Effect of Predation on Shaping Parental Brood Defense and Larval Ontogeny of Convict Cichlids Leading to Population Divergence

Biosciences Department, Minnesota State University Moorhead, Moorhead, MN 56563, USA
Diversity 2020, 12(4), 136; https://0-doi-org.brum.beds.ac.uk/10.3390/d12040136
Submission received: 7 March 2020
/
Revised: 22 March 2020
/
Accepted: 25 March 2020
/
Published: 2 April 2020
(This article belongs to the Special Issue Predators as Agents of Selection and Diversification)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:(1) Predation selects for antipredator competence in prey. For fishes with parental care, brood predators exert selection on the morphological phenotype of offspring, and also exert strong selection pressure to promote parental care behavior of adults. (2) This review summarizes field and lab studies on the ontogeny of antipredator competence in convict cichlids, a freshwater fish with extended biparental care of their free-swimming young. (3) Here, data show that differences in swimming performance between small and large young are exploited by parents when they adopt (smaller) young. Velocity and acceleration of startle responses improves nonlinearly with body size, increasing rapidly at a point when the skeleton rapidly ossifies from cartilage to bone, at the size at which discrimination by adopting parents shifts, and the timing of change in the rate of change in area protected by parents. Convict cichlids in a Nicaraguan lake population showed a similar correlation among these traits, but these traits are delayed relative to Costa Rican fish. (4) Population divergence is likely explained by relatively more intense brood predation in the lake, which selects for different optima of larval antipredator competence and parental brood defense.
1. Introduction
Predation is a major arbiter of natural selection that exerts its effects on prey evolution by pruning variation in behavioral, ontological and morphological phenotypes of prey [1,2,3]. There are many elegant demonstrations of the effects of predation through comparison of prey populations that occur in sympatry or allopatry with predators [4]. In this paper, I summarize a series of previously published papers on the evolutionary ecology of a freshwater fish species, the convict cichlid Amatitlania siquia, that reveal trait divergences in larval swimming performance and parental care between populations exposed to different levels of brood predation.
Unlike forms of parental care practiced by birds and mammals, parental fish generally do not provision their young with food. Parental care in fishes is primarily in the form of defense of developing embryos and newly hatched young against the threat of predators. Moreover, because adult fish are several orders of magnitude larger than their young (unlike birds and mammals), predators of fish larvae and young juveniles pose no threat to parents. Therefore, fish are ideal organisms for studying the effects of predation on the evolution of parental care. Parental care itself is linked to the ontogeny of antipredator competence of the young. There is a direct link between the vulnerability of young to predation, the duration and intensity of care, and the allocation of resources to reproduction. Fish vary widely in all of these traits and parental care behavior lies at the nexus of these life-history trade-offs.
2. Study System
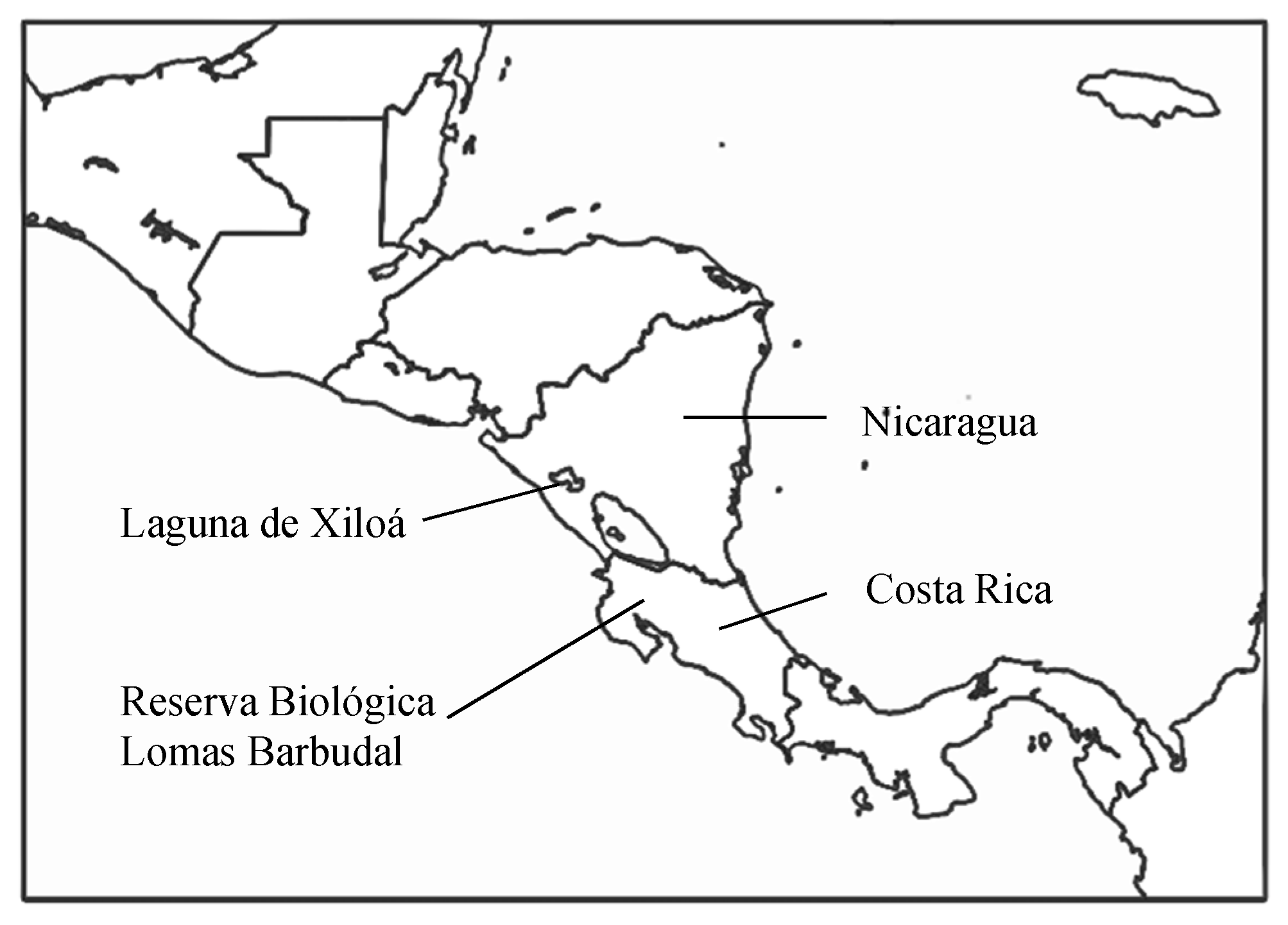
Convict cichlids are biparental freshwater fish endemic to Central America, ranging from Panama to Guatemala [5]. Their taxonomic classification has been revised multiple times in recent years such that the common name “convict cichlid” is more useful than the Latin name. Currently, convict cichlids that occur in Costa Rica and Nicaragua are classified as Amatitlania siquia [6,7]. Most of the data presented here were collected in study populations in clear low-order streams in Lomas Barbudal Biological Reserve, Guanacaste, Costa Rica (Figure 1). A key comparison for the understanding of how predation has shaped life history traits comes from additional data collected in Laguna de Xiloá, a volcanic crater lake in Nicaragua and lab studies based on fish from this population.
3. Mating System of Convict Cichlids in Costa Rican Streams
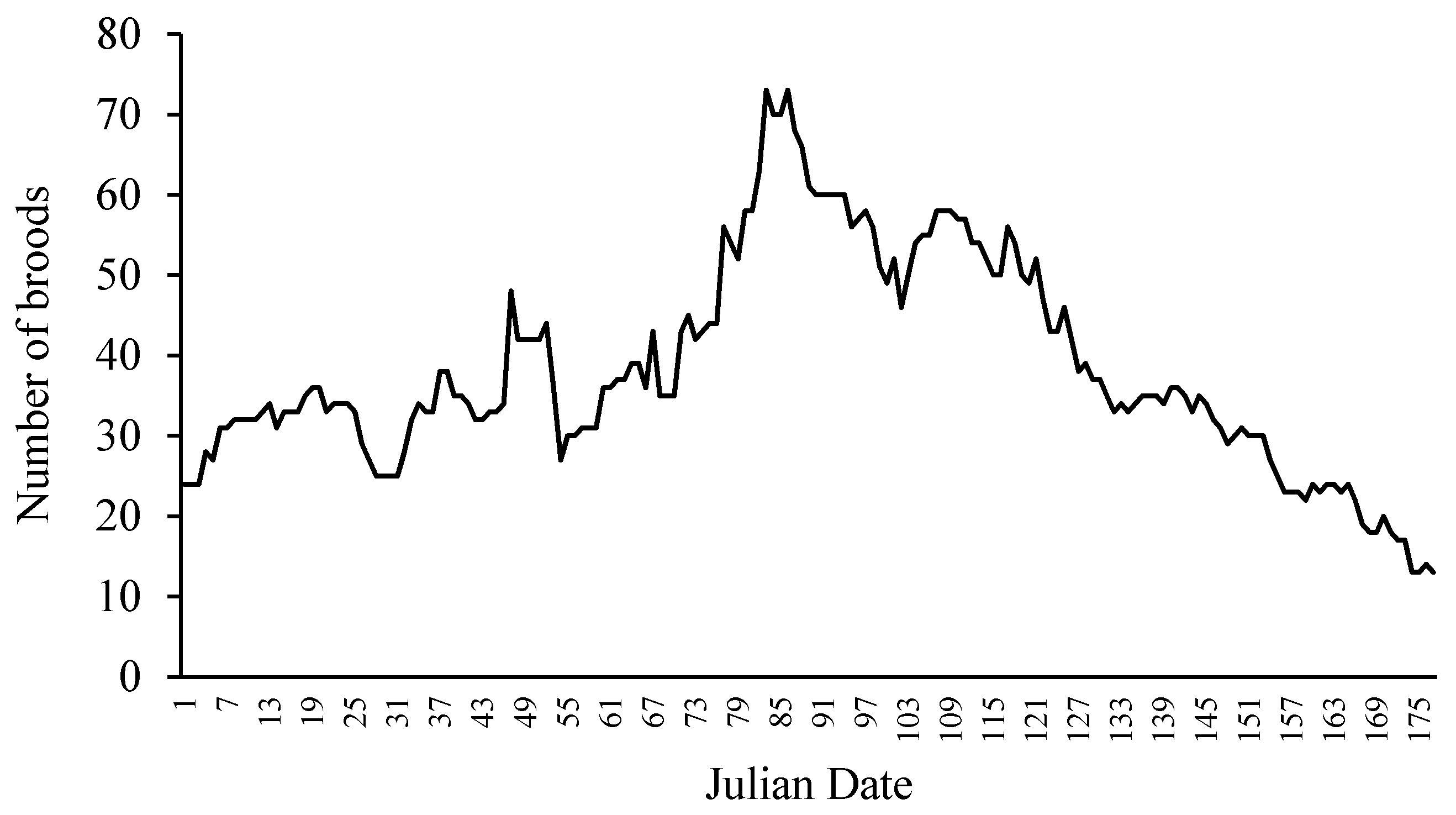
Parental care is tied closely to the mating system. Therefore, a discussion of parental care requires some background information about the mating system. Spawning is seasonal. In Costa Rican streams, spawning activity peaks in the dry season in March–April (Figure 2). The rainy season create spates that scour out sediment and organic debris, and create turbid conditions and violent flows unfavorable for maintaining parent–offspring contact. During the dry season these streams are fed by ground water, flow is gentle and stable, and the water is clear. The dry season is also a time when dry forest trees drop their leaves, which blow into the stream and create a burst of allochthonous production. Convict cichlids engage in leaf-turning behavior to expose the underside of leaves for their own foraging, and when guarding young, they increase leaf turning frequency to provide foraging opportunities for their young [8].
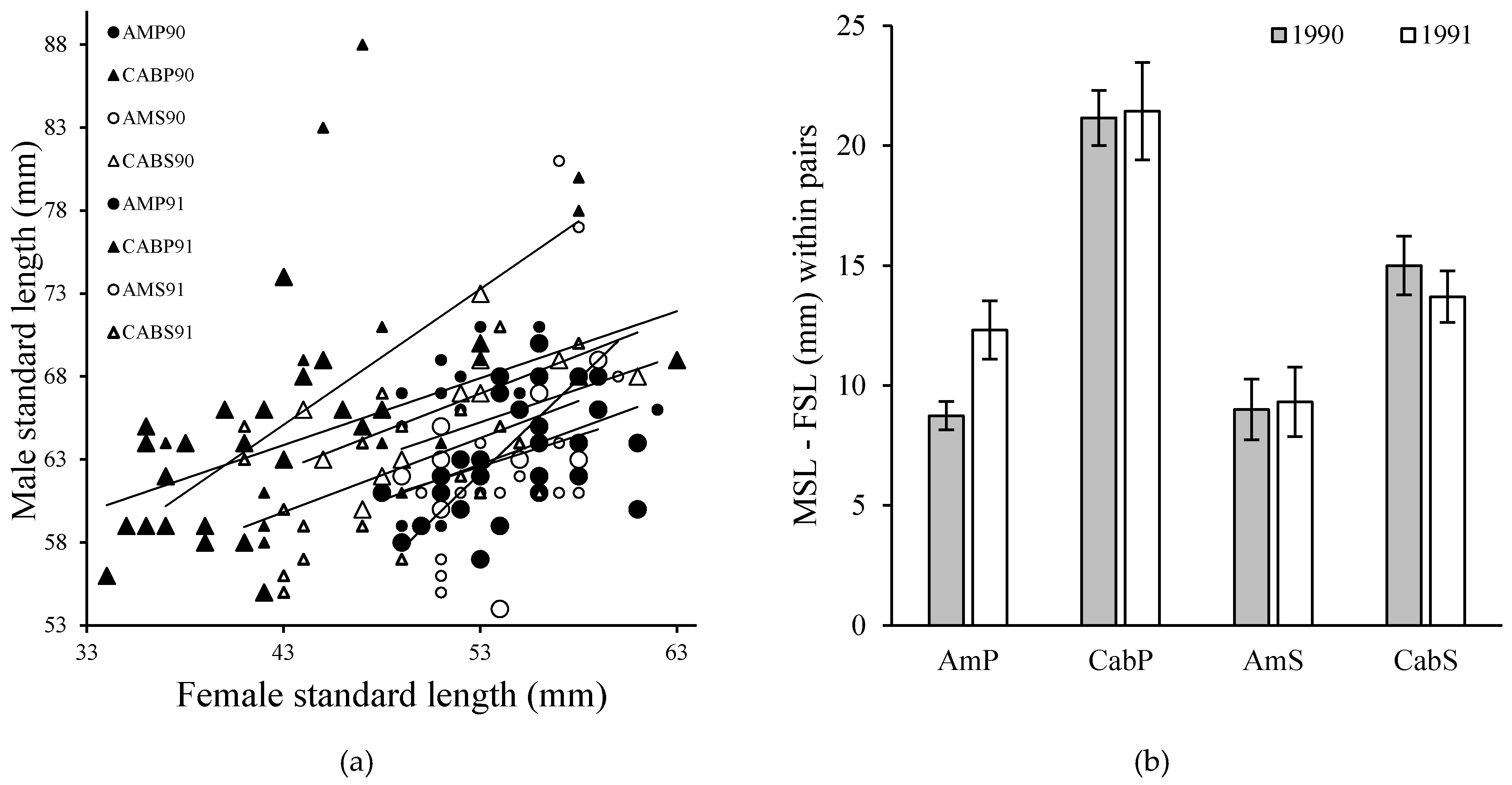
Convict cichlids form monogamous pair bonds and jointly defend a nest cavity from competing conspecific reproductive pairs. Within pairs, males are larger than females and this species shows size-assortative mating (Figure 3). Biparental care is uncommon in fishes. For convict cichlids in this study system, biparental care is more effective than uniparental care in defending against brood predators [9], and allows pairs to defend two territories simultaneously: the mobile territory surrounding the brood, and the fixed territory around the spawning lair. Brooding adults and their offspring return to the lair each night, presumably to take refuge from nocturnal predators such as catfish Rhamdia guatemalensis. During the day, broods roam about the stream bottom foraging on the substrate. About once every 10 min, one parent, usually the male, leaves their partner and young to return to the lair to chase away conspecifics that may be looking to establish themselves there.
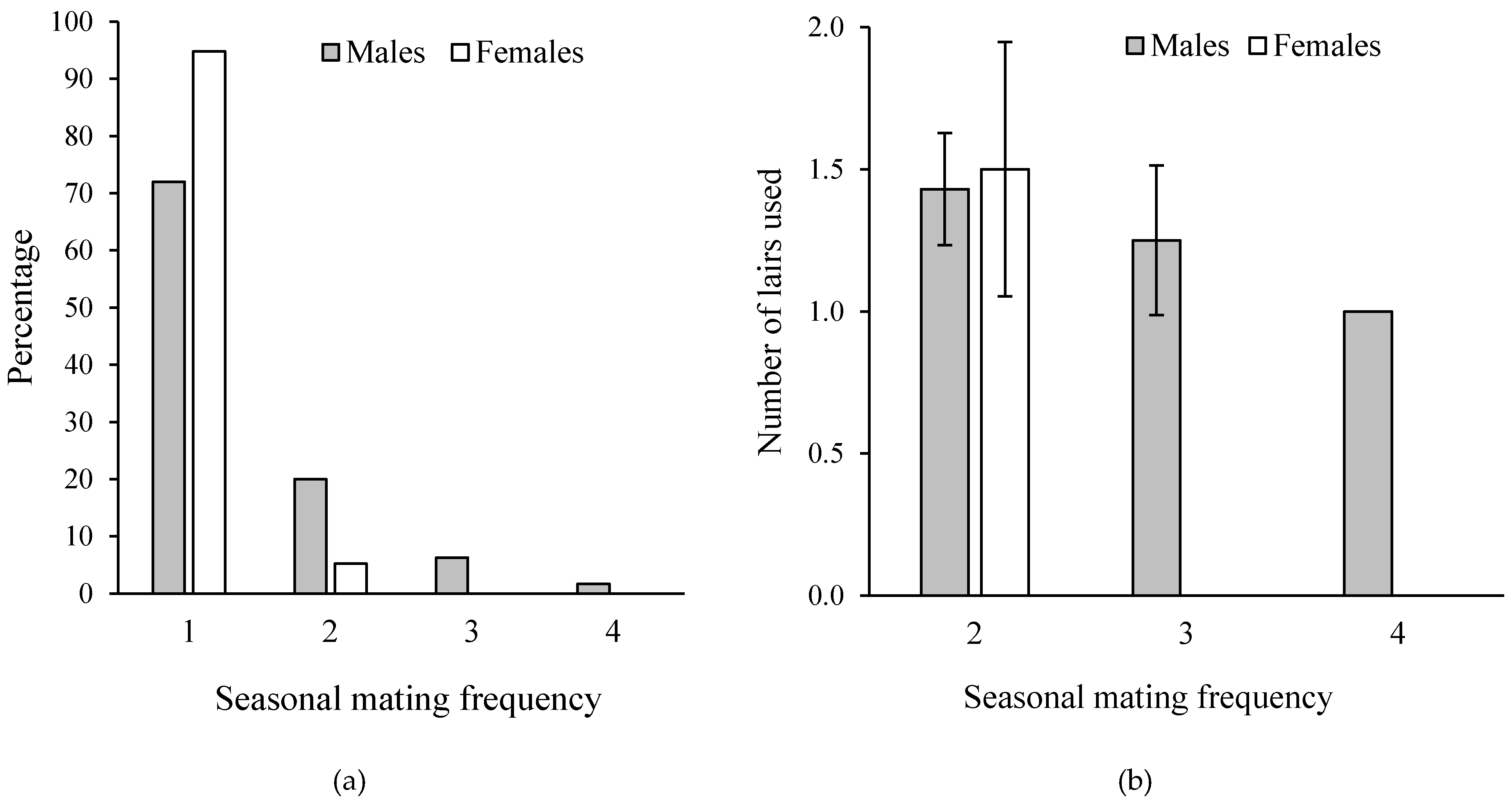
The young emerge from the lair at a size of about 4.5–5.0 mm standard length (SL) and remain associated with parental protection until they are about 10 mm SL, which can take 4–6 weeks depending on the growth rate of fry that varies from site to site [10]. After the young become independent and disperse, the pair bond between the adults dissolves and the male may form a pair bond with a new female and spawn again, often reusing the same lair (Figure 4). Males may spawn up to four times within a 5-month dry season whereas most females spawn only once.
Because only the largest males monopolize spawning lairs and spawn repeatedly in them, the operational sex ratio is skewed, and females compete intrasexually for access to males that possess a lair leading to sexual dichromatism [11]. When not guarding young, males have a dull-colored olive-grey background, often with a dull yellow hue, and dark gray barring. The black and white barring pattern becomes highly contrasting (like the uniforms of prison convicts) when guarding young. Females have three color phases. Similar to males, females are cryptic olive-gray (never with yellow hue) with grey barring when not reproductive, while parental females become highly aposematic with conspicuous bright black and white barring (Figure 5). Gravid females become blackened lending high contrast for gold flecks on the region of their abdomen that swells when gravid with eggs (presumably serving as egg mimics?), yellow pigments in the webbing of the dorsal fin and blue pigment in the gular region [11]. In addition to these color changes, black-phase females actively court parental males that are paired with other females [12] and fight intrasexually for access to males that possess a spawning lair (see video in supplementary materials). Pairs may form before selecting a spawning lair, but often the male already possesses a lair from a previous bout of reproduction.
Eggs are glued to the ceiling of the lair by the female and fertilized by the male. Females do most of the direct care of the embryos while they remain glued to the ceiling of the lair and, in a few days, after they hatch into “free embryos” (eleuthroembryos or “wrigglers”) that form a trembling heap on the floor of the lair. During this time the male patrols the vicinity outside the lair to repel intruders. After a few more days, the wrigglers absorb their yolk and develop their fins and become free-swimming. In Costa Rican streams they emerge from the lair at a size of 4.5–5.0 SL and begin exogenous feeding on the substrate, all the while guarded by both parents. There is genetic [13] and morphological [14] evidence that convict cichlids are socially monogamous but not necessarily genetically monogamous. How the genetic structure of families may affect parental care has not been explored.
4. Parental Brood Defense
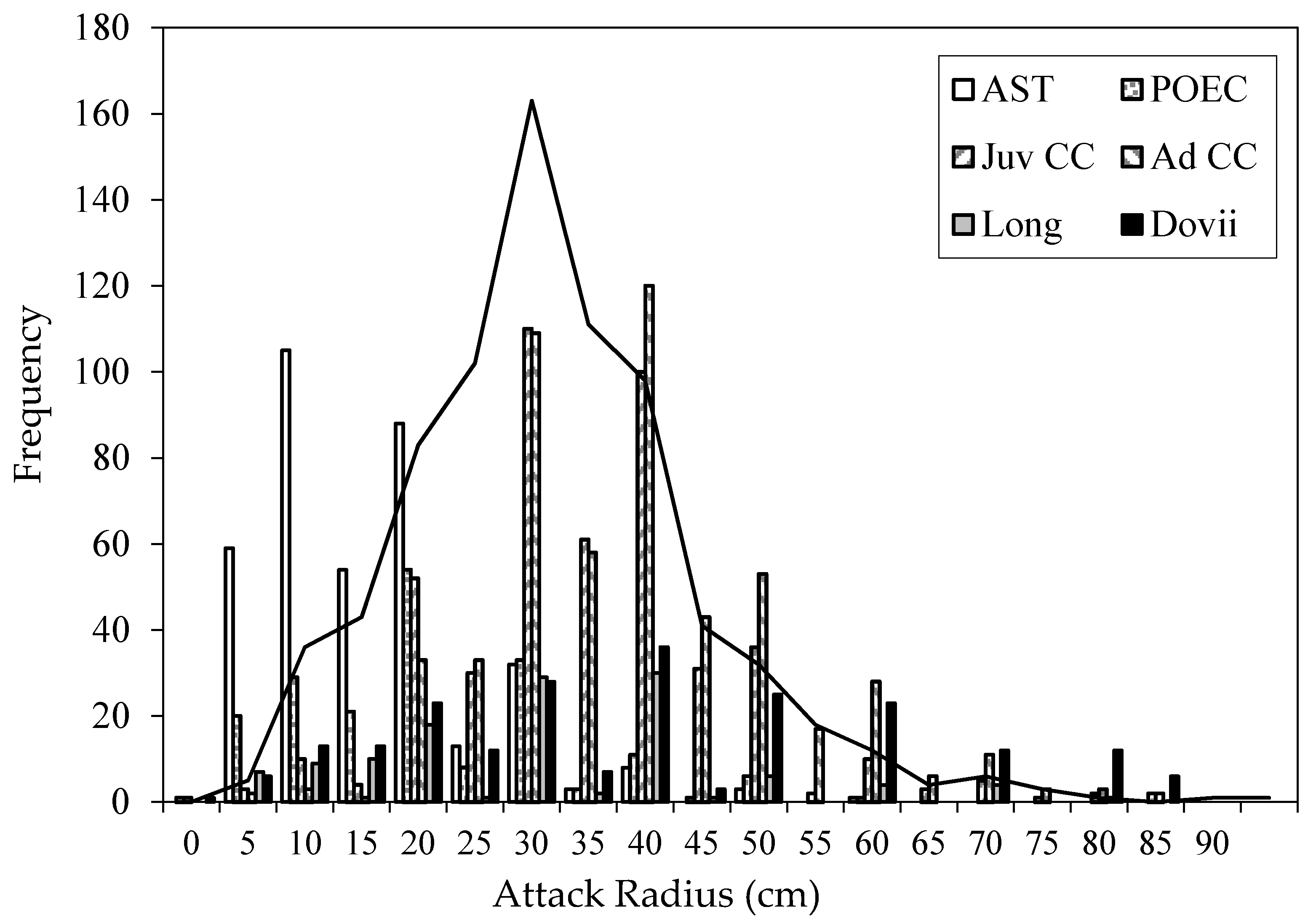
In Costa Rican streams, free-swimming young (“fry”) form a two-dimensional disc on the substrate. Rarely do individual fry rise up into the water column, presumably because river current above the boundary layer would sweep them away, and because interstitial spaces in the substrate are their refuge from brood predators. Parents are positioned centrally, hovering a few centimeters above the substrate and facing in opposite directions from each other, to provide 360° vigilance for the approach of intruders. Not all intruders pose the same degree of threat to the offspring. Poeciliids, especially Poecilia gillii, are common intruders but do not attack cichlid fry. The schooling characin Astyanax fasciatus also pass over broods and are merely nudged away by the parents. Astyanax attack offspring only if the brood is first disturbed by another cause, such as human manipulations of the brood, or an attack by large adult Parachromis dovii. Once an Astyanax attack begins, 50 or more others join in and overwhelm the defenses of the parents, which can disperse fry from the brood area, where they either find their way home using visual and chemical cues [12,15] or join neighboring families [16]. Parental defense attacks are most frequently directed against juvenile cichlids. In the Río Cabuyo and Quebrada Amores system, there were three species of cichlids: convict cichlids, a sand-sifting species Cribroheros longimanus, and the piscivorous P. dovii. Juvenile P. dovii were attacked from the greatest distances from the brood, suggesting that parents consider them as the most serious threat to their young (Figure 6). Indeed, juvenile P. dovii were often observed sneaking up on broods by creeping stealthily forward under the cover of leaf litter and hiding behind pebbles to stage an ambush.
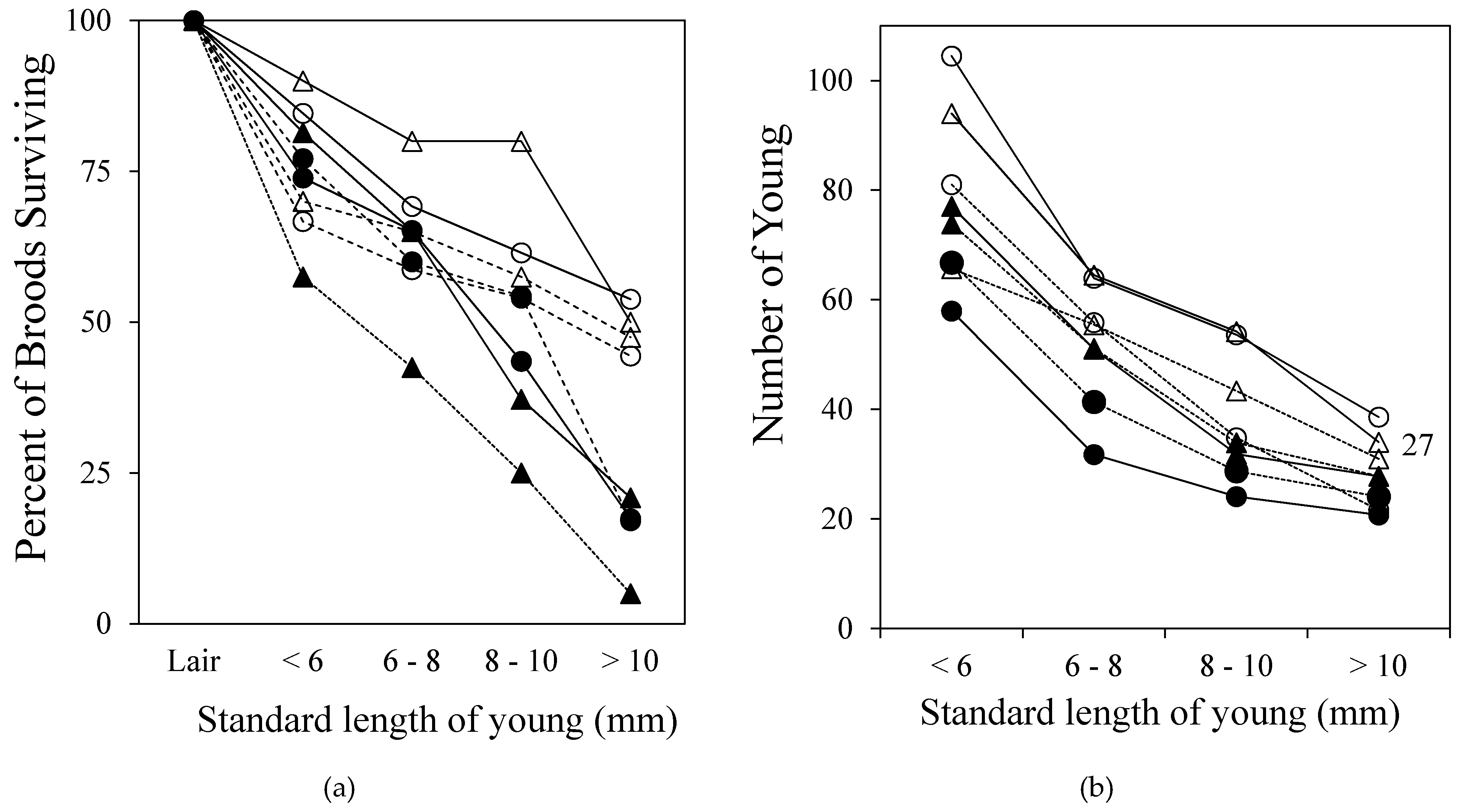
In spite of parental vigilance and brood defense, predators take their toll and reduce the number of young in a brood, and ultimately, the number of broods that persist with surviving offspring to fry independence. Brood success (at least one fry surviving to independence) is strongly linked to habitat (Figure 7a). In shallow sections of the streams (i.e., “stream” sites), brood survival was relatively high (48%) compared to broods reared in the deeper “pool” sections of the river (15%). Contributing factors to this disparity may be that P. dovii spawn in the deep sections of the rivers, thus their juveniles are more abundant there than in the shallow sections of the streams, and because deeper sections of the river are darker and parental detection of approaching brood predators may be impaired under these conditions. For those broods that had young survive to independence, the final number of young averaged 27 despite widely ranging starting numbers, parent size and habitat (Figure 7b).
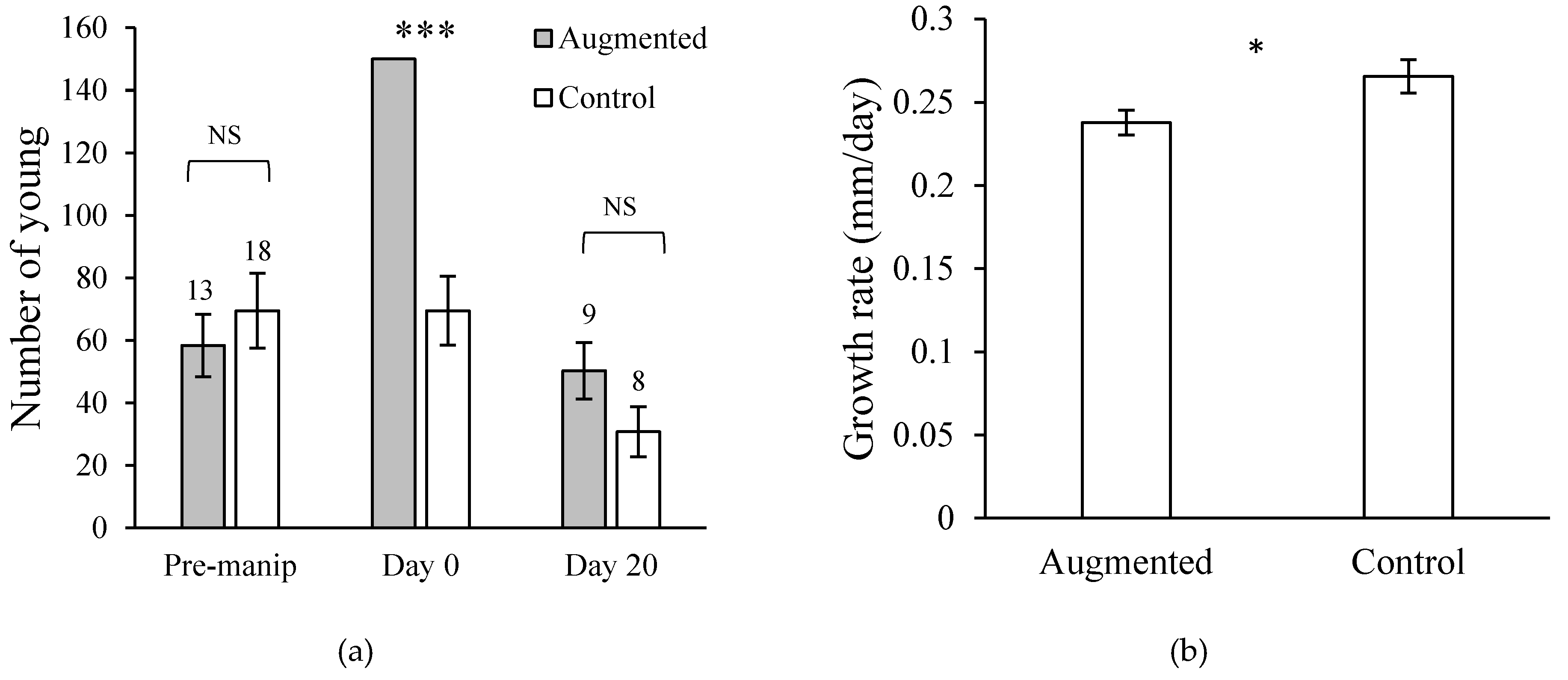
The convergence of offspring number of successful broods to 27 young suggested that there may be constraints on maximum brood size imposed by the logistics of brood defense (Lack’s Hypothesis). This was confirmed by a field manipulation experiment (Figure 8a). This experiment also demonstrated that per capita growth rate of the young was inversely proportional to the number of young in the brood, suggesting that the logistic constraint was the area defined by the effective radius of parental brood defense (Figure 8b). In large broods, young were forced to crowd into the safe area defendable by the parents and therefore foraged less than young in small broods.
5. Broods of Mixed Parentage and Intraspecific Brood Adoption
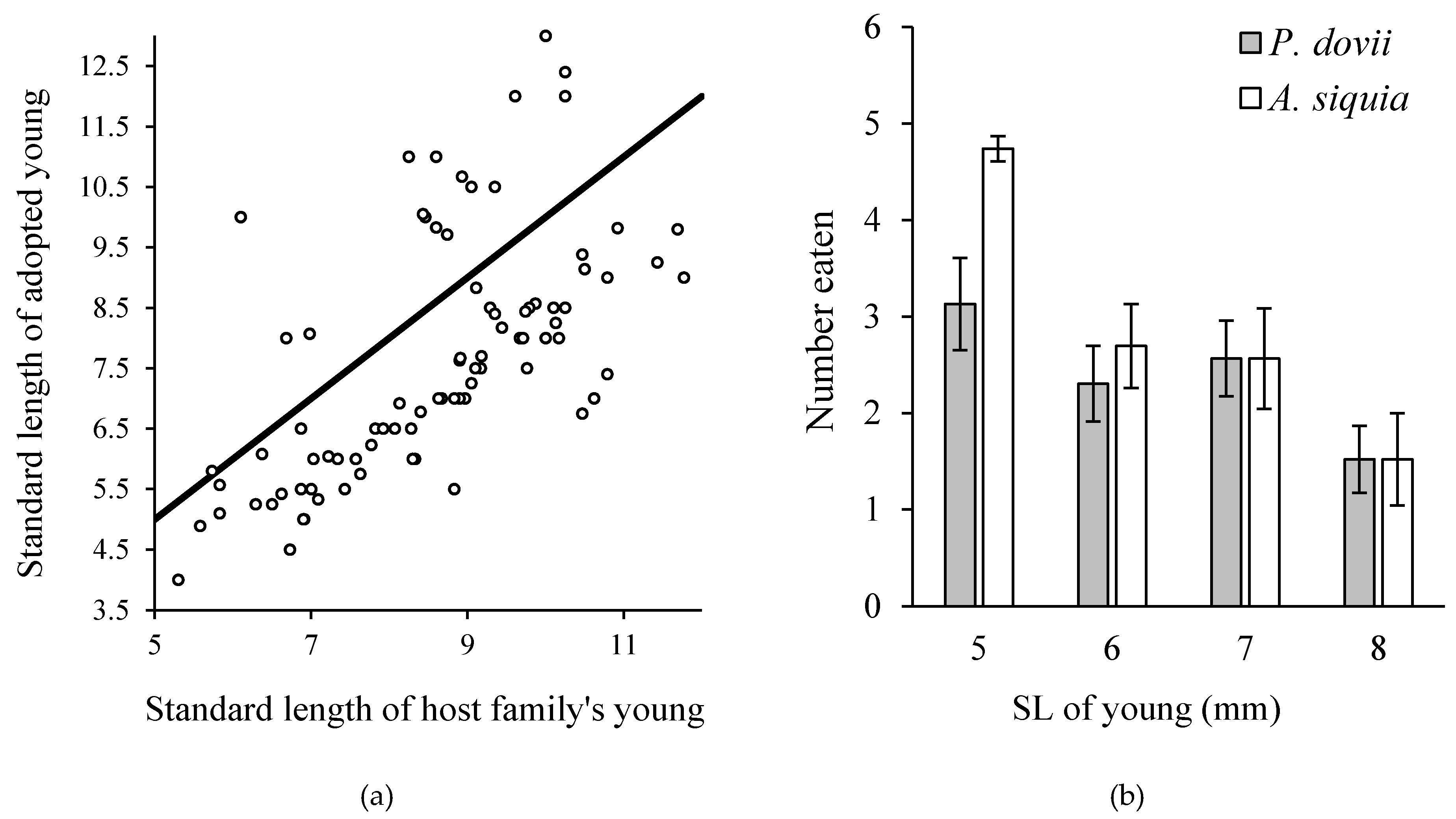
Recent genetic analyses of convict cichlid broods in Río Cabuyo show evidence of multiple maternity, multiple paternity, and wholesale adoption of young unrelated to either parent providing care [13]. An alternative “sneaker” male morph has been observed in Laguna de Xiloá in Nicaragua [14]. Clearly, the mating system of this species is one of social monogamy but not genetic monogamy. In addition to multiple parentage, convict cichlids also add offspring to their brood during the free-swimming stage while the young are being guarded [19]. In tracking the number of young in individual broods over time, it was often observed that the number of young in some broods increased instead of decreasing over time. In these cases, and in others in which the number of young declined, there appeared within the brood two distinct size classes: one that tracked the growth trajectory of the young “related” to the adults providing care, and additional young that were smaller than the young of the host family (Figure 9). Adoption was not a case of misdirected parental care because experimental addition of young that were similar in size or smaller than host young were always successfully adopted, whereas young larger than host young were always rejected [19]. Close examination of natural adoption events revealed that size discrimination occurred only until host young were between 7 and 8 mm SL, and thereafter there was no size bias (Figure 9).
We hypothesized that size-bias in brood adoption was driven by differential swimming performance between small and large young. Predation trials in the lab on broods of mixed sized demonstrated that predators preferentially consumed small fry (Figure 9) and a brood manipulation experiment in the field confirmed that parents that adopt additional young benefit from increasing the survival rate of their “own” young through statistical dilution of predation risk, and through differential predation on smaller adopted young [20].
6. Ontogeny of Antipredator Competence
Burst speed of young at different lengths showed that young became faster swimmers as they grew and developed, but that the rate of increase was not linear (Figure 10). Post-hoc pairwise comparison tests showed that swimming performance rapidly improved after the young reached 7 mm SL, which coincides with the size at which parents no longer discriminate on the basis of size when they adopt new young into their families.
7. Timing of Ossification of the Larval Skeleton from Cartilage to Bone
The skeleton of larval fish is cartilaginous at the time of hatching and then becomes ossified into bone in the transition from larvae to juvenile. Bone is stronger than cartilage and leads to more forceful swimming propulsion. Larvae that had been cleared and stained with alcian blue (cartilage) and alizarin red (bone) allowed us to quantify the timing of the transition from cartilage to bone (Figure 11). Ordination of the resulting data matrix revealed that rapid calcification of the skeleton occurred at around 7 mm SL, which coincides with changes in swimming performance, differential predation and discriminatory brood adoption on the basis of size (Figure 12). Taken together, these data show that predation on larval convict cichlids exerts selection pressure on the timing of developmental events, which affect the growth and survival rates of the young, and shapes the evolution of parental brood defense.
8. Comparison to Convict Cichlids in Laguna de Xiloá, Nicaragua
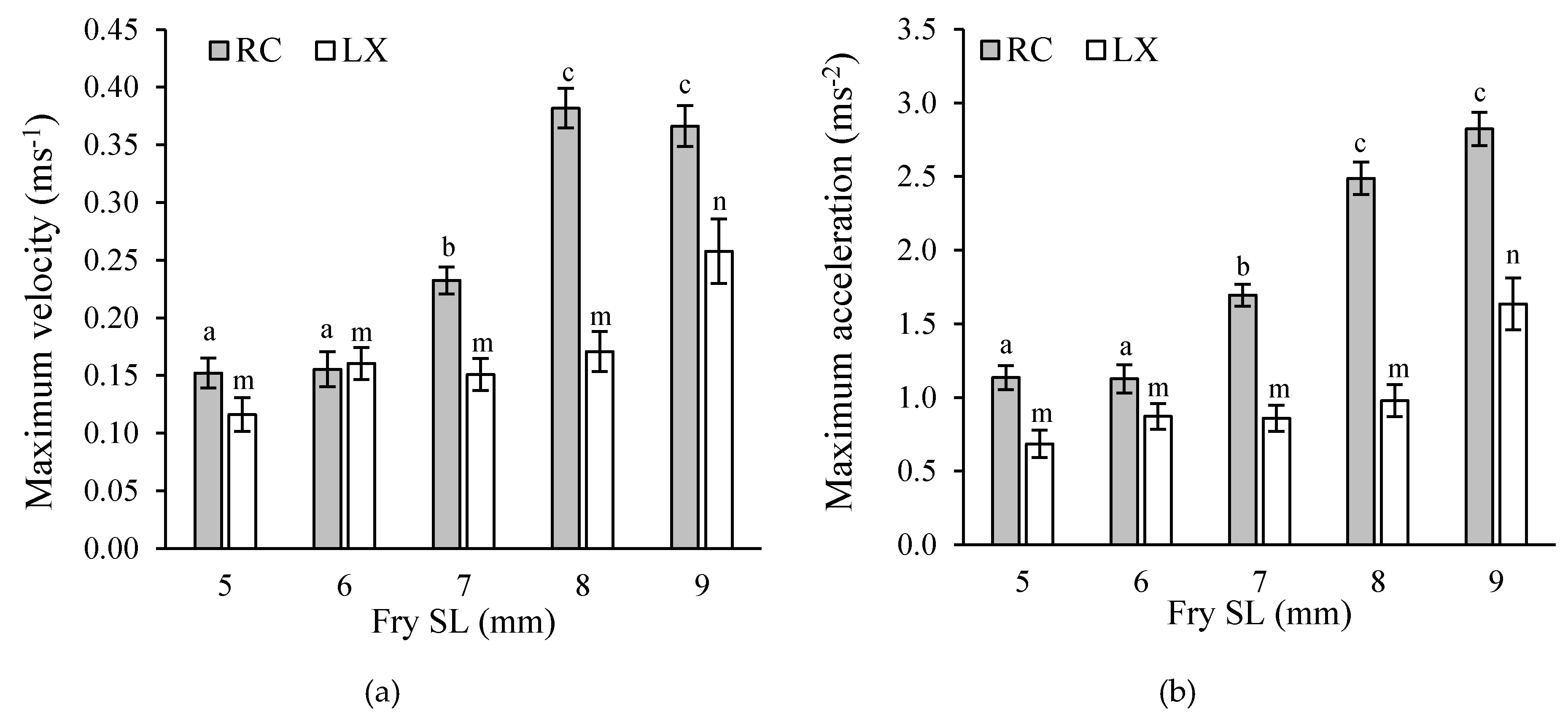
Data from Costa Rican streams indicate a correlation among predation pressure, larval development and parental care. Laguna de Xiloá is a volcanic crater lake in Nicaragua that differs from Costa Rican streams in several ways. It is lentic, it is not shaded by riparian vegetation therefore there is rich autochthonous production, and there is a diverse fish community including the sleeper goby Gobiomorus dormitor, which is a specialist predator on the larvae of cichlids. We collected and preserved larvae from convict cichlid broods from Laguna de Xiloá for analysis of ossification. Ordination of those data revealed that the timing of ossification was different than for fishes in the Rió Cabuyo system, being delayed to between 8 and 9 mm SL (Figure 12). Adults collected from Laguna de Xiloá and bred in the same laboratory tanks used to produce data for the Costa Rican population (i.e., common garden), showed that larvae from Nicaraguan parents showed delayed onset of swimming performance relative to Costa Rican fish. Maximum velocity and acceleration did not noticeably improve until the young were larger than 8 mm SL (Figure 10), which matches well with data on skeletal ossification, and presumably would continue to improve beyond 9 mm SL.
9. Correlation of Parental Brood Defense and Brood Predators
Growth rates of larvae showed that there is a limit to the area that can be effectively defended by two adults. Interacting with this constraint is the swimming speed of the young that enable them to evade an attack by a brood predator. When the young are small and weak-swimming, they occupy a small area that can be effectively guarded. As the young develop and improve in swimming performance, the area occupied by the brood should be expected to expand to reduce intrabrood competition for food. Measurements of brood diameter in the Rió Cabuyo and Laguna de Xiloá confirm that brood area does indeed expand as the young grow and develop into faster swimmers [21].
Change point analysis of brood radius indicates that in the Rió Cabuyo brood radius expands until the young are about 6.45 mm SL, and then levels off, whereas in Laguna de Xiloá brood area remains tight and cohesive until the young reach 7.9 mm SL and then expands in radius thereafter (Figure 13). In both cases, the switch points coincide with site-specific switch points in swimming performance and skeletal ossification. The reason(s) for why brood radius increases then plateaus in Rió Cabuyo but remains small and only later expands in Laguna de Xiloá is not known, but is likely due to a combination of relative predator pressure and relative productivity. Food is richly abundant in Laguna de Xiloá and so too is predator density and effectiveness. The best strategy for larvae (and their parents) in Laguna de Xiloá may be to stay huddled in a tight group for minimum risk to brood predators. They can afford to use this strategy because algal mats and associated epiphytic fauna are abundant in the shallow waters of Laguna de Xiloá where our data were collected. In contrast, the Rió Cabuyo is a low-order stream shaded almost completely by riparian canopy even in the dry season, thus food availability is less abundant. That, in combination with a relatively benign predator community, may favor early expansion of brood radius and therefore early ossification of the skeleton.
10. Discussion
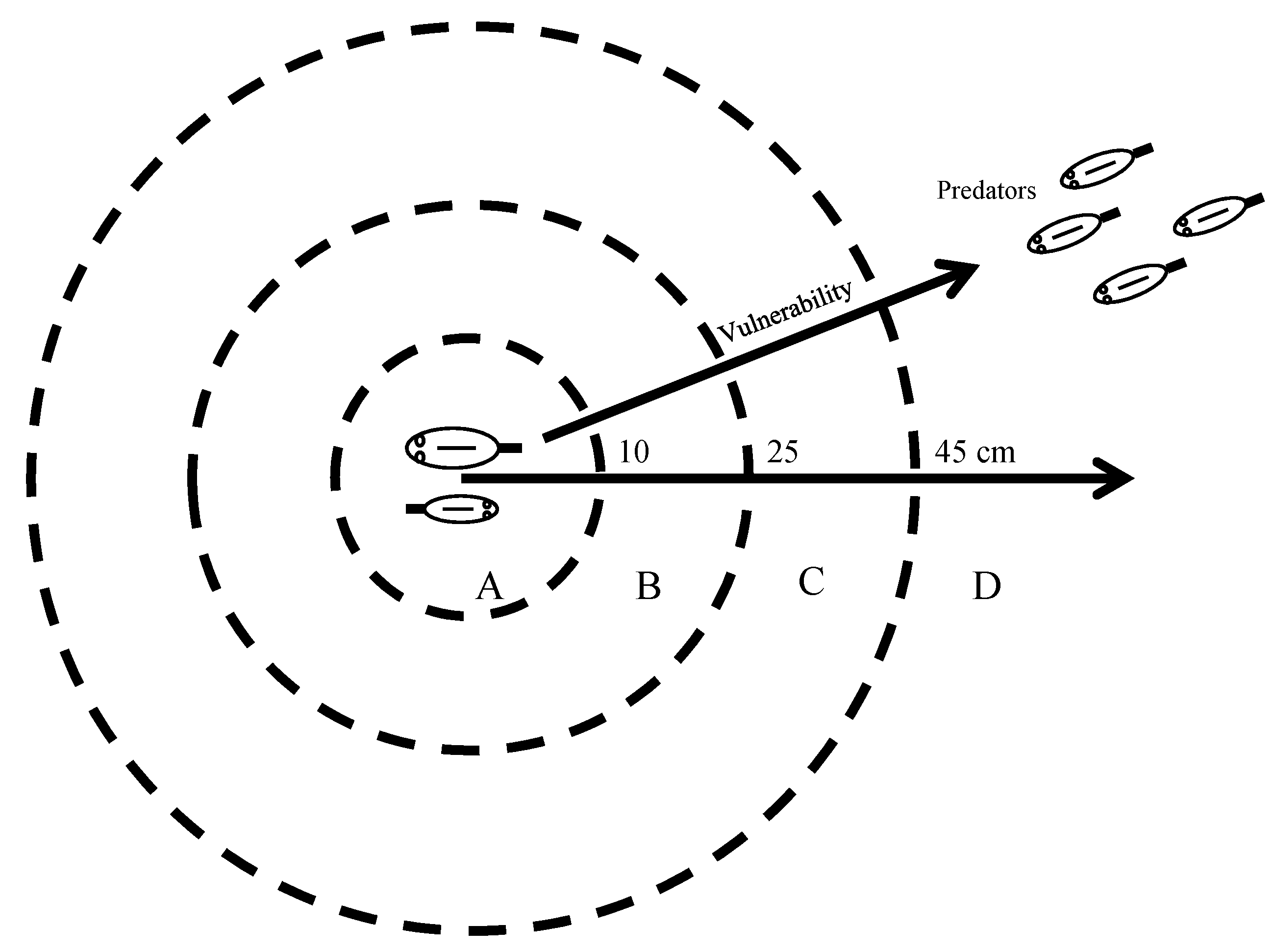
Brood defense is, in part, a matter of geometry (Figure 14). Risk of predation increases with distance from the brood center where the parents are positioned. The zone circumscribed by a radius of two body lengths is a zone of intimidation (zone A in Figure 14). Few attacks occur within this central area because intruders almost never approach the brood center. Aposematic coloration adopted by convict cichlids during the parental phase presumably help the young track the position of their parents, but they also warn intruders of the presence of highly aggressive fish.
Beyond a radius of 10 cm, parental attack rates quickly escalate, peaking for attack radii from 25 and 45 cm (zone C in Figure 14). Attacks with radii larger than 25 cm are usually a quick burst out to intercept the intruder followed by a quick return to the brood center because driving a predator away requires that the parent must temporarily leave their brood vulnerable to attack from the opposite direction. Parents differentially attack species that pose a greater threat to the young, namely juvenile conspecifics and juvenile P. dovii. Juvenile convict cichlids are opportunistic predators, numerically abundant in this system and attacked frequently. P. dovii are piscivores and although not common in this system, judging by attack distances, P. dovii represent a greater threat to cichlid fry than conspecific juveniles do.
Zones of parental protection constrain how far fry can stray from their parents while foraging. There is a trade-off for young between foraging (growth) and risk of predation. This trade-off shifts as swimming performance improves. Brood diameter increased with fry SL independent of the number of young in the brood [17]. Note that brood radii for convict cichlid broods in the Río Cabuyo do not extend beyond 20 cm until after the skeleton has fully ossified. In Laguna de Xiloá the relationship between development of the young and brood radius is the inverse of the pattern observed in the Río Cabuyo. Young in both populations remain in a tight shoal until they attain a length that coincides with skeletal ossification and then begin to stray from their parents. Larval and juvenile convict cichlids in Laguna de Xiloá remain with their parents until they have grown much larger than typical size-at-dispersal observed for broods in Río Cabuyo. This may reflect greater threat of predation and/or richer food, or other unmeasured parameters in the laguna.
The direction of change in ontogenetic timing in response to predation could occur in either direction. If predation selects for fast escape responses early in development, then one might predict earlier onset of calcification. A second possibility is that intense predation risk selects for extended periods of tight shoaling behavior instead of individual escape responses, allowing young to allocate more resources into growth in physical size rather than to ossification of skeletal components. There is support for the latter hypothesis from population differences in pumpkinseed sunfish (Lepomis gibbosus), in a population with faster growth rates showing delayed ossification of cranial bones [22]. Similarly, medaka (Oryzias latipes) from relatively fast-growing populations have fewer skeletal supports for fins and poorer swimming performance that same-sized fish from slow-growing populations [23]. It seems likely that convict cichlids in Laguna de Xiloá have been selected for fast growth and therefore delayed skeletal ossification until they have attained a relatively large size compared to convict cichlids in Río Cabuyo.
11. Conclusions
Convict cichlids in Rio Cabuyó and Laguna de Xiloá are different from one another. Divergence in the timing of skeletal ossification leads to divergent behavioral phenotypes of larvae. We know divergent development is not a result of developmental plasticity or food availability because the ossification data were derived from lab-reared adults held under identical conditions (i.e., a “common garden”). Population differences reflect genetic differences, not environmental ones. Skeletal ossification changes swimming performance and therefore vulnerability to brood predators, which in turn alters selection on parental care behavior in terms of brood defense and fine-grain decision-making, such as when to accept or reject new young that join their brood.
Allopatric population divergences are consistent with incipient speciation. Reproductively, isolation occurs by mechanisms of foraging competitive exclusion and reproductive behaviors including female mate choice, depth and timing of spawning, e.g., [24,25]. Laguna de Xiloá is well known for several examples of sympatric speciation of other cichlid species [26,27,28], a much younger evolutionarily radiation than the celebrated radiations that have occurred in cichlid species of the rift valley lakes in East Africa. Species radiations in cichlids in Laguna de Xiloá, and African rift valley cichlids radiated due to sexual selection or ecological niche partitioning. In contrast, it seems very likely that the cause of divergence in convict cichlid populations in Costa Rica streams and a Nicaraguan lake is predation. Predation is known to lead to population divergences in other systems (e.g., [29,30,31,32]). However, the interacting roles of larval development, larval antipredator competence, and parental care in driving species diversity is not well explored (but see [33,34]). These traits could be added to the list of traits that cause reproductive isolation through suppression of hybrid crosses.
Brood defense is maintained by kin-selected benefits to parents derived from increased offspring survival. Across cichlids, patterns of parental care vary widely in terms of duration of care, whether care is uniparental, biparental or multigenerational, and whether eggs are attached to the substrate or incubated orally [35]. But in every case, predation on eggs, larvae and juveniles is the driver of selection for these behaviors and physiological mechanisms. Cichlids offer a remarkably diverse system with which to explore the relationship among predation, the evolution of parental care and species diversity.
There are several potentially fruitful avenues for future study to explore the extent to which the phenomena described here apply to other populations and other species. The data presented here are based on two populations that differ in many ways, including predation pressure on fry. Inclusion of additional populations will increase resolution of the causal relationships between brood predators, food, temperature, spawning site availability, adult density, etc., that may all shape patterns of parental care. Non-cichlid species that provide care for their young offer an opportunity to test these relationships independent from phylogenetic effects. Ultimately, a goal of this research would be to develop a quantitative framework that integrates life history traits with the environmental factors that shape them.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/1424-2818/12/4/136/s1, Video S1: Video of female-female intrasexual contest over a male.
Funding
The data reported here were funded by the National Science and Research Council of Canada, and internal grants from Minnesota State University Moorhead from the Dille Fund for Excellence, and Faculty Research Grants to BDW.
Acknowledgments
This review summarizes approximately 30 years of research on the evolutionary ecology of convict cichlids in the field and lab. I have had many collaborators in this effort, listed as coauthors in the cited references. Miles Keenleyside deserves special mention for expertly and patiently guiding the early stages of this work. Patricia (Bee) Wisenden was involved throughout the 30+ years of this research as a field assistant and life partner. Gordon and Jutta Frankie made the Costa Rican field work possible. Ken McKaye and Eric van den Berghe made work in Laguna de Xiloá possible, and Eric collected and shipped LX fish to my lab in Minnesota.
Conflicts of Interest
The author declares no conflict of interest.
References
- Lima, S.L.; Dill, L.M. Behavioral decisions made under the risk of predation: A review and prospectus. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Stankowich, T.; Blumstein, D.T. Fear in animals: A meta-analysis and review of risk assessment. Proc. R. Soc. B Biol. Sci. 2005, 272, 2627–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, M.R. The evolutionary consequences of indirect effects. Trends Ecol. Evol. 2013, 28, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Palkovacs, E.P.; Wasserman, B.A.; Kinnison, M.T. Eco-evolutionary trophic dynamics: Loss of top predators drives trophic evolution and ecology of prey. PLoS ONE 2011, 6, e18879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussing, W.A. Peces de las Aguas Continentals de Costa Rica, 2nd ed.; Editorial de la Universidad de Costa Rica: San Jose, CA, USA, 2002; 468p. [Google Scholar]
- Schmitter-Soto, J.J. A systematic revision of the genus Archocentrus (Perciformes: Cichlidae), with the description of two new genera and six new species. Zootaxa 2007, 1603, 1–78. [Google Scholar] [CrossRef] [Green Version]
- Bagley, J.; Matamoros, W.A.; Mcmahan, C.D.; Tobler, M.; Chakrabarty, P.; Johnson, J.B. Phylogeography and species delimitation in convict cichlids (Cichlidae: Amatitlania): Implications for taxonomy and Plio-Pleistocene evolutionary history in Central America. Boil. J. Linn. Soc. 2016, 1, 155–170. [Google Scholar]
- Wisenden, B.D.; Lanfranconi-Izawa, T.L.; Keenleyside, M.H.A. Fin digging and leaf lifting: Examples of parental food provisioning by the cichlid fish Cichlasoma nigrofasciatum. Anim. Behav. 1995, 49, 623–639. [Google Scholar] [CrossRef]
- Wisenden, B.D. Factors affecting male mate desertion in the biparental cichlid fish (Cichlasoma nigrofasciatum) in Costa Rica. Behav. Ecol. 1994, 5, 439–447. [Google Scholar] [CrossRef]
- Wisenden, B.D. Factors affecting reproductive success of convict cichlids in Costa Rican streams. Can. J. Zool. 1994, 72, 2177–2185. [Google Scholar] [CrossRef] [Green Version]
- Wisenden, B.D. Reproductive behaviour in free-ranging convict cichlids. Environ. Biol. Fish. 1995, 43, 121–134. [Google Scholar] [CrossRef]
- Wisenden, B.D.; Snekser, J.L.; Stumbo, A.D.; Leese, J.M. Parental defense of an empty nest after catastrophic brood loss. Anim. Behav. 2008, 76, 2059–2067. [Google Scholar] [CrossRef]
- Lee-Jenkins, S.S.Y.; Smith, M.L.; Wisenden, B.D.; Wong, A.; Godin, J.-G.J. Genetic evidence for mixed broods and extra-pair fertilizations in a socially monogamous biparental cichlid fish. Behaviour 2015, 152, 1507–1526. [Google Scholar] [CrossRef] [Green Version]
- Clotfelter, E.D.; Johnson, E.M.; Anderson, C.; Earley, R.L. Evidence for male alternative reproductive tactics in convict cichlids (Amatitlania siquia) in Lake Xiloá, Nicaragua. Environ. Biol. Fish. 2017, 100, 655–662. [Google Scholar] [CrossRef]
- Wisenden, B.; Dye, T.P. Young convict cichlids use visual information to update olfactory homing cues. Behav. Ecol. Sociobiol. 2008, 63, 443–449. [Google Scholar] [CrossRef]
- Wisenden, B.D.; Mammenga, E.A.; Storseth, C.N.; Berglund, N.J. Odor-tracking by larval convict cichlids and a mechanism for alloparental brood amalgamation. Anim. Behav. 2014, 93, 201–206. [Google Scholar] [CrossRef]
- Wisenden, B.D.; Stumbo, A.D.; Self, P.A.; Snekser, J.; McEwen, D.C.; Wisenden, P.A.; Keenleyside, M.H.A.; Itzkowitz, M.; Brisch, E. Co-evolution of offspring antipredator competence and parental brood defense in convict cichlids. Hydrobiology 2014, 748, 259–272. [Google Scholar] [CrossRef]
- Wisenden, B.D.; Keenleyside, M.H. Brood size and the economy of brood defence: Examining Lack’s hypothesis in a biparental cichlid fish. Environ. Biol. Fishes 1995, 43, 145–151. [Google Scholar] [CrossRef]
- Wisenden, B.; Keenleyside, M. Intraspecific brood adoption in convict cichlids: A mutual benefit. Behav. Ecol. Sociobiol. 1992, 31, 263–269. [Google Scholar] [CrossRef]
- Wisenden, B.D.; Keenleyside, M.H.A. The dilution effect and differential predation following brood adoption in free-ranging convict cichlids (Cichlasoma nigrofasciatum). Ethol. 2010, 96, 203–212. [Google Scholar] [CrossRef]
- Wisenden, B.D.; Stumbo, A.D.; McEwen, D.C.; McIntire, K.; Scheierl, J.; Aasand, J.; North, H.; Gilbertson, J.; Grant, D.; Joseph, F.; et al. Population-specific co-evolution of offspring antipredator competence and parental brood defense in Nicaraguan convict cichlids. Environ. Biol. Fishes 2016, 99, 325–333. [Google Scholar] [CrossRef]
- Arendt, J.D.; Wilson, D.S. Population differences in the onset of cranial ossification in pumpkinseed (Lepomis gibbosus), a potential cost of rapid growth. Can. J. Fish. Aquat. Sci. 2000, 57, 351–356. [Google Scholar] [CrossRef]
- Fujimoto, S.; Kawajiri, M.; Suzuki, Y.; Yamahira, K. Evolution of growth rates under the constraint of growth-development trade-off in a fish. Popul. Ecol. 2011, 54, 275–283. [Google Scholar] [CrossRef]
- Schluter, L.; Rambaut, A. Ecological speciation in postglacial fishes. Philos. Trans. R. Soc. B Boil. Sci. 1996, 351, 807–814. [Google Scholar]
- Seehousen, O. Process and pattern in cichlid radiations—Inferences for understanding unusually high rates of evolutionary diversification. New Phytol. 2015, 207, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Barluenga, M.; Stölting, K.N.; Salzburger, W.; Muschick, M.; Meyer, A. Sympatric speciation in Nicaraguan crater lake cichlid fish. Nature 2006, 439, 719–723. [Google Scholar] [CrossRef]
- Elmer, K.R.; Fan, S.; Gunter, H.; Jones, J.C.; Boekhoff, S.; Kuraku, S.; Meyer, A. Rapid evolution and selection inferred from the transcriptomes of sympatric crater lake cichlid fishes. Mol. Ecol. 2010, 19, 197–211. [Google Scholar] [CrossRef]
- Kautt, A.F.; Elmer, K.R.; Meyer, A. Genomic signatures of divergent selection and speciation patterns in a ‘natural experiment’, the young parallel radiations of Nicaraguan crater lake cichlid fishes. Mol. Ecol. 2012, 21, 4770–4786. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A. The inheritance and development of minnow anti-predator behaviour. Anim. Behav. 1990, 39, 834–842. [Google Scholar] [CrossRef]
- Reznick, D.N.; Butler, M.J.I.V.; Rodd, F.H.; Ross, P. Life-history evolution in guppies (Poecilia reticulata). 6. Differential mortality as a mechanism for natural selection. Evolution 1996, 50, 1651–1660. [Google Scholar]
- Johnson, J.; Belk, M. Predation environment predicts divergent life-history phenotypes among populations of the livebearing fish Brachyrhaphis rhabdophora. Oecologia 2001, 126, 142–149. [Google Scholar] [CrossRef]
- Wund, M.A.; Baker, J.A.; Golub, J.L.; Foster, S.A. The evolution of antipredator behavior following relaxed and reversed selection in Alaskan threespine stickleback fish. Anim. Behav. 2015, 106, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolm, N.; Goodwin, N.B.; Balshine, S.; Reynolds, J.D. Life history evolution in cichlids 1: Revisiting the evolution of life histories in relation to parental care. J. Evol. Biol. 2006, 19, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Kolm, N.; Goodwin, N.B.; Balshine, S.; Reynolds, J.D. Life history evolution in cichlids 2: Directional evolution of the trade-off between egg number and egg size. J. Evol. Biol. 2006, 19, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Keenleyside, M.H.A. Parental care. In Cichlid Fishes: Behaviour, Ecology and Evolution; Chapman and Hall: New York, NY, USA, 1991; pp. 191–208. [Google Scholar]
Figure 1.
Map of Central America indicating the location of Lomas Barbudal Biological Reserve and Laguna de Xiloá.
Figure 1.
Map of Central America indicating the location of Lomas Barbudal Biological Reserve and Laguna de Xiloá.

Figure 2.
The number of breeding pairs actively guarding broods in the dry seasons of 1990 and 1991 within defined study areas in the Río Cabuyo and its tributary Quebrada Amores, located in and adjacent to Lomas Barbudal Biological Reserve, Guanacaste, Costa Rica. Data from [9].
Figure 2.
The number of breeding pairs actively guarding broods in the dry seasons of 1990 and 1991 within defined study areas in the Río Cabuyo and its tributary Quebrada Amores, located in and adjacent to Lomas Barbudal Biological Reserve, Guanacaste, Costa Rica. Data from [9].

Figure 3.
(a) Male standard length (mm) versus female standard length in mated pairs of convict cichlids by 1990 and 1991 in the pool (P) and stream (S) habitat in Río Cabuyo and Quebrada Amores. Large males monopolize spawning sites and large females produce larger clutches leading to size-assortative mating. (b) Overall mean ± SE length difference within a pair was 13.78 ± 0.59 mm (n = 149), and intrapair difference differed significantly by site (F3,145 = 44.25, p < 0.001). Small females were not found spawning in Quebrada Amores as often as they occurred in Río Cabuyo. Data from [10].
Figure 3.
(a) Male standard length (mm) versus female standard length in mated pairs of convict cichlids by 1990 and 1991 in the pool (P) and stream (S) habitat in Río Cabuyo and Quebrada Amores. Large males monopolize spawning sites and large females produce larger clutches leading to size-assortative mating. (b) Overall mean ± SE length difference within a pair was 13.78 ± 0.59 mm (n = 149), and intrapair difference differed significantly by site (F3,145 = 44.25, p < 0.001). Small females were not found spawning in Quebrada Amores as often as they occurred in Río Cabuyo. Data from [10].

Figure 4.
(a) Seasonal mating frequency (Dec–May) for males and females for all sites combined. 28% of males spawned more than once, 5% of females spawned more than once. (b) Mean ± SE number of spawning lairs used as a function of seasonal mating frequency. Males often reused the same site for sequential broods. Data from [11].
Figure 4.
(a) Seasonal mating frequency (Dec–May) for males and females for all sites combined. 28% of males spawned more than once, 5% of females spawned more than once. (b) Mean ± SE number of spawning lairs used as a function of seasonal mating frequency. Males often reused the same site for sequential broods. Data from [11].

Figure 5.
Color phases of convict cichlids. Males alternate between drab olive-gray when non reproductive as in the upper fish in (a), to contrasting black and white when defending young (female is ahead of the male in this image) (b). Females have an additional color phase when gravid and courting males in which the background color become black as in the lower fish in (a), image (c), which provides a striking contrast for gold flecks on the belly and yellow and blue pigments in the fins and throat. Photos of fish in hand by Brian Wisenden, photo of breeding pair (b) by Terence Lee.
Figure 5.
Color phases of convict cichlids. Males alternate between drab olive-gray when non reproductive as in the upper fish in (a), to contrasting black and white when defending young (female is ahead of the male in this image) (b). Females have an additional color phase when gravid and courting males in which the background color become black as in the lower fish in (a), image (c), which provides a striking contrast for gold flecks on the belly and yellow and blue pigments in the fins and throat. Photos of fish in hand by Brian Wisenden, photo of breeding pair (b) by Terence Lee.

Figure 6.
Parental brood defense attacks per 10 min observation period (n = 840 trials) against the characin Astyanax fasciatus (AST), Poecilia gillii (POEC), juvenile convict cichlids (Juv CC), adult convict cichlids (Ad CC), sand-sifter cichlid Cribroheros longimanus (Long), or juveniles of the piscivore Parachromis dovii (Dovii). Line represents overall frequency distribution of parental attacks against brood predators. Data from [10,17].
Figure 6.
Parental brood defense attacks per 10 min observation period (n = 840 trials) against the characin Astyanax fasciatus (AST), Poecilia gillii (POEC), juvenile convict cichlids (Juv CC), adult convict cichlids (Ad CC), sand-sifter cichlid Cribroheros longimanus (Long), or juveniles of the piscivore Parachromis dovii (Dovii). Line represents overall frequency distribution of parental attacks against brood predators. Data from [10,17].

Figure 7.
(a) Percentage of broods surviving at various intervals between emergence from the lair until independence from parental care after the young reach 10 mm SL. (b) Number of young in broods at these same intervals for each site. Although there were site differences in brood size initially, all site-years converged on an average of 27 fry at independence. Solid symbols, pool habitat; open symbols, stream habitat. Circles, Río Cabuyo; triangles, Quebrada Amores. Solid lines, 1990; dashed lines, 1991. Data from [9].
Figure 7.
(a) Percentage of broods surviving at various intervals between emergence from the lair until independence from parental care after the young reach 10 mm SL. (b) Number of young in broods at these same intervals for each site. Although there were site differences in brood size initially, all site-years converged on an average of 27 fry at independence. Solid symbols, pool habitat; open symbols, stream habitat. Circles, Río Cabuyo; triangles, Quebrada Amores. Solid lines, 1990; dashed lines, 1991. Data from [9].

Figure 8.
(a) Lack’s hypothesis is that maximum brood size is limited by the number of offspring that can be reared to independence, not by the physiological capacity to produce eggs. Naturally-occurring broods in the Río Cabuyo were either left unmanipulated (control) or augmented with extra fry to have 150 young. Experimental broods were unable to sustain numerical advantage of the manipulation by the end of 20 days. (b) Individual growth rates of larvae in experimental broods were significantly lower than larvae in control broods due to crowding within the parentally-defended area and therefore experienced increased competition for food. * p < 0.05, *** p < 0.001. Data from [18].
Figure 8.
(a) Lack’s hypothesis is that maximum brood size is limited by the number of offspring that can be reared to independence, not by the physiological capacity to produce eggs. Naturally-occurring broods in the Río Cabuyo were either left unmanipulated (control) or augmented with extra fry to have 150 young. Experimental broods were unable to sustain numerical advantage of the manipulation by the end of 20 days. (b) Individual growth rates of larvae in experimental broods were significantly lower than larvae in control broods due to crowding within the parentally-defended area and therefore experienced increased competition for food. * p < 0.05, *** p < 0.001. Data from [18].

Figure 9.
(a) Size-biased conspecific adoption of broods from neighboring families. Line is y = x. When young are adopted from neighboring families, parents accept young that are smaller than their own and reject those that are larger than their own (data not shown), until the host young are larger than 8 mm SL. This raised the possibility that adopted young are being set up as targets for predators to spare predation on host young, which was confirmed in a subsequent field experiment. (b) Hunting success of juvenile convict cichlids and juvenile P. dovii on groups of convict cichlid fry of mixed sizes. Small fry were eaten significantly more often than large fry. Data from [18].
Figure 9.
(a) Size-biased conspecific adoption of broods from neighboring families. Line is y = x. When young are adopted from neighboring families, parents accept young that are smaller than their own and reject those that are larger than their own (data not shown), until the host young are larger than 8 mm SL. This raised the possibility that adopted young are being set up as targets for predators to spare predation on host young, which was confirmed in a subsequent field experiment. (b) Hunting success of juvenile convict cichlids and juvenile P. dovii on groups of convict cichlid fry of mixed sizes. Small fry were eaten significantly more often than large fry. Data from [18].

Figure 10.
(a) Maximum velocity (m·s−1) and (b) acceleration (m·s−2) for cichlid fry of lengths 5 to 9 mm SL, from parents from Río Cabuyo (RC) in Costa Rica and from Laguna de Xiloá (LX) in Nicaragua. Letters above bars indicate post-hoc pairwise comparison tests across fry sizes within each site. Data from [17,21].
Figure 10.
(a) Maximum velocity (m·s−1) and (b) acceleration (m·s−2) for cichlid fry of lengths 5 to 9 mm SL, from parents from Río Cabuyo (RC) in Costa Rica and from Laguna de Xiloá (LX) in Nicaragua. Letters above bars indicate post-hoc pairwise comparison tests across fry sizes within each site. Data from [17,21].

Figure 11.
(a) Cleared and stained larvae and juveniles with alcian blue (cartilage) and alizarin red (bone) for Río Cabuyo fish sized 5–9 mm SL. (b) Each specimen was imaged at high magnification so that each of 111 skeletal elements could be scored for degree of ossification on a scale of 1–4. Photos by Tony Stumbo. Data from [17].
Figure 11.
(a) Cleared and stained larvae and juveniles with alcian blue (cartilage) and alizarin red (bone) for Río Cabuyo fish sized 5–9 mm SL. (b) Each specimen was imaged at high magnification so that each of 111 skeletal elements could be scored for degree of ossification on a scale of 1–4. Photos by Tony Stumbo. Data from [17].

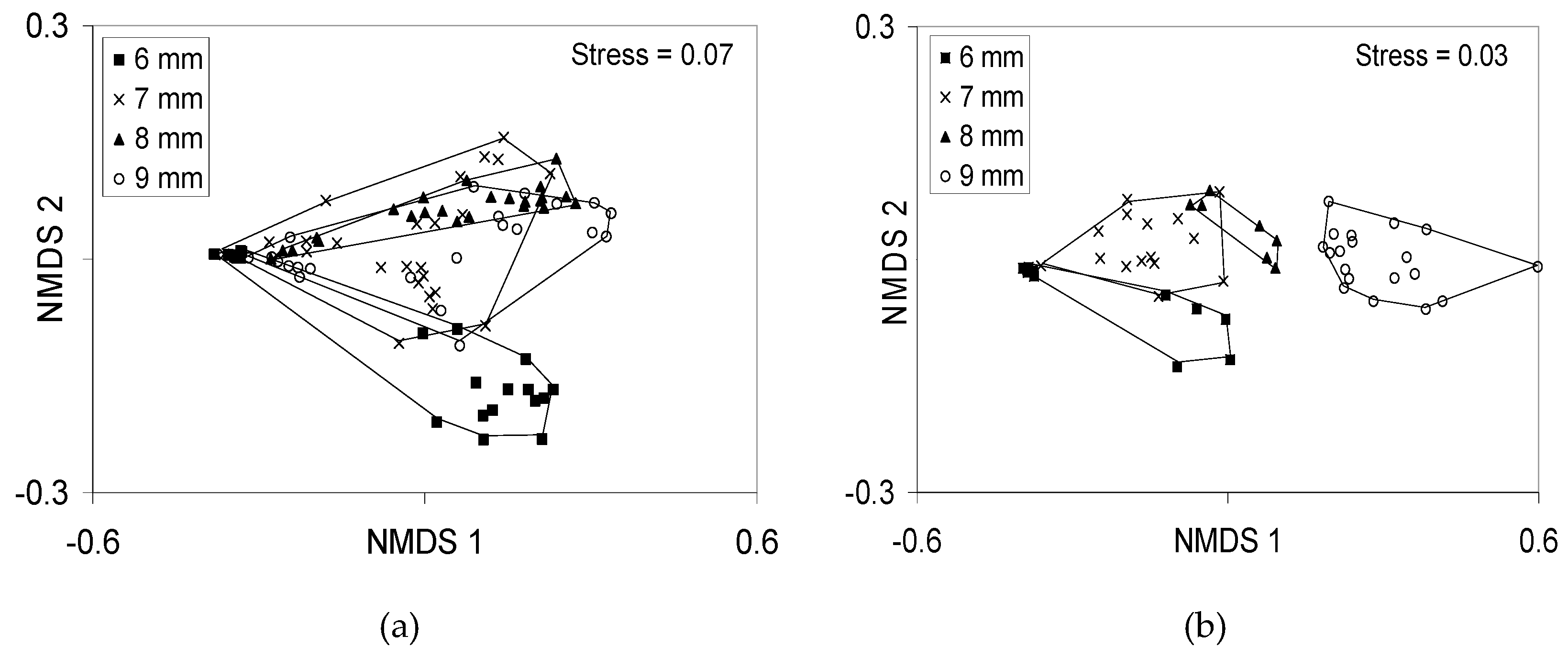
Figure 12.
(a) Non-metric dimensional scaling (NMDS) axes for 111 bones × 125 larvae from Río Cabuyo. There was no variation in 5 mm fry (not shown). Pairwise comparisons revealed that ossification of 6 mm fry < 7 = 8 = 9 mm SL fry. (b) NMDS ordination of images from larvae from lab-F1 adults from Laguna de Xiloá. Again 5 mm SL fish contained no variation (not shown), but the conspicuous shift in ossification was between 8 mm fry and 9 mm fry. Data from [17,21].
Figure 12.
(a) Non-metric dimensional scaling (NMDS) axes for 111 bones × 125 larvae from Río Cabuyo. There was no variation in 5 mm fry (not shown). Pairwise comparisons revealed that ossification of 6 mm fry < 7 = 8 = 9 mm SL fry. (b) NMDS ordination of images from larvae from lab-F1 adults from Laguna de Xiloá. Again 5 mm SL fish contained no variation (not shown), but the conspicuous shift in ossification was between 8 mm fry and 9 mm fry. Data from [17,21].

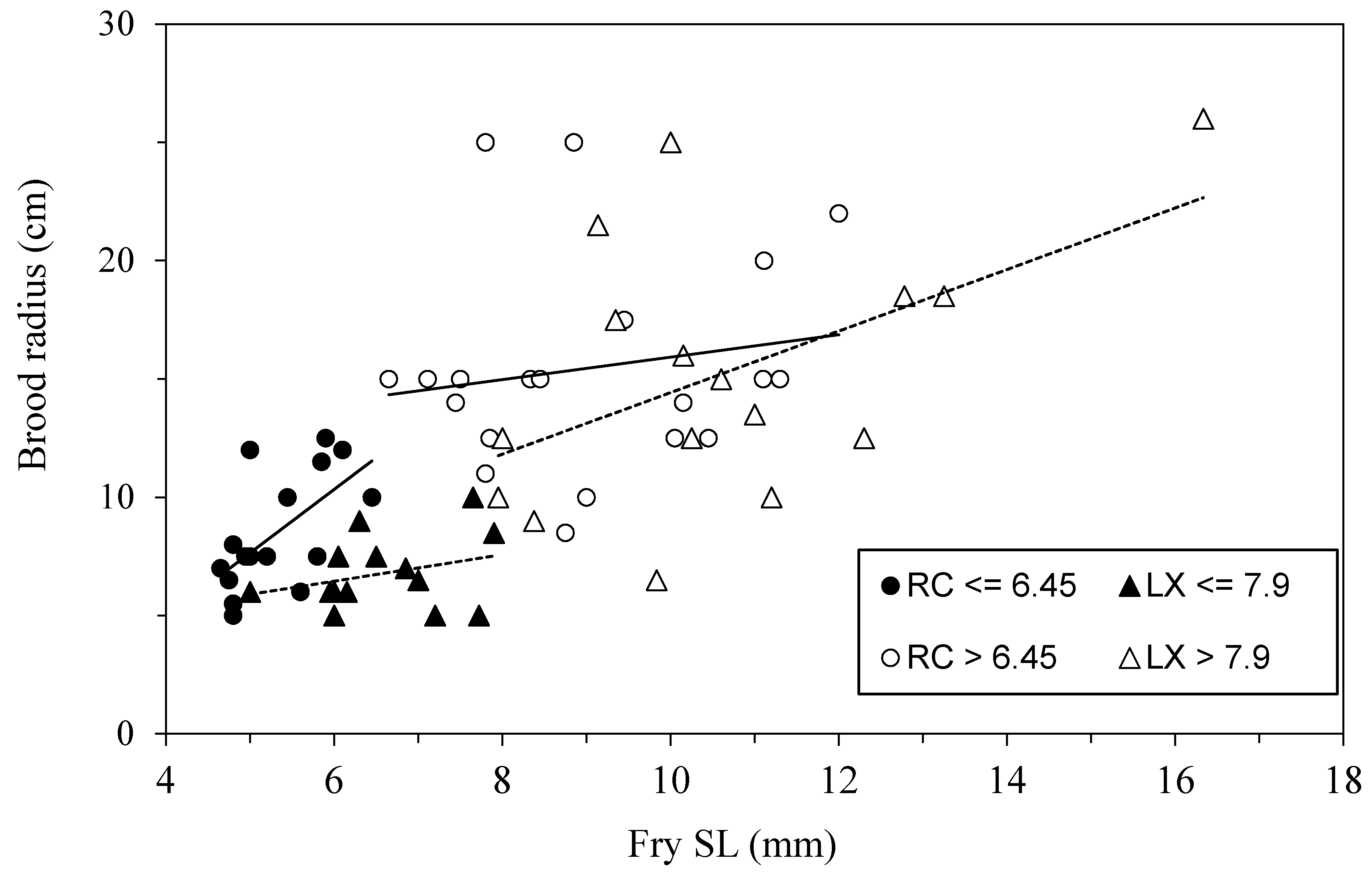
Figure 13.
The radius of broods being guarded in Río Cabuyo (circles) and Laguna de Xiloá (triangles) from time of early emergence from the spawning lair to the point of independence from parental care. Change-point analysis revealed nonlinearity occurred at SL = 6.45 in Río Cabuyo fish and SL = 7.9 for fish in Laguna de Xiloá. These switch points coincide with changes in skeletal ossification or the young, ontogenetic shifts in swimming performance and changes in parental adoption behavior. Data from [21].
Figure 13.
The radius of broods being guarded in Río Cabuyo (circles) and Laguna de Xiloá (triangles) from time of early emergence from the spawning lair to the point of independence from parental care. Change-point analysis revealed nonlinearity occurred at SL = 6.45 in Río Cabuyo fish and SL = 7.9 for fish in Laguna de Xiloá. These switch points coincide with changes in skeletal ossification or the young, ontogenetic shifts in swimming performance and changes in parental adoption behavior. Data from [21].

Figure 14.
Geometry of brood defense in convict cichlids in Río Cabuyo that incorporates data on parental attack distances, brood radius data and the trade-off for fry between maximizing foraging while minimizing risk of predation. A, zone of intimidation where physical presence of parents deters approach of brood predators. Very small fry from Río Cabuyo stay here and small and medium-sized fry from Laguna de Xiloá stay here; B, transition zone where fry can expand into when their swimming performance and predation risk warrants it; C, zone of interception to maintain a predator-free buffer around the brood; D, zone of escalated attack, particularly for juveniles of the piscivore P. dovii. The evolution of parental care in cichlids is a three-way process between brood predators, ontogeny of antipredator competence in the fry, and parental area defense.
Figure 14.
Geometry of brood defense in convict cichlids in Río Cabuyo that incorporates data on parental attack distances, brood radius data and the trade-off for fry between maximizing foraging while minimizing risk of predation. A, zone of intimidation where physical presence of parents deters approach of brood predators. Very small fry from Río Cabuyo stay here and small and medium-sized fry from Laguna de Xiloá stay here; B, transition zone where fry can expand into when their swimming performance and predation risk warrants it; C, zone of interception to maintain a predator-free buffer around the brood; D, zone of escalated attack, particularly for juveniles of the piscivore P. dovii. The evolution of parental care in cichlids is a three-way process between brood predators, ontogeny of antipredator competence in the fry, and parental area defense.

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wisenden, B.D. Effect of Predation on Shaping Parental Brood Defense and Larval Ontogeny of Convict Cichlids Leading to Population Divergence. Diversity 2020, 12, 136. https://0-doi-org.brum.beds.ac.uk/10.3390/d12040136
AMA Style
Wisenden BD. Effect of Predation on Shaping Parental Brood Defense and Larval Ontogeny of Convict Cichlids Leading to Population Divergence. Diversity. 2020; 12(4):136. https://0-doi-org.brum.beds.ac.uk/10.3390/d12040136
Chicago/Turabian StyleWisenden, Brian D. 2020. "Effect of Predation on Shaping Parental Brood Defense and Larval Ontogeny of Convict Cichlids Leading to Population Divergence" Diversity 12, no. 4: 136. https://0-doi-org.brum.beds.ac.uk/10.3390/d12040136
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.