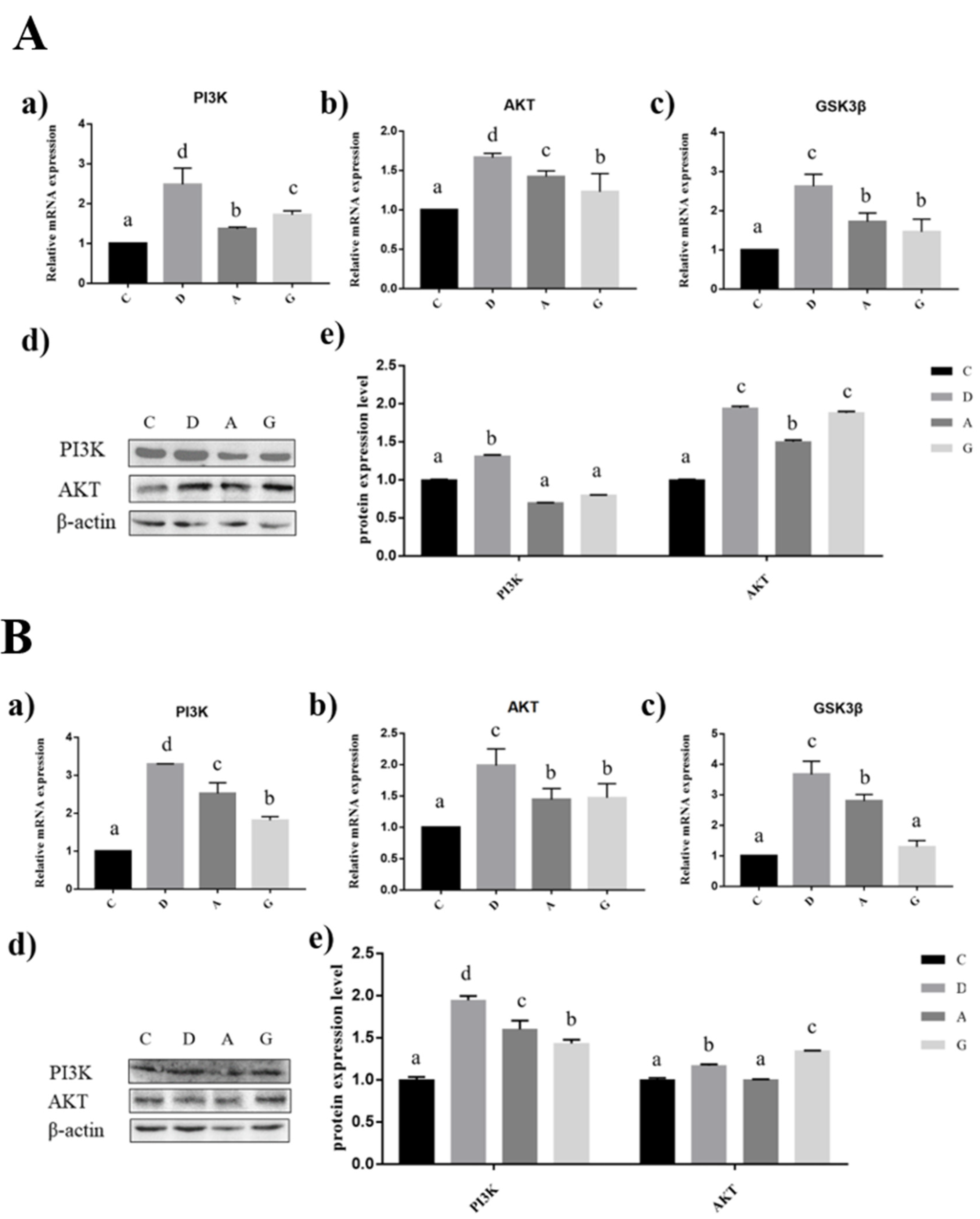
2.6. Effects of Music in Different Tones on the mRNA and Protein Expression Levels of PI3K/AKT Pathways in the Hippocampus and Prefrontal Cortex
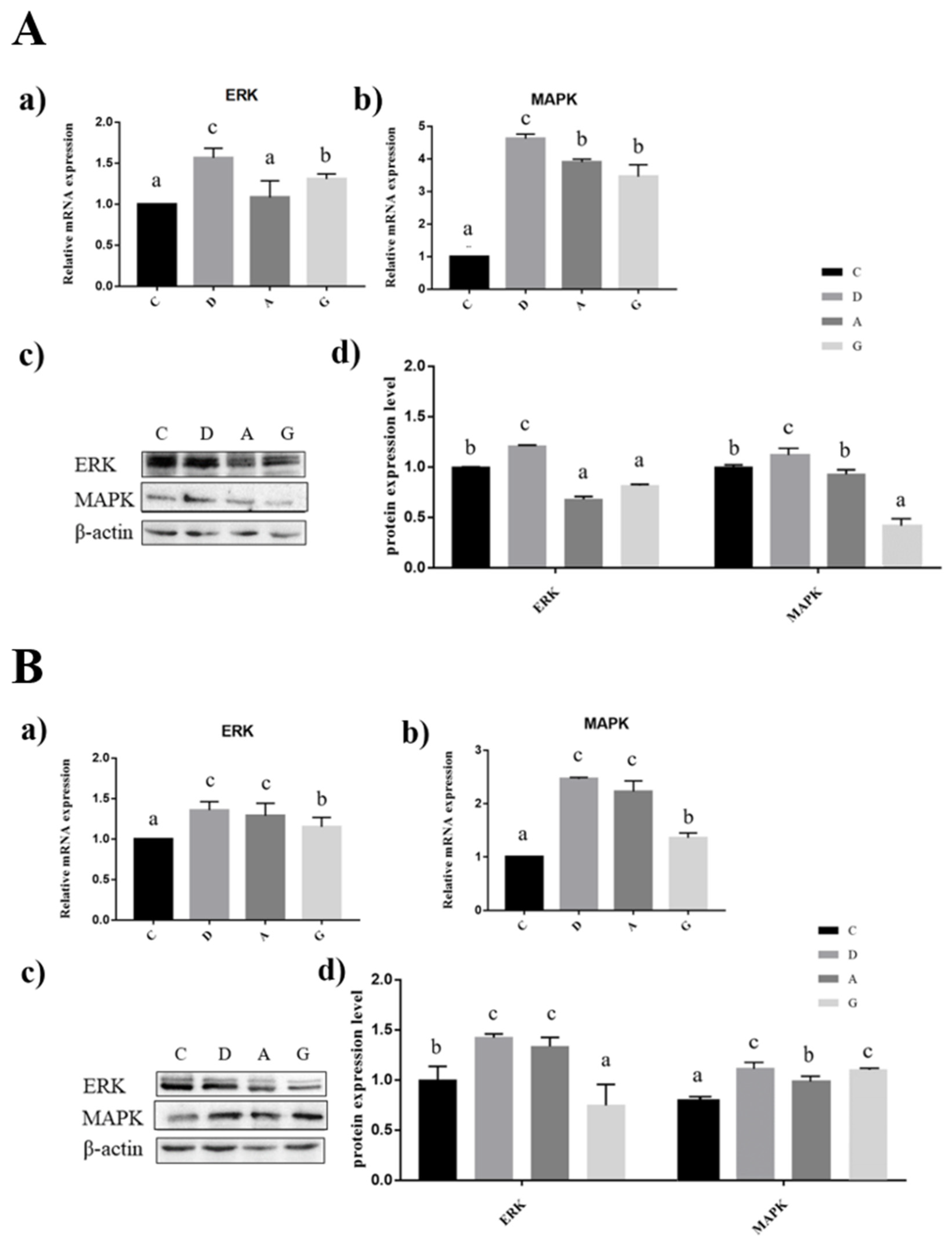
2.7. Effects of Music in Different Tones on the mRNA and Protein Expression Levels of MAPK/ERK Pathway in the Hippocampus and Frontal Cortex
3. Discussion
4. Materials and Methods
4.1. Feeding and Management of Experimental Animals
4.2. Grouping for Experimental Animals
4.3. Sample Collection
4.4. Nissl Staining
4.5. Golgi Staining
4.6. Immunofluorescence
4.7. Real-Time Quantitative PCR (qRT-PCR)
4.8. Western Blot
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kempermann, G. Environmental enrichment, new neurons and the neurobiology of individuality. Nat. Rev. Neurosci. 2019, 20, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Mooney-Leber, S.M.; Brummelte, S. Neonatal pain and reduced maternal care: Early-life stressors interacting to impact brain and behavioral development. Neuroscience 2017, 342, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, M.B.; Shalev, N.; Ramduny, J.; Sotiropoulos, S.N.; Chechlacz, M. Right fronto-parietal networks mediate the neurocognitive benefits of enriched environments. Brain Commun. 2022, 4, fcac080. [Google Scholar] [CrossRef] [PubMed]
- Steele, C.J.; Bailey, J.A.; Zatorre, R.J.; Penhune, V.B. Early Musical Training and White-Matter Plasticity in the Corpus Callosum: Evidence for a Sensitive Period. J. Neurosci. 2013, 33, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.M.; Li, W.H.; Ma, B.; Deng, X.; Zhang, L.; Zhao, L.; Zheng, T.J.; Jing, Y.H. Experiences affect social behaviors via altering neuronal morphology and oxytocin system. Psychoneuroendocrinology 2021, 129, 105247. [Google Scholar] [CrossRef]
- Schloesser, R.J.; Lehmann, M.; Martinowich, K.; Manji, H.K.; Herkenham, M. Environmental enrichment requires adult neurogenesis to facilitate the recovery from psychosocial stress. Mol. Psychiatry 2010, 15, 1152–1163. [Google Scholar] [CrossRef]
- Chrusch, M.J.; Fu, S.; Spanswick, S.C.; Vecchiarelli, H.A.; Patel, P.P.; Hill, M.N.; Dyck, R.H. Environmental Enrichment Engages Vesicular Zinc Signaling to Enhance Hippocampal Neurogenesis. Cells 2023, 12, 883. [Google Scholar] [CrossRef]
- Qu, C.H.; Xu, L.L.; Shen, J.; Li, Y.Q.; Qu, C.J.; Song, H.; Zhang, J.J. Protection of blood-brain barrier as a potential mechanism for enriched environments to improve cognitive impairment caused by chronic cerebral hypoperfusion. Behav. Brain Res. 2020, 379, 112385. [Google Scholar] [CrossRef]
- Xing, Y.S.; Xia, Y.; Kendrick, K.; Liu, X.X.; Wang, M.S.; Wu, D.; Yang, H.; Jing, W.; Guo, D.Q.; Yao, D.Z. Mozart, Mozart rhythm and retrograde Mozart effects: Evidences from behaviors and neurobiology bases. Sci. Rep. 2016, 6, 18744. [Google Scholar] [CrossRef]
- Chen, S.; Liang, T.; Zhou, F.H.; Cao, Y.; Wang, C.; Wang, F.Y.; Li, F.; Zhou, X.; Zhang, J.Y.; Li, C.Q. Regular music exposure in juvenile rats facilitates conditioned fear extinction and reduces anxiety after foot shock in adulthood. Biomed. Res. Int. 2019, 2019, 8740674. [Google Scholar] [CrossRef]
- Habibi, A.; Damasio, A.; Ilari, B.; Veiga, R.; Joshi, A.A.; Leahy, R.M.; Haldar, J.P.; Varadarajan, D.; Bhushan, C.; Damasio, H. Childhood music training induces change in micro and macroscopic brain structure: Results from a longitudinal study. Cereb. Cortex 2018, 28, 4336–4347. [Google Scholar] [CrossRef] [PubMed]
- Kirste, I.; Nicola, Z.; Kronenberg, G.; Walker, T.L.; Liu, R.C.; Kempermann, G. Is silence golden? Effects of auditory stimuli and their absence on adult hippocampal neurogenesis. Brain Struct. Funct. 2015, 220, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Papadakakis, A.; Sidiropoulou, K.; Panagis, G. Music exposure attenuates anxiety-and depression-like behaviors and increases hippocampal spine density in male rats. Behav. Brain Res. 2019, 372, 112023. [Google Scholar] [CrossRef] [PubMed]
- Maguire, M.J. Wired for sound: The effect of sound on the epileptic brain. Seizure 2022, 102, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, M.H.; Chang, H.K.; Lee, T.H.; Lee, H.H.; Shin, M.C.; Shin, M.S.; Won, R.; Shin, H.S.; Kim, C.J. Influence of prenatal noise and music on the spatial memory and neurogenesis in the hippocampus of developing rats. Brain Dev. 2006, 28, 109–114. [Google Scholar] [CrossRef]
- Kim, C.H.; Lee, S.C.; Shin, J.W.; Chung, K.J.; Lee, S.H.; Shin, M.S.; Baek, S.B.; Sung, Y.H.; Kim, C.J.; Kim, K.H. Exposure to music and noise during pregnancy influences neurogenesis and thickness in motor and somatosensory cortex of rat pups. Int. Neurourol. J. 2013, 17, 107–113. [Google Scholar] [CrossRef]
- Lindig, A.M.; McGreevy, P.D.; Crean, A.J. Musical Dogs: A Review of the Influence of Auditory Enrichment on Canine Health and Behavior. Animals 2020, 10, 127. [Google Scholar] [CrossRef]
- Gupte, R.P.; Kadunganattil, S.; Shepherd, A.J.; Merrill, R.; Planer, W.; Bruchas, M.R.; Strack, S.; Mohapatra, D.P. Convergent Phosphomodulation of the Major Neuronal Dendritic Potassium Channel Kv4.2 by Pituitary Adenylate Cyclase-activating Polypeptide. Neuropharmacology 2016, 101, 291–308. [Google Scholar] [CrossRef]
- Nishiyama, J. Plasticity of dendritic spines: Molecular function and dysfunction in neurodevelopmental disorders. Psychiatry Clin. Neurosci. 2019, 73, 541–550. [Google Scholar] [CrossRef]
- Chen, M.; Jiang, Y.; Li, X.; Li, J.; Tu, C.; Zhang, H.; Gong, X.; Yu, G.; Guo, G.; Zhang, J. Enriched environment ameliorates learning and memory deficits in hepatic encephalopathy mice by restoration of the structure of dendrites and dendritic spines. Brain Res. 2023, 1804, 148–264. [Google Scholar] [CrossRef]
- Hruska, M.; Henderson, N.; Le Marchand, S.J.; Jafri, H.; Dalva, M.B. Synaptic nanomodules underlie the organization and plasticity of spine synapses. Nat. Neurosci. 2018, 21, 671–682. [Google Scholar] [CrossRef]
- Foggetti, A.; Baccini, G.; Arnold, P.; Schiffelholz, T.; Wulff, P. Spiny and Non-spiny Parvalbumin-Positive Hippocampal Interneurons Show Different Plastic Properties. Cell Rep. 2019, 27, 3725–3732. [Google Scholar] [CrossRef] [PubMed]
- Leggio, M.G.; Mandolesi, L.; Federico, F.; Spirito, F.; Ricci, B.; Gelfo, F.; Petrosini, L. Environmental enrichment promotes improved spatial abilities and enhanced dendritic growth in the rat. Behav. Brain Res. 2005, 163, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Aghighi Bidgoli, F.; Salami, M.; Talaei, S.A. Environmental enrichment restores impaired spatial memory and synaptic plasticity in prenatally stress exposed rats: The role of GABAergic neurotransmission. Int. J. Dev. Neurosci. 2020, 80, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Stein, L.R.; O’Dell, K.A.; Funatsu, M.; Zorumski, C.F.; Izumi, Y. Short-term environmental enrichment enhances synaptic plasticity in hippocampal slices from aged rats. Neuroscience 2016, 329, 294–305. [Google Scholar] [CrossRef] [PubMed]
- von Bohlen Und Halbach, O.; von Bohlen Und Halbach, V. BDNF effects on dendritic spine morphology and hippocampal function. Cell Tissue Res. 2018, 373, 729–741. [Google Scholar] [CrossRef] [PubMed]
- Mang, C.S.; Campbell, K.L.; Ross, C.J.D.; Boyd, L.A. Promoting neuroplasticity for motor rehabilitation after stroke: Considering the effects of aerobic exercise and genetic variation on brain-derived neurotrophic factor. Phys. Ther. 2013, 93, 1707–1716. [Google Scholar] [CrossRef]
- Martínez-Molina, N.; Siponkoski, S.T.; Särkämö, T. Cognitive efficacy and neural mechanisms of music-based neurological rehabilitation for traumatic brain injury. Ann. N. Y. Acad. Sci. 2022, 1513, 20–32. [Google Scholar] [CrossRef]
- Mualem, O.; Lavidor, M. Music education intervention improves vocal emotion recognition. Int. J. Music. Educ. 2015, 33, 413–425. [Google Scholar] [CrossRef]
- Ohline, S.M.; Abraham, W.C. Environmental enrichment effects on synaptic and cellular physiology of hippocampal neurons. Neuropharmacology 2019, 145, 3–12. [Google Scholar] [CrossRef]
- Braun, K.; Häberle, B.M.; Wittmann, M.T.; Lie, D.C. Enriched environment ameliorates adult hippocampal neurogenesis deficits in Tcf4 haploinsufficient mice. BMC Neurosci. 2020, 21, 50. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.Y.; Yu, K.W.; Zhou, N.Y.; Shen, X.Y.; Tian, S.; Zhang, B.; Wang, Y.Y.; Wu, J.F.; Liu, G.; Jiang, C.Y.; et al. Enriched Environment Elicits Proangiogenic Mechanisms After Focal Cerebral Ischemia. Transl. Stroke Res. 2019, 10, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.M.; Zhao, E.C.; Chen, L.M.; Guo, W.Q.; Zheng, J.Y.; Lin, L.; Wang, F.; Dong, W.G. Effects of early electroacupuncture on expressions of synapsin and PSD-95 in cortex and hippocampus of SAMP8 mice. Chin. J. Inf. Tradit. Chin. Med. 2021, 28, 49–53. [Google Scholar]
- Angelucci, F.; Fiore, M.; Ricci, E.; Padua, L.; Sabino, A.; Tonali, P. Investigating the neurobiology of music: Brain-derived neurotrophic factor modulation in the hippocampus of young adult mice. Behav. Pharmacol. 2007, 18, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, S.; Wadhwa, S. Prenatal auditory stimulation alters the levels of CREB mRNA, p-CREB and BDNF expression in chick hippocampus. Int. J. Dev. Neurosci. 2009, 27, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Chikahisa, S.; Sei, H.; Morishima, M.; Sano, A.; Kitaoka, K.; Nakaya, Y.; Morita, Y. Exposure to music in the perinatal period enhances learning performance and alters BDNF/TrkB signaling in mice as adults. Behav. Brain Res. 2006, 169, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Taherian, N.; Vaezi, G.; Neamati, A.; Etemad, L.; Hojjati, V.; Gorji-Valokola, M. Vitamin B12 and estradiol benzoate improve memory retrieval through activation of the hippocampal AKT, BDNF, and CREB proteins in a rat model of multiple sclerosis. Iran. J. Basic. Med. Sci. 2021, 24, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Poo, M.M. Neurotrophins as synaptic modulators. Nat. Rev. Neurosci. 2001, 2, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Tyler, W.J.; Alonso, M.; Bramham, C.R.; Pozzo-Miller, L.D. From acquisition to consolidation: On the role of brain-derived neurotrophic factor signaling in hippocampal-dependent learning. Learn. Mem. 2002, 9, 224–237. [Google Scholar] [CrossRef]
- Li, W.-J.; Yu, H.; Yang, J.-M.; Gao, J.; Jiang, H.; Feng, M.; Zhao, Y.-X.; Chen, Z.-Y. Anxiolytic effect of music exposure on BDNFMet/Met transgenic mice. Brain Res. 2010, 1347, 71–79. [Google Scholar] [CrossRef]
- Yamada, K.; Nabeshima, T. Brain-derived neurotrophic factor/TrkB signaling in memory processes. J. Pharmacol. Sci. 2003, 91, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Ly, C.; Greb, A.C.; Cameron, L.P.; Wong, J.M.; Barragan, E.V.; Wilson, P.C.; Burbach, K.F.; Soltanzadeh Zarandi, S.; Sood, A.; Paddy, M.R.; et al. Psychedelics Promote Structural and Functional Neural Plasticity. Cell Rep. 2018, 23, 3170–3182. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.; Mogul, A.S.; Taylor-Yeremeeva, E.M.; Khan, A.; Tirabassi, A.D.; Wang, H.Y. Stress Diminishes BDNF-stimulated TrkB Signaling, TrkB-NMDA Receptor Linkage and Neuronal Activity in the Rat Brain. Neuroscience 2021, 473, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Michler, S.A.; Illing, R.B. Molecular plasticity in the rat auditory brainstem: Modulation of expression and distribution of phosphoserine, phospho-CREB and TrkB after noise trauma. Audiol. Neurootol. 2003, 8, 190–206. [Google Scholar] [CrossRef]
- Zheng, K.; An, J.J.; Yang, F.; Xu, W.; Xu, Z.Q.; Wu, J.; Hökfelt, T.G.; Fisahn, A.; Xu, B.; Lu, B. TrkB signaling in parvalbu-min-positive interneurons is critical for gamma-band network synchronization in hippocampus. Proc. Natl. Acad. Sci. USA 2011, 108, 17201–17206. [Google Scholar] [CrossRef]
- Fries, G.R.; Saldana, V.A.; Finnstein, J.; Rein, T. Molecular pathways of major depressive disorder converge on the synapse. Mol. Psychiatry 2023, 28, 284–297. [Google Scholar] [CrossRef]
- Kaplan, D.R.; Miller, F.D. Neurotrophin signal transduction in the nervous system. Curr. Opin. Neurobiol. 2000, 10, 381–391. [Google Scholar] [CrossRef]
- Jin, T.; Zhang, Y.; Botchway, B.O.A.; Zhang, J.; Fan, R.; Zhang, Y.; Liu, X. Curcumin can improve Parkinson’s disease via activating BDNF/PI3k/Akt signaling pathways. Food Chem. Toxicol. 2022, 164, 113091. [Google Scholar] [CrossRef]
- Lai, H.C.; Chang, Q.Y.; Hsieh, C.L. Signal Transduction Pathways of Acupuncture for Treating Some Nervous System Diseases. Evid. Based Complement. Alternat. Med. 2019, 2019, 2909632. [Google Scholar] [CrossRef]
- Lopes, C.M.; Remon, J.I.; Matavel, A.; Sui, J.L.; Keselman, I.; Medei, E.; Shen, Y.; Rosenhouse-Dantsker, A.; Rohacs, T.; Logo-thetis, D.E. Protein kinase A modulates PLC-dependent regulation and PIP2-sensitivity of K+ channels. Channels 2007, 1, 124–134. [Google Scholar] [CrossRef]
- Bai, L.; Zhang, S.; Zhou, X.; Li, Y.; Bai, J. Brain-derived neurotrophic factor induces thioredoxin-1 expression through TrkB/Akt/CREB pathway in SH-SY5Y cells. Biochimie 2019, 160, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N.; Enomoto, A.; Nagai, T.; Takahashi, M.; Yamada, K. Molecular mechanism linking BDNF/TrkB signaling with the NMDA receptor in memory: The role of Girdin in the CNS. Rev. Neurosci. 2016, 27, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yao, L.; Yang, J.; Wang, Z.; Du, G. PI3K/Akt and HIF-1 signaling pathway in hypoxia-ischemia (Review). Mol. Med. Rep. 2018, 18, 3547–3554. [Google Scholar] [CrossRef]
- Pirbhoy, P.S.; Farris, S.; Steward, O. Synaptically driven phosphorylation of ribosomal protein S6 is differentially regulated at active synapses versus dendrites and cell bodies by MAPK and PI3K/mTOR signaling pathways. Learn. Mem. 2017, 24, 341–357. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiang, L.; Wang, C.; Song, Y.; Miao, J.; Miao, M. Protection against acute cerebral ischemia/reperfusion injury by Leonuri Herba Total Alkali via modulation of BDNF-TrKB-PI3K/Akt signaling pathway in rats. Biomed. Pharm. 2021, 133, 111021. [Google Scholar] [CrossRef]
- Falcicchia, C.; Tozzi, F.; Arancio, O.; Watterson, D.M.; Origlia, N. Involvement of p38 MAPK in Synaptic Function and Dys-function. Int. J. Mol. Sci. 2020, 21, 5624. [Google Scholar] [CrossRef]
- Carlini, V.P.; Poretti, M.B.; Rask-Andersen, M.; Chavan, R.A.; Ponzio, M.F.; Sawant, R.S.; de Barioglio, S.R.; Schiöth, H.B.; de Cuneo, M.F. Differential effects of fluoxetine and venlafaxine on memory recognition: Possible mechanisms of action. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 38, 159–167. [Google Scholar] [CrossRef]
- Lewczuk, A.; Boratyńska-Jasińska, A.; Zabłocka, B. Validation of the Reference Genes for Expression Analysis in the Hippocampus after Transient Ischemia/Reperfusion Injury in Gerbil Brain. Int. J. Mol. Sci. 2023, 24, 2756. [Google Scholar] [CrossRef]
- Ji, R.R.; Befort, K.; Brenner, G.J.; Woolf, C.J. ERK MAP kinase activation in superficial spinal cord neurons induces prodynorphin and NK-1 upregulation and contributes to persistent inflammatory pain hypersensitivity. J. Neurosci. 2002, 22, 478–485. [Google Scholar] [CrossRef]
- Li, Y.T.; Pan, L.; Zeng, X.Y.; Zhang, R.X.; Li, X.; Li, J.; Xing, H.J.; Bao, J. Ammonia exposure causes the imbalance of the gut-brain axis by altering gene networks associated with oxidative metabolism, inflammation and apoptosis. Ecotoxicol. Environ. Saf. 2021, 224, 112668. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.T.; Wang, J.X.; Wang, Y.L.; Liu, H.G.; Bao, J. Selenium Alleviates Ammonia-Induced Splenic Cell Apoptosis and Inflammation by Regulating the Interleukin Family/Death Receptor Axis and Nrf2 Signaling Pathway. Biol. Trace Elem. Res. 2022, 201, 1748–1760. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.X.; Wang, J.; Li, Y.T.; Han, Q.; Wang, Y.L.; Liu, H.G.; Bao, J. Organic Selenium Alleviates Ammonia-Mediated Abnormal Autophagy by Regulating Inflammatory Pathways and the Keap1/Nrf2 Axis in the Hypothalamus of Finishing Pigs. Biol. Trace Elem. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.T.; Wang, J.; Xing, H.J.; Bao, J. Selenium Mitigates Ammonia-Induced Neurotoxicity by Suppressing Apoptosis, Immune Imbalance, and Gut Microbiota-Driven Metabolic Disturbance in Finishing Pigs. Biol. Trace Elem. Res. 2022. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Wang, J.; Wang, Y.; Chai, Y.; Li, H.; Miao, D.; Liu, H.; Li, J.; Bao, J. Music with Different Tones Affects the Development of Brain Nerves in Mice in Early Life through BDNF and Its Downstream Pathways. Int. J. Mol. Sci. 2023, 24, 8119. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24098119
Wang J, Wang J, Wang Y, Chai Y, Li H, Miao D, Liu H, Li J, Bao J. Music with Different Tones Affects the Development of Brain Nerves in Mice in Early Life through BDNF and Its Downstream Pathways. International Journal of Molecular Sciences. 2023; 24(9):8119. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24098119
Chicago/Turabian StyleWang, Jing, Jianxing Wang, Yulai Wang, Yiwen Chai, Haochen Li, Deyang Miao, Honggui Liu, Jianhong Li, and Jun Bao. 2023. "Music with Different Tones Affects the Development of Brain Nerves in Mice in Early Life through BDNF and Its Downstream Pathways" International Journal of Molecular Sciences 24, no. 9: 8119. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24098119