Limnology and Aquatic Microbial Ecology of Byers Peninsula: A Main Freshwater Biodiversity Hotspot in Maritime Antarctica


Cavanilles Institute for Biodiversity and Evolutionary Biology, University of Valencia, E-46980 Paterna, Valencia, Spain
*
Author to whom correspondence should be addressed.
Diversity 2019, 11(10), 201; https://0-doi-org.brum.beds.ac.uk/10.3390/d11100201
Submission received: 1 June 2019
/
Revised: 24 September 2019
/
Accepted: 25 September 2019
/
Published: 21 October 2019
(This article belongs to the Special Issue Microbial Diversity in Extreme Environments: Implications for Ecological and Applicative Perspectives)
Abstract
:Here we present a comprehensive review of the diversity revealed by research in limnology and microbial ecology conducted in Byers Peninsula (Livingston Island, South Shetland Islands, Antarctica) during the last two decades. The site constitutes one of the largest ice-free areas within the Antarctic Peninsula region. Since it has a high level of environmental protection, it is less human-impacted compared to other sites within the South Shetland archipelago. The main investigations in Byers Peninsula focused on the physical and chemical limnology of the lakes, ponds, rivers, and wetlands, as well as on the structure of their planktonic and benthic microbial communities, and on the functional ecology of the microbial food webs. Lakes and ponds in Byers range along a productivity gradient that extends from the less productive lakes located upland to the eutrophic coastal lakes. Their planktonic assemblages include viruses, bacteria, a metabolically diverse community of protists (i.e., autotrophs, heterotrophs, and mixotrophs), and a few metazooplankton species. Most of the studies conducted in the site demonstrate the strong influence of the physical environment (i.e., temperature, availability of light, and water) and nutrient availability in structuring these microbial communities. However, top-down biotic processes may occur in summer, when predation by zooplankton can exert a strong influence on the abundance of protists, including flagellates and ciliated protozoa. As a consequence, bacterioplankton could be partly released from the grazing pressure exerted by these protists, and proliferates fueled by external nutrient subsidies from the lake’s catchment. As summer temperatures in this region are slightly above the melting point of water, biotic processes, such as those related to the productivity of lakes during ice-free periods, could become even more relevant as warming induced by climate change progresses. The limnological research carried out at the site proves that Byers Peninsula deserves special attention in the framework of the research in extreme environments. Together with nearby sites, such as Signy Island, Byers Peninsula comprises a featuring element of the Maritime Antarctic region that represents a benchmark area relative to the global distribution and diversity of aquatic microorganisms.
1. Introduction
Antarctica harbors one of the coldest and, in some places, driest climates on Earth, which justifies the dominance of microorganisms as the most diverse components of its biological communities [1]. From an ecological standpoint, the isolation of Antarctica and the simplicity of its biological communities provide a unique “laboratory” for the study of ecological processes [2]. In particular, in the Antarctic freshwater ecosystems, trophic pathways are mostly limited to the microbial loop, although the presence of metazoans suggests a greater trophic complexity [3].
Among the different types of Antarctic habitats, there are coastal locations where temperatures rise above the freezing point during summer and have undergone deglaciation during the last centuries or millennia [4]. They show periglacial landforms where water flows during several months, and aquatic organisms thrive. There is a larger relative amount of these ice-free areas in Maritime Antarctica compared to the Continental region due to its milder climate. In the last two decades, research conducted in both regions has significantly contributed to the knowledge of the composition and functioning of microbial communities in high latitude freshwater ecosystems. This is the case, for instance, of lake-rich areas, such as the McMurdo Dry Valleys [5,6,7], Vestfold Hills [8,9], Larsemann Hills [10,11], and Amery Oasis [12,13], all of them located in the continent, as well as in zones of the Maritime Antarctica, such as Signy Island [14] and King George Island [15,16]. However, at the beginning of the present century, there was still an important lack of limnological information about Byers Peninsula, which had been understudied despite being as valuable as the former from a limnological standpoint [17].
Byers Peninsula was designated as an Antarctic Specially Protected Area (ASPA number 126). This recognition was granted because of the extraordinary biological, geological, and archaeological values of the site, and aims to minimize environmental impacts. Byers Peninsula is only open to visits by small numbers of scientists at a time, with stringent environmental protection measures. The Limnopolar project started to study Byers Peninsula in 2001 with the aim to fill the knowledge gap by studying the ecological processes and the structure of the biological communities of freshwater ecosystems in this area, as well as their response to global warming [17]. The papers by Camacho [3], Toro et al. [18], and Fernández–Valiente et al. [19] were the opening reports of this research program. Thereafter, the Spanish Polar Research program published several studies dealing with the limnology and microbiology of the site, as well as the effect of climate change and other disciplines [17,20]. Our review highlights the studies on the limnology and microbial ecology of Byers Peninsula conducted during the last two decades, most of them within the context of the Limnopolar projects, to highlight the distinctive biological features of the site and to promote its international acceptance as a reference site [17]. Freshwater ecosystems from Byers Peninsula are useful models to understand major microbial processes occurring in polar regions, both functional and evolutionary processes, therefore rendering an important baseline for environmental studies.
2. Landscape and Weather
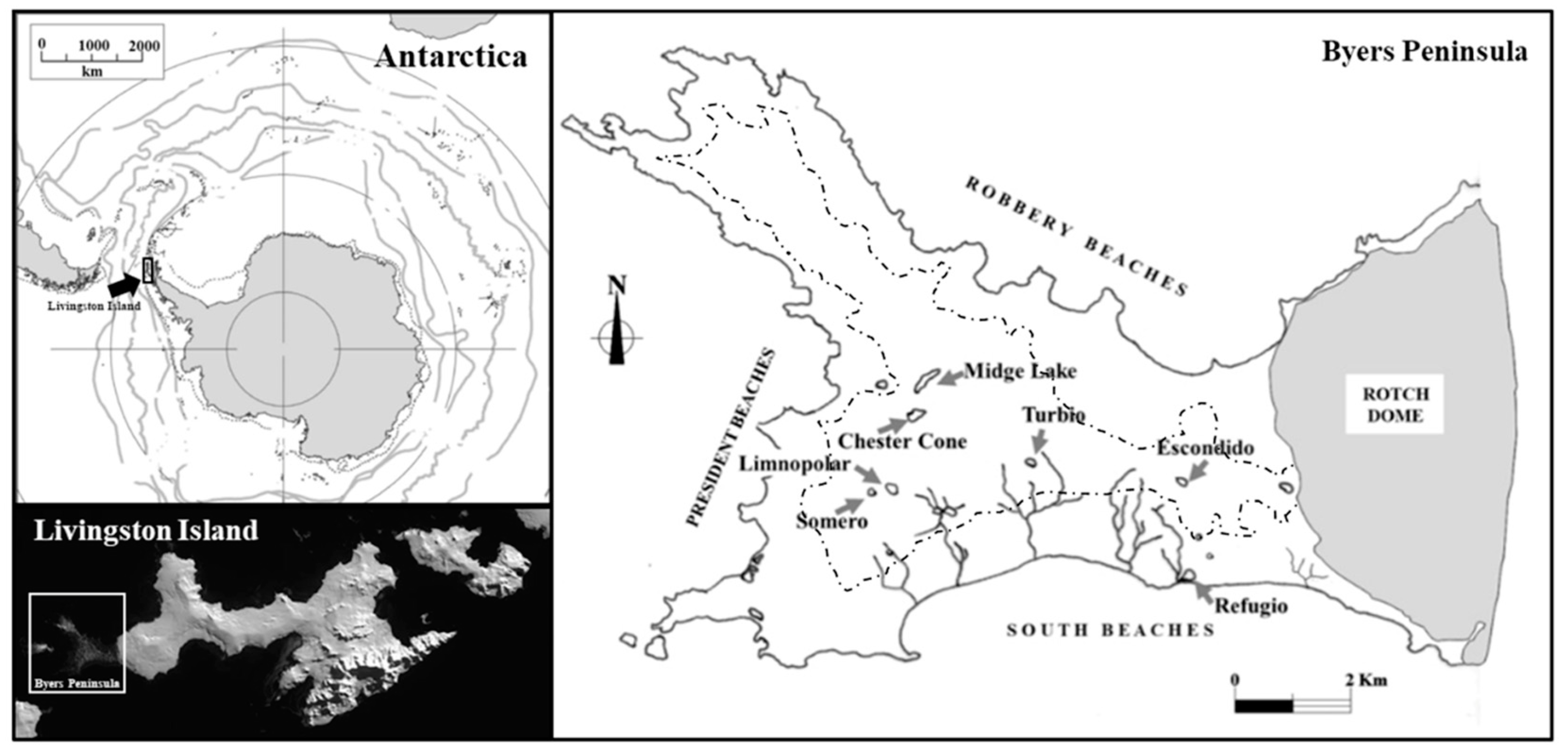
Byers Peninsula is one of the largest (~60 km2) ice-free areas in the region of the Antarctic Peninsula, located at the western side of Livingston Island (62°40′ S, 61° W), one of the main islands in the South Shetlands archipelago (Figure 1). Byers Peninsula is the site, within the maritime Antarctic region, where the rocks of the Mesozoic–Cenozoic magmatic arc complex are more exposed [21]. The drainage pattern is structured by the high rates of ablation occurring during summer. According to Navas et al. [22], there are two main geomorphological areas in Byers (Figure 1). One of them comprises the marine platforms and beaches bordering the coast, whereas the other comprises an elevated platform located between 85 and 100 m a.s.l., the latter showing many erosive and depositional features of glacial origin. The geomorphology of this area is basically determined by periglacial landforms [23], showing many temporary shallow melt-water ponds, as well as permanent lakes (Figure 2). Most of the lakes at the plateau have a bowl-shaped morphology (Kettle lakes). This is a fluvioglacial landform formed during the glacier retreat as the result of the melting of a residual portion of ice, which is later filled with water.
Deposits from the Holocene dominate the beaches, while the upland region is composed by non-marine tuffs of the Lower Cretaceous. In general, the site shows a smooth topography where only a few formations of volcanic origin, such as Chester Cone (188 m) and Cerro Negro (143 m), stand out (Figure 2). Soils with an ornithogenic character, particularly in the coastal region, are also typical in the site [24,25], with a penguin occupation that started approximately 1000 years ago [26].
The weather in Byers is typical of the maritime Antarctic region. It is characterized by cloudy skies, windy conditions, and relatively high precipitation. The average temperatures during summer are commonly above freezing point of water. During winter, minimum temperatures average approximately −10 °C (extreme values of around −25 °C), and maxima are always below 0 °C [27]. Attending to regional atmospheric models, rainfall at the site is around 700 to 1000 mm [28]. Snow accumulates through the winter but mostly melts during summer, although some isolated snow packs persist. The site displays characteristically changeable weather, with rapid changes in winds and barometric pressure. This feature is imposed by alternating wind directions, although westerlies or easterlies dominate.
3. Hydrology, Nutrients Sources, and Environmental Gradients
Byers Peninsula is characterized by the existence of a complex drainage network, where many lakes, ponds, streams, and wetlands appear (Figure 3). The site is particularly rich in wetland areas extensively covered with microbial mats and moss carpets [19]. Attending to the deglaciation chronology deduced from the paleolimnological studies of Björck and co-workers [29], the glacier retreat started in Byers 4500 years before present, with a large part of the region becoming ice-free only 500 years later (Figure 1). This glacier retreat has originated a western-eastern distribution of lakes, with older lakes located towards the west, although the lakes are also influenced by the occurrence of faults and tectonic fractures [4]. In nearly the entire region an active permafrost layer appears until around 50–70 cm depth. Topsoil layers melt and warm during summer, thereby affecting the hydrological process in the catchments [30]. The predominant sandy soils and gravels facilitate groundwater flows. This dynamic activity virtually disappears during winter, when the hydric supply to the catchments is merely limited to the snow precipitation. The periglacial morphometry of the site and an underdeveloped stream network structure are typical of an early stage of landscape evolution [4]. Both surface and groundwater flows over the permafrost feed the lakes, though most lakes show point inflows and outflow. Ice dams formed during winter may close the lake’s outlet, impeding drainage. The rupture, as catastrophic events, of these dams during the summer melting causes sudden decreases in lake depth [18].
As shown in Table 1, there is a gradual ionic and nutrients enrichment in the water of streams and lakes as they are closer to the sea [18,31], which influence the species distribution patterns and the structure of plankton microbial communities [31,32,33]. Sea spray is the main salt source to Byers’ waterbodies, though the occurrence of an active chemical weathering has also been demonstrated [34,35], in such a way that both the lithology and landscape position have an effect on the streams geochemistry. The vegetation coverage may also influence the nutrient content in soils and its runoff towards lakes and streams in the site [36,37]. The shallowest lakes lack benthic mosses, although some of them show phototrophic microbial mats in their catchments or forming a ring along the lake. By contrast, most of the deepest lakes show well-developed or patchy bottom moss carpets of Drepanocladus longifolius. Particularly, microbial mats growing within the lake’s catchment [19] may supply the lakes with inorganic nutrients and organic carbon [38]. These fluxes mainly occur at the beginning of summer, when runoff is higher because of the snow melting. Nutrients might otherwise derive from the wind erosion of the catchment material [39], as also observed in the ephemeral wetlands of the Dry Valleys [40]. However, in coastal areas, and contrasting with inland lakes and streams, marine animals (elephant seals, penguins, etc.) are, in the former, the main nutrient sources by far [31,36], promoting a much higher trophic status. Roughly the entire south and west beaches, where one of the most studied lakes is located (Lake Refugio), are used by seals as a place to rest. On the other hand, an active Gentoo Penguin (Pygoscelis papua) colony is located northwest in the Peninsula, though it does not affect any of the studied lakes. In addition, the within-lake process may also regulate the availability of nutrients in the water column. The runoff is impeded when the lake surface is frozen [41]. The water column mixing linked to the onset of the ice-melting of the lake surface promotes convective movements that contribute to nutrient diffusion through the water column. Conversely, when ice cover melts out completely during the summer, there is no thermal stratification because surface warming is not enough to generate a temperature gradient.
4. Virioplankton
Two viral metagenomic studies have been performed in Lake Limnopolar, one of the most representative lakes of Byers Peninsula, focused on DNA [42] and RNA viruses [43], respectively. Both studies are outstanding as they report an unexpectedly rich genetic diversity of viruses for an Antarctic lake. The first study revealed a high proportion of unknown and single-stranded DNA viruses (Table 2) when the lake was under dim light conditions. However, the dominance of double-stranded viruses such as Phycodnaviridae (which are able of infecting freshwater and marine algae) increased in relative terms after ice melting, when the increases in nutrients and light availability allowed further development of phytoplankton. The seasonal estimation of the abundance of virus-like particles (VLP) performed at different depths in Lake Limnopolar by Rochera et al. [44] match the idea that virus assemblages change simultaneously with the host populations. These authors observed a notable variation of the viral vs. bacterial ratios, similar to those observed in other Antarctic lakes [45], suggesting that viruses abundance is also linked to the lake’s primary production. López–Bueno et al. [43] also reported, for the first time, the occurrence of RNA viruses in an Antarctic lake. They described a RNA virome dominated by the order Picornavirales (Table 2), which is able to infect a wide variety of hosts, but must particularly be associated with microorganisms. Interestingly, and in agreement with the approach of ecological modeling studies conducted in this lake [38], López–Bueno et al. [43] delve on the potential ecological connectivity existing between Lake Limnopolar and the microbial mats in the surrounding catchment, which may provide a pool of allochthonous viruses for the lake.
5. Prokaryotes in Plankton and Lacustrine Sediments
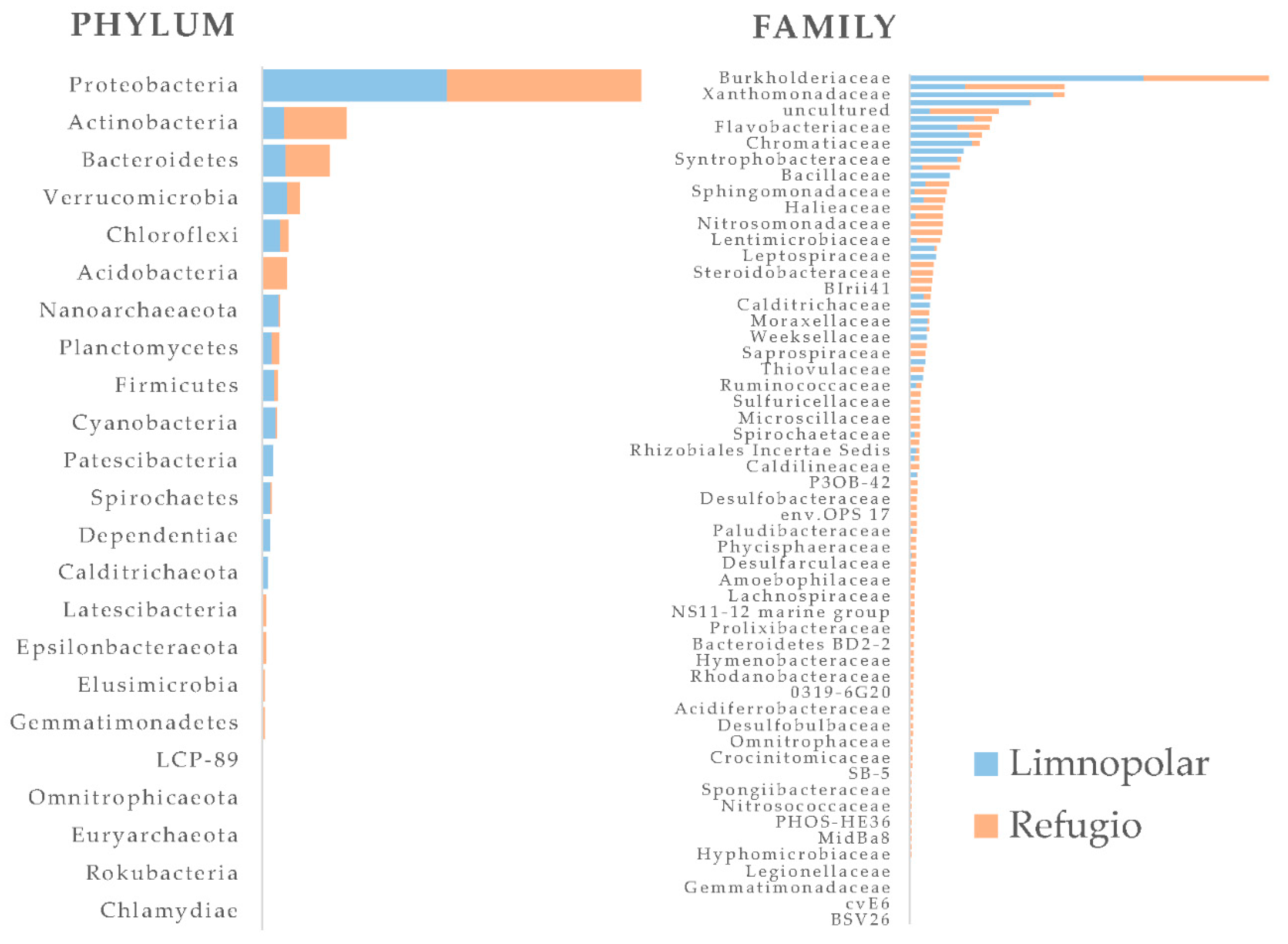
A bacterial metacommunity study carried out in a large set of Patagonian and Antarctic lakes, including some from Byers, showed that trends of bacterioplankton communities vary with latitude but are also reliant on major environmental factors such as the availability of light and nutrients [46]. This implies that both the geographic position of lakes and their local environmental conditions play an important structuring role. The different surveys conducted in Byers’ lakes to assess the bacterioplankton diversity are conclusive on the overall dominance of the phylum Proteobacteria (Figure 4), more specifically the order Betaproteobacteriales, and of the phylum Bacteroidetes [33,47,48]. Among Betaproteobacteria, it is remarkable the occurrence of an endemic clade of Polynucleobacter necessarius, which differs from any other member of this species found elsewhere outside Antarctica [49]. Some members of the bacterial phylum Chloroflexi have otherwise a major role in the eutrophic and impacted coastal lakes [33], particularly in the coastal eutrophic waters (Figure 4). Certain members of this phylum are important in the microbial assemblages of sites impacted by guano inputs, both in lacustrine sediments and soils, respectively [25,50]. Other taxa incidentally present in the bacterioplankton assemblages, particularly in coastal lagoons, are orders such as Clostridiales and Bacillales (Figure 4), which are also common in soils from the site affected by penguin depositions [25], mostly linked to fecal drops from marine animals. Even in the inland lakes, although they show a lower relevance, allochthonous bacterial taxa originating from the fecal drops of skuas and terns occur [33,51]. As observed for plankton, bacteria also dominated largely over Archaea in the different sediments environments studied in Byers [50].
Compared to previous molecular studies performed on the bacterioplankton in Byers [48,51], the taxonomic assignments performed by Picazo et al. [33] were made using the SILVA database of aligned ribosomal RNA gene sequences [52], which suggests some improvements compared to former studies. These involved important changes at high bacterial taxonomic ranks, such as that affecting to the former class Betaproteobacteria, which has now been reclassified as an order within the class Gammaproteobacteria. Regardless of these misalignments, the dominance of specific bacterial groups appears to be a common pattern in nutrient-limited Antarctic lakes ([53] and references therein), [54]). This includes, for instance, a significant abundance of genera, such as Flavobacterium or Polaromonas, as also observed in shallow lakes from the northern Victoria Land, East Antarctica [55].
On the other hand, most data related to the bacterioplankton community composition in Maritime Antarctica come from lakes of Signy Island [53], which are very similar to Byers’ lakes regarding its dependence on allochthonous nutrients inputs, also showing higher bacterial abundance compared to the more nutrient-deprived bacterial communities of the Continental region. Lakes at both locations reveal the dominance of members of globally distributed freshwater bacterial phyla, such as Alpha- and Betaproteobacteria, Actinobacteria, as well the Cytophaga-Flavobacterium-Bacteroidetes group. It is also common in both sites the dominance of the previously mentioned cluster of Polynucleobacter, as well as Achromobacter and Polaromonas, all of them from the family Burkholderiaceae. Likewise, both Sphingomonas and Rhodobacter are also the dominant Alphaproteobacteria in the lakes of both sites. Otherwise, an interesting outcome of Picazo et al. [33] work in Byers is that up to 25% of Zero-radius Operational Taxonomic Units (ZOTUs) detected could not be assigned at the genus level, which would suggest the presence of yet undescribed taxa that could represent locally adapted bacterial groups.
6. Phytoplankton
The abundance of taxon-specific carotenoids deduced by HPLC analyses from samples of different Byers lakes shows the dominance of chlorophytes (featured by specific pigments such as lutein, violaxanthin, neoxanthin, and chlorophyll-b), and both chrysophytes and diatoms (featured by fucoxanthin, diadinoxanthin, and chlorophyll-c) [31]. Microscopic observations of samples from Lake Limnopolar [44] show a community of plastidic nanoflagellates dominated by chrysophytes, such as Ochromonas sp. and Chromulina sp. and, particularly, by different species of Pseudokephyrion (Table 2). Diatom assemblages are exclusively composed by pennate forms (Table 2), mainly from the genera Navicula, Nitzschia, Achnanthes and Fragilaria, most of them reaching the lakes by runoff from the microbial mats within the lake catchment, where these genera are abundant. On the other hand, also within the same size range, non-flagellated Ankistrodesmus antarcticus and Ankyra sp., and the flagellate Chlamydomonas sp., dominated among chlorophytes. Cryptophytes, which are far less abundant, are largely represented by Chroomonas acuta. The dominance of chrysophytes over cryptophytes seems to be a common pattern in Maritime Antarctic lakes, opposite to that observed in Patagonian lakes, as demonstrated by an Illumina high-throughput sequencing study, including some Byers lakes [56].
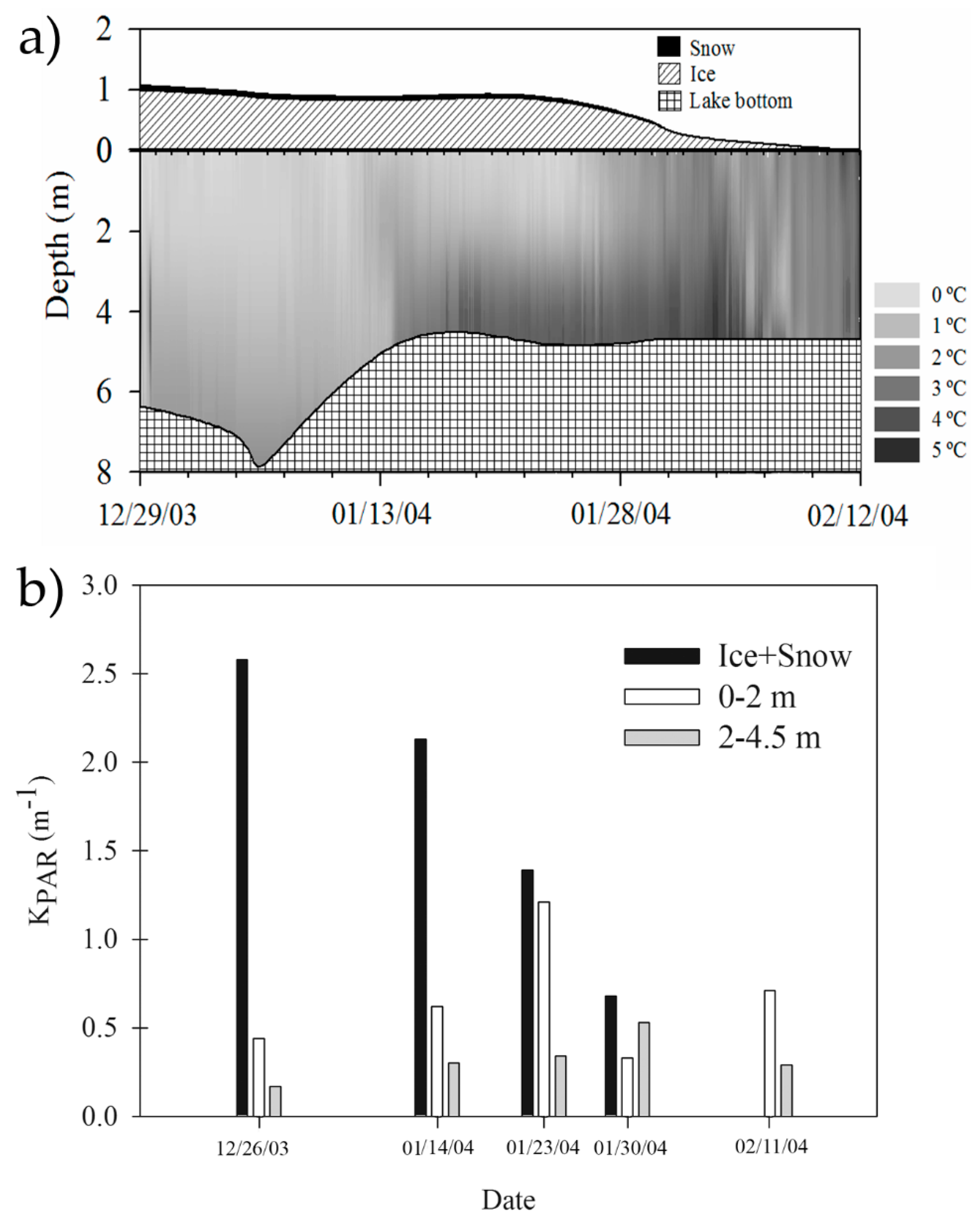
As shown by the trophic variables included in Table 1 (i.e., chlorophyll a, total phosphorus and bacterial abundance), shallow coastal lakes of Byers typically display eutrophic conditions because of the nutrients inputs due to marine animals and sea spray [31]. On the other hand, lakes at the plateau, which are generally deeper, are oligotrophic, with very limited inputs of nutrients. Still, some shallow inland lakes are mesotrophic because of the occurrence of a biologically mediated active nutrient release from the sediments. Furthermore, this trophic gradient, that extends from upland to coastal lakes, favors a progressive dominance of chlorophytes over chrysophytes and diatoms [31]. These findings demonstrate that the availability of nutrients in Byers lakes plays a significant role (especially in summer) in structuring the pelagic communities despite climatic constraints, similarly to non-polar systems. Major changes in the phytoplankton community occur during the transition to the ice-free period, just when the light availability and the fluxes of allochthonous nutrients towards the lakes increase. During this period, Byers’ lakes are highly dynamic and may exhibit significant spatial and temporal heterogeneity. This is related to the lake ice/snow cover phenology during the productive season, which affects the thermal structure and light availability, as illustrated in Figure 5. This heterogeneity was pointed out by Rochera et al. [41,44], who assessed the seasonal variations of both abiotic and biotic characteristics of Lake Limnopolar during three meteorologically contrasting years, as well as by Villaescusa et al. [32,51], who demonstrated the differential vertical distribution of the bacterioplankton assemblages in spite of the absence of summer stratification in the same lake.
In Lake Limnopolar, the highest algal abundance in surface layers usually occurs at the onset of ice melting due to the strong increase in nutrients and light availability, with certain species-specific preferences for light quantity, light quality, or both [41,44]. When the ice is completely covering the lake’s surface, some degree of stratification occurs. During this period, although not extremely abundant, picocyanobacteria proliferate at deep layers [44]. They specifically excite their phycobilin pigments by green light, which penetrates most efficiently. Conversely, other algal groups, such as diatoms, show a rather uniform distribution in the water column [44]. These diatoms, and also loricate chrysophytes, increase their abundance rapidly coinciding with higher water column turbulence just after ice melting, though many diatoms enter the lake with water runoff from biofilms and microbial mats within the catchment.
7. Protozoans and Metazoans
Heterotrophic or mixotrophic flagellated and ciliated protozoa are the main bacterial grazers [57]. In Antarctic lakes, these heterotrophic protists may play a key role when metazoans are scarce or virtually absent due to climate harshness, which is the case of the Antarctic continental region. As shown in Rochera et al. [41], both heterotrophic and autotrophic (or mixotrophic) members occur in the nanoflagellate assemblages from Byers lakes (Table 2). This involves, for instance, the presence of small chrysomonads (3–5 µm long) resembling Spumella and Oikomonas. Ciliate assemblages are mainly composed of small prostomatids and scuticociliates, such as Balanion planctonicum and Cyclidium glaucoma [41,58]. These nanoplanktonic ciliates may present conspicuous populations (up to 2.8 × 103 ind L−1), particularly during ice-free periods when the lake’s productivity is enhanced [41]. On the other hand, epibiotic forms of a peritrichous ciliate with a morphology resembling Vorticella profusely occur attached to copepods (Figure 6), which hypothetically might alternate sessile and free swimming motile phases [59].
Copepods (Boeckella poppei), fairy shrimps (Branchinecta gaini), both shown in Figure 6, and two species of insects (the dipterans Parochlus steinenii and Belgica antarctica), occur extensively in Byers freshwater ecosystems [3,18]. Additionally, the benthic cladoceran Macrothrix ciliata has also been occasionally observed in some of the lakes, representing the first record of this species in Antarctica [18]. Almost all the lakes from Byers show metazoan assemblages characterized by an overwhelming dominance of copepod B. poppei [18,31]. This is in contrast with the scarcity of metazoans in lakes of the continental region. For instance, only some copepods nauplii have been occasionally observed in Lake Joyce of the McMurdo Dry Valleys [60], as well as in lakes from the Amery Oasis [61]. The geographical distribution of B. poppei extends to the lakes of South America and the sub-Antarctic islands [62]. There are palaeoecological records supporting the idea of a post-glacial colonization of the maritime Antarctic region by this species after the Last Glacial Maximum, particularly during the early to mid-Holocene, [63]. Interestingly, this study suggests that it would originate from populations of B. poppei that maintained in continental refuges such as Amery Oasis, which is in contrast with the idea that the colonization of the Maritime Antarctic region would arise from South America. The anostracan Branchinecta gaini appears near the bottom of the deep lakes, with a minor impact on the pelagic compartment, although it is more abundant and active in shallow lakes [18,31]. However, it should be noted that larvae of Branchinecta gaini, that can be abundant around the melting of ice cover when eggs typically hatch out, could also act as plankton feeders, thus adding a higher trophic complexity in the water column. Nevertheless, this should be studied further in Byers to establish a more precise hypothesis.
8. Microbial Mats and Autotrophic Biofilms: Function and Diversity
Microbial mats, dominated by cyanobacteria and diatoms (Table 3), and phototrophic biofilms, extensively occur in Byers Peninsula [18,19,64,65,66]. Other algal groups such as chlorophytes also appear therein, though with much lower abundance, in these benthic communities. They are thin (from mm to a few cm) laminated microbial mats that may show vertically stratified layers differing in its physiological activity and species composition, which also explains their macroscopic differential aspect (Figure 7). In Byers Peninsula, they can be found in streams, puddle soils occupying soft clay sediments, and fluvial planes [37], as well as colonizing the bottom and the shores of very shallow lakes from the central plateau [19].
A great dominance of cyanobacteria is evident by the pigment signatures of Byers mats [19,65,66,67], which show a high abundance of carotenoids such as myxoxanthophyll and scytonemin. The earlier microscopic observations of these mats demonstrated the occurrence of several oscillatorian morphotypes which are also present in other microbial mats in continental Antarctica (Table 3), some of which could be assigned to Phormidium sp. and Leptolyngbya cf. antarctica [19]. The metagenomic study made by Picazo et al. [33] confirmed the occurrence of the families Leptolyngbyaceae and Phormidiaceae in water samples associated with these benthic environments. Nostocacean morphotypes (cf. Tolypothrix and Nostoc cf. commune) are also abundant in nutrient-starved mats, as well as in those showing high dehydration [19,68]. Actually, recent studies performed using molecular techniques in these mats demonstrate the dominance of the family Nostocaceae, comprising nearly half of the Cyanobacterial abundance compared to Oscillatoriaceae and Chroococcaceae [69]. Coccoid chlorophytes (green algae) may also occur in the mats [68], whereas Ulothrix sp. dominated only in permanently flooded stream biofilms [66]. On the other hand, and as observed for microbial mats, cyanobacteria were the main components in the stream biofilms suffering high drought stress. It should be noted; however, that differences observed between studies in the relative abundances of phototrophs in each mat type might respond to seasonal changes during the ice-free periods as suggested by Velázquez et al. [68].
Regarding diatoms diversity in these benthic communities (Table 3), a deep taxonomic study conducted in the site showed that nearly half of the 143 described taxa presented a restricted Antarctic biogeographic distribution [70], although cosmopolitan species such as Fragilaria capucina usually dominated [66]. The genus Fragilaria is also a common colonizer of rivers and brooks in several sub-Antarctic islands [71], and it is known to bloom opportunistically, thereby showing a rapid colonization capability [72] enabling its adaptation to highly dynamic environments such as the freshwater habitats of Byers Peninsula. Other sound taxonomic studies showed a high morphological diversity within particular diatom taxa (i.e., Pinnularia), thus revealing the occurrence of several previously unknown taxa [73,74,75]. Kopalová and Van De Vijver [70] conclude that salinity and nutrients largely explain the distribution of benthic freshwater diatoms in Byers Peninsula, although, particularly in streams, the substrate type can also be a relevant factor explaining the variability in the diatom assemblages [64].
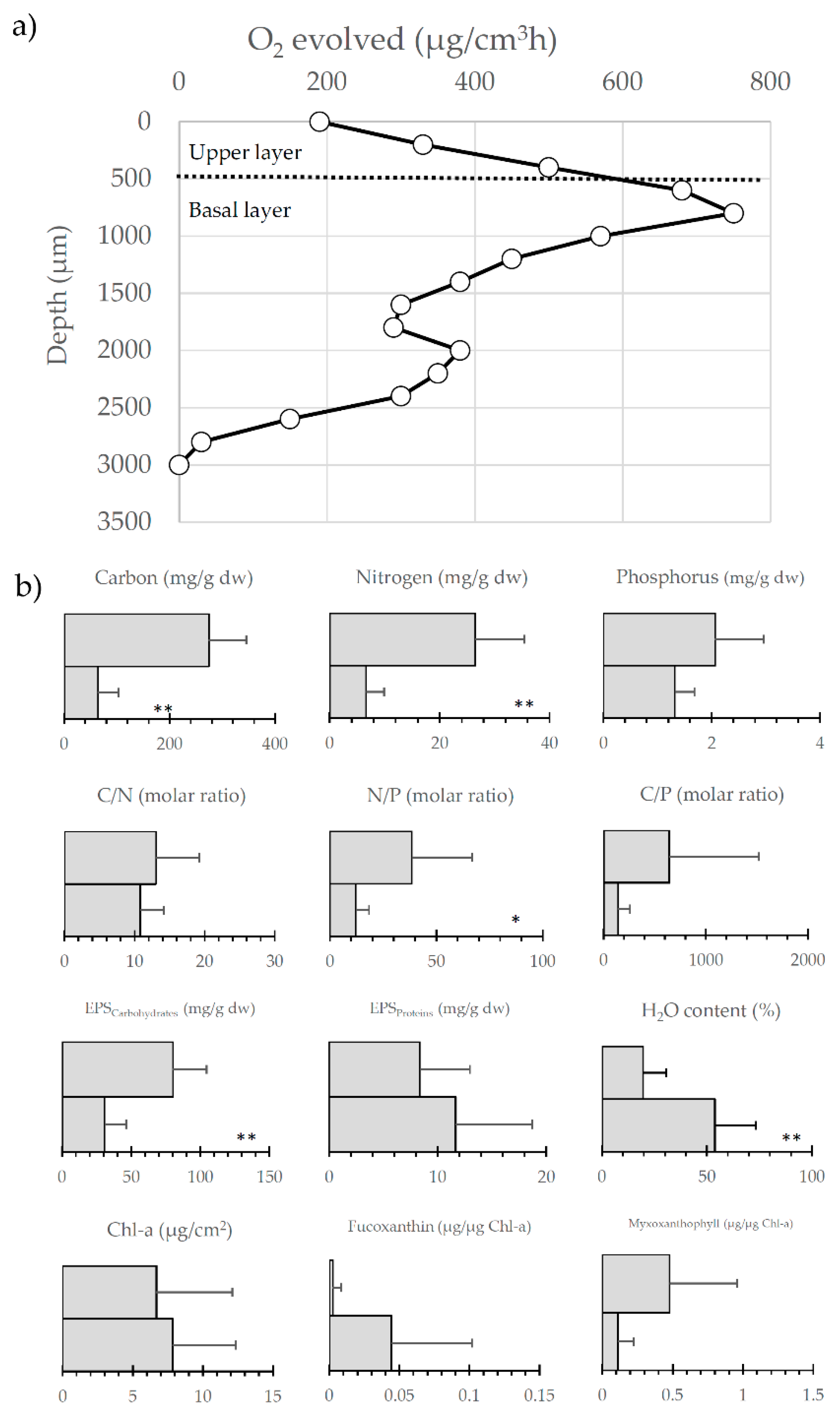
In Byers Peninsula, nutrients are generally supplied in very low amounts to these benthic communities. Accordingly, N2-fixation, performed by diazotrophs such as the aforementioned Nostocaceae, may constitute up to 23% of the assimilated nitrogen [19]. Interestingly, this way of N assimilation, and as well the photosynthetic activity, are greatly regulated by temperature, whereas other metabolisms (i.e., NO3 and urea uptake) appear to be less sensitive to this environmental factor [68]. In general, this nutrient scarcity influences the vertical structure of Byer’s microbial mats [65], whose structure is outlined in Figure 8. Thus, although the typical anoxic layers putatively containing sulphur-reducing and methanogenic bacteria do not appear in these mats, both the vertical profiles of O2 evolution [19] and some biochemical features [65] confirm the occurrence of an upper layer adapted to cope with environmental stressors, such as desiccation and high light radiation (by means of high contents of exopolysaccharides and the UV-screening scytonemin pigment, respectively). Contrarily, there is a more photosynthetically-active basal layer showing a more equilibrated C-N-P stoichiometry (Figure 8). The triggering of decomposition within these mats initiates in the upper layers when environmental conditions are adverse [76]. This is tightly related to the development of saprophytic members of the community, whose biomass increase with the seasonal rise of temperature and the ice melting [68].
Physiological studies carried out with microbial mats and stream biofilms in the site reveal their major role in the biogeochemical cycles, thereby facilitating the transfer of nutrients from the physical environment to the food webs [3,19,38,66,77]. The estimates of the photosynthetic rates in the mats attained in some of the former studies, all based on the use of NaH13CO3 as a tracer, range up to an order of magnitude from 0.74 to 4.2 mg C cm−2 h−1, being the differences directly related to the environmental conditions for the studied communities. Bayesian mixing models built for these microbial mats [69] indicate the occurrence of four trophic levels, namely, primary producers (cyanobacteria and diatoms), primary consumers (rotifers and tardigrades), secondary consumers (nematodes) and decomposers (fungi and heterotrophic bacteria). Interestingly, this study [69] suggests that there is a great coupling among the different microbial guilds, in such way that they are minimizing functional redundancies and optimizing its development for the short growth periods. Velázquez et al. [68] indicated that the heterotrophic microbes of the mats, composed by fungi and bacteria, transfer the microbial production to upper trophic levels. Accordingly, isotopic studies performed in these mats indicate that the nematodes are acting as omnivores, by feeding on bacteria as well as on other consumers [69].
9. Aquatic Food-Web Assembly
In natural systems under high environmental stress, the shortening of the trophic chain length is expected, as well as the prevalence of opportunistic growth strategies. Theoretically, this implies the lack of biological features counteracting environmental constraints, thereby producing erratic biological population dynamics mainly subjected to stochastic effects [78]. In Antarctica, biological interactions are assumed to play a minor role. However, the occurrence of some biological forces might partly rule out this idea. There are different studies stating that a top-down control, through grazing, on the structure of the planktonic microbial food web in the sub-Antarctic and maritime Antarctic lakes, including those from Byers, might exist, even though as a temporary mechanism [3]. As mentioned previously, most Byers lakes show an overwhelming dominance of the copepod Boeckella poppei and, conversely, very low abundance of rotifers, thereby representing a simplest food web complexity compared to lakes of neighboring locations, such as those of Signy Island [79] and King George [80]. This might respond either to control of rotifer populations by copepod predation, to a niche overlap, or even to the absence of fishes in Byers lakes, given that rotifer abundance in Patagonian lakes is relatively higher where fishes are present [81]. The copepod B.poppei behaves as an omnivore in the Antarctic lakes [82], which has also been shown in Byers lakes, showing an advantageous nutritional versatility for the survival in extreme and nutrient starved environments. Given the protozoans assemblages of these lakes and its abundance [18,31,44,58], it can be thought that bacterivory might exert a certain control on bacterial populations, which is more pronounced in oligotrophic systems compared to productive environments [83]. On the contrary, cascade effects from copepods, acting as omnivores and feeding on protozoans, could partly reduce the grazing pressure of protozoans on bacterioplankton [3]. This, together with the external fueling of bacterial production, could explain the high bacterial abundance in such oligotrophic lakes [3]. In fact, a trophic cascade in the plankton is more likely to occur if there are no major cut-offs in plankton size spectra, as it occurs in Lake Limnopolar [44].
In Byers lakes, during the transition to the ice-free periods, the r-strategists seem to dominate phytoplankton assemblages when its production is higher [41,44], with the occurrence of rapidly growing species that require turbulent conditions to thrive (sensu [84]). At this time, inorganic nutrients seem to be rapidly assimilated by inedible algae, which would lead to a large burying of biogenic carbon from the water column because of their high settling rates. Under these circumstances, retention of nutrients in the pelagic compartment could be favored if there are trophic interactions following the microbial loop.
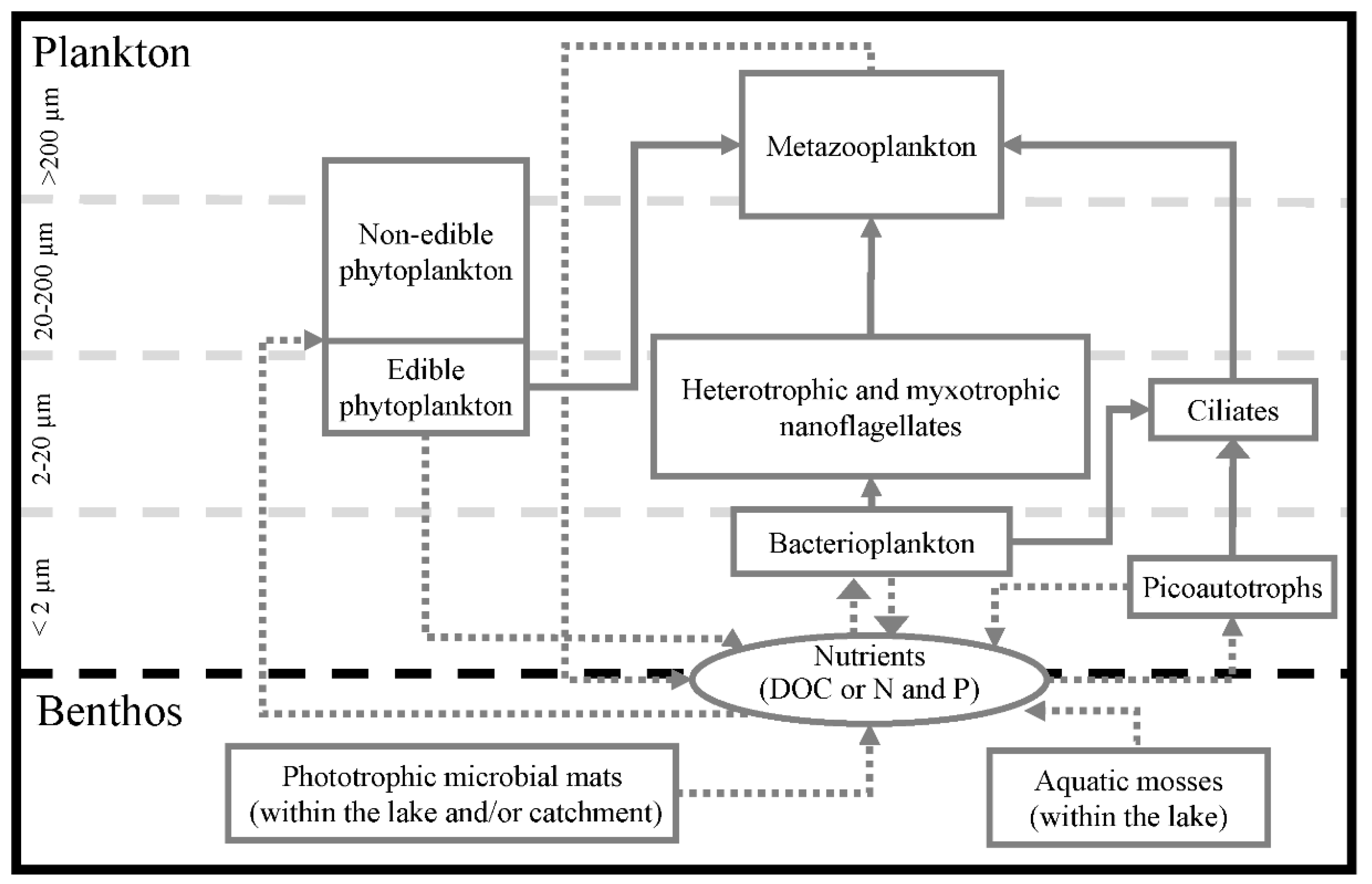
Attending to the modeling of carbon fluxes in Lake Limnopolar (Figure 9), most of the organic matter fueling the heterotrophic metabolism has an external origin, being mainly released by benthic mosses and microbial mats within the catchment [3,38]. This increases the trophic complexity of this lake, as indirectly deduced from other studies [43]. The main carbon source deriving from these mats could be the colloidal fraction of the extracellular polymeric substances (EPS), which includes water-soluble compounds that are exuded to the environment [65,66].
In summary, all these observations point out that biotic interactions may play a key role in structuring biological communities in Byers lakes despite the environmental harshness. In a framework of increasing temperatures that may dwindle the strength of climatic restrictions, a shift in the regional nutrient dynamics might eventually alter the functioning of these communities. However, top-down interactions may also exert a control role.
Supplementary Materials
The following materials are available online at https://0-www-mdpi-com.brum.beds.ac.uk/1424-2818/11/10/201/s1, Interactive Krona charts of Bacteria and Archaea domains represented by sequences of the 16S rRNA gene recovered from inland oligotrophic (Lake Limnopolar: Figure S1) and coastal eutrophic (Lake Refugio: Figure S2) lake surface waters. Files are provided in html format. Figure S1: Krona chart with the taxonomic affiliation of prokaryotes obtained by massive sequencing of the 16S rRNA from surface waters of the inland oligotrophic Lake Limnopolar; Figure S2: Krona chart with the taxonomic affiliation of prokaryotes obtained by massive sequencing of the 16S rRNA from surface waters of the coastal eutrophic Lake Refugio.
Author Contributions
Conceptualization, C.R., A.C.; Writing, Original Draft Preparation, C.R.; Writing, Review & Editing, A.C.; Funding Acquisition, A.C.
Funding
This study was supported by grant CGL2005-06549-C02-02/ANT, funded by the Spanish Ministry of Education and Science, whereas the current work of both authors is funded by the project CLIMAWET (CGL2015-69557-R) of the Spanish Ministry of Economy and Competitiveness, both granted to A.C. and co-financed by European FEDER funds, “A way to make Europe”.
Acknowledgments
We are indebted to the UTM (Maritime Technology Unit, CSIC) and Las Palmas crew (Spanish Navy) who provided logistic support to make possible research in Byers, as well as to our colleagues from the Limnopolar Research team, who shared with us all most limnological research performed in Byers.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hughes, K.A.; Cowan, D.A.; Wilmotte, A. Protection of Antarctic microbial communities—‘Out of sight, out of mind’. Front. Microbiol. 2015, 6, 151. [Google Scholar] [CrossRef] [PubMed]
- Camacho, A.; Rochera, C.; Villaescusa, J.A.; Velázquez, D.; Toro, M.; Rico, E.; Fernández-Valiente, E.; Justel, A.; Bañón, M.; Quesada, A. Maritime Antarctic lakes as sentinels of climate change. Int. J. Des. Nat. Ecodyn. 2012, 7, 239–250. [Google Scholar] [CrossRef]
- Camacho, A. Planktonic microbial assemblages and the potential effects of metazooplankton predation on the food web of lakes from the maritime Antarctica and sub-Antarctic islands. Rev. Environ. Sci. Biotechnol. 2006, 5, 167–185. [Google Scholar] [CrossRef]
- Mink, S.; López-Martínez, J.; Maestro, A.; Garrote, J.; Ortega, J.A.; Serrano, E.; Durán, J.J.; Schmid, T. Insights into deglaciation of the largest ice-free area in the South Shetland Islands (Antarctica) from quantitative analysis of the drainage system. Geomorphology 2014, 225, 4–24. [Google Scholar] [CrossRef]
- Priscu, J.C.; Fritsen, C.H.; Adams, E.E.; Giovannoni, S.J.; Paerl, H.W.; McKay, C.P.; Doran, P.T.; Gordon, D.A.; Lanoil, B.D.; Pinckney, J.L. Perennial Antarctic Lake Ice: An Oasis for Life in a Polar Desert. Science 1998, 208, 2095–2098. [Google Scholar] [CrossRef] [PubMed]
- Roberts, E.C.; Laybourn-Parry, J. Mixotrophic cryptophytes and their predators in the Dry Valley lakes of Antarctica. Freshw. Biol. 1999, 41, 737–746. [Google Scholar] [CrossRef]
- Karr, E.A.; Sattley, W.M.; Jung, D.O.; Madigan, M.T.; Achenbach, L.A. Remarkable diversity of phototrophic purple bacteria in a permanently frozen Antarctic lake. Appl. Environ. Microbiol. 2003, 69, 4910–4914. [Google Scholar] [CrossRef]
- Bowman, J.P.; McCammon, S.A.; Rea, S.M.; McMeekin, T.A. The microbial composition of three limnologically disparate hypersaline Antarctic lakes. FEMS Microbiol. Lett. 2006, 183, 81–88. [Google Scholar] [CrossRef]
- Yau, S.; Lauro, F.M.; DeMaere, M.Z.; Brown, M.V.; Thomas, T.; Raftery, M.J.; Andrews-Pfannkoch, C.; Lewis, M.; Hoffman, J.M.; Gibson, J.A.; et al. Virophage control of antarctic algal host–virus dynamics. Proc. Natl. Acad. Sci. USA 2011, 108, 6163–6168. [Google Scholar] [CrossRef]
- Sabbe, K.; Verleyen, E.; Hodgson, D.A.; Vanhoutte, K.; Vyverman, W. Benthic diatom flora of freshwater and saline lakes in the Larsemann Hills and Rauer Islands, East-Antarctica. Antarct. Sci. 2003, 15, 227–248. [Google Scholar] [CrossRef]
- Taton, A.; Grubisic, S.; Balthasart, P.; Hodgson, D.A.; Laybourn-Parry, J.; Wilmotte, A. Biogeographical distribution and ecological ranges of benthic cyanobacteria in East Antarctic lakes. FEMS Microbiol. Ecol. 2006, 57, 272–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laybourn-Parry, J.; Quayle, W.C.; Henshaw, T.; Ruddell, A.; Marchant, H.J. Life on the edge: The plankton and chemistry of Beaver Lake, an ultraoligotrophic epishelf lake, Antarctica. Freshw. Biol. 2001, 46, 1205–1217. [Google Scholar] [CrossRef]
- Cremer, H.; Gore, D.; Hultzsch, N.; Melles, M.; Wagner, B. The diatom flora and limnology of lakes in the Amery Oasis, East Antarctica. Polar Biol. 2004, 27, 513–531. [Google Scholar] [CrossRef]
- Pearce, D.A. The structure and stability of the bacterioplankton community in Antarctic freshwater lakes, subject to extremely rapid environmental change. FEMS Microbiol. Ecol. 2005, 53, 61–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinocur, A.; Maidana, N.I. Spatial and temporal variations in moss-inhabiting summer diatom communities from Potter Peninsula (King George Island, Antarctica). Polar Biol. 2010, 33, 443–455. [Google Scholar] [CrossRef]
- Callejas, C.; Gill, P.R.; Catalan, A.I.; Azziz, G.; Castro-Sowinski, S.; Batista, S. Phylotype diversity in a benthic cyanobacterial mat community on King George Island, maritime Antarctica. World J. Microbiol. Biotechnol. 2011, 27, 1507–1512. [Google Scholar] [CrossRef]
- Quesada, A.; Camacho, A.; Rochera, C.; Velàzquez, D. Byers Peninsula: A reference site for coastal, terrestrial and limnetic ecosystem studies in maritime Antarctica. Polar Sci. 2009, 3, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Toro, M.; Camacho, A.; Rochera, C.; Rico, E.; Bañón, M.; Fernández-Valiente, E.; Marco, E.; Justel, A.; Avendaño, M.C.; Ariosa, Y.; et al. Limnological characteristics of the freshwater ecosystems of Byers Peninsula, Livingston Island, in maritime Antarctica. Polar Biol. 2007, 30, 635–649. [Google Scholar] [CrossRef]
- Fernández-Valiente, E.; Camacho, A.; Rochera, C.; Rico, E.; Vincent, W.F.; Quesada, A. Community structure and physiological characterization of microbial mats in Byers Peninsula, Livingston Island (SouthShetland Islands, Antarctica). FEMS Microbiol. Ecol. 2007, 59, 377–385. [Google Scholar] [CrossRef]
- Benayas, L.; Pertierra, L.; Tejedo, P.; Lara, F.; Bermudez, O.; Hughes, K.A.; Quesada, A. A review of scientific research trends within ASPA No. 126 Byers Peninsula, South Shetland Islands, Antarctica. Antarct. Sci. 2013, 25, 128–145. [Google Scholar] [CrossRef] [Green Version]
- Hathway, B.; Lomas, S.A. The Upper Jurassic-Lower Cretaceous Byers Group, South Shetland Islands, Antarctica: Revised stratigraphy and regional correlations. Cretac. Res. 1998, 19, 43–67. [Google Scholar] [CrossRef]
- Navas, A.; Soto, J.; López-Martínez, J. Radionuclides in soils of Byers Peninsula, South Shetland Islands, Western Antarctica. Appl. Radiat. Isot. 2005, 62, 809–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Martínez, J.; Serrano, E.; Schmid, T.; Mink, S.; Linés, C. Periglacial processes and landforms in the South Shetland Islands (northern Antarctic Peninsula region). Geomorphology 2012, 155–156, 62–79. [Google Scholar] [CrossRef]
- Moura, P.A.; Francelino, M.R.; Schaefer, C.E.G.R.; Simas, F.N.B.; de Mendoça, B.A.F. Distribution and characterization of soils and landform relationships in Byers Peninsula, Livingston Island, Maritime Antarctica. Geomorphology 2012, 155–156, 45–54. [Google Scholar] [CrossRef]
- Santamans, A.C.; Boluda, R.; Picazo, A.; Gil, C.; Ramos-Miras, J.; Tejedo, P.; Pertierra, L.R.; Benayas, J.; Camacho, A. Soil features in rookeries of Antarctic penguins reveal sea to land biotransport of chemical pollutants. PLoS ONE 2017, 12, e0181901. [Google Scholar] [CrossRef] [PubMed]
- Emslie, S.D.; Baumann, K.; van Tuinen, M. Late Holocene occupation of Gentoo Penguins (Pygoscelis papua) on Byers Peninsula, Livingston Island, Antarctica. Polar Biol. 2010, 34, 283–290. [Google Scholar] [CrossRef]
- Bañón, M.; Justel, A.; Velázquez, D.; Quesada, A. Regional weather survey on Byers Peninsula, Livingston Island, South Shetland Islands, Antarctica. Antarct. Sci. 2013, 25, 146–156. [Google Scholar] [CrossRef]
- Van Lipzig, N.P.M.; King, J.C.; Lachlan-Cope, T.A.; Van den Broeke, M.R. Precipitation, sublimation and snow drift in the Antarctic Peninsula regionfrom a regional atmospheric model. J. Geophys. Res. 2004, 109, D24106. [Google Scholar] [CrossRef]
- Björck, S.; Olsson, S.; Ellis-Evans, C.; Håkansson, H.; Humlum, O.; De Lirio, J.M. Late Holocene palaeoclimatic records from lake sediments on James Ross Island, Antarctica. Palaeogeogr. Palaeocl. 1996, 121, 195–220. [Google Scholar] [CrossRef]
- Oliva, M.; Hrbacek, F.; Ruiz-Fernández, J.; de Pablo, M.A.; Vieira, G.; Ramos, M.; Antoniades, D. Active layer dynamics in three topographically distinct lake catchments in Byers Peninsula (Livingston Island, Antarctica). Catena 2017, 149, 548–559. [Google Scholar] [CrossRef]
- Rochera, C.; Toro, M.; Rico, E.; Fernández-Valiente, E.; Villaescusa, J.A.; Picazo, A.; Quesada, A.; Camacho, A. Structure of planktonic microbial communities along a trophic gradient in lakes of Byers Peninsula. Antarct. Sci. 2013, 25, 277–287. [Google Scholar] [CrossRef]
- Villaescusa, J.A.; Casamayor, E.O.; Rochera, C.; Velázquez, D.; Chicote, A.; Quesada, A.; Camacho, A. A close link between bacterial community composition and environmental heterogeneity in maritime Antarctic lakes. Int. Microbiol. 2010, 13, 67–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picazo, A.; Rochera, C.; Villaescusa, J.A.; Miralles-Lorenzo, J.; Velázquez, D.; Quesada, A.; Camacho, A. Bacterioplankton community composition along environmental gradients in lakes from Byers Peninsula (Maritime Antarctica) as determined by Next-Generation Sequencing. Front. Microbiol. 2019, 10, 908. [Google Scholar] [CrossRef] [PubMed]
- Navas, A.; López-Martínez, J.; Casas, J.; Machín, J.; Durán, J.J.; Serrano, E.; Cuchi, J.-A.; Mink, S. Soil characteristics on varying lithological substrates in the South Shetland Islands, maritime Antarctica. Geoderma 2008, 144, 123–139. [Google Scholar] [CrossRef] [Green Version]
- Lyons, W.B.; Welch, K.A.; Welch, S.A.; Camacho, A.; Rochera, C.; Michaud, L.; de Wit, R.; Carey, A.E. Geochemistry of streams from Byers Peninsula, Livingston Island. Antarct. Sci. 2013, 25, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Cabrerizo, A.; Dachs, J.; Barceló, D.; Jones, K.C. Influence of organic matter content and human activities on the occurrence of organic pollutants in Antarctic soils, lichens, grass, and mosses. Environ. Sci. Technol. 2012, 46, 1396–1405. [Google Scholar] [CrossRef]
- Otero, X.L.; Fernández, S.; de Pablo Hernandez, M.A.; Nizoli, E.C.; Quesada, A. Plant communities as a key factor in biogeochemical processes involving micronutrients (Fe, Mn, Co, and Cu) in Antarctic soils (Byers Peninsula, maritime Antarctica). Geoderma 2013, 195–196, 145–154. [Google Scholar] [CrossRef]
- Villaescusa, J.A.; Jørgensen, S.E.; Rochera, C.; Velázquez, D.; Quesada, A.; Camacho, A. Carbon dynamics modelization and biological community sensitivity to temperature in an oligotrophic freshwater Antarctic lake. Ecol. Model. 2016, 319, 21–30. [Google Scholar] [CrossRef]
- Fassnacht, S.R.; Toro, M.; Meiman, P.J.; Whitt, Z.C. The effect of aeolian deposition on the surface roughness of melting snow, Byers Peninsula, Antarctica. Hydrol. Process. 2010, 24, 2007–2013. [Google Scholar] [CrossRef]
- Moorhead, D.L. Mesoscale dynamics of ephemeral wetlands in the Antarctic Dry Valleys: Implications to production and distribution of organic matter. Ecosystems 2007, 10, 87–95. [Google Scholar] [CrossRef]
- Rochera, C.; Justel, A.; Fernández-Valiente, E.; Bañón, M.; Rico, E.; Toro, M.; Camacho, A.; Quesada, A. Interannual meteorological variability and its effects on a lake from maritime Antarctica. Polar Biol. 2010, 33, 1615–1628. [Google Scholar] [CrossRef]
- López-Bueno, A.; Tamames, J.; Velázquez, D.; Moya, A.; Quesada, A.; Alcamí, A. High diversity of the viral community from an Antarctic lake. Science 2009, 326, 858–861. [Google Scholar] [CrossRef] [PubMed]
- López-Bueno, A.; Rastrojo, A.; Peiró, R.; Arenas, M.; Alcamí, A. Ecological connectivity shapes quasispecies structure of RNA viruses in an Antarctic lake. Mol. Ecol. 2015, 24, 4812–4825. [Google Scholar] [CrossRef] [Green Version]
- Rochera, C.; Quesada, A.; Toro, M.; Rico, E.; Camacho, A. Plankton assembly in an ultra-oligotrophic Antarctic lake over the summer transition from the ice-cover to ice-free period: A size spectra approach. Polar Sci. 2017, 11, 72–82. [Google Scholar] [CrossRef]
- Madan, N.J.; Marshall, W.A.; Laybourn-Parry, J. Virus and microbial loop dynamics over an annual cycle in three contrasting Antarctic lakes. Freshw. Biol. 2005, 50, 1291–1300. [Google Scholar] [CrossRef]
- Schiaffino, M.R.; Unrein, F.; Gasol, J.M.; Massana, R.; Balague, V.; Izaguirre, I. Bacterial community structure in a latitudinal gradient of lakes: The roles of spatial versus environmental factors. Freshw. Biol. 2011, 56, 1973–1991. [Google Scholar] [CrossRef]
- Villaescusa, J.A.; Rochera, C.; Velázquez, D.; Rico, E.; Quesada, A.; Camacho, A. Bacterioplankton summer dynamics in a maritime Antarctic lake. Limnetica 2013, 32, 253–268. [Google Scholar]
- Papale, M.; Rizzo, C.; Villescusa, J.A.; Rochera, C.; Camacho, A.; Michaud, L.; Lo Giudice, A. Prokaryotic assemblages in the maritime Antarctic Lake Limnopolar (Byers Peninsula, South Shetland Islands). Extremophiles 2017, 21, 947–961. [Google Scholar] [CrossRef]
- Hahn, M.W.; Koll, U.; Jezberová, J.; Camacho, A. Global phylogeography of pelagic Polynucleobacter bacteria: Restricted geographic distribution of subgroups, isolation by distance and influence of climate. Environ. Microbiol. 2015, 17, 829–840. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Michaud, L.; Lo Giudice, A.; Lentini, V.; Rochera, C.; Camacho, A.; Luciana Maugeri, T. Prokaryotic community in lacustrine sediments of Byers Peninsula (Livingston Island, Maritime Antarctica). Microb. Ecol. 2016, 71, 387–400. [Google Scholar] [CrossRef]
- Villaescusa, J.A.; Casamayor, E.O.; Rochera, C.; Quesada, A.; Michaud, L.; Camacho, A. Heterogeneous vertical structure of the bacterioplankton community in a non-stratified Antarctic lake. Antarct. Sci. 2013, 5, 229–238. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and ‘‘All-species Living Tree Project (LTP)’’ taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef]
- Pearce, D.A.; Galand, P.E. Microbial biodiversity and biogeography. In Polar Lakes and Rivers, Limnology of Arctic and Antarctic Aquatic Ecosystems, 1st ed.; Vincent, W., Laybourn-Parry, J., Eds.; Oxford University Press: New York, NY, USA, 2008; pp. 213–230. [Google Scholar] [CrossRef]
- Valdespino-Castillo, P.M.; Cerqueda-García, D.; Espinosa, A.C.; Batista, S.; Merino-Ibarra, M.; Tas, N.; Alcántara-Hernández, R.J.; Falcón, L.I. Microbial distribution and turnover in Antarctic microbial mats highlight the relevance of heterotrophic bacteria in low-nutrient environments. FEMS Microbiol. Ecol. 2018, 94, fiy129. [Google Scholar] [CrossRef]
- Michaud, L.; Caruso, C.; Mangano, S.; Interdonato, F.; Bruni, V.; Lo Giudice, A. Predominance of Flavobacterium, Pseudomonas, and Polaromonas within the prokaryotic community of freshwater shallow lakes in the northern Victoria Land, East Antarctica. FEMS Microbiol. Ecol. 2012, 82, 391–404. [Google Scholar] [CrossRef]
- Schiaffino, M.R.; Lara, E.; Fernández, L.D.; Balagué, V.; Singer, D.; Seppey, C.C.W.; Massana, R.; Izaguirre, I. Microbial eukaryote communities exhibit robust biogeographical patterns along a gradient of Patagonian and Antarctic lakes. Environ. Microbiol. 2016, 18, 5249–5264. [Google Scholar] [CrossRef] [Green Version]
- Simek, K.; Bobkova, J.; Macek, M.; Nedoma, J.; Psenner, R. Ciliate grazing on picoplankton in a eutrophic reservoir during the summer phytoplankton maximum: A study at the species and community level. Limnol. Oceanogr. 1995, 40, 1077–1090. [Google Scholar] [CrossRef]
- Petz, W.; Valbonesi, A.; Schiftner, U.; Quesada, A.; Ellis-Evans, J.C. Ciliate biogeography in Antarctic and Arctic freshwater ecosystems: Endemism or global distribution of species? FEMS Microb. Ecol. 2007, 59, 396–408. [Google Scholar] [CrossRef]
- Lynn, D.H. The Ciliated Protozoa: Characterization, Classification and Guide to the Literature, 3rd ed.; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Roberts, E.C.; Priscu, J.C.; Laybourn-Parry, J. Microplankton dynamics in a perennially ice-covered Antarctic lake—Lake Hoare. Freshw. Biol. 2004, 49, 853–869. [Google Scholar] [CrossRef]
- Bayly, I.A.E.; Gibson, J.A.E.; Wagner, B.; Swadling, K.M. Taxonomy, ecology and zoogeography of two East Antarctic freshwater calanoid copepod species: Boeckella poppei and Gladioferens antarcticus. Antarct. Sci. 2003, 15, 439–448. [Google Scholar] [CrossRef]
- Menu-Marque, S.; Morrone, J.J.; De Mitrovich, C.L. Distributional patterns of the South American species of Boeckella (Copepoda: Centropagidae): A track analysis. J. Crustac. Biol. 2000, 20, 262–272. [Google Scholar] [CrossRef]
- Gibson, J.A.E.; Bayly, I.A.E. New insights into the origins of crustaceans of Antarctic lakes. Antarct. Sci. 2007, 19, 157–164. [Google Scholar] [CrossRef]
- Pla-Rabes, S.; Toro, M.; Van De Vijver, B.; Rochera, C.; Villaescusa, J.A.; Camacho, A.; Quesada, A. Stability and endemicity of benthic diatom assemblages from different substrates in a maritime stream on Byers Peninsula, Livingston Island, Antarctica: The role of climate variability. Antarct. Sci. 2013, 25, 254–269. [Google Scholar] [CrossRef]
- Rochera, C.; Villaescusa, J.A.; Velázquez, D.; Fernández-Valiente, E.; Quesada, A.; Camacho, A. Vertical structure of bi-layered microbial mats from Byers Peninsula, Maritime Antarctica. Antarct. Sci. 2013, 25, 270–276. [Google Scholar] [CrossRef]
- Rochera, C.; Fernández-Valiente, E.; Van de Vijver, B.; Rico, E.; Toro, M.; Vincent, W.F.; Quesada, A.; Camacho, A. Community structure and photosynthetic activity of benthic biofilms from a waterfall in the maritime Antarctica. Polar Biol. 2013, 36, 1709–1722. [Google Scholar] [CrossRef]
- Camacho, A.; Rochera, C.; Hennebelle, R.; Ferrari, C.; Quesada, A. Total mercury and methyl-mercury contents and accumulation in polar microbial mats. Sci. Total Environ. 2015, 509–510, 145–153. [Google Scholar] [CrossRef]
- Velázquez, D.; Jungblut, A.D.; Rochera, C.; Rico, E.; Camacho, A.; Quesada, A. Trophic interactions in microbial mats on Byers Peninsula, maritime Antarctica. Polar Biol. 2017, 40, 1115–1126. [Google Scholar] [CrossRef]
- Almela, P.; Velázquez, D.; Rico, E.; Justel, A.; Quesada, A. Carbon pathways through the food web of a microbial mat from Byers Peninsula, Antarctica. Front. Microbiol. 2019, 10, 628. [Google Scholar] [CrossRef]
- Kopalová, K.; Van De Vijver, B. Structure and ecology of freshwater benthic diatom communities from Byers Peninsula, Livingston Island, South Shetland Islands. Antarct. Sci. 2013, 25, 239–253. [Google Scholar] [CrossRef]
- Van de Vijver, B.; Beyens, L. Freshwater diatoms from Ile de la Possession (Crozet Archipelago, Sub-Antarctica): An ecological assessment. Polar Biol. 1999, 22, 178–188. [Google Scholar] [CrossRef]
- Lotter, A.F.; Bigler, C. Do diatoms in the Swiss Alps reflect the length of ice-cover? Aquat. Sci. 2000, 62, 125–141. [Google Scholar] [CrossRef] [Green Version]
- Van De Vijver, B.; Agius, J.T.; Gibson, J.A.E.; Quesada, A. An unusual spine-bearing Pinnularia species from the Antarctic Livingston Island (South Shetland Islands). Diatom Res. 2009, 24, 431–441. [Google Scholar] [CrossRef]
- Van De Vijver, B.; Zidarova, R. Five new taxa in the genus Pinnularia sectio Distantes (Bacillariophyta) from Livingston Island (South Shetland Islands). Phytotaxa 2011, 24, 39–50. [Google Scholar] [CrossRef]
- Zidarova, R.; Kopalová, K.; Van De Vijver, B. The genus Pinnularia (Bacillariophyta) excluding the section Distantes on Livingston Island (South Shetland Islands) with the description of twelve new taxa. Phytotaxa 2012, 44, 11–37. [Google Scholar] [CrossRef]
- Velázquez, D.; López-Bueno, A.; Aguirre de Cárcer, D.; de los Ríos, A.; Alcamí, A.; Quesada, A. Ecosystem function decays by fungal outbreaks in Antarctic microbial mats. Sci. Rep. 2016, 6, 22954. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, D.; Rochera, C.; Camacho, A.; Quesada, A. Temperature effects on carbon and nitrogen metabolism in some Maritime Antarctic freshwater phototrophic communities. Polar Biol. 2011, 34, 1045–1055. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecology: The Experimental Analysis of Distribution and Abundance, 5th ed.; Benjamin Cummings: Menlo Park, CA, USA, 2001. [Google Scholar]
- Butler, H.G.; Edworthy, M.G.; Ellis-Evans, J.C. Temporal plankton dynamics in an oligotrophic maritime Antarctic lake. Freshw. Biol. 2000, 43, 215–230. [Google Scholar] [CrossRef]
- Pociecha, A.; Dumont, H.J. Life cycle of Boeckella poppei Mrazek and Branchinecta gaini Daday (King George Island, South Shetlands). Polar Biol. 2008, 31, 245–248. [Google Scholar] [CrossRef]
- Reissig, M.; Trochine, C.; Queimaliños, C.; Balseiro, E.; Modenutti, B. Impact of fish introduction on planktonic food webs in lakes of the Patagonian Plateau. Biol Conserv. 2006, 132, 437–447. [Google Scholar] [CrossRef]
- Butler, H.G.; Atkinson, A.; Gordon, M. Omnivory and predation impact of the calanoid copepod Boeckella poppei in a maritime Antarctic lake. Polar Biol. 2005, 28, 815–821. [Google Scholar] [CrossRef]
- Gasol, J.M.; Pedros-Aliò, C.; Vaqué, D. Regulation of bacterial assemblages in oligotrophic plankton systems: Results from experimental and empirical approaches. Anton. Leeuw. J. Microb. 2002, 81, 435–452. [Google Scholar] [CrossRef]
- Reynolds, C.S. Phytoplankton periodicity: The interactions of form, function and environmental variability. Freshw. Biol. 1984, 14, 111–142. [Google Scholar] [CrossRef]
Figure 1.
Location of Byers Peninsula within the Maritime Antarctica, and main lakes within the Peninsula. The aerial view of Livingston Island shows the glacier (Rotch Dome showed) retreat in its westernmost part, which corresponds to Byers Peninsula. The dashed line delimits the coastal and central plateau area.
Figure 1.
Location of Byers Peninsula within the Maritime Antarctica, and main lakes within the Peninsula. The aerial view of Livingston Island shows the glacier (Rotch Dome showed) retreat in its westernmost part, which corresponds to Byers Peninsula. The dashed line delimits the coastal and central plateau area.

Figure 2.
Photographs of some representative lakes from Byers Peninsula: (a) Lake Limnopolar; (b) Lake Cerro Negro; (c) Lake Somero; (d) Lake Refugio; (e) Lake Turbio; (f) Chester Cone Lake.
Figure 2.
Photographs of some representative lakes from Byers Peninsula: (a) Lake Limnopolar; (b) Lake Cerro Negro; (c) Lake Somero; (d) Lake Refugio; (e) Lake Turbio; (f) Chester Cone Lake.

Figure 3.
Landscape photographs showing components of the drainage network of Byers Peninsula.

Figure 4.
Normalized reads assigned to prokaryotic phyla and families obtained by massive sequencing of the 16S rRNA encoding DNA gene recovered from surface waters of the inland oligotrophic Lake Limnopolar and the coastal eutrophic Lake Refugio. Data redrawn from Picazo et al. [33]. Interactive Krona charts are provided as Supplementary Material. The complete taxonomic affiliation of families shown in the figure can be accessed in the interactive krona charts provided as Supplementary Material for Lake Limnopolar (Figure S1) and Lake Refugio (Figure S2).
Figure 4.
Normalized reads assigned to prokaryotic phyla and families obtained by massive sequencing of the 16S rRNA encoding DNA gene recovered from surface waters of the inland oligotrophic Lake Limnopolar and the coastal eutrophic Lake Refugio. Data redrawn from Picazo et al. [33]. Interactive Krona charts are provided as Supplementary Material. The complete taxonomic affiliation of families shown in the figure can be accessed in the interactive krona charts provided as Supplementary Material for Lake Limnopolar (Figure S1) and Lake Refugio (Figure S2).

Figure 5.
Major physical changes occurring in the water column of Lake Limnopolar during the 2003/04 summer transition from the ice-cover to ice-free period. (a) Time-course of snow/ice cover and the temperature regime in Lake Limnopolar obtained with a thermistors chain; (b) Changes in the extinction coefficient of the photosynthetic active radiation (KPAR) in the snow/ice cap and in the water column. Dates as month/day/year. Redrawn from Rochera et al. [41].
Figure 5.
Major physical changes occurring in the water column of Lake Limnopolar during the 2003/04 summer transition from the ice-cover to ice-free period. (a) Time-course of snow/ice cover and the temperature regime in Lake Limnopolar obtained with a thermistors chain; (b) Changes in the extinction coefficient of the photosynthetic active radiation (KPAR) in the snow/ice cap and in the water column. Dates as month/day/year. Redrawn from Rochera et al. [41].

Figure 6.
Microscope image showing two main metazoans present in Byers lakes. (a) Adult of the copepod Boeckella poppei; (b) Adults of the fairy shrimp Branchinecta gaini; (c–e) Pictures showing epibiotic ciliates and diatoms attached to the surface of Boeckella poppei. Note that the scale of the photographs is different. Bar scales are included to point out the different size of B. poppei and B. gaini.
Figure 6.
Microscope image showing two main metazoans present in Byers lakes. (a) Adult of the copepod Boeckella poppei; (b) Adults of the fairy shrimp Branchinecta gaini; (c–e) Pictures showing epibiotic ciliates and diatoms attached to the surface of Boeckella poppei. Note that the scale of the photographs is different. Bar scales are included to point out the different size of B. poppei and B. gaini.

Figure 7.
Photographs of different microbial mats studied in Byers Peninsula. (a,b) soil mat; (c,d) pond mat; (e,f) stream mat (names according to Fernandez-Valiente et al. [19]).
Figure 7.
Photographs of different microbial mats studied in Byers Peninsula. (a,b) soil mat; (c,d) pond mat; (e,f) stream mat (names according to Fernandez-Valiente et al. [19]).

Figure 8.
Functional structure of a typical microbial mat from Byers Peninsula: (a) Vertical profiles of oxygen evolution within the mat measured with microelectrodes; (b) Allocation, in the surface and basal layers, of different structural characteristics. Variables are major nutrients and their molar ratios, exopolysaccharides (EPS), both carbohydrates and proteins, percentage of water content, and main photosynthetic pigments. The latter include taxa specific carotenoids for the main photosynthetic organisms, fucoxanthin, as the main marker for diatoms, and myxoxanthophyll, as the main marker for cyanobacteria. One or two asterisks indicate statistical differences with significance at p < 0.05 and p < 0.01 respectively, between both layers.
Figure 8.
Functional structure of a typical microbial mat from Byers Peninsula: (a) Vertical profiles of oxygen evolution within the mat measured with microelectrodes; (b) Allocation, in the surface and basal layers, of different structural characteristics. Variables are major nutrients and their molar ratios, exopolysaccharides (EPS), both carbohydrates and proteins, percentage of water content, and main photosynthetic pigments. The latter include taxa specific carotenoids for the main photosynthetic organisms, fucoxanthin, as the main marker for diatoms, and myxoxanthophyll, as the main marker for cyanobacteria. One or two asterisks indicate statistical differences with significance at p < 0.05 and p < 0.01 respectively, between both layers.

Figure 9.
Conceptual model of the Lake Limnopolar planktonic microbial food web and dissolved nutrients transfers. Solid arrows represent phagotrophic organic matter transfers from the donor compartment to the consumer. Dotted arrows represent dissolved nutrients transfer. Planktonic organisms are depicted based on their size range.
Figure 9.
Conceptual model of the Lake Limnopolar planktonic microbial food web and dissolved nutrients transfers. Solid arrows represent phagotrophic organic matter transfers from the donor compartment to the consumer. Dotted arrows represent dissolved nutrients transfer. Planktonic organisms are depicted based on their size range.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Main limnological characteristics and average ± standard deviation of some variables of representative lakes from Byers Peninsula. Average values represent data compiled from surveys conducted in these lakes during different austral summers, from 2001/2002 to 2008/2009. The Universal Transverse Mercator (UTM) coordinates format is the World Geodetic System 1984 (WGS-84), from the International Map of the World (IMW) sheet numbering SP20.
Table 1.
Main limnological characteristics and average ± standard deviation of some variables of representative lakes from Byers Peninsula. Average values represent data compiled from surveys conducted in these lakes during different austral summers, from 2001/2002 to 2008/2009. The Universal Transverse Mercator (UTM) coordinates format is the World Geodetic System 1984 (WGS-84), from the International Map of the World (IMW) sheet numbering SP20.
| Lake | Chester Cone | Midge Lake | Limnopolar | Escondido | Turbio | Somero | Refugio |
|---|---|---|---|---|---|---|---|
| X-UTM | 597,500 | 597,700 | 597,100 | 599,475 | 598,000 | 596,800 | 602,200 |
| Y-UTM | 3,053,550 | 3,054,150 | 3,052,200 | 3,052,650 | 3,051,800 | 3,052,150 | 3,050,550 |
| Maximum depth (m) | 5.0 | 8.2 | 5.5 | 4.5 | 7.8 | 0.5 | 0.5 |
| Lake Surface (Km2) | 0.039 | 0.054 | 0.023 | 0.022 | 0.021 | 0.011 | 0.016 |
| Catchment size (Km2) | 0.09 | 0.27 | 0.58 | 0.08 | 0.58 | 0.06 | 0.12 |
| Conductivity (µS/cm) | 58 ± 6 | 68 ± 12 | 65 ± 9 | 60 ± 7 | 60 ± 17 | 81 ± 14 | 148 ± 31 |
| Total phosphorus (µM) | 0.28 ± 0.21 | 0.24 ± 0.17 | 0.39 ± 0.18 | 0.28 ± 0.04 | 1.27 ± 0.92 | 1.02 ± 0.49 | 16.84 ± 6.16 |
| Bacterioplankton (cells/mL) × 106 | 0.99 ± 0.39 | 0.98 ± 0.36 | 1.39 ± 0.86 | 1.50 ± 0.41 | 2.26 ± 2.19 | 3.27 ± 1.61 | 8.20 ± 2.68 |
| Chlorophyll a (µg/L) | 0.08 ± 0.01 | 0.14 ± 0.06 | 0.10 ± 0.07 | 0.38 ± 0.17 | 0.37 ± 0.19 | 0.94 ± 0.66 | 23.41 ± 9.98 |
Table 2.
Dominant taxa, per groups, in the water column and benthos of Lake Limnopolar during the ice- free summer periods. Data compiled from [18,31,33,42,43,44].
| Group | Dominant Taxa | |
|---|---|---|
| DNA Virioplankton | Phycodnaviridae, Caudovirales, Mimiviridae | |
| RNA Virioplankton | Dicistroviridae, Secoviridae, Marnaviridae, Iflaviridae, Potyviridae, Picornaviridae and Tombusviridae | |
| Heterotrophic bacterioplankton (Families) | Burkholderiaceae, Sporichthyaceae, Flavobacteriaceae, Chromatiaceae, Chitinophagaceae, Xanthomonadaceae, Sphingomonadaceae | |
| Autotrophic picoplankton | Picocyanobacteria, Prasinophytes | |
| Plastidic nanoflagellates (genera) | Ochromonas, Chromulina, Pseudokephyrion. | |
| Heterotrophic nanoflagellates (genera) | Spumella, Oikomonas | |
| Microphytoplankton (genera) | Pennate diatoms | Navicula, Nitzschia, Achnanthes, Fragilaria |
| Chlorophytes | Ankistrodesmus, Ankyra, Chlamydomonas | |
| Cryptophytes | Chroomonas | |
| Nanoplanktonic ciliates | Balanion planctonicum, Cyclidium glaucoma | |
| Rotifers (genera) | Notholca | |
| Copepods | Boeckella poppei | |
| Cladocerans | Macrothrix ciliata | |
| Anostraca | Branchinecta gainii | |
| Chironomids | Parochlus steinenii, Belgica antarctica | |
| Oligochaetes | Lumbricillus healyae | |
| Bryophytes | Drepanocladus longifolius | |
Table 3.
¡ Dominant taxa, per groups, observed in microbial mats from of Peninsula Byers Peninsula. Data compiled from [19,65,66,67,68,69,73,74,75].
| Group | Dominant Taxa | |
|---|---|---|
| RNA Viruses | Bacillariornavirus, Picornavirales | |
| DNA Viruses | Circoviridae, Nanoviridae, Microviridae, Geminiviridae | |
| Bacteria | Alphaproteobacteria, Bacteroidetes, Cloroflexi, Actinobacteria | |
| Fungi | Ascomycota, Basidiomycta, Chytridiomycota | |
| Phototrophs | Cyanobacteria | Leptolyngbya, Oscillatoria, Phormidium, Porphyrosiphon, Nostoc |
| Diatoms | Fragilaria, Nitzschia, Gomphonema, Planothidium, Navicula | |
| Chlorophytes | Ulothrix, Klebsormidium, Chlamydomonas, Chloromonas | |
| Chrysophytes | Ochromonas | |
| Nematodes | Plectidae, Monhysteridae | |
| Tardigrades | Hypsibiidae, Macrobioitidae, Calohypsibiidae | |
| Rotifers | Philodinidae, Lecanidae, Lindiidae | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rochera, C.; Camacho, A. Limnology and Aquatic Microbial Ecology of Byers Peninsula: A Main Freshwater Biodiversity Hotspot in Maritime Antarctica. Diversity 2019, 11, 201. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100201
AMA Style
Rochera C, Camacho A. Limnology and Aquatic Microbial Ecology of Byers Peninsula: A Main Freshwater Biodiversity Hotspot in Maritime Antarctica. Diversity. 2019; 11(10):201. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100201
Chicago/Turabian StyleRochera, Carlos, and Antonio Camacho. 2019. "Limnology and Aquatic Microbial Ecology of Byers Peninsula: A Main Freshwater Biodiversity Hotspot in Maritime Antarctica" Diversity 11, no. 10: 201. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100201
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.