
Neurotrophins and Trk Neurotrophin Receptors in the Retina of Adult Killifish (Nothobranchius guentheri)
 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
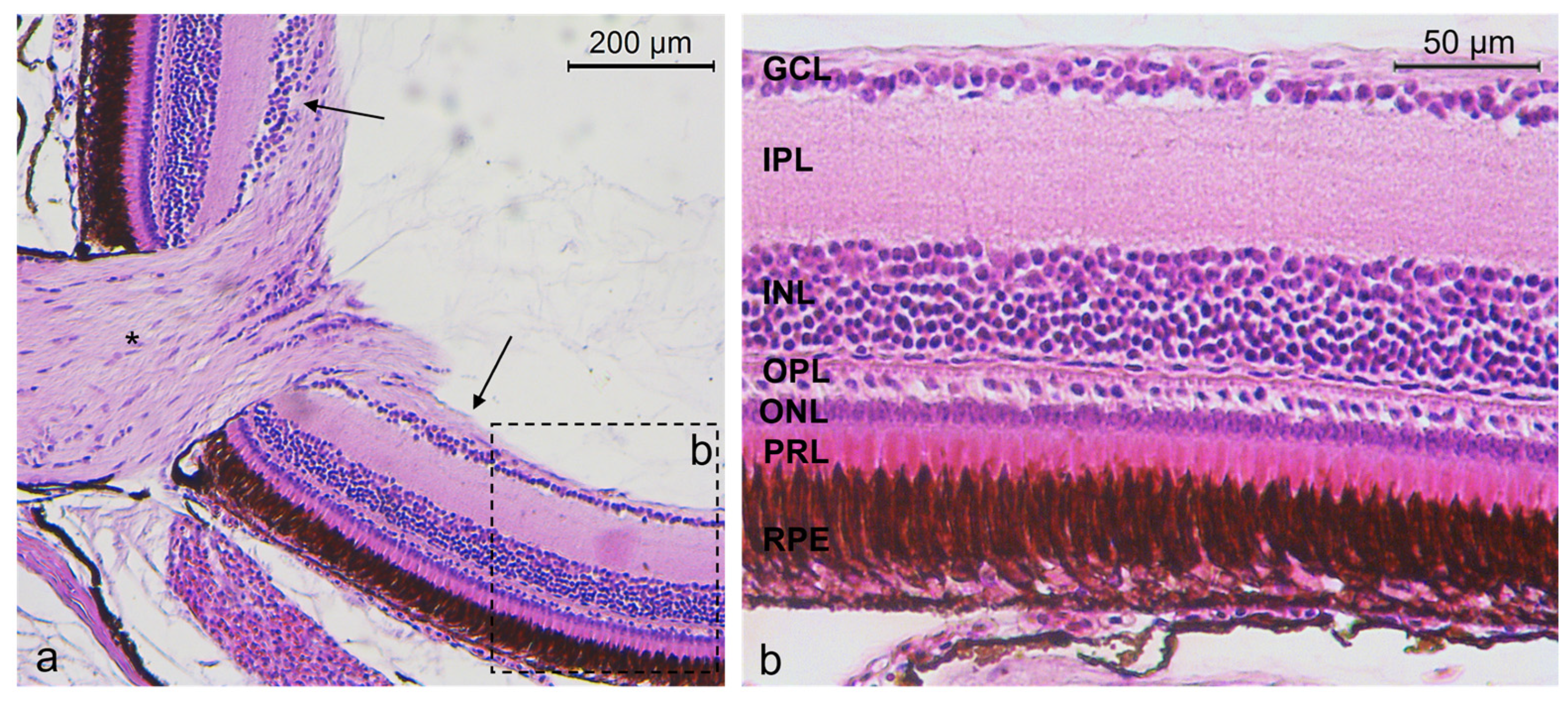
2.1. Histology of N. guentheri Retina
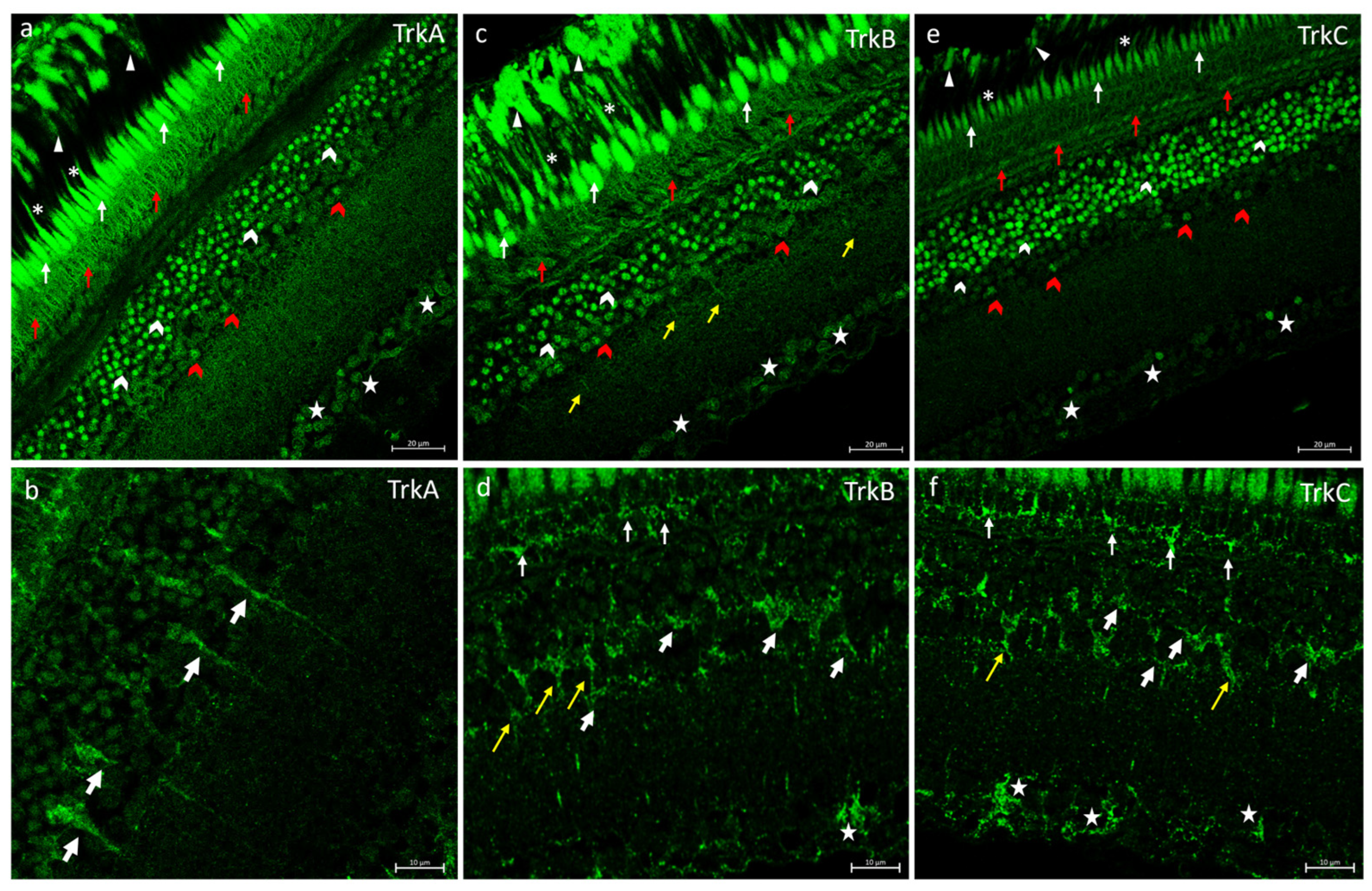
2.2. Trks Immunofluorescences in N. guentheri Retina
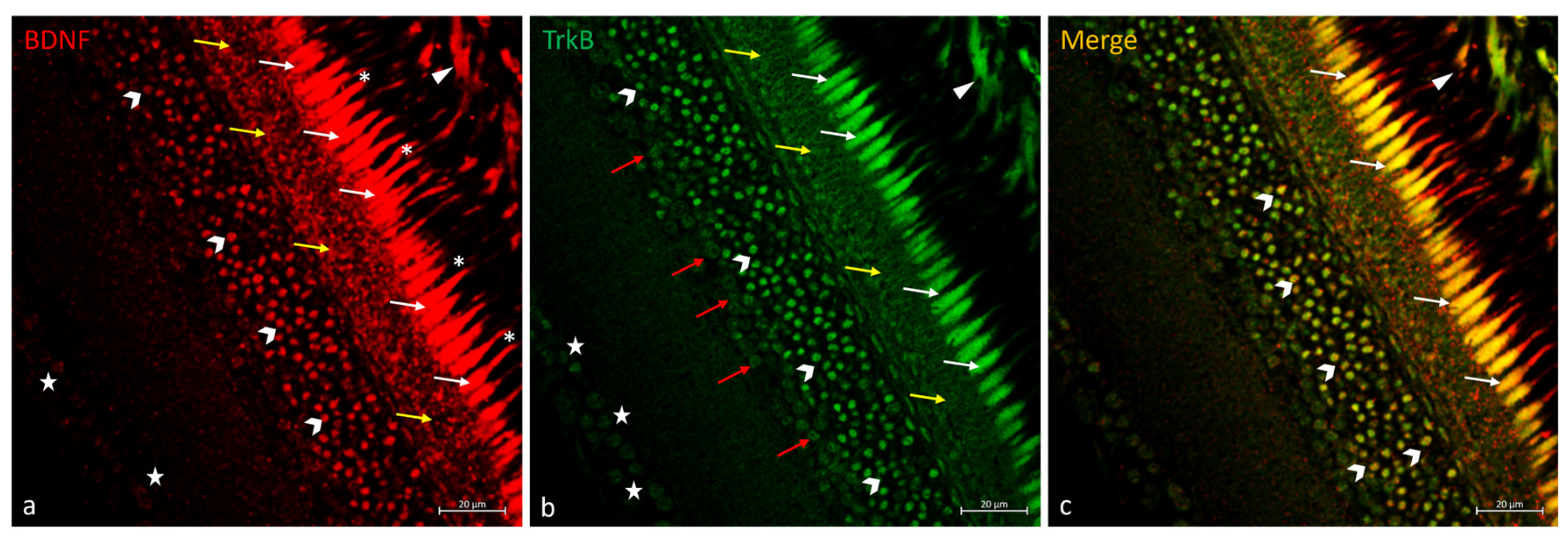
2.3. Double Immunofluorescences of BDNF and TrkB in N. guentheri Retina
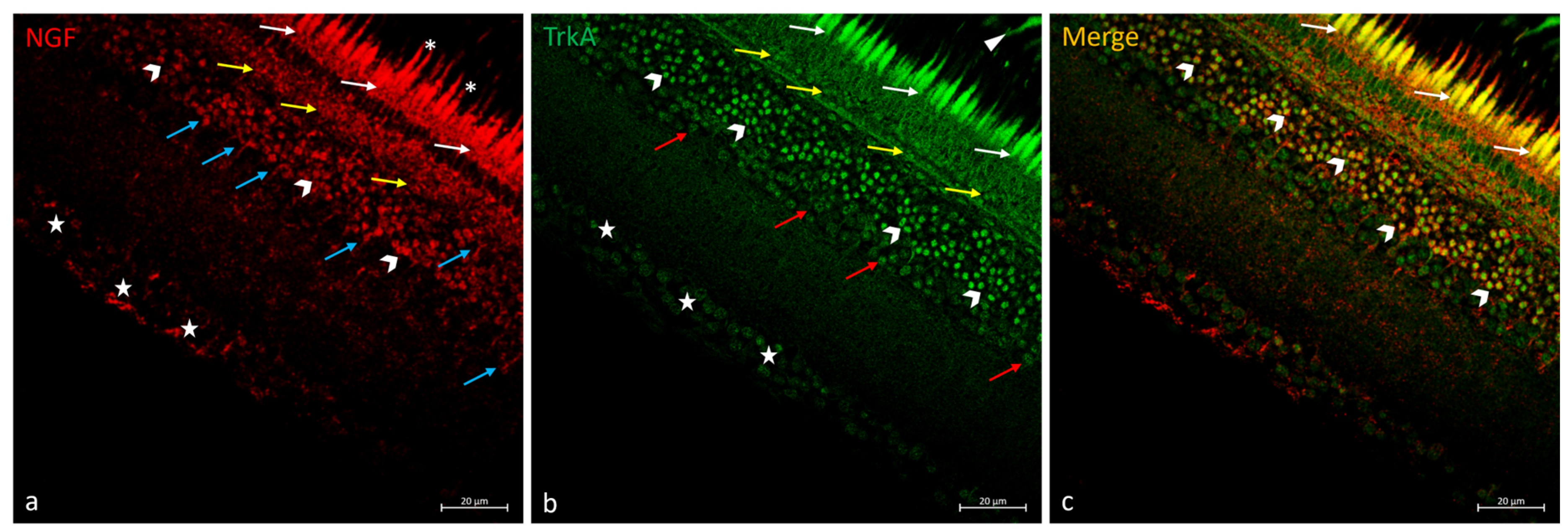
2.4. Double Immunofluorescences of NGF and TrkA in N. guentheri Retina
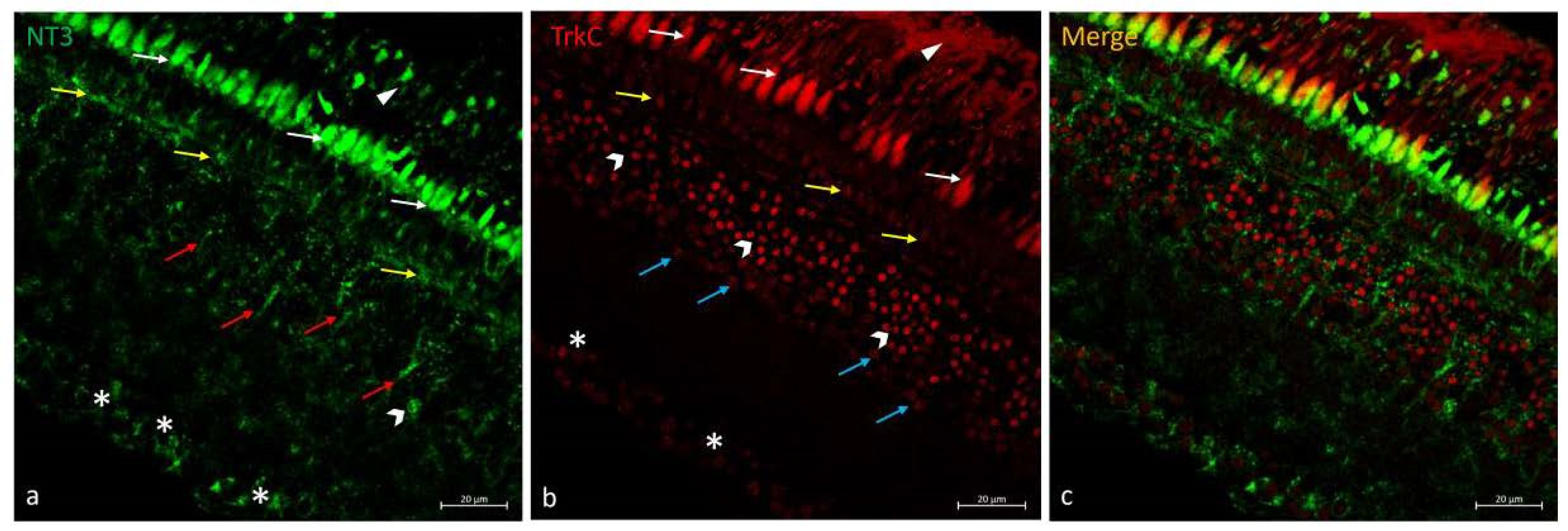
2.5. Double Immunofluorescences of NT-3 and TrkC in N. guentheri Retina
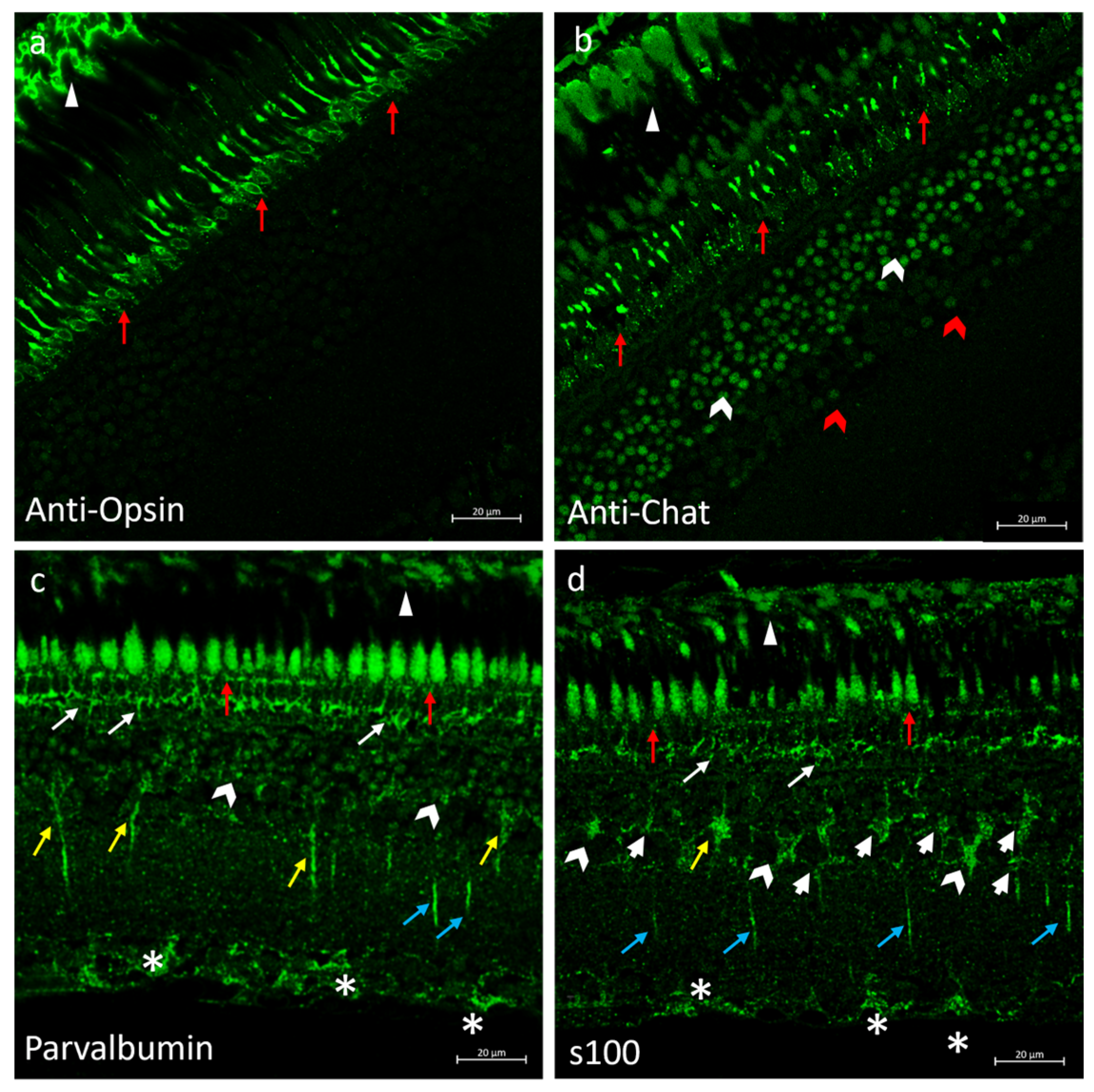
2.6. Immunofluorescences of Anti-Opsin, Anti-Chat, Parvalbumin and s100p in N. guentheri Retina
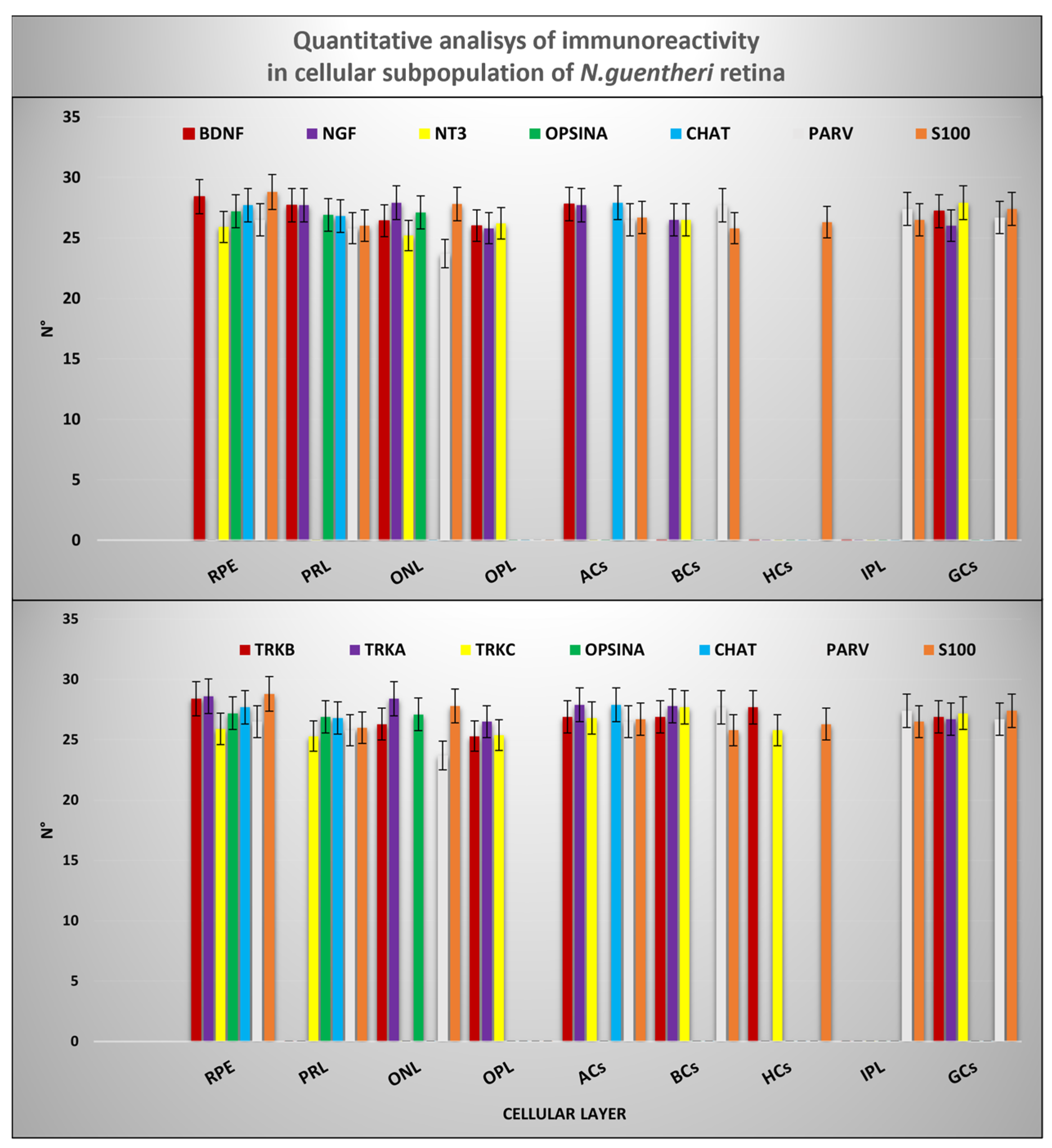
2.7. Quantitative Analysis
3. Discussion
4. Materials and Methods
4.1. Fish and Tissue Treatment
4.2. Optical Microscopy
4.3. Immunohistochemistry
4.3.1. TrkA, TrkB, TrkC, Anti-Opsin, Anti-Chat, Parvalbumin and s100p Immunofluorescences
4.3.2. NTs/Trks System Double Immunofluorescences
4.3.3. Confocal Laser Scanning Microscope
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Nikiforov-Nikishin, D.L.; Irkha, V.A.; Kochetkov, N.I.; Kalita, T.L.; Nikiforov-Nikishin, A.L.; Blokhin, E.E.; Antipov, S.S.; Makarenkov, D.A.; Zhavnerov, A.N.; Glebova, I.A.; et al. Some Aspects of Development and Histological Structure of the Visual System of Nothobranchius guentheri. Animals 2021, 11, 2755. [Google Scholar] [CrossRef]
- Ostrander, G.K.; Hopkins, J. The Laboratory Fish; Elsevier: Amsterdam, The Netherlands, 2000. [Google Scholar]
- Catania, S.; Germana, A.; Cabo, R.; Ochoa-Erena, F.; Guerrera, M.; Hannestad, J.; Represa, J.; Vega, J. Neurotrophin and Trk neurotrophin receptors in the inner ear of Salmo salar and Salmo trutta. J. Anat. 2007, 210, 78–88. [Google Scholar] [CrossRef] [PubMed]
- García-Suárez, O.; Germanà, A.; Hannestad, J.; Pérez-Pérez, M.; Esteban, I.; Naves, F.J.; Vega, J.A. Changes in the expression of the nerve growth factor receptors TrkA and p75LNGR in the rat thymus with ageing and increased nerve growth factor plasma levels. Cell Tissue Res. 2000, 301, 225–234. [Google Scholar] [CrossRef]
- García-Suárez, O.; Pérez-Pérez, M.; Germanà, A.; Esteban, I.; Germanà, G. Involvement of growth factors in thymic involution. Microsc. Res. Tech. 2003, 62, 514–523. [Google Scholar] [CrossRef]
- Germanà, A.; Marino, F.; Guerrera, M.C.; Campo, S.; de Girolamo, P.; Montalbano, G.; Germanà, G.P.; Ochoa-Erena, F.J.; Ciriaco, E.; Vega, J.A. Expression and distribution of S100 protein in the nervous system of the adult zebrafish (Danio rerio). Microsc. Res. Tech. 2008, 71, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Germana, A.; Catania, S.; Cavallaro, M.; González-Martínez, T.; Ciriaco, E.; Hannestad, J.; Vega, J. Immunohistochemical localization of BDNF-, TrkB-and TrkA-like proteins in the teleost lateral line system. J. Anat. 2002, 200, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D. Neurotrophic Factors: An Overview. In Neurotrophic Factors: Methods and Protocols; Skaper, S.D., Ed.; Springer: New York, NY, USA, 2018; pp. 1–17. [Google Scholar] [CrossRef]
- Aragona, M.; Porcino, C.; Guerrera, M.C.; Montalbano, G.; Laurà, R.; Cometa, M.; Levanti, M.; Abbate, F.; Cobo, T.; Capitelli, G.; et al. The BDNF/TrkB Neurotrophin System in the Sensory Organs of Zebrafish. Int. J. Mol. Sci. 2022, 23, 2621. [Google Scholar] [CrossRef] [PubMed]
- Aragona, M.; Porcino, C.; Guerrera, M.C.; Montalbano, G.; Laurà, R.; Levanti, M.; Abbate, F.; Cobo, T.; Capitelli, G.; Calapai, F.; et al. Localization of BDNF and Calretinin in Olfactory Epithelium and Taste Buds of Zebrafish (Danio rerio). Int. J. Mol. Sci. 2022, 23, 4696. [Google Scholar] [CrossRef] [PubMed]
- Hannestad, J.; Marino, F.; Germanà, A.; Catania, S.; Abbate, F.; Ciriaco, E.; Vega, J. Trk neurotrophin receptor-like proteins in the teleost Dicentrarchus labrax. Cell Tissue Res. 2000, 300, 1–9. [Google Scholar] [CrossRef]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Carmignoto, G.; Comelli, M.C.; Candeo, P.; Cavicchioli, L.; Yan, Q.; Merighi, A.; Maffei, L. Expression of NGF receptor and NGF receptor mRNA in the developing and adult rat retina. Exp. Neurol. 1991, 111, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Dergham, P.; Nedev, H.; Xu, J.; Galan, A.; Rivera, J.C.; ZhiHua, S.; Mehta, H.M.; Woo, S.B.; Sarunic, M.V.; et al. Chronic and Acute Models of Retinal Neurodegeneration TrkA Activity Are Neuroprotective whereas p75NTR Activity Is Neurotoxic through a Paracrine Mechanism. J. Biol. Chem. 2010, 285, 39392–39400. [Google Scholar] [CrossRef] [PubMed]
- Frade, J.M.; Barde, Y.-A. Microglia-Derived Nerve Growth Factor Causes Cell Death in the Developing Retina. Neuron 1998, 20, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Frade, J.M.; Bovolenta, P.; Rodríguez-Tébar, A. Neurotrophins and other growth factors in the generation of retinal neurons. Microsc. Res. Tech. 1999, 45, 243–251. [Google Scholar] [CrossRef]
- Cohen-Cory, S.; Lom, B. Neurotrophic regulation of retinal ganglion cell synaptic connectivity: From axons and dendrites to synapses. Int. J. Dev. Biol. 2004, 48, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Takano, M.; Horie, H.; Iijima, Y.; Dezawa, M.; Sawada, H.; Ishikawa, Y. Brain-derived Neurotrophic Factor Enhances Neurite Regeneration from Retinal Ganglion Cells in Aged Human Retina in vitro. Exp. Eye Res. 2002, 74, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.A.; Sparrow, J.; Cai, B.; Monroe, J.; Mikawa, T.; Hempstead, B.L. TrkB/BDNF signaling regulates photoreceptor progenitor cell fate decisions. Dev. Biol. 2006, 299, 455–465. [Google Scholar] [CrossRef]
- de la Rosa, E.J.; Arribas, A.; Frade, J.M.; Rodri’guez-Te’bar, A. Role of neurotrophins in the control of neural development: Neurotrophin-3 promotes both neuron differentiation and survival of cultured chick retinal cells. Neuroscience 1994, 58, 347–352. [Google Scholar] [CrossRef]
- Gleeson, M.; Connaughton, V.; Arneson, L.S. Induction of hyperglycaemia in zebrafish (Danio rerio) leads to morphological changes in the retina. Acta Diabetol. 2007, 44, 157–163. [Google Scholar] [CrossRef]
- Gestri, G.; Link, B.A.; Neuhauss, S.C. The visual system of zebrafish and its use to model human ocular diseases. Dev. Neurobiol. 2012, 72, 302–327. [Google Scholar] [CrossRef]
- Seritrakul, P.; Gross, J.M. Genetic and epigenetic control of retinal development in zebrafish. Curr. Opin. Neurobiol. 2019, 59, 120–127. [Google Scholar] [CrossRef]
- Tarboush, R.; Chapman, G.B.; Connaughton, V.P. Ultrastructure of the distal retina of the adult zebrafish, Danio rerio. Tissue Cell 2012, 44, 264–279. [Google Scholar] [CrossRef]
- D’Angelo, L.; Avallone, L.; Cellerino, A.; de Girolamo, P.; Paolucci, M.; Varricchio, E.; Lucini, C. Neurotrophin-4 in the brain of adult Nothobranchius furzeri. Ann. Anat.-Anat. Anz. 2016, 207, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Braunbeck, T.; Lammer, E. Fish embryo toxicity assays. Ger. Fed. Environ. Agency 2006, 298, 7–119. [Google Scholar]
- Kroll, W. Morphological and behavioral embryology and spontaneous diapause in the African killifish, Aphyosemion gardneri. Environ. Biol. Fishes 1984, 11, 21–28. [Google Scholar] [CrossRef]
- Tozzini, E.T.; Baumgart, M.; Battistoni, G.; Cellerino, A. Adult neurogenesis in the short-lived teleost Nothobranchius furzeri: Localization of neurogenic niches, molecular characterization and effects of aging. Aging Cell 2012, 11, 241–251. [Google Scholar] [CrossRef]
- Vanhunsel, S.; Beckers, A.; Moons, L. Designing neuroreparative strategies using aged regenerating animal models. Ageing Res. Rev. 2020, 62, 101086. [Google Scholar] [CrossRef]
- Johns, P.R. Growth of the adult goldfish eye. III. Source of the new retinal cells. J. Comp. Neurol. 1977, 176, 343–357. [Google Scholar] [CrossRef]
- Raymond, P.A.; Barthel, L.K.; Bernardos, R.L.; Perkowski, J.J. Molecular characterization of retinal stem cells and their niches in adult zebrafish. BMC Dev. Biol. 2006, 6, 36. [Google Scholar] [CrossRef]
- Hrbek, T.; Larson, A. The Evolution of Diapause in the Killifish Family Rivulidae (Atherinomorpha, Cyprinodontiformes): A Molecular Phylogenetic and Biogeographic Perspective. Evolution 1999, 53, 1200–1216. [Google Scholar] [CrossRef]
- Vanhunsel, S.; Bergmans, S.; Beckers, A.; Etienne, I.; Van Houcke, J.; Seuntjens, E.; Arckens, L.; De Groef, L.; Moons, L. The killifish visual system as an in vivo model to study brain aging and rejuvenation. npj Aging Mech. Dis. 2021, 7, 22. [Google Scholar] [CrossRef]
- Cellerino, A.; Valenzano, D.R.; Reichard, M. From the bush to the bench: The annual Nothobranchius fishes as a new model system in biology. Biol. Rev. 2016, 91, 511–533. [Google Scholar] [CrossRef]
- Lucas-Sánchez, A.; Almaida-Pagán, P.F.; Mendiola, P.; de Costa, J. Nothobranchius as a model for aging studies. A review. Aging Dis 2013, 5, 281–291. [Google Scholar] [CrossRef]
- Valenzano, D.R.; Sharp, S.; Brunet, A. Transposon-mediated transgenesis in the short-lived African killifish Nothobranchius furzeri, a vertebrate model for aging. G3 Genes Genomes Genet. 2011, 1, 531–538. [Google Scholar] [CrossRef]
- Hartmann, N.; Englert, C. A microinjection protocol for the generation of transgenic killifish (Species: Nothobranchius furzeri). Dev. Dyn. 2012, 241, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Tozzini, E.T.; Cellerino, A. Nothobranchius annual killifishes. EvoDevo 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, X.; Feng, W.; Li, G.; Su, F.; Zhang, S. Differential expression of aging biomarkers at different life stages of the annual fish Nothobranchius guentheri. Biogerontology 2012, 13, 501–510. [Google Scholar] [CrossRef]
- Wang, X.; Shang, X.; Luan, J.; Zhang, S. Identification, expression and function of apolipoprotein E in annual fish Nothobranchius guentheri: Implication for an aging marker. Biogerontology 2014, 15, 233–243. [Google Scholar] [CrossRef]
- Mhalhel, K.; Briglia, M.; Aragona, M.; Porcino, C.; Abbate, F.; Guerrera, M.C.; Laurà, R.; Krichen, Y.; Guerbej, H.; Germanà, A.; et al. Nothobranchius as a model for anorexia of aging research: An evolutionary, anatomical, histological, immunohistochemical, and molecular study. Ann. Anat.-Anat. Anz. 2023, 250, 152116. [Google Scholar] [CrossRef]
- Jackson, G.R.; De Leon Ortega, J.; Girkin, C.; Rosenstiel, C.E.; Owsley, C. Aging-related changes in the multifocal electroretinogram. J. Opt. Soc. Am. A 2002, 19, 185–189. [Google Scholar] [CrossRef]
- Owsley, C.; Jackson, G.R.; Cideciyan, A.V.; Huang, Y.; Fine, S.L.; Ho, A.C.; Maguire, M.G.; Lolley, V.; Jacobson, S.G. Psychophysical Evidence for Rod Vulnerability in Age-Related Macular Degeneration. Investig. Ophthalmol. Vis. Sci. 2000, 41, 267–273. [Google Scholar]
- Owsley, C.; Jackson, G.R.; White, M.; Feist, R.; Edwards, D. Delays in rod-mediated dark adaptation in early age-related maculopathy. Ophthalmology 2001, 108, 1196–1202. [Google Scholar] [CrossRef]
- Curcio, C.A.; Drucker, D.N. Retinal ganglion cells in Alzheimer’s disease and aging. Ann. Neurol. 1993, 33, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Hollyfield, J.G. Aging of the human retina. Differential loss of neurons and retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 1992, 33, 1–17. [Google Scholar]
- Berkowitz, B.A.; Grady, E.M.; Roberts, R. Confirming a prediction of the calcium hypothesis of photoreceptor aging in mice. Neurobiol. Aging 2014, 35, 1883–1891. [Google Scholar] [CrossRef] [PubMed]
- Bonnel, S.; Mohand-Said, S.; Sahel, J.-A. The aging of the retina. Exp. Gerontol. 2003, 38, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Cunea, A.; Powner, M.B.; Jeffery, G. Death by color: Differential cone loss in the aging mouse retina. Neurobiol. Aging 2014, 35, 2584–2591. [Google Scholar] [CrossRef] [PubMed]
- Fabre, M.; Mateo, L.; Lamaa, D.; Baillif, S.; Pagès, G.; Demange, L.; Ronco, C.; Benhida, R. Recent Advances in Age-Related Macular Degeneration Therapies. Molecules 2022, 27, 5089. [Google Scholar] [CrossRef] [PubMed]
- Goyal, M.; Bordt, A.S.; Neitz, J.; Marshak, D.W. Trogocytosis of neurons and glial cells by microglia in a healthy adult macaque retina. Sci. Rep. 2023, 13, 633. [Google Scholar] [CrossRef]
- Nadal-Nicolás, F.M.; Vidal-Sanz, M.; Agudo-Barriuso, M. The aging rat retina: From function to anatomy. Neurobiol. Aging 2018, 61, 146–168. [Google Scholar] [CrossRef] [PubMed]
- Nag, T.C. Müller cell vulnerability in aging human retina: Implications on photoreceptor cell survival. Exp. Eye Res. 2023, 235, 109645. [Google Scholar] [CrossRef]
- Sachdeva, M.M. Retinal Neurodegeneration in Diabetes: An Emerging Concept in Diabetic Retinopathy. Curr. Diabetes Rep. 2021, 21, 65. [Google Scholar] [CrossRef]
- Mirzaei, N.; Shi, H.; Oviatt, M.; Doustar, J.; Rentsendorj, A.; Fuchs, D.-T.; Sheyn, J.; Black, K.L.; Koronyo, Y.; Koronyo-Hamaoui, M. Alzheimer’s retinopathy: Seeing disease in the eyes. Front. Neurosci. 2020, 14, 921. [Google Scholar] [CrossRef]
- Vandenabeele, M.; Veys, L.; Lemmens, S.; Hadoux, X.; Gelders, G.; Masin, L.; Serneels, L.; Theunis, J.; Saito, T.; Saido, T.C.; et al. The AppNL-G-F mouse retina is a site for preclinical Alzheimer’s disease diagnosis and research. Acta Neuropathol. Commun. 2021, 9, 6. [Google Scholar] [CrossRef]
- Veys, L.; Vandenabeele, M.; Ortuño-Lizarán, I.; Baekelandt, V.; Cuenca, N.; Moons, L.; De Groef, L. Retinal α-synuclein deposits in Parkinson’s disease patients and animal models. Acta Neuropathol. 2019, 137, 379–395. [Google Scholar] [CrossRef]
- Chader, G.J.; Taylor, A. Preface: The Aging Eye: Normal Changes, Age-Related Diseases, and Sight-Saving Approaches. Investig. Ophthalmol. Vis. Sci. 2013, 54, ORSF1–ORSF4. [Google Scholar] [CrossRef]
- Cao, W.; Li, T. COVID-19: Towards understanding of pathogenesis. Cell Res. 2020, 30, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Nations, U. World Population Ageing 2019 Division; United Nations Department of Economic and Social Affairs: New York, NY, USA, 2020; p. 64. [Google Scholar]
- Salvi, S.M.; Akhtar, S.; Currie, Z. Ageing changes in the eye. Postgrad. Med. J. 2006, 82, 581. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Genade, T.; Benedetti, M.; Terzibasi, E.; Roncaglia, P.; Valenzano, D.R.; Cattaneo, A.; Cellerino, A. Annual fishes of the genus Nothobranchius as a model system for aging research. Aging Cell 2005, 4, 223–233. [Google Scholar] [CrossRef]
- Terzibasi, E.; Valenzano, D.R.; Cellerino, A. The short-lived fish Nothobranchius furzeri as a new model system for aging studies. Exp. Gerontol. 2007, 42, 81–89. [Google Scholar] [CrossRef]
- Valenzano, D.R.; Cellerino, A. Resveratrol and the Pharmacology of Aging: A New Vertebrate Model to Validate an Old Molecule. Cell Cycle 2006, 5, 1027–1032. [Google Scholar] [CrossRef]
- Baumgart, M.; Groth, M.; Priebe, S.; Savino, A.; Testa, G.; Dix, A.; Ripa, R.; Spallotta, F.; Gaetano, C.; Ori, M.; et al. RNA-seq of the aging brain in the short-lived fish N. furzeri—Conserved pathways and novel genes associated with neurogenesis. Aging Cell 2014, 13, 965–974. [Google Scholar] [CrossRef]
- Hartmann, N.; Reichwald, K.; Wittig, I.; Dröse, S.; Schmeisser, S.; Lück, C.; Hahn, C.; Graf, M.; Gausmann, U.; Terzibasi, E.; et al. Mitochondrial DNA copy number and function decrease with age in the short-lived fish Nothobranchius furzeri. Aging Cell 2011, 10, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Bollaerts, I.; Veys, L.; Geeraerts, E.; Andries, L.; De Groef, L.; Buyens, T.; Salinas-Navarro, M.; Moons, L.; Van Hove, I. Complementary research models and methods to study axonal regeneration in the vertebrate retinofugal system. Brain Struct. Funct. 2018, 223, 545–567. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Normando, E.M.; Shah, P.A.; De Groef, L.; Cordeiro, M.F. Oculo-visual abnormalities in Parkinson’s disease: Possible value as biomarkers. Mov. Disord. 2018, 33, 1390–1406. [Google Scholar] [CrossRef] [PubMed]
- Telegina, D.V.; Kolosova, N.G.; Kozhevnikova, O.S. Immunohistochemical localization of NGF, BDNF, and their receptors in a normal and AMD-like rat retina. BMC Med. Genom. 2019, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Pezet, S.; McMahon, S.B. Neurotrophins: Mediators and Modulators of Pain. Annu. Rev. Neurosci. 2006, 29, 507–538. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, L.; Li, Z.; Zhang, X.; Wu, Y.; Yang, H.; Min, B.; Zhang, X.; Ma, D.; Lu, Y. Thinner changes of the retinal nerve fiber layer in patients with mild cognitive impairment and Alzheimer’s disease. BMC Neurol. 2015, 15, 14. [Google Scholar] [CrossRef] [PubMed]
- Michalski, D.; Preißler, H.; Hofmann, S.; Kacza, J.; Härtig, W. Decline of microtubule-associated protein tau after experimental stroke in differently aged wild-type and 3xTg mice with Alzheimer-like alterations. Neuroscience 2016, 330, 1–11. [Google Scholar] [CrossRef]
- Parnell, M.; Guo, L.; Abdi, M.; Cordeiro, M.F. Ocular Manifestations of Alzheimer’s Disease in Animal Models. Int. J. Alzheimer’s Dis. 2012, 2012, 786494. [Google Scholar] [CrossRef]
- Cellerino, A.; Kohler, K. Brain-derived neurotrophic factor/neurotrophin-4 receptor TrkB is localized on ganglion cells and dopaminergic amacrine cells in the vertebrate retina. J. Comp. Neurol. 1997, 386, 149–160. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2017, 114, 1752–1761. [Google Scholar] [CrossRef]
- Lein, E.S.; Hohn, A.; Shatz, C.J. Dynamic regulation of BDNF and NT-3 expression during visual system development. J. Comp. Neurol. 2000, 420, 1–18. [Google Scholar] [CrossRef]
- Hallböök, F. Evolution of the vertebrate neurotrophin and Trk receptor gene families. Curr. Opin. Neurobiol. 1999, 9, 616–621. [Google Scholar] [CrossRef]
- Heinrich, G.; Lum, T. Fish neurotrophins and Trk receptors. Int. J. Dev. Neurosci. 2000, 18, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.C.; Marazzi, G.; Sandell, J.H.; Heinrich, G. Five Trk Receptors in the Zebrafish. Dev. Biol. 1995, 169, 745–758. [Google Scholar] [CrossRef] [PubMed]
- Caminos, E.; Becker, E.; Martín-Zanca, D.; Vecino, E. Neurotrophins and their receptors in the tench retina during optic nerve regeneration. J. Comp. Neurol. 1999, 404, 321–331. [Google Scholar] [CrossRef]
- Abed, E.; Corbo, G.; Falsini, B. Neurotrophin Family Members as Neuroprotectants in Retinal Degenerations. BioDrugs 2015, 29, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Inanc Tekin, M.; Sekeroglu, M.A.; Demirtas, C.; Tekin, K.; Doguizi, S.; Bayraktar, S.; Yilmazbas, P. Brain-Derived Neurotrophic Factor in Patients With Age-Related Macular Degeneration and Its Correlation With Retinal Layer Thicknesses. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2833–2840. [Google Scholar] [CrossRef]
- Taslipinar Uzel, A.G.; Ugurlu, N.; Toklu, Y.; ÇIçek, M.; Boral, B.; Sener, B.; ÇaGil, N. Relationship between stages of diabetic retinopathy and levels of Brain-Derived Neurotrophic Factor in aqueous humor and serum. Retina 2020, 40, 121–125. [Google Scholar] [CrossRef]
- Kimura, Y.; Sumiyoshi, M. Olive leaf extract and its main component oleuropein prevent chronic ultraviolet B radiation-induced skin damage and carcinogenesis in hairless mice. J. Nutr. 2009, 139, 2079–2086. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.J.; Harman, C.D.; Viswanathan, S. Effects of optic nerve injury, glaucoma, and neuroprotection on the survival, structure, and function of ganglion cells in the mammalian retina. J. Physiol. 2008, 586, 4393–4400. [Google Scholar] [CrossRef]
- Bessero, A.-C.; Clarke, P.G.H. Neuroprotection for optic nerve disorders. Curr. Opin. Neurol. 2010, 23, 10–15. [Google Scholar] [CrossRef]
- Zhang, X.; Bao, S.; Hambly, B.D.; Gillies, M.C. Vascular endothelial growth factor-A: A multifunctional molecular player in diabetic retinopathy. Int. J. Biochem. Cell Biol. 2009, 41, 2368–2371. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Liu, C.; Yang, M.; Li, H.; Zhang, F.; Zhang, W.; Zhang, J. Neurotrophic factor and Trk signaling mechanisms underlying the promotion of motor recovery after acute spinal cord injury in rats. Exp. Ther. Med. 2017, 14, 652–656. [Google Scholar] [CrossRef]
- Gatta, C.; Castaldo, L.; Cellerino, A.; de Girolamo, P.; Lucini, C.; D’Angelo, L. Brain derived neurotrophic factor in the retina of the teleost N. furzeri. Ann. Anat.-Anat. Anz. 2014, 196, 192–196. [Google Scholar] [CrossRef]
- D’Angelo, L.; Castaldo, L.; Cellerino, A.; de Girolamo, P.; Lucini, C. Nerve growth factor in the adult brain of a teleostean model for aging research: Nothobranchius furzeri. Ann. Anat.-Anat. Anz. 2014, 196, 183–191. [Google Scholar] [CrossRef]
- de Girolamo, P.; Leggieri, A.; Palladino, A.; Lucini, C.; Attanasio, C.; D’Angelo, L. Cholinergic System and NGF Receptors: Insights from the Brain of the Short-Lived Fish Nothobranchius furzeri. Brain Sci. 2020, 10, 394. [Google Scholar] [CrossRef]
- Garcia, T.B.; Hollborn, M.; Bringmann, A. Expression and signaling of NGF in the healthy and injured retina. Cytokine Growth Factor Rev. 2017, 34, 43–57. [Google Scholar] [CrossRef]
- Theiss, C.; Güntürkün, O. Distribution of BDNF, NT-3, trkB and trkC in the developing retino-tectal system of the pigeon (Columba livia). Anat. Embryol. 2001, 204, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.; Monzón-Mayor, M.; Romero-Alemán, M.M.; Yanes, C. Distribution of neurotrophin-3 during the ontogeny and regeneration of the lizard (Gallotia galloti) visual system. Dev. Neurobiol. 2008, 68, 31–44. [Google Scholar] [CrossRef]
- Duprey-Díaz, M.V.; Blagburn, J.M.; Blanco, R.E. Neurotrophin-3 and TrkC in the frog visual system: Changes after axotomy. Brain Res. 2003, 982, 54–63. [Google Scholar] [CrossRef]
- Llamosas, M.M.; Cernuda-Cernuda, R.; Huerta, J.J.; Vega, J.A.; García-Fernández, J.M. Neurotrophin receptors expression in the developing mouse retina: An immunohistochemical study. Anat. Embryol. 1997, 195, 337–344. [Google Scholar] [CrossRef]
- Grishanin, R.N.; Yang, H.; Liu, X.; Donohue-Rolfe, K.; Nune, G.C.; Zang, K.; Xu, B.; Duncan, J.L.; LaVail, M.M.; Copenhagen, D.R.; et al. Retinal TrkB receptors regulate neural development in the inner, but not outer, retina. Mol. Cell. Neurosci. 2008, 38, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Nag, T.C.; Wadhwa, S. Developmental expression of calretinin immunoreactivity in the human retina and a comparison with two other EF-hand calcium-binding proteins. Neuroscience 1999, 91, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Germanà, A.; Sánchez-Ramos, C.; Guerrera, M.C.; Calavia, M.; Navarro, M.; Zichichi, R.; García-Suárez, O.; Pérez-Piñera, P.; Vega, J.A. Expression and cell localization of brain-derived neurotrophic factor and TrkB during zebrafish retinal development. J. Anat. 2010, 217, 214–222. [Google Scholar] [CrossRef]
- Peinado-Ramon, P.; Salvador, M.; Villegas-Perez, M.P.; Vidal-Sanz, M. Effects of axotomy and intraocular administration of NT-4, NT-3, and brain-derived neurotrophic factor on the survival of adult rat retinal ganglion cells. A quantitative in vivo study. Investig. Ophthalmol. Vis. Sci. 1996, 37, 489–500. [Google Scholar]
- Bennett, J.L.; Zeiler, S.R.; Jones, K.R. Patterned Expression of BDNF and NT-3 in the Retina and Anterior Segment of the Developing Mammalian Eye. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2996–3005. [Google Scholar]
- Das, I.; Hempstead, B.L.; Macleish, P.R.; Sparrow, J.R. Immunohistochemical analysis of the neurotrophins BDNF and NT-3 and their receptors trk B, trk C, and p75 in the developing chick retina. Vis. Neurosci. 1997, 14, 835–842. [Google Scholar] [CrossRef]
- Colucci-D’Amato, L.; Speranza, L.; Volpicelli, F. Neurotrophic Factor BDNF, Physiological Functions and Therapeutic Potential in Depression, Neurodegeneration and Brain Cancer. Int. J. Mol. Sci. 2020, 21, 7777. [Google Scholar] [CrossRef]
- Kishi, S.; Slack, B.E.; Uchiyama, J.; Zhdanova, I.V. Zebrafish as a Genetic Model in Biological and Behavioral Gerontology: Where Development Meets Aging in Vertebrates—A Mini-Review. Gerontology 2009, 55, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Ramos, C.; Bonnin-Arias, C.; Guerrera, M.C.; Calavia, M.; Chamorro, E.; Montalbano, G.; López-Velasco, S.; López-Muñiz, A.; Germanà, A.; Vega, J.A. Light regulates the expression of the BDNF/TrkB system in the adult zebrafish retina. Microsc. Res. Tech. 2013, 76, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Colucci, P.; Giannaccini, M.; Baggiani, M.; Kennedy, B.N.; Dente, L.; Raffa, V.; Gabellini, C. Neuroprotective Nanoparticles Targeting the Retina: A Polymeric Platform for Ocular Drug Delivery Applications. Pharmaceutics 2023, 15, 1096. [Google Scholar] [CrossRef]
- Mattern, L.; Otten, K.; Miskey, C.; Fuest, M.; Izsvák, Z.; Ivics, Z.; Walter, P.; Thumann, G.; Johnen, S. Molecular and Functional Characterization of BDNF-Overexpressing Human Retinal Pigment Epithelial Cells Established by Sleeping Beauty Transposon-Mediated Gene Transfer. Int. J. Mol. Sci. 2022, 23, 12982. [Google Scholar] [CrossRef] [PubMed]
- Rocco, M.L.; Balzamino, B.O.; Petrocchi Passeri, P.; Micera, A.; Aloe, L. Effect of purified murine NGF on isolated photoreceptors of a rodent developing retinitis pigmentosa. PLoS ONE 2015, 10, e0124810. [Google Scholar] [CrossRef]
- Harper, M.M.; Boese, E.A.; Kardon, R.H.; Ledolter, J.; Kuehn, M.H. High Correlation between Glaucoma Treatment with Topical Prostaglandin Analogs and BDNF Immunoreactivity in Human Retina. Curr. Eye Res. 2021, 46, 739–745. [Google Scholar] [CrossRef]
- Balzamino, B.O.; Biamonte, F.; Esposito, G.; Marino, R.; Fanelli, F.; Keller, F.; Micera, A. Characterization of NGF, trkANGFR, and p75NTR in Retina of Mice Lacking Reelin Glycoprotein. Int. J. Cell Biol. 2014, 2014, 725928. [Google Scholar] [CrossRef]
- Martin, K.L.; Lighton, J.R. Aerial CO2 and O2 exchange during terrestrial activity in an amphibious fish, Alticus kirki (Blenniidae). Copeia 1989, 1989, 723–727. [Google Scholar] [CrossRef]
- Cellerino, A.; Arango-González, B.A.; Kohler, K. Effects of brain-derived neurotrophic factor on the development of NADPH-diaphorase/nitric oxide synthase-positive amacrine cells in the rodent retina. Eur. J. Neurosci. 1999, 11, 2824–2834. [Google Scholar] [CrossRef]
- Yu, J.Y.; Jeong, D.E.; Joo, J.Y.; Kim, S.T. Brain-derived neurotrophic factor levels and macular ganglion cell-inner plexiform layer thickness in macular telangiectasia type 2. Int. Ophthalmol. 2023, 43, 1927–1933. [Google Scholar] [CrossRef] [PubMed]
- Zanellato, A.; Comelli, M.C.; Dal Toso, R.; Carmignoto, G. Developing Rat Retinal Ganglion Cells Express the Functional NGF Receptor p140trkA. Dev. Biol. 1993, 159, 105–113. [Google Scholar] [CrossRef]
- Park, M.; Kim, H.-M.; Shin, H.-A.; Lee, S.-H.; Hwang, D.-Y.; Lew, H. Human Pluripotent Stem Cell-Derived Neural Progenitor Cells Promote Retinal Ganglion Cell Survival and Axon Recovery in an Optic Nerve Compression Animal Model. Int. J. Mol. Sci. 2021, 22, 12529. [Google Scholar] [CrossRef]
- Roberti, G.; Mantelli, F.; Macchi, I.; Massaro-Giordano, M.; Centofanti, M. Nerve Growth Factor Modulation of Retinal Ganglion Cell Physiology. J. Cell. Physiol. 2014, 229, 1130–1133. [Google Scholar] [CrossRef]
- Lambert, W.S.; Clark, A.F.; Wordinger, R.J. Neurotrophin and Trk expression by cells of the human lamina cribrosa following oxygen-glucose deprivation. BMC Neurosci. 2004, 5, 51. [Google Scholar] [CrossRef]
- D’angelo, L. Brain Atlas of an Emerging Teleostean Model: Nothobranchius furzeri. Anat. Rec. 2013, 296, 681–691. [Google Scholar] [CrossRef]
- D’Angelo, L.; De Girolamo, P.; Lucini, C.; Terzibasi, E.T.; Baumgart, M.; Castaldo, L.; Cellerino, A. Brain-derived neurotrophic factor: mRNA expression and protein distribution in the brain of the teleost Nothobranchius furzeri. J. Comp. Neurol. 2014, 522, 1004–1030. [Google Scholar] [CrossRef]
- Daly, C.; Shine, L.; Heffernan, T.; Deeti, S.; Reynolds, A.L.; O’Connor, J.J.; Dillon, E.T.; Duffy, D.J.; Kolch, W.; Cagney, G.; et al. A Brain-Derived Neurotrophic Factor Mimetic Is Sufficient to Restore Cone Photoreceptor Visual Function in an Inherited Blindness Model. Sci. Rep. 2017, 7, 11320. [Google Scholar] [CrossRef]
- Aragona, M.; Porcino, C.; Guerrera, M.C.; Montalbano, G.; Levanti, M.; Abbate, F.; Laurà, R.; Germanà, A. Localization of Neurotrophin Specific Trk Receptors in Mechanosensory Systems of Killifish (Nothobranchius guentheri). Int. J. Mol. Sci. 2021, 22, 10411. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; Ciriaco, E.; Germanà, A. Morphology of the tongue dorsal surface of gilthead seabream (Sparus aurata). Microsc. Res. Tech. 2012, 75, 1666–1671. [Google Scholar] [CrossRef] [PubMed]
- Lauriano, E.; Guerrera, M.; Laurà, R.; Capillo, G.; Pergolizzi, S.; Aragona, M.; Abbate, F.; Germanà, A. Effect of light on the calretinin and calbindin expression in skin club cells of adult zebrafish. Histochem. Cell Biol. 2020, 154, 495–505. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Capillo, G.; Icardo, J.M.; Fernandes, J.M.O.; Kiron, V.; Kuciel, M.; Zuwala, K.; Guerrera, M.C.; Aragona, M.; Germana’, A.; et al. Neuroepithelial cells (NECs) and mucous cells express a variety of neurotransmitters and neurotransmitter receptors in the gill and respiratory air-sac of the catfish Heteropneustes fossilis (Siluriformes, Heteropneustidae): A possible role in local immune defence. Zoology 2021, 148, 125958. [Google Scholar] [CrossRef]
- Capillo, G.; Zaccone, G.; Cupello, C.; Fernandes, J.M.O.; Viswanath, K.; Kuciel, M.; Zuwala, K.; Guerrera, M.C.; Aragona, M.; Icardo, J.M.; et al. Expression of acetylcholine, its contribution to regulation of immune function and O2 sensing and phylogenetic interpretations of the African butterfly fish Pantodon buchholzi (Osteoglossiformes, Pantodontidae). Fish Shellfish. Immunol. 2021, 111, 189–200. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies Investigated | Specific Antibodies for Retinal Cells | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BDNF | TrkB | NGF | TrkA | NT-3 | TrkC | Opsin | Chat | Parv | S100p | |
| Mean ± ∆σ in RPE | 28.4 ± 2.15 *** | 27.8 ± 1.95 * | ꟷ | 28.6 ± 2.45 *** | 25.9 ± 2.46 *** | 25.9 ± 2.85 *** | 27.2 ± 1.24 ** | 26.3 ± 1. 83 *** | 26.5 ± 2.57 *** | 28.8 ± 5.11 *** |
| Mean ± ∆σ in PRL (outer segment) | 27.7 ± 2.49 * | ꟷ | 27.7 ± 2.49 * | ꟷ | ꟷ | 25.3 ± 3.13 *** | 26.9 ± 3. 82 * | 26.8 ± 2. 40 ** | 25.8 ± 2.35 *** | 26 ± 3.66 *** |
| Mean ± ∆σ in PRL (inner segment) | 26.4 ± 2.9 ** | 26.3 ± 2.86 ** | 27.9 ± 2.80 * | 28.4 ± 2.74 *** | 25.2 ± 3.6 ** | ꟷ | 27.1 ± 4. 19 ** | ꟷ | 23.7 ± 3.95 *** | 27.8 ± 5.97 *** |
| Mean ± ∆σ in OPL | 26 ± 3.66 *** | 25.3 ± 3.13 *** | 25.8 ± 2.35 *** | 26.5 ± 2.57 *** | 26.2 ± 2.08 *** | 25.4 ± 2.15 ** | ꟷ | ꟷ | ꟷ | ꟷ |
| Mean ± ∆σ of ACs | 27.8 ± 2.31 * | 26.9 ± 2.58 *** | 27.7 ± 2.49 * | 27.9 ± 2.79 * | ꟷ | 26.8 ± 2. 52 *** | ꟷ | 27.9 ± 3. 04 * | 26.5 ± 5.48 *** | 26.7 ± 4.64 *** |
| Mean ± ∆σ of BCs | ꟷ | 26.9 ± 2.36 *** | 26.5 ± 2.57 *** | 27.8 ± 3.65 * | 26.5 ± 2.57 *** | 27.7 ± 2.14 *** | ꟷ | ꟷ | 27.7 ± 5.36 *** | 25.8 ± 4.6 *** |
| Mean ± ∆σ of HCs | ꟷ | 27.7 ± 2.49 * | ꟷ | ꟷ | ꟷ | 25.8 ± 2.67 *** | ꟷ | ꟷ | ꟷ | 26.3 ± 4 ** |
| Mean ± ∆σ in IPL | ꟷ | ꟷ | ꟷ | ꟷ | ꟷ | ꟷ | ꟷ | ꟷ | 27.4 ± 4.8 ** | 26.5 ± 5.12 *** |
| Mean ± ∆σ in GCs | 27.2 ± 1.24 ** | 26.9 ± 1. 42 ** | 26 ± 2.89 * | 26.7 ± 3. 40 *** | 27.9 ± 2.38 * | 27.2 ± 2.62 ** | ꟷ | ꟷ | 26.7 ± 4.42 *** | 27.4 ± 4.84 *** |
| Species | N. guentheri * | Zebrafish | Mouse | Human | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Antibodies | BDNF | NGF | NT-3 | Trks | BDNF | NGF | NT-3 | Trks | BDNF | NGF | NT-3 | Trks | BDNF | NGF | NT-3 | Trks | |||||||||
| A | B | C | A | B | C | A | B | C | A | B | C | ||||||||||||||
| Cell layers | RPE | + | n/a | + | + | + | + | + | + | − | + | + | + | + | + | n/a | + | + | − | + | n/a | n/a | + | + | − |
| R. | [10,81,105,107,108,109,110,111,112] | [17,96,101] | [97,113] | ||||||||||||||||||||||
| PRL | + | + | + | + | + | + | + | + | − | + | + | + | + | + | + | + | + | + | + | n/a | n/a | + | + | + | |
| R. | [10,81,110,111,112] | [95,101,108,114] | [97,115] | ||||||||||||||||||||||
| OPL | − | − | − | − | − | − | + | + | − | + | + | + | + | n/a | + | + | − | + | + | n/a | n/a | + | + | + | |
| R. | [10,81,110,111,112] | [95,108] | [97,115] | ||||||||||||||||||||||
| INL | + | + | + | − | − | − | + | + | − | + | + | + | + | + | + | + | + | − | + | n/a | n/a | + | + | + | |
| R. | [10,81,110,111,112] | [95,108,116] | [97,115] | ||||||||||||||||||||||
| HCs | − | − | − | − | − | − | + | + | − | + | + | + | + | + | n/a | + | + | + | n/a | n/a | n/a | + | + | + | |
| R. | [10,81,110,111,112] | [95,108] | [97] | ||||||||||||||||||||||
| BCs | − | + | + | + | + | − | + | + | − | + | + | + | + | + | n/a | − | − | − | n/a | n/a | n/a | n/a | n/a | n/a | |
| R. | [10,81,110,111,112] | [95,101,108,114,116] | |||||||||||||||||||||||
| ACs | + | + | + | + | + | + | + | + | − | + | + | + | + | n/a | + | + | − | − | + | n/a | n/a | + | + | + | |
| R. | [10,110,111,112,117] | [95,108] | [97,118] | ||||||||||||||||||||||
| IPL | − | − | − | − | − | − | + | + | − | + | + | + | + | n/a | n/a | + | + | + | + | n/a | n/a | + | + | + | |
| R. | [10,81,102,110,111,112] | [95,108] | [97,119] | ||||||||||||||||||||||
| GCL | + | + | + | + | + | + | + | + | − | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| R. | [10,81,102,110,111,112] | [17,95,96,107,120] | [97,121,122,123] | ||||||||||||||||||||||
| Primary Antibodies | Supplier | Catalogue Number | Source | Dilution | Antibody ID |
|---|---|---|---|---|---|
| BDNF | Sigma-Aldrich, Inc., St. Louis, MO, USA | AB1534SP | rabbit | 1:100 | AB_90748 |
| NGF | Sigma-Aldrich, Inc., St. Louis, MO, USA | AB1526 | rabbit | 1:100 | AB_90733 |
| NT-3 (A4) | Santa Cruz Biotechnology, Inc., Dallas, TX, USA | sc-518099 | mouse | 1:100 | |
| TrkA (Y32Ex) | Santa Cruz Biotechnology, Inc., Dallas, TX, USA | sc-80398 | mouse | 1:100 | AB_1130726 |
| TrkB (F-1) | Santa Cruz Biotechnology, Inc., Dallas, TX, USA | sc-377218 | mouse | 1:100 | AB_2801499 |
| TrkC (798) | Santa Cruz Biotechnology, Inc., Dallas, TX, USA | sc-117 | rabbit | 1:100 | AB_632560 |
| Anti-Opsin Clone RET-P1 | Sigma-Aldrich, Inc., St. Louis, MO, USA | O4886 | mouse | 1:100 | AB_260838 |
| Anti-Chat | Sigma-Aldrich, Inc., St. Louis, MO, USA | AMAB91130 | mouse | 1:100 | AB_2665812 |
| Parvalbumin clone PA235 | Sigma-Aldrich, Inc., St. Louis, MO, USA | P-3171 | mouse | 1:1000 | AB_2313693 |
| S100p | Dako Agilent, Santa Clara, CA, USA | Z0311 | rabbit | 1:100 | AB_10013383 |
| Secondary Antibody | Supplier | Catalogue Number | Source | Dilution | Antibody ID |
| Anti-rabbit IgG (H + L) Alexa Fluor 594 | Molecular Probes, Invitrogen, Waltham, MA, USA | A32754 | Donkey | 1:300 | AB_2762827 |
| Anti-rabbit IgG (H + L) Alexa Fluor 488 | Molecular Probes, Invitrogen, Waltham, MA, USA | A-11008 | goat | 1:300 | AB_143165 |
| Anti-mouse IgG (H + L) Alexa Fluor 488 | Molecular Probes, Invitrogen, Waltham, MA, USA | A-11001 | goat | 1:300 | AB_2534069 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porcino, C.; Mhalhel, K.; Briglia, M.; Cometa, M.; Guerrera, M.C.; Germanà, P.G.; Montalbano, G.; Levanti, M.; Laurà, R.; Abbate, F.; et al. Neurotrophins and Trk Neurotrophin Receptors in the Retina of Adult Killifish (Nothobranchius guentheri). Int. J. Mol. Sci. 2024, 25, 2732. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25052732
Porcino C, Mhalhel K, Briglia M, Cometa M, Guerrera MC, Germanà PG, Montalbano G, Levanti M, Laurà R, Abbate F, et al. Neurotrophins and Trk Neurotrophin Receptors in the Retina of Adult Killifish (Nothobranchius guentheri). International Journal of Molecular Sciences. 2024; 25(5):2732. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25052732
Chicago/Turabian StylePorcino, Caterina, Kamel Mhalhel, Marilena Briglia, Marzio Cometa, Maria Cristina Guerrera, Patrizia Germana Germanà, Giuseppe Montalbano, Maria Levanti, Rosaria Laurà, Francesco Abbate, and et al. 2024. "Neurotrophins and Trk Neurotrophin Receptors in the Retina of Adult Killifish (Nothobranchius guentheri)" International Journal of Molecular Sciences 25, no. 5: 2732. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25052732