Analysis of Encystment, Excystment, and Cyst Structure in Freshwater Eutardigrade Thulinius ruffoi (Tardigrada, Isohypsibioidea: Doryphoribiidae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Methods
3. Results
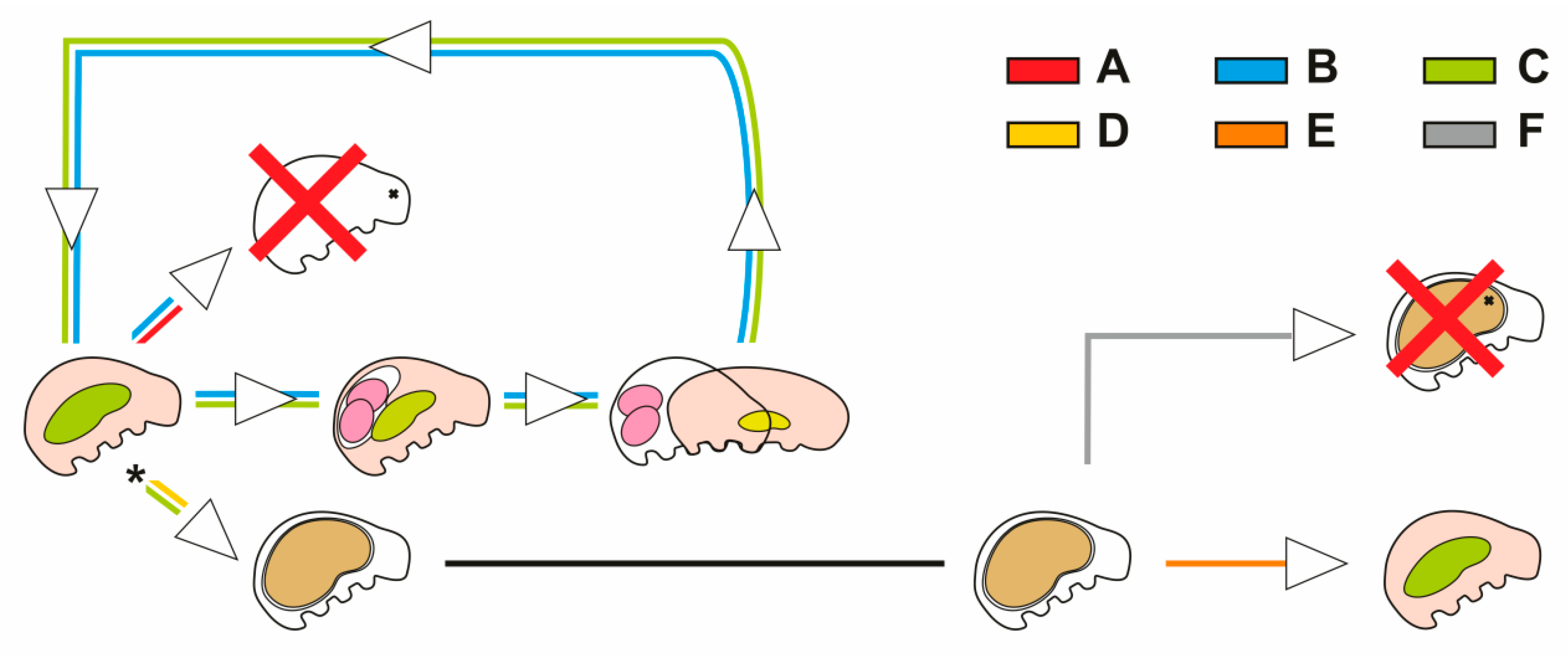
3.1. Encystment and Excystment
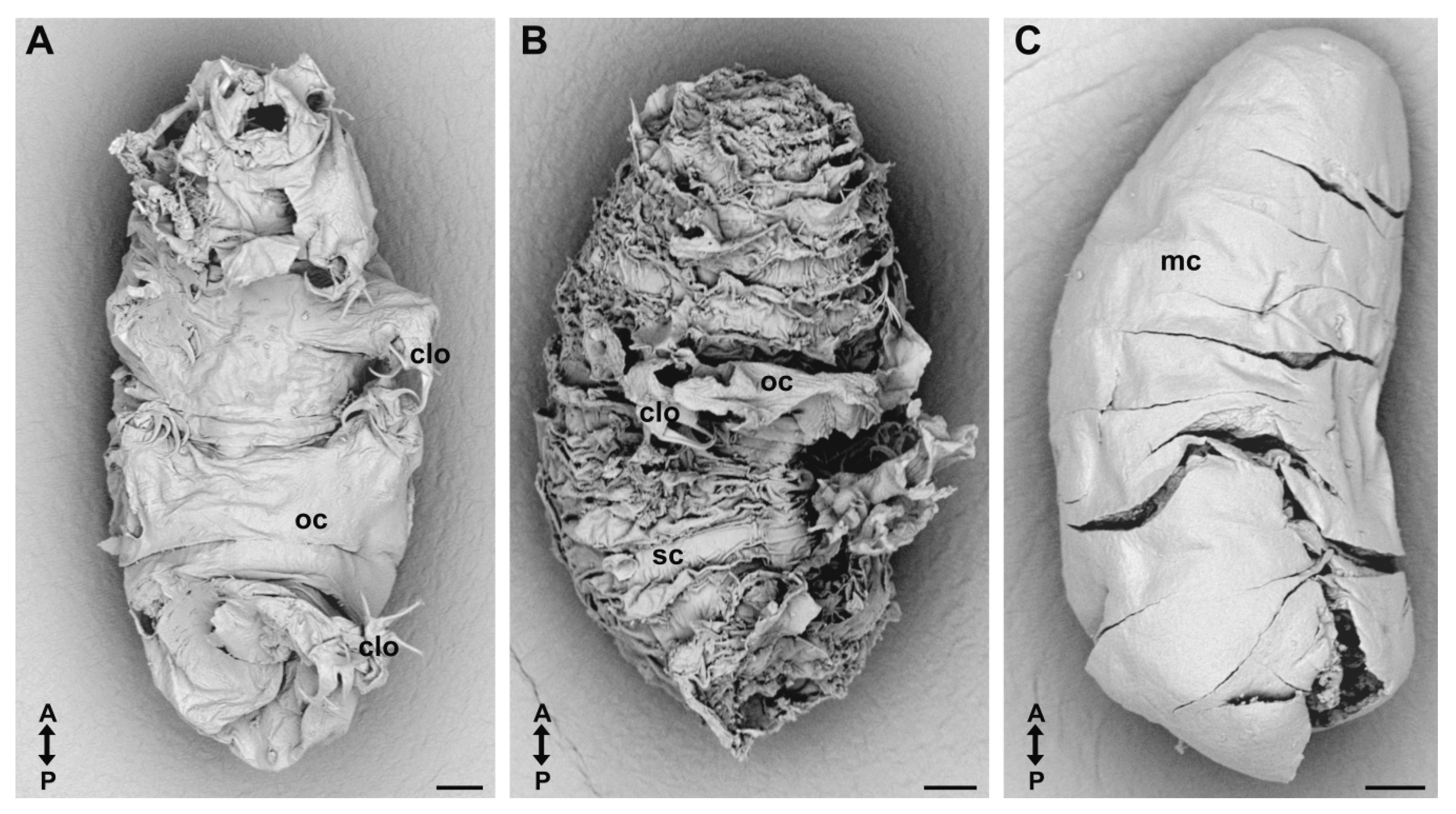
3.2. Cuticular Capsule
3.3. Buccal-Pharyngeal Apparatus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Padilla, P.A.; Ladage, M.L. Suspended animation, diapause and quiescence. Cell Cycle 2012, 11, 1672–1679. [Google Scholar] [CrossRef] [Green Version]
- Gorr, T.A. Hypometabolism as the ultimate defence in stress response: how the comparative approach helps understanding of medically relevant questions. Acta Physiol. 2017, 219, 409–440. [Google Scholar] [CrossRef] [PubMed]
- Guidetti, R.; Boschini, D.; Altiero, T.; Bertolani, R.; Rebecchi, L. Diapause in tardigrades: a study of factors involved in encystment. J. Exp. Biol. 2008, 211, 2296–2302. [Google Scholar] [CrossRef] [Green Version]
- Møbjerg, N.; Halberg, K.A.; Jørgensen, A.; Persson, D.; Bjørn, M.; Ramløv, H.; Kristensen, R.M. Survival in extreme environments — on the current knowledge of adaptations in tardigrades. Acta Physiol. 2011, 202, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Guidetti, R.; Rebecchi, L. Chapter 17 — Phylum Tardigrada. In Thorp and Covich’s Freshwater Invertebrates, 4th ed.; Thorp, J.H., Rogers, D.C., Eds.; Elsevier: London, UK, 2015; Volume 1, pp. 347–380. [Google Scholar]
- Guidetti, R.; Altiero, T.; Rebecchi, L. On dormancy strategies in tardigrades. J. Insect Physiol. 2011, 57, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Grohme, M.; Frohme, M.; Schnölzer, M.; Dandekar, T.; Reuter, D.; Schill, R.O. New insights into anhydrobiotic organisms: Perspectives for preservation and stabilization of biological material. Comp. Biochem. Physiol. Part A 2008, 151, 31–36. [Google Scholar] [CrossRef]
- Jönsson, K.I.; Rabbow, E.; Schill, R.O.; Harms-Ringdahl, M.; Rettberg, P. Tardigrades survive exposure to space in low Earth orbit. Curr. Biol. 2008, 18, R729–R731. [Google Scholar] [CrossRef] [Green Version]
- Schill, R.O.; Mali, B.; Dandekar, T.; Schnölzer, M.; Reuter, D.; Frohme, M. Molecular mechanisms of tolerance in tardigrades: New perspectives for preservation and stabilization of biological material. Biotechnol. Adv. 2009, 27, 348–352. [Google Scholar] [CrossRef]
- Varghese, A.C.; Nagy, Z.P.; Agarwal, A. Current trends, biological foundations and future prospects of oocyte and embryo cryopreservation. Reprod. Biomed. Online 2009, 19, 126–140. [Google Scholar] [CrossRef]
- Sloan, D.; Batista, R.A.; Loeb, A. The resilience of life to astrophysical events. Sci. Rep. 2017, 7, 5419. [Google Scholar] [CrossRef]
- Guidetti, R.; Møbjerg, N. Environmental Adaptations: Encystment and Cyclomorphosis. In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Zoological Monographs; Springer: Cham, Switzerland, 2018; Volume 2, pp. 249–271. [Google Scholar]
- Bertolani, R.; Guidetti, R.; Jönsson, K.I.; Altiero, T.; Boschini, D.; Rebecchi, L. Experiences with dormancy in tardigrades. J. Limnol. 2004, 63, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Guidetti, R.; Boschini, D.; Rebecchi, L.; Bertolani, R. Encystment processes and the “Matrioshka-like stage” in a moss-dwelling and in a limnic species of eutardigrades (Tardigrada). Hydrobiologia 2006, 558, 9–21. [Google Scholar] [CrossRef]
- Clausen, L.K.B.; Andersen, K.N.; Hygum, T.L.; Jørgensen, A.; Møbjerg, N. First record of cysts in the tidal tardigrade Echiniscoides sigismundi. Helgol. Mar. Res. 2014, 68, 531–537. [Google Scholar] [CrossRef] [Green Version]
- von Wenck, W. Entwicklungs geschichtliche Untersuchungen an Tardigraden (Macrobiotus lacustris Duj.). Zool. Jahrb. Abt. Für Anat. Ontog. Tiere 1914, 37, 465–514. [Google Scholar]
- Marcus, E. Spinnentiere oder Arachoides. IV. Bärtichen (Tardigrada). In Die Tierreich Deutschlands und der angrenzenden Meeresteile; Dahl, F., Ed.; Gustav Fischer: Jena, Germany, 1928; Volume 12, pp. 1–230. [Google Scholar]
- Marcus, E. Tardigrada. In HG Bronn’s Klassen und Ordnungen des Tierreichs; Bronn, H.G., Ed.; Akademische Verlagsgesellschaft: Leipzig, Germany, 1929; Volume 5, pp. 1–608. [Google Scholar]
- Ramazzotti, G. Tardigradi in terreni prativi. Atti Soc. Ital. Sci. Nat. Mus. Civ. Stor. Nat. Milano 1959, 98, 199–210. [Google Scholar]
- Manicardi, G.C. Two new species of soil moss eutardigrades (Tardigrada) from Canada. Can. J. Zool. 1989, 67, 2282–2285. [Google Scholar] [CrossRef]
- Biserov, V.I. A new genus and three new species of tardigrades (Tardigrada: Eutardigrada) from the USSR. Boll. Zool. 1992, 59, 95–103. [Google Scholar] [CrossRef] [Green Version]
- McInnes, S.J.; Pugh, P.J. Zonation in Antarctic lake-dwelling benthic meiofauna, with emphasis on the Tardigrada. Zool. Anz. 1999, 238, 283–288. [Google Scholar]
- Stark, C.; Kristensen, R.M. Tardigrades in the soil of Greenland. Ber. Polarforsch. 1999, 330, 44–63. [Google Scholar]
- Schill, R.O.; Huhn, F.; Köhler, H.-R. The first record of tardigrades (Tardigrada) from the Sinai Peninsula, Egypt. Zool. Middle East 2007, 42, 83–88. [Google Scholar] [CrossRef]
- Perry, E.S.; Miller, W.R.; Lindsay, S. Looking at tardigrades in a new light: using epifluorescence to interpret structure. J. Microsc. 2015, 257, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Morek, W.; Stec, D.; Gąsiorek, P.; Schill, R.O.; Kaczmarek, Ł.; Michalczyk, Ł. An experimental test of eutardigrade preparation methods for light microscopy. Zool. J. Linn. Soc. 2016, 178, 785–793. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.J. Dormancy in Rotifers. Trans. Am. Microsc. Soc. 1974, 93, 490–513. [Google Scholar] [CrossRef]
- Ricci, C. Dormancy patterns in rotifers. Hydrobiologia 2001, 446, 1–11. [Google Scholar] [CrossRef]
- Fielding, M.J. Observations on the length of dormancy in certain plant infecting nematodes. Proc. Helminthol. Soc. Wash. 1951, 18, 110–112. [Google Scholar]
- Hengherr, S.; Schill, R.O. Dormant stages in freshwater bryozoans — An adaptation to transcend environmental constraints. J. Insect Physiol. 2011, 57, 595–601. [Google Scholar] [CrossRef]
- Danks, H.V. Insect Dormancy: An Ecological Perspective; Biological Survey of Canada (Terrestrial Artropods): Ottawa, QC, Canada, 1987. [Google Scholar]
- Dahms, H.-U. Dormancy in the Copepoda—An overview. Hydrobiologia 1995, 306, 199–211. [Google Scholar] [CrossRef]
- Lauterborn, R. Demonstrationen aus der Fauna des Oberrheins und seiner Umgebung. Verhandlungen Dtsch. Zool. Ges. 1906, 16, 265–268. [Google Scholar]
- Murray, J. The encystment of Macrobiotus. Zoologist 1907, 11, 4–11. [Google Scholar]
- Murray, J. Some Tardigrada of the Sikkim Himalaya. J. R. Microsc. Soc. 1907, 27, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Murray, J. Scottish Tardigrada, collected by the Lake Survey. Trans. R. Soc. Edinb. 1907, 45, 641–668. [Google Scholar] [CrossRef] [Green Version]
- Murray, J. Encystment of Tardigrada. Trans. R. Soc. Edinb. 1907, 45, 837–854. [Google Scholar] [CrossRef] [Green Version]
- Murray, J. Tardigrada. In British Antarctic Expedition, 1907–1909: Under the Command of Sir E.H. Shackleton, C.V.O.: Reports on the Scientific Investigations: Biology; Murray, J., Ed.; William Heinemann: London, UK, 1910; Volume 1, pp. 81–185. [Google Scholar]
- Heinis, F. Systematik und Biologie der moosbewohnenden Rhizopoden, Rotatorien und Tardigradender Umgebung von Basel mit Berücksichtigung der übrigen Schweiz. Arch. Hydrobiol. Planktonkd. 1910, 5, 40–115. [Google Scholar]
- Węglarska, B. Hypsibius (Isohypsibius) smreczynskii spec. nov., a new species of freshwater tardigrade. Zesz. Nauk. Uniw. Jagiellonsk. Pr. Zool. 1970, 16, 107–114. [Google Scholar]
- Westh, P.; Kristensen, R.M. Ice formation in the freeze-tolerant eutardigrades Adorybiotus coronifer and Amphibolus nebulosus studied by differential scanning calorimetry. Polar Biol. 1992, 12, 693–699. [Google Scholar] [CrossRef]
- Rebecchi, L.; Bertolani, R. Maturative pattern of ovary and testis in eutardigrades of freshwater and terrestrial habitats. Invertebr. Reprod. Dev. 1994, 26, 107–117. [Google Scholar] [CrossRef]
- Hansen, J.G.; Kristensen, R.M.; Bertolani, R.; Guidetti, R. Comparative analyses of Bertolanius species (Eohypsibiidae; Eutardigrada) with the description of Bertolanius birnae sp. nov. from northern polar regions. Polar Biol. 2017, 40, 123–140. [Google Scholar] [CrossRef]
- Węglarska, B. On the encystation in Tardigrada. Zool. Pol. 1957, 8, 315–325. [Google Scholar]
- Szymanska, B. Encystment in the Tardigrade Dactylobiotus dispar (Murray, 1907)(Tardigrada: Eutardigrada). Part I. Observation of living animals and structure of cyst. Zool. Pol. 1995, 40, 91–102. [Google Scholar]
- Rost-Roszkowska, M.M.; Poprawa, I. Ultrastructure of the midgut epithelium in Dactylobiotus dispar (Tardigrada: Eutardigrada) during encystation. Zool. Pol. 2008, 53, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J.; Katholm, A.K. A study of the genus Amphibolus from Disko Island with special attention on the life cycle of Amphibolus nebulosus (Eutardigrada: Eohypsibiidae). In Arctic Biology Field Course, Quqertarsuaq, 2002; Hansen, J.G., Ed.; Zoological Museum University of Copenhagen: Copenhagen, Denmark, 2002; pp. 129–163. [Google Scholar]
- Pigoń, A.; Węglarska, B. The respiration of Tardigrada: a study in animal anabiosis. Bull. Acad. Polon. Sci. 1953, 1, 69–72. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janelt, K.; Poprawa, I. Analysis of Encystment, Excystment, and Cyst Structure in Freshwater Eutardigrade Thulinius ruffoi (Tardigrada, Isohypsibioidea: Doryphoribiidae). Diversity 2020, 12, 62. https://0-doi-org.brum.beds.ac.uk/10.3390/d12020062
Janelt K, Poprawa I. Analysis of Encystment, Excystment, and Cyst Structure in Freshwater Eutardigrade Thulinius ruffoi (Tardigrada, Isohypsibioidea: Doryphoribiidae). Diversity. 2020; 12(2):62. https://0-doi-org.brum.beds.ac.uk/10.3390/d12020062
Chicago/Turabian StyleJanelt, Kamil, and Izabela Poprawa. 2020. "Analysis of Encystment, Excystment, and Cyst Structure in Freshwater Eutardigrade Thulinius ruffoi (Tardigrada, Isohypsibioidea: Doryphoribiidae)" Diversity 12, no. 2: 62. https://0-doi-org.brum.beds.ac.uk/10.3390/d12020062