

Potential Applications of Thermophilic Bacteriophages in One Health

School of Veterinary Medicine, Texas Tech University, Amarillo, TX 79106, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(9), 8222; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24098222
Submission received: 31 March 2023
/
Revised: 26 April 2023
/
Accepted: 2 May 2023
/
Published: 4 May 2023
(This article belongs to the Special Issue Thermophilic and Hyperthermophilic Microbes and Enzymes 2.0)
Abstract
:Bacteriophages have a wide range of applications such as combating antibiotic resistance, preventing food contamination for food safety, and as biomarkers to indirectly assess the quality of water. Additionally, bacteriophage components (endolysins and coat proteins) have a lot of applications in food processing, vaccine design, and the delivery of cargo to the body. Therefore, bacteriophages/components have a multitude of applications in human, plant/veterinary, and environmental health (One Health). Despite their versatility, bacteriophage/component use is mostly limited to temperatures within 4–40 °C. This limits their applications (e.g., in food processing conditions, pasteurization, and vaccine design). Advances in thermophilic bacteriophage research have uncovered novel thermophilic endolysins (e.g., ΦGVE2 amidase and MMPphg) that can be used in food processing and in veterinary medicine. The endolysins are thermostable at temperatures > 65 °C and have broad antimicrobial activities. In addition to thermophilic endolysins, enzymes (DNA polymerase and ligases) derived from thermophages have different applications in molecular biology/biotechnology: to generate DNA libraries and develop diagnostics for human and animal pathogens. Furthermore, coat proteins from thermophages are being explored to develop virus-like particle platforms with versatile applications in human and animal health. Overall, bacteriophages, especially those that are thermophilic, have a plethora of applications in One Health.
1. Introduction
Bacteriophages or phages are viruses that infect bacteria. It is estimated that more than 1031 bacteriophages are present on earth; bacteriophages are thus considered to be the most abundant organisms on earth [1,2,3,4,5,6]. They can be identified/isolated from wherever their host bacteria are present; this includes, but is not limited to, the soil, oceans and lakes, humans and animals, and hot springs and hydrothermal vents. Phages can be grouped into three groups based on the temperature at which their hosts bacteria grow. Phages that infect bacteria with an optimum growth temperature of 37 °C are known as mesophilic phages, while those that infect bacteria that grow at temperatures up to 75 °C are referred to as thermophilic phages. Those that infect bacteria that grow at >75 °C are known as hyperthermophilic phages. Thermophilic and hyperthermophilic phages will be referred to in this review as thermophages. Phages in general have a wide range of applications in human, animal, and plant health as well as in food safety.
1.1. One Health and the Application of Mesophilic Phages
The food and animal industries are globalized with a supply chain that has greatly enhanced people’s welfare. However, the globalized nature of the supply chain can contribute to the spread of foodborne and zoonotic diseases; Salmonella, Listeria, E. coli O157, Shiga toxin-producing E. coli O104:H4, hepatitis A, Norovirus, etc., have been linked to imported foods [7,8,9]. Thus, the world’s population is vulnerable to foodborne pathogens (including those that are resistant to antibiotics) as well as emerging and re-emerging pathogens (reviewed in [10]). For example, the outbreaks of zoonotic diseases such as SARS, bird flu, Ebola, COVID-19, etc., within the last two decades reminds us that a solution to human health problems (for example, zoonotic diseases, foodborne diseases, antibiotic resistance, and the environment) should be cross-species and multi-disciplinary in nature. To tackle this problem, an interdisciplinary concept “One Health” was created, which is a collaborative effort to achieve optimal health outcomes based on the interconnection between humans, animals, food and agriculture, and the environment [11]. Bacteriophages have contributed and continue to contribute towards the efforts of One Health as described below:
- (a)
- Mesophilic phages have been used within the last decades to combat antibiotic resistance in humans and animals [12,13]. Bacterial resistance to antibiotics is on the rise and alternative approaches are needed to control infections (reviewed in [14,15]). Phages have been used successfully to treat antibiotic-resistant bacterial infections such as: Acinetobacter baumannii infection in a 68-year-old diabetic patient [16], Staphylococcus aureus-associated chronic rhinosinusitis infection [17], and colistin-only or ceftazidime-sensitive Pseudomonas aeruginosa infections in patients with septicemia or with an infected aortic graft [18,19]. Phages have also been used to significantly reduce the concentration of bacteria (Salmonella, Campylobacter) in chickens, Staphylococcus in bovine, and Yersinia, Salmonella, and Bordetella in pigs (reviewed in [20,21]); thus, phages can be used as an alternative treatment for bacterial infections (especially those with antibiotic resistance) in humans and animals. Other studies in plants have also shown that phages can be used as an alternative to copper-based pesticides, which can contaminate the environment, to treat/combat antibiotic resistance in pathogenic plants [22,23,24].
- (b)
- Phages can be used in food safety to prevent the contamination of refrigerated and non-refrigerated food by bacteria [13,25,26,27,28]. For example, an E. coli phage, Tequatrovirus EP01, has the potential to reduce the concentration of Salmonella Enteritidis, E. coli O157:H7, E. coli O114:K90, and E. coli O142:K86 in contaminated meat and milk by 2.18–6.55 log10 CFU/sample [27]. In addition to this, a lytic enzyme (endolysin, lysin PhiH5) derived from a Staphylococcus aureus phage (PhiH5) has the potential to kill Staphylococcus aureus in pasteurized milk [29]. Moreover, GBS B30 endolysin from a streptococcus phage has the potential to lyse three Streptococcus spp. (S. agalactiae, S. dysgalactiae, and S. uberis) associated with mastitis in cattle [30].
- (c)
- (d)
- Phages have been used as biomarkers to indirectly assess the quality of drinking water [34,35]. For example, using T7 phage-conjugated magnetic beads, 1 × 104 colony forming units (cfu)/mL of E. coli can be detected in drinking water in 2.5 h; additionally, 10 cfu/mL can be detected using the same system after 6 h of sample pre-enrichment [34]. Other assays, non-phage-based, can detect 1 cfu/mL of E. coli (but in 12.5 h [36]) and 2 × 106 cfu/mL (in less than 1 h but it is less sensitive [37]). In addition to this, non-phage-based assays require advanced instruments and training.
- (e)
- (f)
- VLPs can be loaded with fluorophores for the imaging of cancer and fibrin clots in vivo (reviewed in [41]).
Thus, phages play a critical role in the One Health approach. They can be used to improve the health of humans, animals, and plants (which both humans and animals dependent on); they can also be used to improve the safety of our food as well as to assess the quality of water, in the environment, for human and animal consumption. Despite the importance/potential application of mesophilic phages in One Health, there are some challenges associated with (that limit) their use.
1.2. Limitation of Mesophilic Phages
Mesophilic phages or their components (e.g., endolysins, virus-like particles) cannot maintain their integrity and/or function at environmental conditions above or below their optimum growth temperature (of host bacteria) and thus limit their application/use in One Health as described below.
- (a)
- Endolysins (e.g., lysin PhiH5, LysB4, GBS B30 endolysin) derived from mesophilic phages have the potential to lyse bacteria [29,30,42,43]. However, they lose activity or are inactivated at higher temperatures (such as 55–63 °C for 30 min, 72 °C for 15 min, −20 °C for 30 min) and at pH < 4. Thus, such endolysins cannot tolerate food processing conditions such as pasteurization, nor could they be effective (if applied topically) in treating antibiotic-resistant mastitis-causing bacteria (such as Staphylococcus aureus, Streptococcus spp.) in the field, especially in areas where temperatures rise above 30 °C.
- (b)
- Virus-like particles (VLPs), derived from mesophilic phages and used as platforms to design vaccines against other infectious agents, are not stable at room temperature [44] and thus require refrigeration. Virus-like particles are empty shells derived from the capsid or envelop proteins of viruses [45]. They lack viral genomes and thus, they are not infectious. In addition to phage-derived VLPs, other viral-derived VLPs have been used as vaccines against viruses (human papillomaviruses, hepatitis B virus, and hepatitis A virus) from which the VLPs are derived [46,47,48,49]. In addition to this, they have also been used as platforms to display and enhance the immunogenicity of less-immunogenic peptide antigens from other infectious agents. The display of foreign peptides on the surface of VLPs enhances the immunogenicity of the peptides and offers protection against infection by the infectious agents from which the peptides were derived [32,50,51,52]. However, VLPs derived from mesophilic viruses have some limitations.
- (i)
- Like most vaccines, VLP-based vaccines require refrigeration, thus limiting their distribution/use in countries with limited cool-chain infrastructure. Excellent examples are COVID-19 vaccines. Although the vaccines are not VLP-based, they still require refrigeration; 100 million doses of the vaccines were rejected in 2021 by poorer nations due to thermostability-/expiration-related issues [53,54]. VLP-based vaccines are likely to face similar challenges during a pandemic. It is also worth mentioning that a refrigeration requirement for vaccines can also lead to the accidental freezing of vaccines (for facilities that have them); for example, it is estimated that ~14–35% of refrigerated vaccines are accidentally exposed to freezing temperatures during transportation/storage, thus decreasing their efficacy [55,56].
- (ii)
- Pre-existing antibodies, in human and animal populations, against a platform may affect efficacy. Although pre-existing antibodies to phages that infect bacteria that colonize humans/animals (in a natural setting) have not been reported [57], pre-existing antibodies against some phages/vaccine platforms (in experimental studies) have the potential to attenuate efficacy [58]. In a study, in an experimental setting, pre-existing A3R and 676Z phage-specific IgA antibodies in the gut of mice were shown to reduce the bioavailability of the phages in the gut [59]; a reduction in phage bioavailability is likely to adversely affect phage therapy.
- (iii)
- Lastly, VLPs derived from mesophilic bacteria have a limitation on the quantity of cargo (drugs, vaccine adjuvants such as toll-like receptor ligands, fluorophores, etc.) that can be loaded into the inner core of the VLPs for delivery into the body/cells. Most phage VLPs (MS2, QB, PP7) with icosahedral structures used for cargo delivery are ~28 nm in diameter [39,40,60], thus limiting the quantity of cargo that can be loaded into them.
To circumvent some of these problems, enzymes for food processing and structural proteins for the development of VLP vaccine platforms can be derived from thermophilic/hyperthermophilic phages; the bacteria that they infect are known as thermophilic bacteria.
2. Overview of Thermophages
2.1. Diversity of Thermophages
Thermophages can be isolated from hot springs, including hot springs on land [61,62] and submarine alkaline freshwater, as well as coastal springs, deep-sea hydrothermal vents [63], compost piles, dairy environments, soil, industrial hot waters, etc. [64]. Regardless of where they are isolated, thermophages differ in their specificity for the bacterial host they infect, genome/particle size, shape, and structure (Table 1). Some thermophages can infect a wide variety of thermophilic bacterial species, whereas others can infect only a limited number of thermophilic bacteria. For example, phages P23–45 and P74–26B can infect Thermus aquatic, Thermus flavus, Thermus thermophilus HB8, and Thermus sp. ATCC 31,674 [62], while the filamentous phage, φOH3, seems to infect only the HB8 strain of Thermus thermophilus (and not other strains such as HB27, AT62, TMY, etc.) [65].
Thermophage particles come in different shapes (Figure 1); some have an icosahedral capsid (oval head without a tail and are called tail-less phages) [66] while others are oval with a tail (tailed phages) [67]. In addition to tail-less and tailed thermophages, some thermophages are filamentous in shape [65,68,69]. Their genome sizes are also diverse; genome sizes range from 5.2 kb to 152.4 kb [26,65,70], and virion particles are within 60–130 nm wide (for oval-shaped thermophages) to 910 nm long (for filamentous thermophages [69]. Their genomes can be linear [71], circular [72], single-stranded [65], or double-stranded [66,71]. The GC content of the genome ranges from 32 to 68%. Most thermophages infect and replicate in bacteria with optimum growth temperatures from 55 to 70 °C (Table 1).

{kind=link}
Table 1.
Diversity of thermophilic phages.
| Phage | Phage Genome | Genome Size (kb) | Morphology | Size of Phage | Isolation Origin | Host Bacteria Isolated from | Optimum/Cultured Temperature of Host Bacteria * (°C) | Optimum/Cultured pH of Host Bacteria | References |
|---|---|---|---|---|---|---|---|---|---|
| P74–26 | Double-stranded (ds)DNA | 83.32 | Oval capsid (head) and a tail | Head: 824 Å Tail: ~800 nm | Kamchatka peninsula hot springs, Russia | Thermus thermophilus (T. thermophilus) | 65 | 6.0 to 7.0 | [73,74] |
| φOH3 | Single-stranded (ss)DNA | 5.7 | Filamentous | 830 nm long and 8 nm wide | Obama hot spring, Japan | T. thermophilus HB8 | 70 | 7.0 | [65] |
| ϕNS11 | dsDNA | 57 | Oval capsid with spike-like structures | Diameter: 60–75 nm | Beppu hot springs, Japan | A. acidocaldarius strain TA6 | 60 to 65 | 3.5 | [62,64,75] |
| TP-84 | dsDNA | 47.7 | Oval capsid and a non-contractile tail | Head: 53 × 30 nm Tail: 131 nm long and 3–5 nm wide | Greenhouse soil | Geobacillus stearothermophilus | 55 to 60 | 6.5 | [64,76] |
| TS2126 | dsDNA | ~90 | ** | ** | Hot tap water in Iceland | Thermus scotoductus | 65 | 7.5 | [77,78] |
| ΦGVE2 | dsDNA | 40.9 | ** | ** | Deep-sea hydrothermal fields in the Pacific | Geobacillus sp. E263 | 60 to 65 | 7.0 | [63,64,79] |
| MMP17 | dsDNA | 33.5 to 39.5 | Oval capsid and a tail | Head: 42 nm Tail: 120 nm long and 17 nm wide | Eryuan hot spring, China | Meiothermus | 55 to 60 | 6.0 to 7.0 | [80] |
| P23–77 | dsDNA | 17.04 | Icosahedral capsid with spikes on the vertices, and an internal lipid membrane, no tail | Diameter: 78 nm | Alkaline hot springs, New Zealand | T. thermophilus | 70 | 6.0 to 7.0 | [72] |
| ΦIN93 | dsDNA | 19.6 | Icosahedral | Diameter: ~130 nm | Hot spring soil, Japan | Thermus aquaticus | 70 | 6.0 to 7.0 | [61] |
| PH75 | ssDNA | 6.5 | Filamentous | Length: 910 ± 17 nm | Hot spring, New Zealand | T. thermophilus | 70 | 8.3 | [62,69] |
| ϕTMA | dsDNA | 151.5 | Icosahedral capsid and a contractile tail | Head: ~125 nm Tail: ~175 nm | Atagawa hot spring, Japan | T. thermophilus | 65 | 6.0 to 7.0 | [26] |
| ϕYS40 | dsDNA | 152.4 | Oval capsid and a contractile tail | Head: 125 nm Tail: 178 nm long and 27 nm wide | Atagawa hot spring, Japan | T. thermophilus | 65 | 6.0 to 7.0 | [26,81,82] |
| RM378 | dsDNA | 129.9 | Isometric capsid and a tail | Head: 85–95 nm Tail: 150 nm | Slightly saline, geothermal environments, and hot springs, Iceland | Rhodothermus marinus | 65 | 7.2 | [70] |
* They can also grow at higher temperatures but at lower rate; ** No information available.
2.2. Features That Confer Thermostability in Extreme Hot Environments
- (a)
- The capsid: The stability of the capsid of thermophages is crucial for their survival in host bacteria, as well as in the environment, following their release from the host (i.e., until they infect the next bacteria). The organization/geometry of capsomeres (coat proteins), which make up the capsid, contributes to the stability of the capsid. It is believed that the capsid of thermophage P74–26 is stable at 80 °C (unlike that of a mesophilic lambda phage heated at the same temperature) due to the fact that its coat proteins are topologically “tied” together by lassos [74]. The thermostability of the capsid thus protects the genome from environmental stress such as high temperature and low/high pH. For example, thermophage ϕNS11 (PhiNS11) remains stable at 60 °C (pH 2–5), while its naked DNA is degraded at 55 °C (pH 4) [84]; ϕNS11 infects acidophilic thermophilic bacteria and thus, it also considered to be an acidophilic phage.
- (b)
- The guanine–cytosine (GC) content: The GC content of nucleic acids of thermophages also contributes to the thermostability of their genomes at high temperatures. For example, the GC content of thermophage P23–77 and ΦIN93 is 68% and 66%, respectively [66,85], while that of mesophilic phages (e.g., Salmonella phage BIS20 and E. coli lambda phage) is 53% and 50%, respectively [86,87]. Given its higher GC contents, higher temperatures are required to break the three hydrogen bonds between guanine and cytosine than the two hydrogen bonds between thymine and adenine in mesophilic phages. This thus enhances the thermostability of genomes of thermophages compared to those of mesophilic phages.
- (c)
- Osmolarity: Inorganic ions (calcium, Ca2+, or potassium, K+) in salts enhance the stability of thermophages (the head, tail, and DNA). Positive ions help neutralize negative phosphate charges on sugar molecules (on nucleic acid) that repel each other, thus stabilizing DNA. As a matter of fact, the supplementation of thermophage (e.g., TP-84) growth media with calcium or potassium ions enhances survival/titers by 18–10,000-fold at 65 °C [76,88]. On the contrary, the chelation of these ions with EDTA (ethylenediaminetetraacetic acid) or the addition of phosphate (additional negative ions that promote repulsion) leads to the dissociation of the tail from the head of thermophage TP-84 [89].
Overall, the osmolarity of a host bacterium and its environment, the GC content of thermophages, and the geometry/tethering of capsomeres on the capsid all contribute to the stability of thermophages and, subsequently, their ability to thrive at high temperatures.
3. Application of Thermophages in One Health
While live thermophages cannot be used to control bacterial infections in humans, animals, or pathogenic plants (due to their inability to infect/replicate in bacteria that infect humans, animals, and plants), their components (enzymes, capsid proteins) can be used in molecular biology to improve the health of humans and animals, as well as in food safety. As mentioned above, components, such as endolysins and virus-like particles, derived from mesophilic phages lose activity (with time) at room temperature and above; this thus limits their application in food processing, in treating antibiotic-resistant mastitis-causing bacteria in the field, and in treating pathogenic plants, as well as their applications as platforms for vaccine design. These limitations can be overcome by using enzymes (endolysins and viral polymerases) and coat proteins, derived from thermophages.
3.1. Thermophage Enzymes in Molecular Biology
Enzymes derived from thermophages have a lot of applications in molecular biology. For example, PyroPhage 3173 DNA Pol polymerase used in reverse transcription (RT) polymerase chain reaction (PCR) was derived from a thermophage genomic library generated using samples collected from Yellowstone hot springs [90]. RT-PCR/PCR is normally performed using a two-enzyme system: a reverse transcriptase derived from MMLV (Moloney Murine Leukemia Virus) and Taq polymerase derived from thermophilic bacteria (Thermus aquaticus). While MMLV RT can convert RNA to cDNA for PCR, it lacks thermostability and thus, Taq polymerase, which is active at temperatures up to 95 °C for 40 min (reviewed in [91]), is used as an enzyme for PCR after reverse transcription. With the expression and purification of PyroPhage 3173 DNA Pol polymerase (thermostable at 94 °C for 11 min) from thermophages, RT-PCR/PCR can now be performed using a single enzyme. The enzyme catalyzes both reverse transcription and DNA amplification in a single reaction [90,92]. A modified version of PyroPhage 3173 DNA Pol polymerase, OmniAmp Pol, has been developed and used in point of care (POC) diagnostic tests in LAMP (loop-mediated isothermal amplification) assays to detect human and animal infectious agents such as swine influenza virus, porcine circovirus-2, West Nile virus, Ebola virus, Crimean–Congo hemorrhagic fever, Bovine viral diarrhea virus, Edwardsiella ictaluri, Bacillus atrophaeus, and Staphylococcus aureus. It is worth mentioning that the POC LAMP assays using OmniAmp Pol amplified target genes in <30 min compared to RT-LAMP using Bst polymerase (which takes about 20% longer) [93].
In addition to the above enzymes, another thermophage enzyme (Phage TS2126 ligase) isolated from a thermophage TS2126 [77] is very crucial in molecular biology; the enzyme is stable at 60–65 °C. Phage TS2126 ligase (marketed as CirLigaseTM) is used to circularize single-stranded DNA for many applications (rolling-circle replication/transcription) [94]. The enzyme has been used to generate DNA libraries for next-generation sequencing [95]. In addition to Phage TS2126 ligase, RM378 RNA ligase 1 (derived from thermophage RM378) ligates both ssDNA and RNA molecules. Given its thermostability features (optimum temperature 60–64 °C), it is used in the RNA-ligase-mediated rapid amplification of cDNA ends [96], and the adenylation of small RNA sequencing adapters [97]. Thus, phage TS2126 ligase, PyroPhage 3173 DNA Pol polymerase, and its modified version, OmniAmp Pol, have the potential to make a significant contribution to One Health, especially in human and animal health. The enzymes can contribute to the sequencing of the genomes of humans and animals. They can also contribute towards the sequencing of their microbiome, thus contributing to the diagnosis of new pathogens with unknown treatment.
3.2. Thermophage Enzymes in Veterinary Medicine
Enzymes isolated from different thermophages are stable at high temperatures and therefore have potential applications in veterinary medicine and One Health in general. For example, an endolysin (ΦGVE2 amidase) isolated from a deep-sea thermophilic bacteriophage, Geobacillus virus E2 (ΦGVE2), is thermostable at temperatures up to 80 °C. The fusion of the endolysin with an endolysin from a Clostridium perfringens phage (recombinant fusion endolysin called PlyGVE2CpCWB) lyses Clostridium perfringens [98]. Clostridium perfringens is a foodborne pathogen (infects both humans and animals) and is associated with diarrhea in humans, cattle, horses, pigs, chickens, goats, etc. [99,100]; thus, it is present in fecal matter and in soil, where it can form endospores (resistant to environmental stress). Given the stability/activity of PlyGVE2CpCWB endolysin (95% activity after a 30 min incubation at 50 °C), the enzyme can be added to animal feed/water or used in animal facilities to prevent contamination with Clostridium perfringens; it can also be used in food processing to prevent the contamination of chicken/meat with the bacteria.
Recently, another thermostable endolysin (MMPphg), derived from thermophage MMP17, has been expressed and purified [101]. MMPphg is thermostable at temperatures up to 65 °C for 30 min (80% of its activity retained). Unlike other endolysins which are efficacious mostly against Gram-positive bacteria (reviewed in [102]), MMPphg has antimicrobial activities against both Gram-negative and Gram-positive bacteria including antibiotic-resistant strains. At 37 °C for 1 h, the enzyme lysed Escherichia coli O157, Staphylococcus aureus (including resistant strain KMUST1606BL1486), Salmonella enterica serovar Enteritidis, Salmonella enterica serovar Typhi, Salmonella enterica serovar Paratyphi B, Shigella dysenteriae, and Klebsiella pneumoniae (this includes nine antibiotic-resistant strains). Bacterial cells in antimicrobial assays were reduced by 0.96–3.4 log10 [101]. Given the broad spectrum of antimicrobial activity and the thermostability of endolysin MMPphg, the enzyme could be used to treat Staphylococcus aureus infections associated with bovine mastitis.
3.3. Thermophage Coat Proteins and Potentials for Vaccine Platforms
Vaccines play a crucial role in protecting the health of humans and animals, making them an essential component of One Health. Coat proteins derived from thermophages can serve as an alternative source of structural proteins to develop virus-like particle platforms for vaccine design (i.e., display antigens from human and animal infectious agents) and to deliver cargo into the body/cells. P23–77 and ΦIN93 are thermophages with coat proteins with great potential to develop VLP platforms for vaccine design. P23–77 and ΦIN93 were isolated from thermophilic bacteria with an optimum growth temperature of 70 °C [61,72]. P23–77 has a diameter of 78 nm, while ΦIN93 has a diameter of 130 nm. The GC contents of the capsid proteins are very high (~66%); the coat proteins of P23–77 are thermostable at a temperature > 80 °C [103,104], and its capsid is composed of 6 nm-thick multilayered proteins [72]. Given the aforementioned features, VLPs derived from these two thermophages (P23–77 and ΦIN93) are likely thermostable at high temperatures, and their inner cores can accommodate large quantities of cargo.
Studies have shown that the capsid of P23–77 consists of two major coat proteins (VP16 and VP17) and one minor coat protein (VP11) [66,104]; VP11 is a ~22 KD protein, VP16 is ~19 KD, and VP17 is a ~32 KD protein. It is estimated that the viral capsid of P23–77 is assembled from 147 copies of VP11, 1080 copies of VP16, and 540 copies of VP17 [103,104]. The capsid protein of ΦIN93, on the other hand, is believed to be composed of two putative coat proteins (ORF13 and ORF14) based on sequence homology with the VP16 and VP17 of P23–77. ORF13 has 80% sequence identity with VP16, while ORF14 has 73% sequence identity with the VP17 of P23–77; additionally, the structures of coat proteins from both viruses are similar [61,105]. A sequence in thermophage ΦIN93 that is homologous to VP11 (minor capsid protein of P23–77) has not been identified, suggesting that the capsid protein of ΦIN93 is composed of only two coat proteins, ORF13 and ORF14. Attempts have been made within the last few years to assess the potential of coat proteins from P23–77 [104] and ΦIN93 [61,105] to assemble into VLPs. Pawlowski et al. [104] expressed and purified (separately from E. coli) VP11, VP16, and VP17 coat proteins; a mixture of the three purified coat proteins showed that they can form complexes in vitro. More recently, in a PhD dissertation, Yadav [106] successfully co-expressed (for the first time) truncated versions of VP16 and VP17 coat proteins together with VP11; co-expressed proteins formed concentric cycles, but their diameters were less than 70 nm. Recently, Zhai et al. [105] co-expressed ORF13 and OFR14 coat proteins of ΦIN93 (full-length and truncated versions) in E. coli. Similar to those of VP11, VP16, and VP17, the co-expression of the truncated versions of ORF13 and OFR14 formed oval structures with diameters (~75 nm to ~100 nm) smaller than those of the authentic ΦIN93 virus (130 nm). More studies are required to authenticate these oval structures and to further assess whether other structural proteins are required for the coat proteins to form complexes or assemble into VLPs.
4. Conclusions and Perspectives for the Future
Bacteriophages, especially their components (lytic enzymes and coat proteins), have a plethora of applications in One Health. Mesophilic phages can be used to combat antibiotic-resistant bacteria in humans, animals, and plants, and also in food processing (especially vegetables that may be contaminated with fecal water). In addition to live phages, lytic enzymes, especially those derived from thermophages (with greater thermostability), can be used in food processing, in treating water for livestock that may be contaminated with pathogenic bacteria, and in treating bacteria-associated mastitis in cattle. Temperature is very critical in the livestock industry; mesophilic phage components may present challenges for development and utilization in high-temperature environments, which is relatively common in animal rearing and product processing. While a lot of studies have demonstrated the efficacy of thermophilic endolysins (e.g., MMPphg, ΦGVE2 amidase) against a select group of bacteria (including Escherichia coli O157, Staphylococcus aureus, Clostridium perfringens, Salmonella enterica, Klebsiella pneumoniae, etc.) [98,101], more studies are required to assess their efficacy against other bacteria of veterinary importance such as Streptococcus infections (associated with bovine mastitis), Bacillus anthracis (associated with anthrax and forms endospores), etc. In addition to this, the ability of endolysin MMPphg to control Paenibacillus larvae (especially its spores) among other pathogenic honeybee bacteria should also be assessed. Paenibacillus larvae is a Gram-positive bacterium associated with fatal disease in the larvae of honeybees; it is present worldwide, with a high prevalence in the US and Europe [107,108]. While an endolysin (PlyPL123) isolated from a mesophilic phage (phiIBB_Pl23) lyses the bacterium (without toxicity to larvae from bees), the endolysin loses activity within a week at room temperature [109]. Thus, PlyPL123 endolysin may not be applicable in the field, especially where temperatures rise above 25 °C. An effective thermostable endolysin that is appliable in the field is needed to prevent the use of antibiotics in honeybee production, thus preventing its introduction into humans. Thermophage components have promising application prospects in the field of human and veterinary health.
Coat proteins derived from thermophages also have a lot of applications in One Health. They can be used to develop thermostable vaccines, to deliver cargo to specific cells, and for in vivo imaging. If successfully developed, VLPs, especially from ΦIN93 (with a diameter of 130 nm), will have the capacity to be loaded with more cargo (vaccine adjuvants, imaging fluorophores) for targeted delivery to cells. The diameter of ΦIN93 is almost twice the size of thermophilic P23–77 (78 nm) and more than four times the size of mesophilic MS2 VLPs (~28 nm), which is widely used in the field to deliver cargo [110,111,112]. The sizes of P23–77 and ΦIN93 have additional benefits as platforms for vaccine design/immunization. Studies have shown that nanoparticles less than 200 nm, such as VLPs, can directly enter the lymphatic vessels from the site of immunization (without association with injection-site antigen-presenting cells) and drain freely to the lymph nodes [113,114]. With this in mind, VLPs derived from P23–77 and ΦIN93 are likely to enter directly into the lymphatic vessels and drain efficiently to lymph nodes. Another benefit of VLPs derived from P23–77 and ΦIN93 is the fact that the public, including animals, lack pre-existing antibodies against these thermophages. Thus, VLPs derived from P23–77 and ΦIN93 are likely to be highly immunogenic in humans and animals. They can therefore be used to display foreign peptides on their surfaces, thus enhancing the immunogenicity of the peptides.
While thermophilic endolysins and their coat proteins have enormous potential in One Health, some challenges lie ahead. For example, the application of endolysins to treat gut bacterial infections in humans and animals may be limited, given challenges to deliver them to the gut without inactivation by gastric juice, which is highly acidic. To overcome this challenge, endolysins from a double extremophile ϕNS11 should be explored. As mentioned above, ϕNS11 infects a thermophilic bacterium that grows in an acidic environment (pH 3) [64,115]. Thus, endolysins derived from this phage are likely to resist the pH of gastric juice. Furthermore, it is not clear whether bacteria will develop resistance, in the near future, to endolysins, as they have to antibiotics. Additionally, the efficacy of ϕNS11 endolysins against endospores also needs to be explored.
Author Contributions
Conceptualization, H.L. and E.T.; Writing—Original Draft Preparation, H.L. and E.T.; Writing—review and editing, H.L., M.K. and E.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by start-up funding from the School of Veterinary Medicine at Texas Tech University and by grant number 7R15AI146982–02 from the US National Institutes of Health (National Institute of Allergy and Infectious Diseases). The content is solely the responsibility of the authors and does not necessarily represent the views of the National Institutes of Health.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
Ebenezer Tumban is a co-inventor of an L2-bacteriophage VLP-related patent managed by the University of New Mexico in accordance with its conflict of interest policies.
References
- Clokie, M.R.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Keen, E.C. A century of phage research: Bacteriophages and the shaping of modern biology. BioEssays News Rev. Mol. Cell. Dev. Biol. 2015, 37, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Comeau, A.M.; Hatfull, G.F.; Krisch, H.M.; Lindell, D.; Mann, N.H.; Prangishvili, D. Exploring the prokaryotic virosphere. Res. Microbiol. 2008, 159, 306–313. [Google Scholar] [CrossRef] [PubMed]
- João, J.; Lampreia, J.; Prazeres, D.M.F.; Azevedo, A.M. Manufacturing of bacteriophages for therapeutic applications. Biotechnol. Adv. 2021, 49, 107758. [Google Scholar] [CrossRef]
- Kumar, S.; Pattnaik, R.; Sahu, P.S.; Bhattacharya, B. Bacteriophage and Virus Engineering. In Microbial Engineering for Therapeutics; Suar, M., Misra, N., Dash, C., Eds.; Springer Nature: Singapore, 2022; pp. 123–144. [Google Scholar]
- Salmond, G.P.; Fineran, P.C. A century of the phage: Past, present and future. Nat. Rev. Microbiol. 2015, 13, 777–786. [Google Scholar] [CrossRef]
- Gould, L.H.; Kline, J.; Monahan, C.; Vierk, K. Outbreaks of Disease Associated with Food Imported into the United States, 1996–2014. Emerg. Infect. Dis. 2017, 23, 525–528. [Google Scholar] [CrossRef]
- Sarno, E.; Pezzutto, D.; Rossi, M.; Liebana, E.; Rizzi, V. A Review of Significant European Foodborne Outbreaks in the Last Decade. J. Food Prot. 2021, 84, 2059–2070. [Google Scholar] [CrossRef]
- USDA. Salmonella Accounted for Nearly 80 Percent of Pathogen Violations in U.S. Food Imports from 2002 to 2019. Available online: https://www.ers.usda.gov/data-products/chart-gallery/gallery/chart-detail/?chartId=103529 (accessed on 18 March 2022).
- Wu, D.; Elliott, C.; Wu, Y. Food Safety Strategies: The One Health Approach to Global Challenges and China’s Actions. China CDC Wkly. 2021, 3, 507–513. [Google Scholar] [CrossRef]
- World Health Organization. One Health. Available online: https://www.who.int/europe/initiatives/one-health (accessed on 28 March 2023).
- Zalewska-Piatek, B.; Piatek, R. Bacteriophages as Potential Tools for Use in Antimicrobial Therapy and Vaccine Development. Pharmaceuticals 2021, 14, 331. [Google Scholar] [CrossRef]
- Gigante, A.; Atterbury, R.J. Veterinary use of bacteriophage therapy in intensively-reared livestock. Virol. J. 2019, 16, 155. [Google Scholar] [CrossRef]
- Moghadam, M.T.; Amirmozafari, N.; Shariati, A.; Hallajzadeh, M.; Mirkalantari, S.; Khoshbayan, A.; Jazi, F.M. How Phages Overcome the Challenges of Drug Resistant Bacteria in Clinical Infections. Infect. Drug Resist. 2020, 13, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage therapy: An alternative to antibiotics in the age of multi-drug resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162–173. [Google Scholar] [CrossRef]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and Use of Personalized Bacteriophage-Based Therapeutic Cocktails To Treat a Patient with a Disseminated Resistant Acinetobacter baumannii Infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef] [PubMed]
- Ooi, M.L.; Drilling, A.J.; Morales, S.; Fong, S.; Moraitis, S.; Macias-Valle, L.; Vreugde, S.; Psaltis, A.J.; Wormald, P.J. Safety and Tolerability of Bacteriophage Therapy for Chronic Rhinosinusitis Due to Staphylococcus aureus. JAMA Otolaryngol.-Head Neck Surg. 2019, 145, 723–729. [Google Scholar] [CrossRef]
- Chan, B.K.; Turner, P.E.; Kim, S.; Mojibian, H.R.; Elefteriades, J.A.; Narayan, D. Phage treatment of an aortic graft infected with Pseudomonas aeruginosa. Evol. Med. Public Health 2018, 2018, 60–66. [Google Scholar] [CrossRef]
- Jennes, S.; Merabishvili, M.; Soentjens, P.; Pang, K.W.; Rose, T.; Keersebilck, E.; Soete, O.; François, P.-M.; Teodorescu, S.; Verween, G.; et al. Use of bacteriophages in the treatment of colistin-only-sensitive Pseudomonas aeruginosa septicaemia in a patient with acute kidney injury—A case report. Crit. Care 2017, 21, 129. [Google Scholar] [CrossRef] [PubMed]
- Mosimann, S.; Desiree, K.; Ebner, P. Efficacy of phage therapy in poultry: A systematic review and meta-analysis. Poult. Sci. 2021, 100, 101472. [Google Scholar] [CrossRef]
- Loponte, R.; Pagnini, U.; Iovane, G.; Pisanelli, G. Phage Therapy in Veterinary Medicine. Antibiotics 2021, 10, 421. [Google Scholar] [CrossRef]
- Nakayinga, R.; Makumi, A.; Tumuhaise, V.; Tinzaara, W. Xanthomonas bacteriophages: A review of their biology and biocontrol applications in agriculture. BMC Microbiol. 2021, 21, 291. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Ye, M.; Zhang, X.T.; Sun, M.M.; Zhang, Z.Y.; Chao, H.Z.; Huang, D.; Wan, J.Z.; Zhang, S.T.; Jiang, X.; et al. Comparing polyvalent bacteriophage and bacteriophage cocktails for controlling antibiotic-resistant bacteria in soil-plant system. Sci. Total Environ. 2019, 657, 918–925. [Google Scholar] [CrossRef]
- Holtappels, D.; Fortuna, K.; Lavigne, R.; Wagemans, J. The future of phage biocontrol in integrated plant protection for sustainable crop production. Curr. Opin. Biotechnol. 2021, 68, 60–71. [Google Scholar] [CrossRef]
- Ramos-Vivas, J.; Elexpuru-Zabaleta, M.; Samano, M.L.; Barrera, A.P.; Forbes-Hernández, T.Y.; Giampieri, F.; Battino, M. Phages and Enzybiotics in Food Biopreservation. Molecules 2021, 26, 5138. [Google Scholar] [CrossRef] [PubMed]
- Tamakoshi, M.; Murakami, A.; Sugisawa, M.; Tsuneizumi, K.; Takeda, S.; Saheki, T.; Izumi, T.; Akiba, T.; Mitsuoka, K.; Toh, H.; et al. Genomic and proteomic characterization of the large Myoviridae bacteriophage ϕTMA of the extreme thermophile Thermus thermophilus. Bacteriophage 2011, 1, 152–164. [Google Scholar] [CrossRef]
- Zhou, Y.Q.; Li, L.; Han, K.O.; Wang, L.P.; Cao, Y.J.; Ma, D.X.; Wang, X.Y. A Polyvalent Broad-Spectrum Escherichia Phage Tequatrovirus EP01 Capable of Controlling Salmonella and Escherichia coli Contamination in Foods. Viruses 2022, 14, 286. [Google Scholar] [CrossRef]
- Liu, A.; Liu, Y.; Peng, L.; Cai, X.; Shen, L.; Duan, M.; Ning, Y.; Liu, S.; Li, C.; Liu, Y.; et al. Characterization of the narrow-spectrum bacteriophage LSE7621 towards Salmonella Enteritidis and its biocontrol potential on lettuce and tofu. LWT 2020, 118, 108791. [Google Scholar] [CrossRef]
- Obeso, J.M.; Martínez, B.; Rodríguez, A.; García, P. Lytic activity of the recombinant staphylococcal bacteriophage PhiH5 endolysin active against Staphylococcus aureus in milk. Int. J. Food Microbiol. 2008, 128, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Donovan, D.M.; Dong, S.; Garrett, W.; Rousseau, G.M.; Moineau, S.; Pritchard, D.G. Peptidoglycan hydrolase fusions maintain their parental specificities. Appl. Environ. Microbiol. 2006, 72, 2988–2996. [Google Scholar] [CrossRef]
- Zhai, L.; Yadav, R.; Kunda, N.K.; Anderson, D.; Bruckner, E.; Miller, E.K.; Basu, R.; Muttil, P.; Tumban, E. Oral immunization with bacteriophage MS2-L2 VLPs protects against oral and genital infection with multiple HPV types associated with head & neck cancers and cervical cancer. Antivir. Res. 2019, 166, 56–65. [Google Scholar] [PubMed]
- Stander, J.; Chabeda, A.; Rybicki, E.P.; Meyers, A.E. A Plant-Produced Virus-Like Particle Displaying Envelope Protein Domain III Elicits an Immune Response Against West Nile Virus in Mice. Front. Plant Sci. 2021, 12, 738619. [Google Scholar] [CrossRef]
- Basu, R.; Zhai, L.K.; Contreras, A.; Tumban, E. Immunization with phage virus-like particles displaying Zika virus potential B-cell epitopes neutralizes Zika virus infection of monkey kidney cells. Vaccine 2018, 36, 1256–1264. [Google Scholar] [CrossRef]
- Chen, J.H.; Alcaine, S.D.; Jiang, Z.W.; Rotello, V.M.; Nugen, S.R. Detection of Escherichia coli in Drinking Water Using T7 Bacteriophage-Conjugated Magnetic Probe. Anal. Chem. 2015, 87, 8977–8984. [Google Scholar] [CrossRef]
- Bayat, F.; Didar, T.; Hosseinidoust, Z. Emerging investigator series: Bacteriophages as nano engineering tools for quality monitoring and pathogen detection in water and wastewater. Environ. Sci. Nano 2021, 8, 367–389. [Google Scholar] [CrossRef]
- Ripp, S.; Jegier, P.; Johnson, C.M.; Brigati, J.R.; Sayler, G.S. Bacteriophage-amplified bioluminescent sensing of Escherichia coli O157:H7. Anal. Bioanal. Chem. 2008, 391, 507–514. [Google Scholar] [CrossRef]
- Boyaci, I.H.; Aguilar, Z.P.; Hossain, M.; Halsall, H.B.; Seliskar, C.J.; Heineman, W.R. Amperometric determination of live Escherichia coli using antibody-coated paramagnetic beads. Anal. Bioanal. Chem. 2005, 382, 1234–1241. [Google Scholar] [CrossRef]
- Das, S.; Yau, M.K.; Noble, J.; de Pascalis, L.; Finn, M.G. Transport of Molecular Cargo by Interaction with Virus-Like Particle RNA. Angew. Chem. Int. Ed. Engl. 2022, 61, e202111687. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Nolte, R.J.; Cornelissen, J.J. Virus-based nanocarriers for drug delivery. Adv. Drug Deliv. Rev. 2012, 64, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Sokullu, E.; Soleymani Abyaneh, H.; Gauthier, M.A. Plant/Bacterial Virus-Based Drug Discovery, Drug Delivery, and Therapeutics. Pharmaceutics 2019, 11, 211. [Google Scholar] [CrossRef] [PubMed]
- Pumpens, P.; Renhofa, R.; Dishlers, A.; Kozlovska, T.; Ose, V.; Pushko, P.; Tars, K.; Grens, E.; Bachmann, M.F. The True Story and Advantages of RNA Phage Capsids as Nanotools. Intervirology 2016, 59, 74–110. [Google Scholar] [CrossRef] [PubMed]
- Jończyk, E.; Kłak, M.; Międzybrodzki, R.; Górski, A. The influence of external factors on bacteriophages—Review. Folia Microbiol. 2011, 56, 191–200. [Google Scholar] [CrossRef]
- Son, B.; Yun, J.; Lim, J.A.; Shin, H.; Heu, S.; Ryu, S. Characterization of LysB4, an endolysin from the Bacillus cereus-infecting bacteriophage B4. BMC Microbiol. 2012, 12, 33. [Google Scholar] [CrossRef]
- Caldeira, J.C.; Peabody, D.S. Thermal stability of RNA phage virus-like particles displaying foreign peptides. J. Nanobiotechnol. 2011, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Zhai, L.; Tumban, E. Virus-like Particle-Based L2 Vaccines against HPVs: Where Are We Today? Viruses 2019, 12, 18. [Google Scholar] [CrossRef]
- GlaxoSmithKline. Package Insert & Patient Information of Human Papillomavirus Bivalent (Types 16 and 18) Vaccine–Cervarix. Available online: https://www.fda.gov/media/78013/download (accessed on 23 March 2023).
- Merck Sharp & Dohme LLC. Package Insert of Human Papillomavirus 9-Valent Vaccine—Gardasil 9. Available online: https://www.fda.gov/media/90064/download (accessed on 28 April 2023).
- Gara, N.; Abdalla, A.; Rivera, E.; Zhao, X.; Werner, J.M.; Liang, T.J.; Hoofnagle, J.H.; Rehermann, B.; Ghany, M.G. Durability of antibody response against hepatitis B virus in healthcare workers vaccinated as adults. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2015, 60, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Y.; Chen, Z.P.; Wang, S.Y.; Pan, H.R.; Wang, Z.F.; Zhang, Q.F.; Shen, L.Z.; Zheng, X.P.; Yan, C.F.; Lu, M.; et al. Safety and immunogenicity of hepatitis E vaccine in elderly people older than 65 years. Vaccine 2019, 37, 4581–4586. [Google Scholar] [CrossRef] [PubMed]
- Prentoe, J.; Janitzek, C.M.; Velázquez-Moctezuma, R.; Soerensen, A.; Jørgensen, T.; Clemmensen, S.; Soroka, V.; Thrane, S.; Theander, T.; Nielsen, M.A.; et al. Two-component vaccine consisting of virus-like particles displaying hepatitis C virus envelope protein 2 oligomers. NPJ Vaccines 2022, 7, 148. [Google Scholar] [CrossRef]
- Tumban, E.; Peabody, J.; Tyler, M.; Peabody, D.S.; Chackerian, B. VLPs displaying a single L2 epitope induce broadly cross-neutralizing antibodies against human papillomavirus. PLoS ONE 2012, 7, e49751. [Google Scholar] [CrossRef]
- Basu, R.; Zhai, L.; Rosso, B.; Tumban, E. Bacteriophage Qβ virus-like particles displaying Chikungunya virus B-cell epitopes elicit high-titer E2 protein antibodies but fail to neutralize a Thailand strain of Chikungunya virus. Vaccine 2020, 38, 2542–2550. [Google Scholar] [CrossRef]
- Guarascio, F. Poorer Nations Reject over 100 mln COVID-19 Vaccine Doses as Many Near Expiry. Available online: https://www.reuters.com/business/healthcare-pharmaceuticals/more-than-100-million-covid-19-vaccines-rejected-by-poorer-nations-dec-unicef-2022-01-13/ (accessed on 14 January 2022).
- Poorer Nations Forced to Dump Close-to-Expiry COVID Vaccines. Available online: https://www.aljazeera.com/news/2022/1/13/poorer-nations-dump-close-to-expiry-covid-vaccines-unicef (accessed on 13 January 2022).
- Edstam, J.S.; Dulmaa, N.; Tsendjav, O.; Dambasuren, B.; Densmaa, B. Exposure of hepatitis B vaccine to freezing temperatures during transport to rural health centers in Mongolia. Prev. Med. 2004, 39, 384–388. [Google Scholar] [CrossRef]
- Matthias, D.M.; Robertson, J.; Garrison, M.M.; Newland, S.; Nelson, C. Freezing temperatures in the vaccine cold chain: A systematic literature review. Vaccine 2007, 25, 3980–3986. [Google Scholar] [CrossRef]
- Kasman, L.M. Barriers to coliphage infection of commensal intestinal flora of laboratory mice. Virol. J. 2005, 2, 34. [Google Scholar] [CrossRef]
- Łusiak-Szelachowska, M.; Zaczek, M.; Weber-Dąbrowska, B.; Międzybrodzki, R.; Kłak, M.; Fortuna, W.; Letkiewicz, S.; Rogóż, P.; Szufnarowski, K.; Jończyk-Matysiak, E.; et al. Phage neutralization by sera of patients receiving phage therapy. Viral Immunol. 2014, 27, 295–304. [Google Scholar] [CrossRef]
- Majewska, J.; Kaźmierczak, Z.; Lahutta, K.; Lecion, D.; Szymczak, A.; Miernikiewicz, P.; Drapała, J.; Harhala, M.; Marek-Bukowiec, K.; Jędruchniewicz, N.; et al. Induction of Phage-Specific Antibodies by Two Therapeutic Staphylococcal Bacteriophages Administered per os. Front. Immunol. 2019, 10, 2607. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Sun, Y.; Zhao, R. Establishment of MicroRNA delivery system by PP7 bacteriophage-like particles carrying cell-penetrating peptide. J. Biosci. Bioeng. 2017, 124, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, I.; Yamashita, N.; Yokota, A. Isolation and characterization of bacteriophage induced from a new isolate of Thermus aquaticus. Microbiol. Cult. Coll. Off. Publ. Jpn. Soc. Cult. Collect. 1995, 11, 133–138. [Google Scholar]
- Yu, M.X.; Slater, M.R.; Ackermann, H.W. Isolation and characterization of Thermus bacteriophages. Arch. Virol. 2006, 151, 663–679. [Google Scholar] [CrossRef]
- Liu, B.; Wu, S.; Song, Q.; Zhang, X.; Xie, L. Two novel bacteriophages of thermophilic bacteria isolated from deep-sea hydrothermal fields. Curr. Microbiol. 2006, 53, 163–166. [Google Scholar] [CrossRef]
- Łubkowska, B.; Jeżewska-Frąckowiak, J.; Sobolewski, I.; Skowron, P.M. Bacteriophages of Thermophilic ‘Bacillus Group’ Bacteria—A Review. Microorganisms 2021, 9, 1522. [Google Scholar] [CrossRef]
- Nagayoshi, Y.; Kumagae, K.; Mori, K.; Tashiro, K.; Nakamura, A.; Fujino, Y.; Hiromasa, Y.; Iwamoto, T.; Kuhara, S.; Ohshima, T.; et al. Physiological Properties and Genome Structure of the Hyperthermophilic Filamentous Phage φOH3 Which Infects Thermus thermophilus HB8. Front. Microbiol. 2016, 7, 50. [Google Scholar] [CrossRef]
- Jalasvuori, M.; Jaatinen, S.T.; Laurinavicius, S.; Ahola-Iivarinen, E.; Kalkkinen, N.; Bamford, D.H.; Bamford, J.K. The closest relatives of icosahedral viruses of thermophilic bacteria are among viruses and plasmids of the halophilic archaea. J. Virol. 2009, 83, 9388–9397. [Google Scholar] [CrossRef]
- Cheepudom, J.; Lee, C.C.; Cai, B.; Meng, M. Isolation, characterization, and complete genome analysis of P1312, a thermostable bacteriophage that infects Thermobifida fusca. Front. Microbiol. 2015, 6, 959. [Google Scholar] [CrossRef]
- Höög, J.O.; Eklund, H.; Jörnvall, H. A single-residue exchange gives human recombinant beta beta alcohol dehydrogenase gamma gamma isozyme properties. Eur. J. Biochem. 1992, 205, 519–526. [Google Scholar] [CrossRef]
- Pederson, D.M.; Welsh, L.C.; Marvin, D.A.; Sampson, M.; Perham, R.N.; Yu, M.; Slater, M.R. The protein capsid of filamentous bacteriophage PH75 from Thermus thermophilus. J. Mol. Biol. 2001, 309, 401–421. [Google Scholar] [CrossRef] [PubMed]
- Hjorleifsdottir, S.; Aevarsson, A.; Hreggvidsson, G.O.; Fridjonsson, O.H.; Kristjansson, J.K. Isolation, growth and genome of the Rhodothermus RM378 thermophilic bacteriophage. Extremophiles 2014, 18, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhou, F.; Wu, S.; Xu, Y.; Zhang, X. Genomic and proteomic characterization of a thermophilic Geobacillus bacteriophage GBSV1. Res. Microbiol. 2009, 160, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Jaatinen, S.T.; Happonen, L.J.; Laurinmäki, P.; Butcher, S.J.; Bamford, D.H. Biochemical and structural characterisation of membrane-containing icosahedral dsDNA bacteriophages infecting thermophilic Thermus thermophilus. Virology 2008, 379, 10–19. [Google Scholar] [CrossRef]
- Minakhin, L.; Goel, M.; Berdygulova, Z.; Ramanculov, E.; Florens, L.; Glazko, G.; Karamychev, V.N.; Slesarev, A.I.; Kozyavkin, S.A.; Khromov, I.; et al. Genome comparison and proteomic characterization of Thermus thermophilus bacteriophages P23-45 and P74-26: Siphoviruses with triplex-forming sequences and the longest known tails. J. Mol. Biol. 2008, 378, 468–480. [Google Scholar] [CrossRef]
- Stone, N.P.; Demo, G.; Agnello, E.; Kelch, B.A. Principles for enhancing virus capsid capacity and stability from a thermophilic virus capsid structure. Nat. Commun. 2019, 10, 4471. [Google Scholar] [CrossRef]
- Sakaki, Y.; Yamada, K.; Oshima, M.; Oshima, T. Bacteriophage phiNS11: A lipid-containing phage of acidophilic thermophilic bacteria. II. Purification and some properties of the phage. J. Biochem. 1977, 82, 1451–1456. [Google Scholar] [CrossRef]
- Skowron, P.M.; Kropinski, A.M.; Zebrowska, J.; Janus, L.; Szemiako, K.; Czajkowska, E.; Maciejewska, N.; Skowron, M.; Łoś, J.; Łoś, M.; et al. Correction: Sequence, genome organization, annotation and proteomics of the thermophilic, 47.7-kb Geobacillus stearothermophilus bacteriophage TP-84 and its classification in the new Tp84virus genus. PLoS ONE 2018, 13, e0196798. [Google Scholar] [CrossRef]
- Blondal, T.; Thorisdottir, A.; Unnsteinsdottir, U.; Hjorleifsdottir, S.; Aevarsson, A.; Ernstsson, S.; Fridjonsson, O.H.; Skirnisdottir, S.; Wheat, J.O.; Hermannsdottir, A.G.; et al. Isolation and characterization of a thermostable RNA ligase 1 from a Thermus scotoductus bacteriophage TS2126 with good single-stranded DNA ligation properties. Nucleic Acids Res. 2005, 33, 135–142. [Google Scholar] [CrossRef]
- Kristjánsson, J.K.; Hjörleifsdóttir, S.; Marteinsson, V.T.; Alfredsson, G.A. Thermus scotoductus, sp. nov., a Pigment-Producing Thermophilic Bacterium from Hot Tap Water in Iceland and Including Thermus sp. X-1. Syst. Appl. Microbiol. 1994, 17, 44–50. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, X. Deep-sea thermophilic Geobacillus bacteriophage GVE2 transcriptional profile and proteomic characterization of virions. Appl. Microbiol. Biotechnol. 2008, 80, 697–707. [Google Scholar] [CrossRef]
- Lin, L.; Han, J.; Ji, X.; Hong, W.; Huang, L.; Wei, Y. Isolation and characterization of a new bacteriophage MMP17 from Meiothermus. Extremophiles 2011, 15, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Naryshkina, T.; Liu, J.; Florens, L.; Swanson, S.K.; Pavlov, A.R.; Pavlova, N.V.; Inman, R.; Minakhin, L.; Kozyavkin, S.A.; Washburn, M.; et al. Thermus thermophilus bacteriophage phiYS40 genome and proteomic characterization of virions. J. Mol. Biol. 2006, 364, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Sakaki, Y.; Oshima, T. Isolation and characterization of a bacteriophage infectious to an extreme thermophile, Thermus thermophilus HB8. J. Virol. 1975, 15, 1449–1453. [Google Scholar] [CrossRef]
- Li, W.; Jing, Z.; Wang, S.; Li, Q.; Xing, Y.; Shi, H.; Li, S.; Hong, Z. P22 virus-like particles as an effective antigen delivery nanoplatform for cancer immunotherapy. Biomaterials 2021, 271, 120726. [Google Scholar] [CrossRef]
- Zablocki, O.; van Zyl, L.; Trindade, M. Biogeography and taxonomic overview of terrestrial hot spring thermophilic phages. Extremophiles 2018, 22, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, I.; Yanase, H. The Genomic Structure of Thermus Bacteriophage ϕIN93. J. Biochem. 2009, 146, 775–785. [Google Scholar] [CrossRef]
- Sattar, S.; Ullah, I.; Khanum, S.; Bailie, M.; Shamsi, B.; Ahmed, I.; Shah, S.T.A.; Javed, S.; Ghafoor, A.; Pervaiz, A.; et al. Phenotypic characterization and genome analysis of a novel Salmonella Typhimurium phage having unique tail fiber genes. Sci. Rep. 2022, 12, 5732. [Google Scholar] [CrossRef]
- Conroy, R.S.; Koretsky, A.P.; Moreland, J. Lambda exonuclease digestion of CGG trinucleotide repeats. Eur. Biophys. J. 2010, 39, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Ljunger, C.; Edebo, M. The influence of inorganic ions on the heat stability of a thermophilic bacteriophage. Physiol. Plant. 1972, 27, 182–186. [Google Scholar] [CrossRef]
- Bassel, A.; Shaw, M.; Campbell, L.L. Dissociation by chelating agents and substructure of the thermophilic bacteriophage TP84. J. Virol. 1971, 7, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Moser, M.J.; DiFrancesco, R.A.; Gowda, K.; Klingele, A.J.; Sugar, D.R.; Stocki, S.; Mead, D.A.; Schoenfeld, T.W. Thermostable DNA polymerase from a viral metagenome is a potent RT-PCR enzyme. PLoS ONE 2012, 7, e38371. [Google Scholar] [CrossRef] [PubMed]
- Van Pelt-Verkuil, E.; van Belkum, A.; Hays, J.P. Principles and Technical Aspects of PCR Amplification; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Heller, R.C.; Chung, S.; Crissy, K.; Dumas, K.; Schuster, D.; Schoenfeld, T.W. Engineering of a thermostable viral polymerase using metagenome-derived diversity for highly sensitive and specific RT-PCR. Nucleic Acids Res. 2019, 47, 3619–3630. [Google Scholar] [CrossRef]
- Chander, Y.; Koelbl, J.; Puckett, J.; Moser, M.J.; Klingele, A.J.; Liles, M.R.; Carrias, A.; Mead, D.A.; Schoenfeld, T.W. A novel thermostable polymerase for RNA and DNA loop-mediated isothermal amplification (LAMP). Front. Microbiol. 2014, 5, 395. [Google Scholar] [CrossRef]
- CircLigase™ ssDNA Ligase and CircLigase™ II ssDNA Ligase, Lucigen. Available online: https://us.vwr.com/store/product/22371898/circligasetm-ssdna-ligase-and-circligasetm-ii-ssdna-ligase-lucigen (accessed on 28 March 2023).
- Ren, H.; Xi, Y.; Li, Z.; Zhang, D.; Huang, F.; Fang, X.; Zhao, X.; Zhang, X.; Chen, A.; Chen, T.; et al. Novel target capture DNA library preparation method using CircLigase-mediated hook ligation. New Biotechnol. 2020, 59, 44–50. [Google Scholar] [CrossRef]
- Blondal, T.; Hjorleifsdottir, S.H.; Fridjonsson, O.F.; Aevarsson, A.; Skirnisdottir, S.; Hermannsdottir, A.G.; Hreggvidsson, G.O.; Smith, A.V.; Kristjansson, J.K. Discovery and characterization of a thermostable bacteriophage RNA ligase homologous to T4 RNA ligase 1. Nucleic Acids Res. 2003, 31, 7247–7254. [Google Scholar] [CrossRef]
- Lama, L.; Ryan, K. Adenylylation of small RNA sequencing adapters using the TS2126 RNA ligase I. RNA 2016, 22, 155–161. [Google Scholar] [CrossRef]
- Swift, S.M.; Seal, B.S.; Garrish, J.K.; Oakley, B.B.; Hiett, K.; Yeh, H.Y.; Woolsey, R.; Schegg, K.M.; Line, J.E.; Donovan, D.M. A Thermophilic Phage Endolysin Fusion to a Clostridium perfringens-Specific Cell Wall Binding Domain Creates an Anti-Clostridium Antimicrobial with Improved Thermostability. Viruses 2015, 7, 3019–3034. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Vidal, J.E.; McClane, B.A.; Gurjar, A.A. Clostridium Perfringens Toxins Involved in Mammalian Veterinary Diseases. Open Toxinol. J. 2010, 2, 24–42. [Google Scholar] [CrossRef]
- CDC. Clostridium Perfringens Toxins Involved in Mammalian Veterinary Diseases. Available online: https://www.cdc.gov/foodsafety/diseases/clostridium-perfringens.html (accessed on 24 March 2023).
- Wang, F.; Xiong, Y.; Xiao, Y.; Han, J.; Deng, X.; Lin, L. MMPphg from the thermophilic Meiothermus bacteriophage MMP17 as a potential antimicrobial agent against both Gram-negative and Gram-positive bacteria. Virol. J. 2020, 17, 130. [Google Scholar] [CrossRef] [PubMed]
- Love, M.J.; Bhandari, D.; Dobson, R.C.J.; Billington, C. Potential for Bacteriophage Endolysins to Supplement or Replace Antibiotics in Food Production and Clinical Care. Antibiotics 2018, 7, 17. [Google Scholar] [CrossRef]
- Rissanen, I.; Grimes, J.M.; Pawlowski, A.; Mäntynen, S.; Harlos, K.; Bamford, J.K.; Stuart, D.I. Bacteriophage P23-77 capsid protein structures reveal the archetype of an ancient branch from a major virus lineage. Structure 2013, 21, 718–726. [Google Scholar] [CrossRef]
- Pawlowski, A.; Moilanen, A.M.; Rissanen, I.A.; Määttä, J.A.; Hytönen, V.P.; Ihalainen, J.A.; Bamford, J.K. The Minor Capsid Protein VP11 of Thermophilic Bacteriophage P23-77 Facilitates Virus Assembly by Using Lipid-Protein Interactions. J. Virol. 2015, 89, 7593–7603. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Anderson, D.; Bruckner, E.; Tumban, E. Novel expression of coat proteins from thermophilic bacteriophage ΦIN93 and evaluation for assembly into virus-like particles. Protein Expr. Purif. 2021, 187, 105932. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R. Efficacy of Mixed MS2-L2 VLPs against Six HPV Types and the Development & Evaluation of Viral Structural Proteins for Assembly into VLPs. Ph.D. Thesis, Michigan Technological University, Houghton, MI, USA, 2021. [Google Scholar]
- Morrissey, B.J.; Helgason, T.; Poppinga, L.; Fünfhaus, A.; Genersch, E.; Budge, G.E. Biogeography of Paenibacillus larvae, the causative agent of American foulbrood, using a new multilocus sequence typing scheme. Environ. Microbiol. 2015, 17, 1414–1424. [Google Scholar] [CrossRef]
- Schäfer, M.O. Introduction on Major Honey Bee Diseases in Europe. In Proceedings of the OIE Virtual Regional Workshop on Honey Bee Diseases in Asia and the Pacific, Online, 24 August 2021. [Google Scholar]
- Oliveira, A.; Leite, M.; Kluskens, L.D.; Santos, S.B.; Melo, L.D.; Azeredo, J. The First Paenibacillus larvae Bacteriophage Endolysin (PlyPl23) with High Potential to Control American Foulbrood. PLoS ONE 2015, 10, e0132095. [Google Scholar]
- Aanei, I.L.; Francis, M.B. Dual Surface Modification of Genome-Free MS2 Capsids for Delivery Applications. Methods Mol. Biol. 2018, 1776, 629–642. [Google Scholar]
- Aanei, I.L.; Glasgow, J.E.; Capehart, S.L.; Francis, M.B. Encapsulation of Negatively Charged Cargo in MS2 Viral Capsids. Methods Mol. Biol. 2018, 1776, 303–317. [Google Scholar]
- Ashley, C.E.; Carnes, E.C.; Phillips, G.K.; Durfee, P.N.; Buley, M.D.; Lino, C.A.; Padilla, D.P.; Phillips, B.; Carter, M.B.; Willman, C.L.; et al. Cell-specific delivery of diverse cargos by bacteriophage MS2 virus-like particles. ACS Nano 2011, 5, 5729–5745. [Google Scholar] [CrossRef]
- Cowan, P.W.; O’Grady, N. Treatment of the severe Class III edentulous patient with implant-retained prostheses. Quintessence Int. 1990, 21, 553–557. [Google Scholar] [PubMed]
- Manolova, V.; Flace, A.; Bauer, M.; Schwarz, K.; Saudan, P.; Bachmann, M.F. Nanoparticles target distinct dendritic cell populations according to their size. Eur. J. Immunol. 2008, 38, 1404–1413. [Google Scholar] [CrossRef] [PubMed]
- Sakaki, Y.; Oshima, T. A new lipid-containing phage infecting acidophilic thermophilic bacteria. Virology 1976, 75, 256–259. [Google Scholar] [CrossRef] [PubMed]
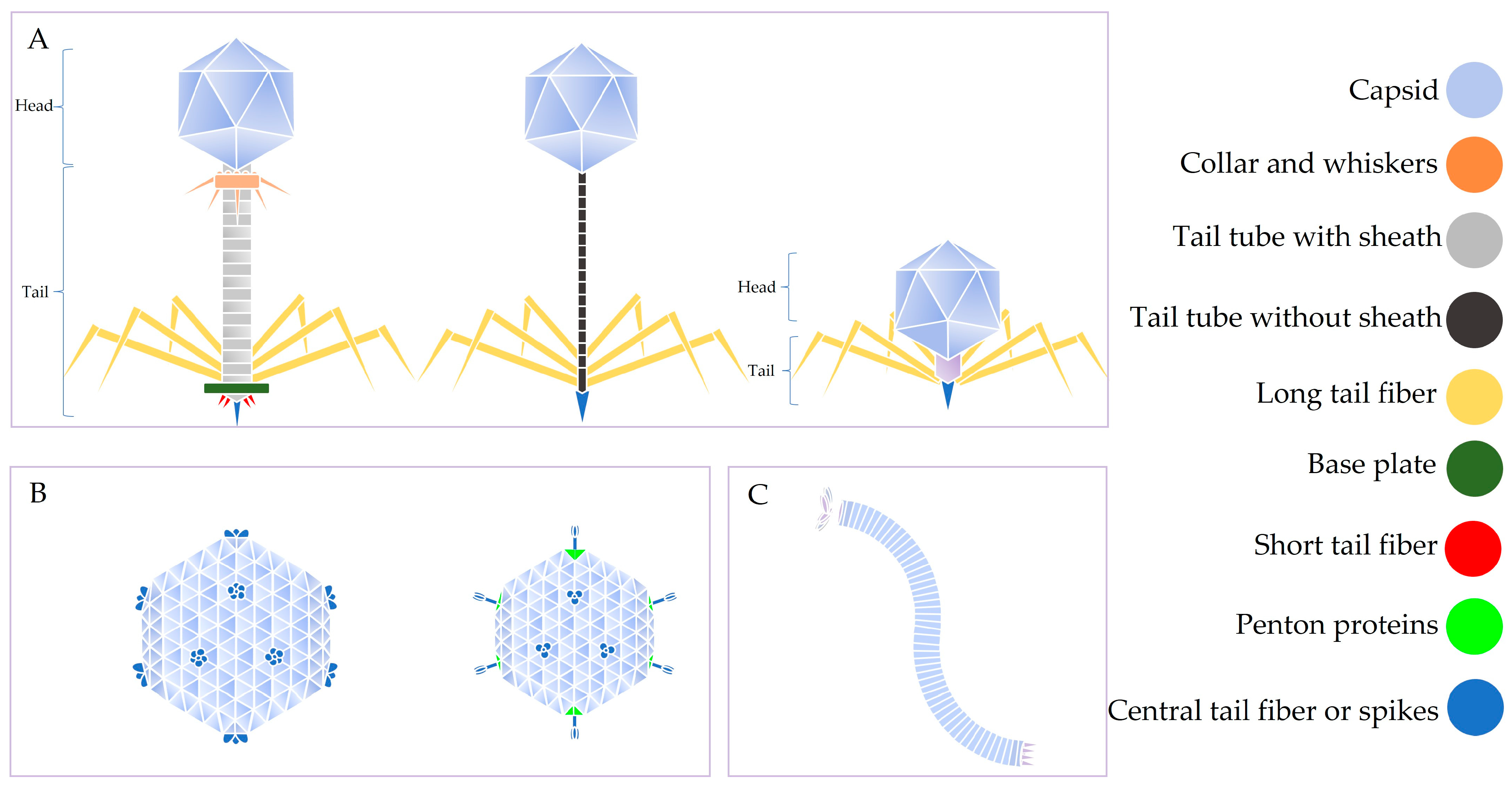
Figure 1.
Diversity in the morphology of thermophages. (A) Tailed phages, (B) tail-less phages, (C) filamentous phage. In thermophages that consist of a head and a tail, the tail tube and associated proteins (tail fiber, base plate, etc.) are used for the delivery of the genome to a bacterial cell, while the associated proteins are used for recognition and attachment of the thermophage to bacterial cells, respectively. The tail and associated proteins are not required for the capsid to assemble. The capsid (surrounding the genome) is composed of coat proteins and confers thermostability to thermophages. In some thermophages, the capsid is made up of only one type of coat protein, while in some thermophages, it is made up of more than one coat protein (major and minor capsid proteins). Expression of coat proteins derived from other tailed phages (i.e., mesophilic phages), in an expression vector, in a suitable bacterial host gives rise to VLPs [83].
Figure 1.
Diversity in the morphology of thermophages. (A) Tailed phages, (B) tail-less phages, (C) filamentous phage. In thermophages that consist of a head and a tail, the tail tube and associated proteins (tail fiber, base plate, etc.) are used for the delivery of the genome to a bacterial cell, while the associated proteins are used for recognition and attachment of the thermophage to bacterial cells, respectively. The tail and associated proteins are not required for the capsid to assemble. The capsid (surrounding the genome) is composed of coat proteins and confers thermostability to thermophages. In some thermophages, the capsid is made up of only one type of coat protein, while in some thermophages, it is made up of more than one coat protein (major and minor capsid proteins). Expression of coat proteins derived from other tailed phages (i.e., mesophilic phages), in an expression vector, in a suitable bacterial host gives rise to VLPs [83].

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, H.; Kheirvari, M.; Tumban, E. Potential Applications of Thermophilic Bacteriophages in One Health. Int. J. Mol. Sci. 2023, 24, 8222. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24098222
AMA Style
Liu H, Kheirvari M, Tumban E. Potential Applications of Thermophilic Bacteriophages in One Health. International Journal of Molecular Sciences. 2023; 24(9):8222. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24098222
Chicago/Turabian StyleLiu, Hong, Milad Kheirvari, and Ebenezer Tumban. 2023. "Potential Applications of Thermophilic Bacteriophages in One Health" International Journal of Molecular Sciences 24, no. 9: 8222. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24098222
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.