The Deep-Sea Genus Coronarctus (Tardigrada, Arthrotardigrada) in Brazil, South-Western Atlantic Ocean, with the Description of Three New Species

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Coronarctus dissimilis sp. nov.
3.1.1. Diagnosis
3.1.2. Type Locality
3.1.3. Type Material and Type Repository
3.1.4. Etymology
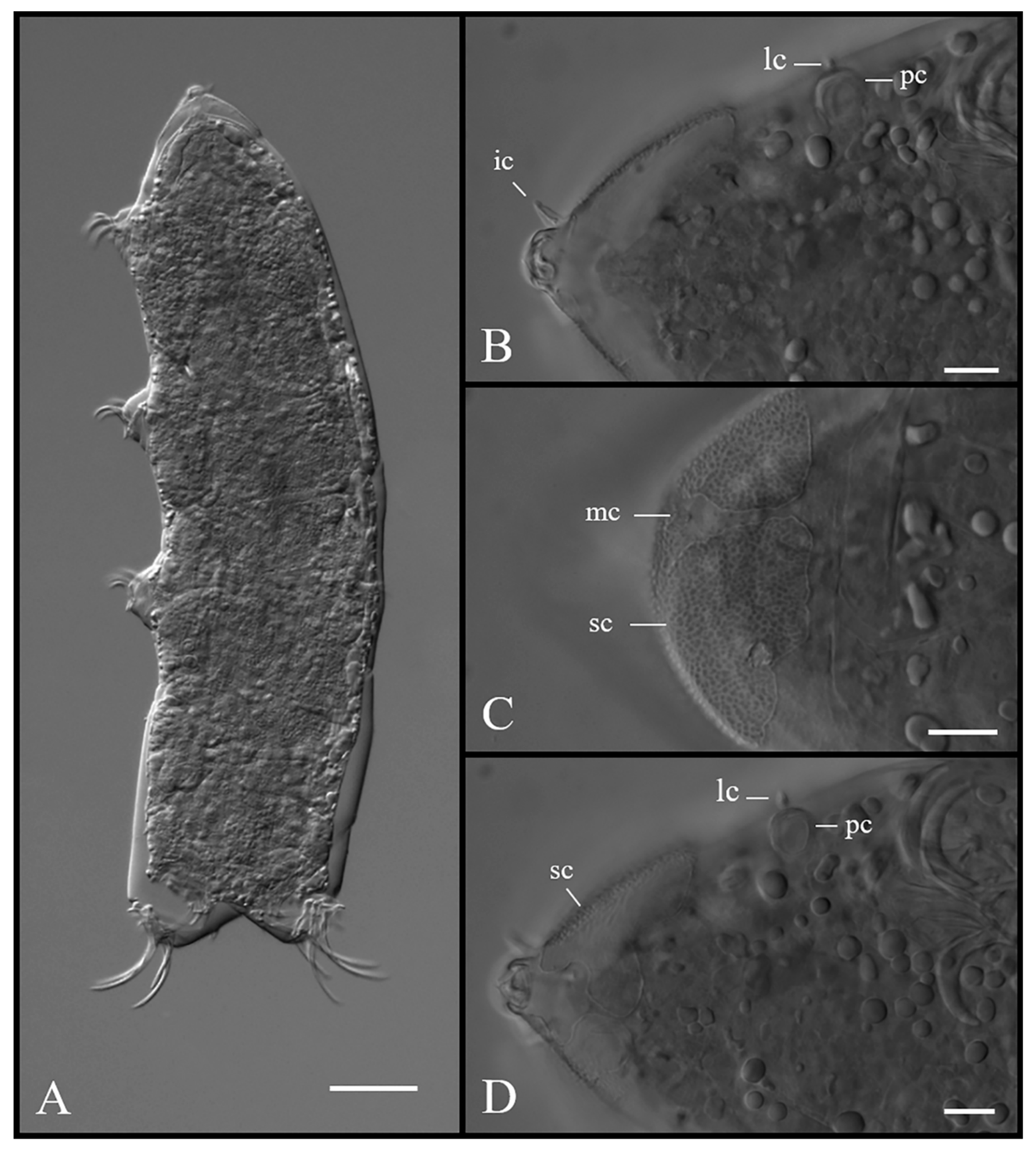
3.1.5. Description of Holotype
3.1.6. Remarks
3.1.7. Differential Diagnosis
3.2. Coronarctus laubieri Renaud-Mornant, 1987
3.2.1. Material Examined
3.2.2. Remarks
3.3. Coronarctus neptunus sp. nov.
3.3.1. Diagnosis
3.3.2. Type Material and Type Repository
3.3.3. Type Locality
3.3.4. Etymology
3.3.5. Description of Holotype
3.3.6. Remarks
3.3.7. Differential Diagnosis
3.4. Coronarctus yurupari sp. nov.
3.4.1. Diagnosis
3.4.2. Type Locality:
3.4.3. Type Material and Type Repository
3.4.4. Etymology
3.4.5. Description of the Holotype
3.4.6. Remarks
3.4.7. Differential Diagnosis
- -
- From C. tenellus, since C. yurupari has coxal spine-like processes on legs IV (absent in C. tenellus) and apical accessory spines on internal and external claws on all legs and additional mediodorsal spine on claws of legs IV (apical accessory spines on external claws and the additional spine are absent in C. tenellus);
- -
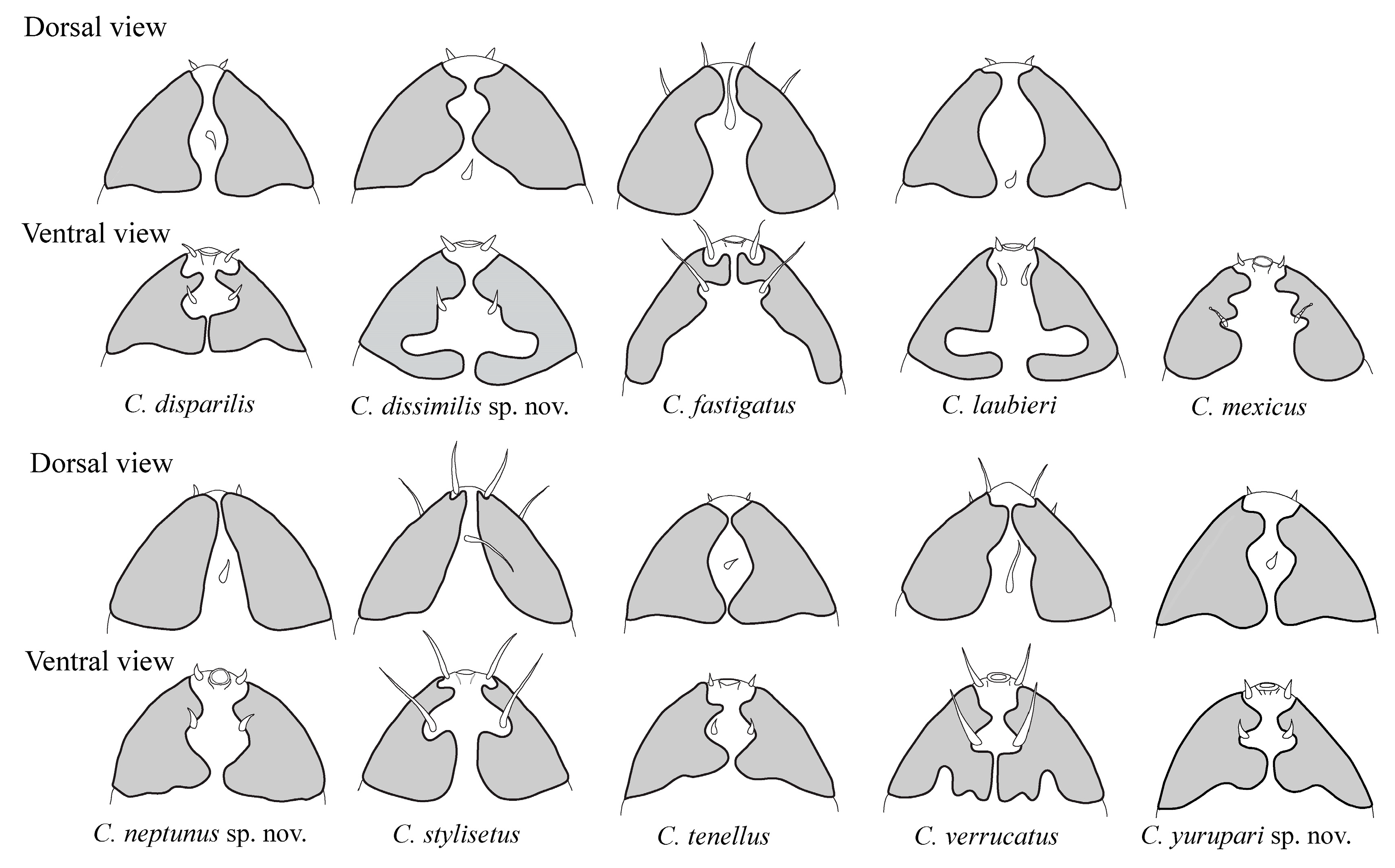
- From C. disparilis, since C. yurupari has the additional spine on legs IV located mediodorsally, while in C. disparilis it is located on the ventral side of the claws. Apical accessory spines are present on all claws of legs I–III in the new species while lacking on external claws in C. disparilis. Moreover, the primary clavae are pedunculated in C. yurupari sp. nov. and subspherical in C. disparilis. The two species also differ in the shape of internal edges of the posterior lobes of secondary clavae that are circular in the new species and squarish in C. disparilis (Figure 5).
4. Discussion
A Dichotomous Key for the Identification of Coronarctus Species
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kristensen, R.M.; Hansen, J.G. Tardigrada (water bears). In Marine Parasitology; Rhode, K., Ed.; CSIRO publishing: Clayton, Australia, 2005; pp. 230–235. [Google Scholar]
- Fontoura, P.; Bartels, P.J.; Jørgensen, A.; Kristensen, R.M.; Hansen, J.G. A dichotomous key to the genera of the Marine Heterotardigrades (Tardigrada). Zootaxa 2017, 4294, 1–45. [Google Scholar] [CrossRef]
- Nelson, D.R.; Bartels, P.J.; Guil, N. Tardigrade Ecology. In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Zoological Monographs 2; Springer Nature Switzerland AG: Cham, Switzerland, 2018; Chapter 7; pp. 163–210. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Bartels, P.J.; Roszkowska, M.; Nelson, D.R. The Zoogeography of Marine Tardigrada. Zootaxa 2015, 4037, 1–189. [Google Scholar] [CrossRef] [Green Version]
- Degma, P.; Guidetti, R. Tardigrade Taxa. In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Zoological Monographs 2; Springer Nature Switzerland AG: Cham, Switzerland, 2018; Chapter 15; pp. 371–409. [Google Scholar] [CrossRef]
- Actual checklist of Tardigrada species (36th Edition: 1-09-2019). 2009–2019. Available online: https://iris.unimore.it/handle/11380/1178608#.XlXnUHERXIU (accessed on 12 September 2019).
- Bartels, P.J.; Apodaca, J.J.; Mora, C.; Nelson, D.R. A global biodiversity estimate of a poorly known taxon: Phylum Tardigrada. Zool. J. Linn. Soc. 2016, 178, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Thiel, H. Quantitative Untersuchungen über die Meiofauna des Tiefseebodens. Veröffentlichungen des Instituts für Meeresforschung in Bremerhaven 1966, 2, 131–148. [Google Scholar]
- Kristensen, R.M.; Sørensen, M.V.; Hansen, J.G.; Zeppilli, D. A new species of Neostygarctus (Arthrotardigrada) from the Condor Seamount in the Azores, Northeast Atlantic. Mar. Biodivers. 2015, 45, 453–467. [Google Scholar] [CrossRef]
- Tchesunov, A.V. A new tardigrade species of the genus Neostygarctus Grimaldi de Zio et al., 1982 (Tardigrada, Arthrotardigrada) from the Great Meteor Seamount, Northeast Atlantic. Eur. J. Taxon. 2018, 479, 1–17. [Google Scholar] [CrossRef]
- Fujimoto, S.; Hansen, J.G. Revision of Angursa (Arthrotardigrada: Styraconyxidae) with the description of a new species from Japan. Eur. J. Taxon. 2019, 510, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Renaud-Mornant, J. Deux nouveaux Angursa Pollock, 1979, du domaine abyssal (Tardigrada, Arthrotardigrada). Tethys 1981, 10, 161–164. [Google Scholar]
- Da Rocha, C.M.C.; Santos, E.C.L.; Gomes, E.L., Jr.; Moura, J.R.; Silva, L.G.S.; Barbosa, D.F. New records of marine tardigrades from Brazil. J. Limnol. 2013, 72, 102–107. [Google Scholar] [CrossRef]
- Menechella, A.G.; Bulnes, V.N.; Cazzaniga, N.J. Two new species of Batillipes (Tardigrada, Arthrotardigrada, Batillipedidae) from the Argentinean Atlantic coast, and key to all known species. Mar. Biodivers. 2018, 48, 239–247. [Google Scholar] [CrossRef]
- Santos, E.; da Rocha, C.M.C.; Gomes, E., Jr.; Fontoura, P. Three new Batillipes species (Arthrotardigrada: Batillipedidae) from the Brazilian coast. Zootaxa 2017, 4243, 483–502. [Google Scholar] [CrossRef] [PubMed]
- Gomes, E., Jr.; Santos, E.; da Rocha, C.M.C.; Santos, P.J.P.; Fontoura, P. A new species of Ligiarctus (Tardigrada, Heterotardigrada) from the Brazilian continental shelf, Southwestern Atlantic Ocean. Mar. Biodivers. 2018, 48, 5–12. [Google Scholar] [CrossRef]
- Zeppilli, D.; Sarrazin, J.; Leduc, D.; Martinéz-Arbizu, P.; Fontaneto, D.; Fontanier, C.; Gooday, A.J.; Kristensen, R.M.; Ivanenko, V.N.; Sørensen, M.V.; et al. Is the meiofauna a good indicator for climate change and anthropogenic impacts? Mar. Biodivers. 2015, 45, 505–535. [Google Scholar] [CrossRef] [Green Version]
- Zeppilli, D.; Leduc, D.; Fontanier, C.; Fontaneto, D.; Fuchs, S.; Gooday, A.J.; Goineau, A.; Ingels, J.; Ivanenko, V.N.; Kristensen, R.M.; et al. Characteristics of meiofauna in extreme marine ecosystems: A review. Mar. Biodivers. 2018, 48, 35–71. [Google Scholar] [CrossRef] [Green Version]
- Vanreusel, A.; Hilario, A.; Ribeiro, P.A.; Menot, L.; Arbizu, P.M. Threatened by mining, polymetallic nodules are required to preserve abyssal epifauna. Sci. Rep. 2016, 6, 26808. [Google Scholar] [CrossRef]
- Renaud-Mornant, J. Bathyal and abyssal Coronarctidae (Tardigrada), description of new species and phylogenetical significance. In Biology of Tardigrades. Selected Symposia and Monographs U.Z.I; Bertolani, R., Ed.; 1. Mucchi Editore: Modena, Italy, 1987; pp. 229–252. [Google Scholar]
- Hansen, J.G.; Jørgensen, A.; Kristensen, R.M. Preliminary studies of the tardigrade fauna of the Faroe Bank. Zool. Anz. 2001, 240, 385–393. [Google Scholar] [CrossRef]
- Hansen, J.G. The deep sea elements of the Faroe Bank tardigrade fauna with a description of two new species. J. Limnol. 2007, 66, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Rubal, M.; Fontoura, P.; Veiga, P. Effects of sediment organic enrichment on a high diversity tardigrade assemblage in the Ría de Muros (NW Spain). In Book of Abstracts, Proceedings of the SeventIMCO - Seventeenth International Meiofauna Conference, Évora, Portugal, 7–12 July 2019; Adão, H., Vicente, C., Sroczyńska, K., Espada, M., Alvim, P., Costa, M., Vieira, S., Eds.; Abstract 5999, Special Publication; University of Évora: Évora, Portugal, 2019; ISBN 978-989-8550-97-2. [Google Scholar]
- Romano, F., III; Gallo, M.; D’Addabbo, R.; Accogli, G.; Bagueley, J.; Montagna, P. Deep-sea tardigrades in the northern Gulf of Mexico with a description of a new species of Coronarctidae (Tardigrada: Arthrotardigrada), Coronarctus mexicus. J. Zool. Syst. Evol. Res. 2011, 49, 48–52. [Google Scholar] [CrossRef]
- Noda, H. The first occurrence of coronarctid tardigrade from the shallow water. Zool. Sci. 1989, 6, 1220. [Google Scholar]
- Renaud-Mornant, J. Une nouvelle famille de Tardigrades marins abyssaux: Coronarctidae fam. nov. (Heterotardigrada). C. R. Acad. Sci. 1974, 278, 3087–3090. [Google Scholar]
- Renaud-Mornant, J. Deep Sea Tardigrada from "Meteor" Indian Ocean Expedition. "Meteor" Forsh.-Ergebnisse 1975, 21, 54–61. [Google Scholar]
- Fujimoto, S.; Jørgensen, A.; Hansen, J.G. A molecular approach to arthrotardigrade phylogeny (Heterotardigrada, Tardigrada). Zool Scr. 2017, 46, 496–505. [Google Scholar] [CrossRef]
- Møbjerg, N.; Jørgensen, A.; Kristensen, R.M.; Neves, R.C. Morphology and Functional Anatomy. In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Zoological Monographs 2; Springer Nature Switzerland AG: Cham, Switzerland, 2018; Chapter 2; pp. 57–94. [Google Scholar] [CrossRef]
- Renaud-Mornant, J. Dévelopement postembryonnaire du Tardigrade abyssal Coronarctus laubieri Renaud-Mornant, 1987 (Tardigrada, Heterotardigrada). Bull. Mus. Natl. Hist. Nat. 1988, 10, 327–345. [Google Scholar]
- Faurby, S.; Jørgensen, A.; Kristensen, R.M.; Funch, P. Phylogeography of North Atlantic intertidal tardigrades: Refugia, cryptic speciation and the history of the Mid-Atlantic Islands. J. Biogeogr. 2011, 38, 1613–1624. [Google Scholar] [CrossRef]
- Faurby, S.; Jørgensen, A.; Kristensen, R.M.; Funch, P. Distribution and speciation in marine intertidal tardigrades: Testing the roles of climatic and geographical isolation. J. Biogeogr. 2012, 39, 1596–1607. [Google Scholar] [CrossRef]
- Perry, E.S.; Rawson, P.; Ameral, N.J.; Miller, W.R.; Miller, J.D. Echiniscoides rugostellatus a new marine tardigrade from Washington U.S.A. (Heterotardigrada: Echiniscoidea: Echiniscoididae: Echiniscoidinae). Proc. Biol. Soc. Wash. 2018, 131, 182–193. [Google Scholar] [CrossRef]
- Santos, E.; Veiga, P.; Rubal, M.; Bartels, P.J.; Da Rocha, C.M.C.; Fontoura, P. Batillipes pennaki Marcus, 1946 (Arthrotardigrada: Batillipedidae): Deciphering a species complex. Zootaxa 2019, 4648, 549–567. [Google Scholar] [CrossRef]
- Hansen, J.G.; Kristensen, R.M.; Jørgensen, A. The armoured marine tardigrades (Arthrotardigrada, Tardigrada). Sci. Dan. 2012, 2, 1–91. [Google Scholar]
- Bartels, P.J.; Fontoura, P.; Nelson, D.R. Marine tardigrades of the Bahamas with the description of two new species and updated keys to the species of Anisonyches and Archechiniscus. Zootaxa 2018, 4420, 43–70. [Google Scholar] [CrossRef]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes-Júnior, E.; Santos, É.; da Rocha, C.M.C.; Santos, P.J.P.; Fontoura, P. The Deep-Sea Genus Coronarctus (Tardigrada, Arthrotardigrada) in Brazil, South-Western Atlantic Ocean, with the Description of Three New Species. Diversity 2020, 12, 63. https://0-doi-org.brum.beds.ac.uk/10.3390/d12020063
Gomes-Júnior E, Santos É, da Rocha CMC, Santos PJP, Fontoura P. The Deep-Sea Genus Coronarctus (Tardigrada, Arthrotardigrada) in Brazil, South-Western Atlantic Ocean, with the Description of Three New Species. Diversity. 2020; 12(2):63. https://0-doi-org.brum.beds.ac.uk/10.3390/d12020063
Chicago/Turabian StyleGomes-Júnior, Edivaldo, Érika Santos, Clélia M.C. da Rocha, Paulo J. P. Santos, and Paulo Fontoura. 2020. "The Deep-Sea Genus Coronarctus (Tardigrada, Arthrotardigrada) in Brazil, South-Western Atlantic Ocean, with the Description of Three New Species" Diversity 12, no. 2: 63. https://0-doi-org.brum.beds.ac.uk/10.3390/d12020063