Diversity and Origin of the Central Mexican Alpine Flora
by

Victor W. Steinmann
1, 
Libertad Arredondo-Amezcua
2,
Rodrigo Alejandro Hernández-Cárdenas
3 and
Yocupitzia Ramírez-Amezcua
2,* 1
Facultad de Ciencias Naturales, Universidad Autónoma de Querétaro, Av. de las Ciencias s/n, Del. Sta. Rosa Jáuregui, Querétaro 76230, Mexico
2
Private Practice, Pátzcuaro, Michoacán 61600, Mexico
3
Herbario Metropolitano, División de Ciencias Biológicas y de la Salud, Departamento de Biología, Universidad Autónoma Metropolitana-Iztapalapa, Avenida San Rafael Atlixco #186, Colonia Vicentina, Iztapalapa, Ciudad de México 09340, Mexico
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(1), 31; https://0-doi-org.brum.beds.ac.uk/10.3390/d13010031
Submission received: 16 November 2020
/
Revised: 21 December 2020
/
Accepted: 5 January 2021
/
Published: 15 January 2021
(This article belongs to the Special Issue Diversity, Ecology and Conservation of Alpine Plants)
Abstract
:Alpine vegetation is scarce in central Mexico (≈150 km2) and occurs on the 11 highest peaks of the Trans-Mexican Volcanic Belt (TMVB). Timberline occurs at (3700) 3900 m, and at 4750 m vascular plants cease to exist. The alpine vascular flora comprises 237 species from 46 families and 130 genera. Asteraceae (44), Poaceae (42), and Caryophyllaceae (21) possess 45% of the species; none of the remaining families have more than 10 species. Four species are strict endemics, and eight others are near endemics. Thirteen species are restricted to alpine vegetation but also occur outside the study area. Seventy-seven species are endemic to Mexico, 35 of which are endemic to the TMVB. In terms of biogeography, the strongest affinities are with Central or South America. Fifteen species are also native to the Old World. Size of the alpine area seems to not be the determining factor for its floristic diversity. Instead, the time since and extent of the last volcanic activity, in addition to the distance from other alpine islands, appear to be important factors affecting diversity. There is evidence for upward vegetational shifts having occurred during the last century.
1. Introduction
Mexico is characterized by the widespread presence of mountains, and landscapes throughout the country are enhanced by the scenic arrangement of countless peaks and valleys. There are four primary montane regions in the country: the Sierra Madre Oriental, the Sierra Madre Occidental, the Sierra Madre del Sur, and the Trans-Mexican Volcanic Belt (TMVB). Mexico’s mountainous terrain has long been recognized as playing a crucial role in the genesis of the country’s rich flora [1]. Depending on location, these mountains house an impressive assemblage of ecosystems, from sweltering lowland tropical rain forests and seasonally dry deciduous forest at lower elevations through foggy montane cloud forests on the upper slopes to a mosaic of temperate woodlands composed of junipers, firs, pines, and oaks at higher elevations [1]. At the summits of a handful of the highest peaks, the forests open to the frigid alpine ecosystem, a biome characterized by a reduced, fragmented distribution and extreme climatic conditions. It occurs in mountainous areas above the limit of continuous forest (timberline) and below the nival zone where vascular plant growth ceases [1,2]. The term alpine applies to any low-stature vegetation above the climatic timberline worldwide and is not restricted to the European Alps from where the name originated [3]. As a consequence of the harsh conditions they must endure, alpine plants are typically small and grow close to the ground and/or in dense clumps [3]. They also possess various adaptations to cold, such as the ability to metabolize at low temperatures, dormancy during the winter, clonal vegetative growth, relatively large underground root and/or rhizome systems which store carbohydrates throughout the winter, and perennating buds that lie at or below the ground surface [4].
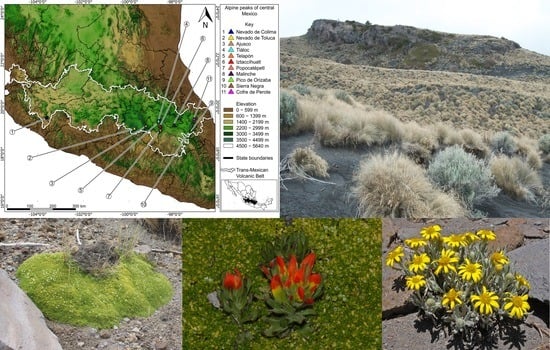
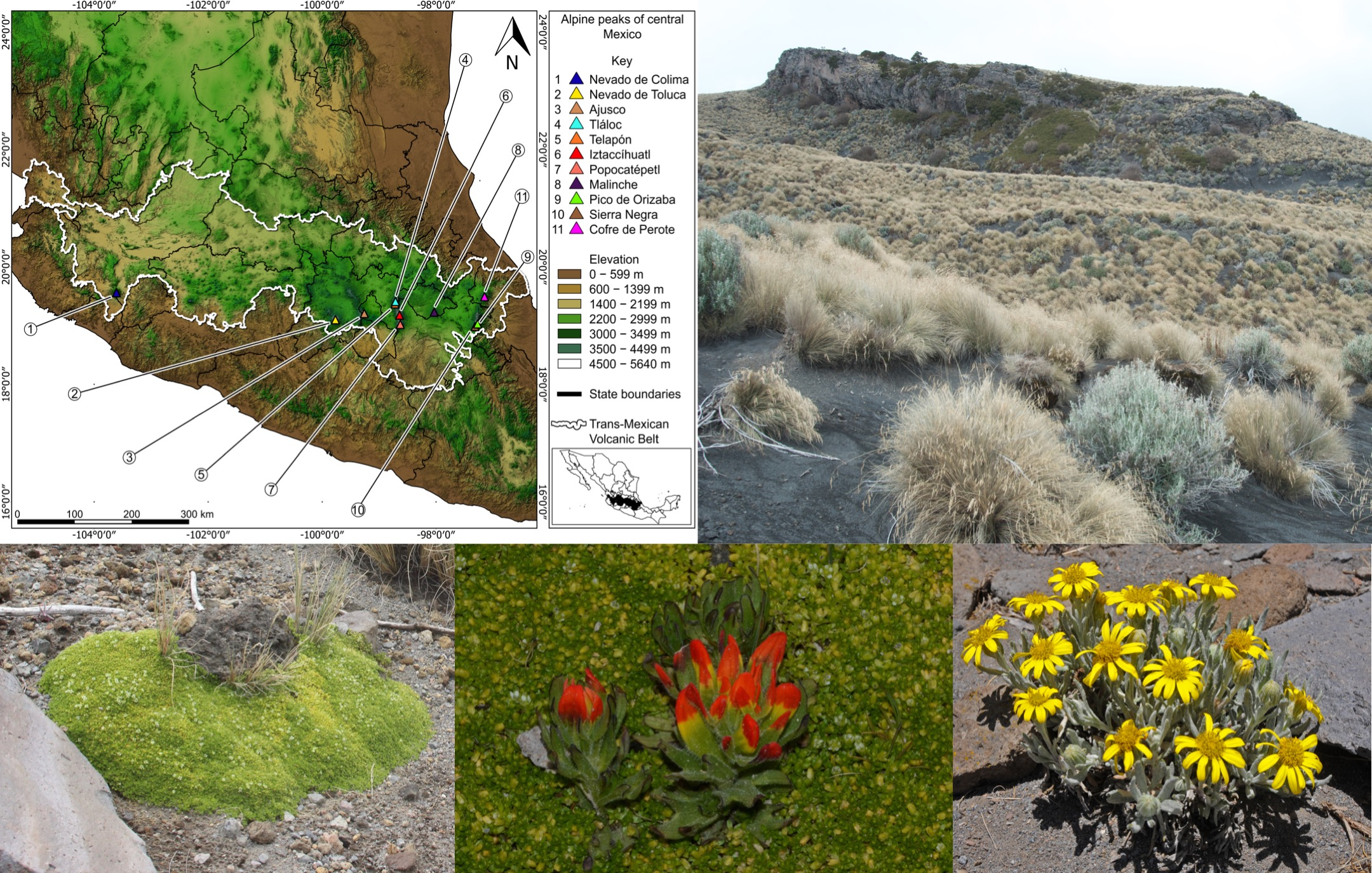
Alpine ecosystems occupy a mere 3% of the world’s land area [3]. This ecosystem is even more restricted in Mexico, and we estimate that less than 0.0001% of the country is covered by alpine vegetation. It is found on the summit of the Tacaná Volcano in Chiapas [5] and a handful of peaks along the Sierra Madre Oriental in the states of Coahuila, Nuevo León, and Tamaulipas [6,7,8]. However, the ecosystem reaches its greatest extent in the Trans-Mexican Volcanic Belt, an active volcanic zone that extends east-west across country from the state of Veracruz to the states of Jalisco and Nayarit [9]. Within this region, the alpine ecosystem forms an archipelago of small patches on the summits of only a handful of the highest mountains (Figure 1).
Mexican alpine regions have mean annual temperatures from 3 to 5 °C and can experience frosts throughout the year. Sporadic winter snowstorms also occur, and snowpack is present during part of the year [1]. Annual precipitation at the lower limits of alpine vegetation on the Nevado de Toluca (4120 m) is 1243 mm and concentrated during the summer monsoon months of May to October [10]. Following García [11], the climate is classified as E(T)H: cold, humid, and tundralike. These regions also experience low atmospheric pressure, substantial insolation, and strong winds, thus resulting in high evaporation [1]. In tropical alpine areas, such as those of central and southern Mexico, diurnal temperature variation can be more extreme than the variation throughout the year, and surface temperature fluctuations of 70 °C have been reported from the Pico de Orizaba [12]. This regular, drastic change in temperature poses additional challenges for tropical alpine plants [13] and has led to the characterization of tropical alpine ecosystems as having “summer every day, winter every night” [14,15].
The elevation of timberline varies throughout the world, depending primarily on latitude, with elevations decreasing towards the poles and increasing near the Equator [3]. The demarcation between forests and the alpine ecosystem is often well marked (Figure 2B). However, sometimes there is an ecotone where the forest gradually becomes less dense and opens to alpine vegetation with increasingly scattered and smaller trees. Even within the limited alpine areas of Mexico, there are many distinct plant communities and associations, often on a single volcano [8,10]. Figure 2 depicts some central Mexican alpine landscapes.
Although the alpine ecosystem is now restricted to a small number of high-elevation refuges, this has not always been so. During the Pleistocene (2.5 million–1700 BP), it underwent a series of expansions and contractions correlated with glacial and interglacial cycles [16,17]. In fact, during the last glacial period, the Wisconsin Glacial Episode (approximately 75,000–11,700 BP), vast expanses of northern and central Mexico were likely covered with alpine vegetation, and it is believed to have occurred at elevations 1000–1300 m lower than present [18]. Due to their association with extremely cold climates, alpine ecosystems throughout the world are greatly at risk from global warming [19,20,21,22,23,24], and those of Mexico are no exception. This was supported by ecological niche models of alpine endemic or near-endemic species from the TMVB [25], where clear reduction of suitable habitat in the future was predicted. Other threats besides climate change include tourism, livestock grazing, fire, and other anthropogenic activities like deforestation and construction of water-retaining ditches [26,27].
The floras of some of the central Mexican alpine peaks have been studied, e.g., Iztaccíhuatl, Popocatépetl, and the Nevado de Toluca [10]; La Malinche [28]; and the Cofre de Perote [29]. There is also a treatment of the alpine ferns of the TMVB [30]. However, most of the alpine summits and their plants have yet to be thoroughly explored, e.g., Pico de Orizaba, Nevado de Colima, and Tláloc. Furthermore, there is no synthesis on the central Mexican alpine flora in its entirety. Our current research documents the composition of the central Mexican alpine vegetation by investigating the local floras of the individual volcanoes. In addition, we analyze the floristic affinities of the central Mexican alpine flora in order to better understand its origin and the overall distributions of its species and provide a description of the plant communities occurring across the elevational gradient. Finally, we discuss endemism and mention species considered to have conservation concerns. Our primary objective is to provide baseline data that can be used for the management of this fragile ecosystem as the imminent threat of global warming becomes a reality.
2. Materials and Methods
Eleven volcanoes in central Mexico have alpine vegetation at their summits, and all of them lie within the TMVB (Figure 1, Table 1). These are the Nevado de Colima, Nevado de Toluca, El Ajusco, Iztaccíhuatl, Popocatépetl, Telapón, Tláloc, La Malinche, Pico de Orizaba (including Cerro Chichimeco), Sierra Negra, and Cofre de Perote. General information about the geology, natural history, and accessibility of these volcanoes can be found in Neyra [31]. We followed Beaman’s [2] concept of timberline, and only open areas above timberline are included; grasslands and meadows occurring at lower elevations in openings within otherwise continuous forest were not considered. In general, the alpine vegetation was restricted to elevations of 3900 m and higher, with the only notable exception being El Ajusco. This volcano is remarkable among those of the TMVB because timberline occurs at approximately 3700 m, or about 200 m lower than on the other volcanoes. We are uncertain as to the causes of this anomaly.
A preliminary floristic list was prepared using different treatments, e.g., La Vegetación del Cofre de Perote [29], Flora Novo-Galiciana [32], Flora Fanerogámica de Valle de México [33], Biodiversidad del Parque Nacional La Malinche [28], Vegetación Alpina de los Volcanes Popocatépetl, Iztaccíhuatl y Nevado de Toluca [10], and some fascicles of Flora de Veracruz were also consulted, such as Grossulariaceae [34] and Gentianaceae [35], to name a few. The holdings of the following herbaria were thoroughly examined: ENCB, FCME, IBUG, IEB, MEXU, MSC, UAMIZ, and XAL (acronyms according to Thiers [36]). A few specimens from ASU, NY, POM, and US were also included. In addition, bimonthly field trips were conducted during two years (September 2012 to August 2014) to make observations on the plant communities and collect material for the floristic study; sporadic collecting expeditions have occurred since then, and all peaks were visited numerous times throughout the year. The collections were made and processed following the specimen preparation techniques detailed by Lot and Chiang [37]. The majority of the vouchers are deposited in IEB, but some are in MEXU or QMEX. Observations on iNaturalist were examined in November 2020. Material was determined using various taxonomy treatments, as well as comparing the specimens with previously identified material in the herbaria IEB and MEXU.
Classification and species circumscription of the ferns follow Mickel and Smith [38], Sigel et al. [39], and PPG I [40]. Familial classification of the gymnosperms and angiosperms is based on APG IV [41]. An annotated checklist of all taxa was prepared, and here we present one voucher specimen from each of the volcanoes where it occurs. The overall distributions of the species were determined by the examination of pertinent literature and herbarium collections, as well as the records available through the Global Biodiversity Information Facility [42] (gbif.org) and other electronic resources such as Tropicos [43] (tropicos.org) and iNaturalist [44] (inaturalist.org). Adventive taxa are indicated as so. Synonyms are included only when the name has been applied frequently to plants from the study area within the last few decades. Species that have been reported from the area but not confirmed by our collections or herbarium specimens are listed in a section of excluded species. Growth forms were characterized from the examination of herbarium specimens, as well as plants in the field, using the following general categories: annual (herbaceous plant not surviving more than a single growing season), perennial herb (herbaceous plant persisting various years), subshrub/shrub (plant woody at least towards the base, often branched at the base, and either less than 5 m tall and/or having a primary axis with a diameter of less than 10 cm at 1.3 m from ground level), and tree (woody plant 5 m tall or more, with a primary axis having a diameter of more than 10 cm at 1.3 m from ground level). In addition, particular habitat preferences such as aquatic, palustrine, and saxicolous are given for some species. A search was conducted to determine which species are listed in the International Union for Conservation of Nature Red List [45] (iucnredlist.org) and the official Mexican standard which evaluates and regulates wild species risk of extinction “NOM-059-SEMARNAT-2010” [46]. The extent of alpine vegetation at each site was calculated using images from Google Earth [47] (google.com/earth) and coupled with field observations.
3. Results
3.1. Diversity and Community Structure
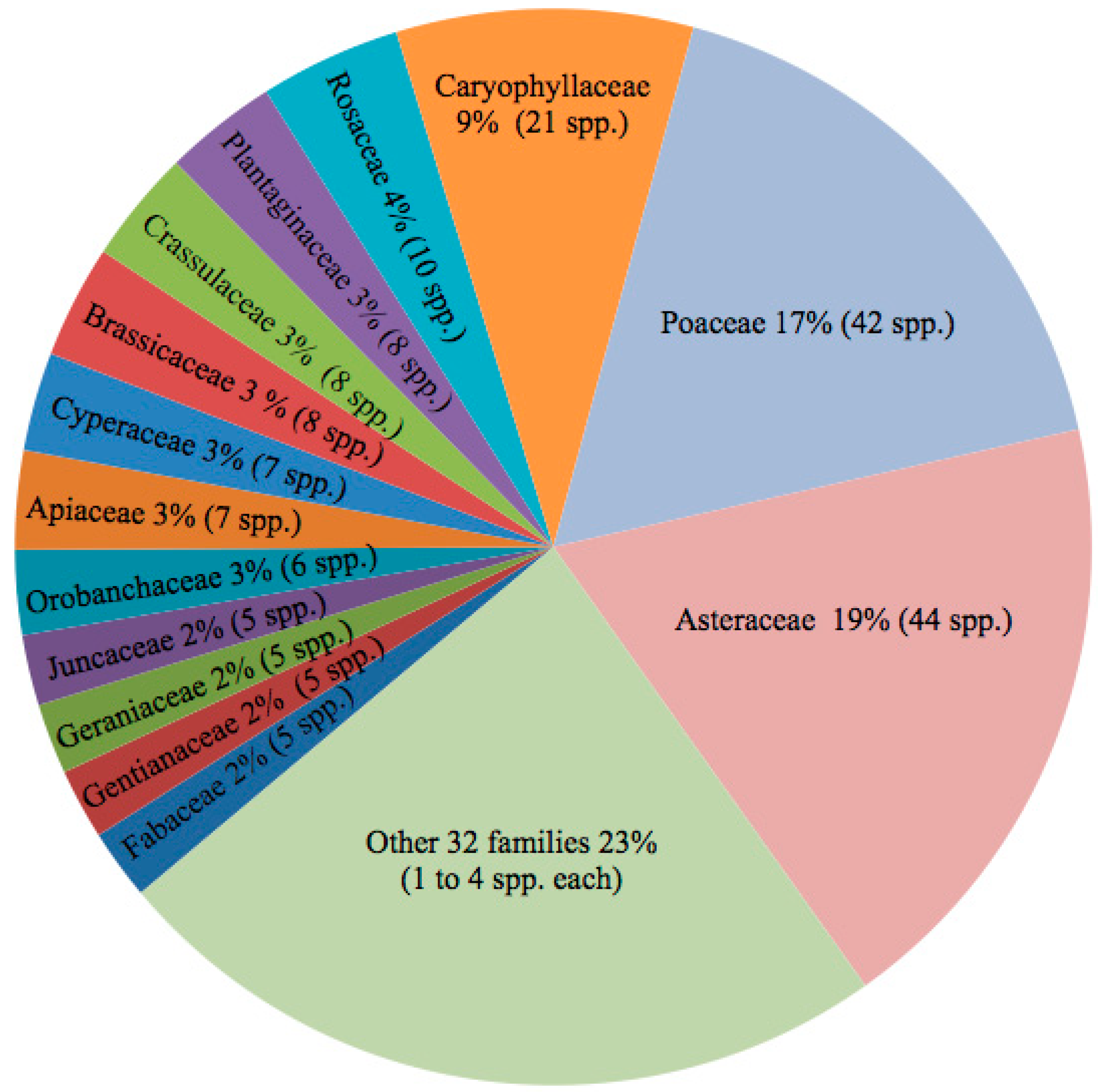
Between 2012 and 2018, approximately 1500 collections were made, many of these with one to several duplicates. An additional 2200 previously collected herbarium specimens were examined, primarily from ENCB, MEXU, and MSC, and nearly 450 observations on iNaturalist were also reviewed. We document 46 families, 130 genera, and 237 species of vascular plants (Table A1). More detailed information about distribution, collectors, vouchers from the different volcanoes (including herbarium) and in some cases synonyms, ecological preference and/or conservation status can be consulted in the Supplementary Material. Some representative species are depicted in Figure 3. The ferns consist of five families, 11 genera, and 12 species, whereas there are two families, three genera, and three species of gymnosperms. The remaining 39 families, 116 genera, and 222 species are angiosperms. The most diverse families are Asteraceae (44 spp.), Poaceae (42), and Caryophyllaceae (21) (see Figure 4). The most speciose genera are Muhlenbergia (8), Cerastium (8, including one introduced), Senecio (7, one introduced), Arenaria (7), Poa (7, two introduced), and Carex (6). The remaining genera are all represented by five species or fewer, and 19 families and 85 genera have a single species in the region. One family, six genera, and 12 species are introduced.
We were unable to verify by fieldwork, herbarium studies, or iNaturalist observations the presence of 25 species that were previously reported from the study area. These are listed in Appendix B. Some are misapplications, e.g., Berberis schiedeana Schltdl. [48], whereas others are unlikely to actually be present, e.g., Pinus montezumae Lamb. In some instances, the taxa may occur in the area, but we have not been able to verify their presence.
The size of the alpine sites varies greatly, with the largest being the Pico de Orizaba (48.83 km2) and the smallest being Telapón (0.03 km2); in total, there are only about 150 km2 of alpine vegetation in central Mexico (Table 1). The diversity among the alpine peaks is greatly disparate (Table A1 and Table 1). It ranges from 170 species on Iztaccíhuatl to 31 species on the Nevado de Colima. Only nine species are found on all of the volcanoes: Pseudognaphalium liebmannii, Draba jorullensis, Juniperus monticola fo. compacta, Lupinus montanus, Luzula racemosa, Pinus hartwegii, Agrostis tolucensis, Trisetum spicatum, and Alchemilla vulcanica. In contrast, 68 (32%) of the native species occur in a single alpine area. Thirty-six of these are restricted to Iztaccíhuatl, and all of the areas have at least one species not shared with any other alpine region (Table A1).
Diversity and structure change along the gradient from lower to upper alpine zones on the peaks that have enough elevational variation to exhibit such trends. Although the composition depends on the plants of the individual site, the patterns are similar throughout central Mexico. Timberline occurs at approximately 3900 to 4000 m with the exception of El Ajusco, where it is at about 3700 m. Just above timberline, it is common to encounter scattered trees of Pinus hartwegii at all of the alpine sites. The only other tree in the region is Salix paradoxa; rare individuals have been found on El Ajusco, but it is not known elsewhere in the study area. A single individual of Abies religiosa was found on the Sierra Negra at 4271 m; the plant was only two meters tall being a young, nonreproductive individual isolated outside its common distribution. The tree line or tree limit, the elevation at which there are trees of P. hartwegii is around 4200 m, and the tree species limit, the elevation at which there are juvenile individuals of P. hartwegii, is around 4550 m.
The lower alpine areas are dominated by a dense assemblage of tussock grasses, and the most common of these are Muhlenbergia quadridentata, Calamagrostis orizabae, C. tolucensis, and Festuca tolucensis. Other characteristic species occurring in the lower tussock grasslands include Eryngium proteiflorum, Cirsium nivale, Laennecia schiedeana, Pseudognaphalium liebmannii, Robinsonecio gerberifolius, Senecio mairetianus, S. procumbens, S. roseus, Phacelia platycarpa, Draba jorullensis, D. nivicola, Arenaria bryoides, A. oresbia, A. reptans, Cerastium orintales, C. ramigerum, Polysticum speciosissimum, Gaultheria myrsinoides, Vaccinium caespitosum, Lupinus montanus, Luzula racemosa, Calandrinia acaulis, Castilleja tolucensis, Penstemon gentianoides, Agrostis tolucensis, Festuca hephaestophila, Poa calycina, Tristeum spicatum, Alchemilla vulcanica, Potentilla richardii, and Cystopteris fragilis.
On the rocky outcrops and cliffs that are common throughout the study area, Berberis alpina, Juniperus monticola fo. compacta, Sedum goldmanii, Echeveria secunda, and Chionolaena lavandulifolia are frequent. Wetlands are scarce in the alpine vegetation of central Mexico, but they do occur on some of the peaks. The crater of the Nevado de Toluca has two lakes, and various seeps are found on Iztaccíhuatl. Aquatic and semiaquatic herbs are Elatine brachysperma, Eleocharis acicularis, Ranunculus dorianus, Colobanthus quitensis, Cotula mexicana, Gentiana ovatiloba, Luzula caricina, Plantago tubulosa, and Muhlenbergia orophila.
Almost all of the species occur between 3900–4200 m. This includes the endemic species which first appear just above timberline. From 4200 to 4500 m, there is considerable reduction in the species present, and the vegetation also becomes sparser, with the tussock grasses being replaced by smaller caespitose grasses and an abundance of low-growing forbs. Woody plants can extend up to about 4550 m, but most reach their limit at approximately 4300 m. Shrubs occurring at this elevation include Berberis alpina, Chionolaena lavandulifolia, Juniperus monticola fo. compacta, Oxylobus arbutifolius, Ribes ciliatum, and Senecio cinerarioides. Only about 10% of the alpine species occur above 4500 m, and almost all of these are herbaceous perennials: Alchemilla vulcanica, Arenaria bryoides, Castilleja tolucensis, Cerastium ramigerum, Chionolaena lavandulifolia, Draba jorullensis, D. nivicola, Festuca tolucensis, Lupinus montanus, Luzula racemosa, Plantago tolucensis, Pseudognaphalium liebmannii, Robinsonecio gerberifolius, Senecio procumbens, and S. roseus. However, on the Pico de Orizaba, an individual of Pinus hartwegii was found at 4550 m.
Between 4700 and 4750 m, plants are scattered and infrequent, and species found there are Agrostis tolucensis, Calamagrostis schiedeana, Draba nivicola, Festuca livida, Poa conglomerata, and Trisetum spicatum. Vascular plant growth ceases at about 4750 m in central Mexico, and only three volcanoes reach this elevation: Iztaccíhuatl, Popocatépetl, and Pico de Orizaba. Above this, the landscape is composed of glaciers and barren rocky slopes. Due to an outbreak of volcanic activity during the last twenty years, we were unable to explore the upper portions of Popocatépetl where certainly much of the upper alpine vegetation has been impacted by the frequent eruptions.
3.2. Endemism
The Mexican alpine flora includes 76 species (32%) that are exclusive to the country, with 36 of these (15%) restricted to the TMVB (Table 2). The only genus endemic to the country is Mexerion (Table 3). There are four species known only from the study area: Castilleja tolucensis, Cerastium purpusii, Draba nivicola, and Plantago tolucensis. None of these are limited to a single volcano, but Cerastium purpusii is restricted to Iztaccíhuatl and the Nevado de Toluca. Draba nivicola is the most widespread of the alpine endemics, occurring on nine volcanoes. Castilleja tolucensis is found on seven volcanoes, and Plantago tolucensis on six. Iztaccíhuatl is the only area to have all the endemic alpine species. In contrast, El Ajusco and the Nevado de Colima lack endemics. In addition to the four strict endemics, there are eight near-endemic species that occur primarily in the alpine vegetation but extend into adjacent coniferous forests at elevations above 3500 m. These include Tauschia alpina (lower elevation of 3600 m), Chaerophyllum tolucanum (3900 m), Cerastium orithales (3600 m), Cerastium ramigerum (3600 m), Carex hermannii (3600 m), Gentiana perpusilla (3500 m), Agrostis calderoniae (3600 m), and Potentilla richardii (3600 m).
3.3. Phytogeography
The general distributions of all species are provided in Table A1, and some more detailed distributions are in the Supplementary Material. Species restricted to alpine vegetation but ranging beyond central Mexico include: Chionolaena lavandulifolia (Tacaná Volcano and Central America), Gamochaeta standleyii (Guatemala), Robinsonecio gerberifolius (Guatemala), Arenaria bryoides (Tacaná Volcano and Central America), Arenaria parvifolia (South America), Luzula vulcanica (South America), Plantago tubulosa (South America), Festuca livida (Guatemala), Poa calycina var. mathewsii (South America), Poa chamaeclinos (South America), Poa gymnantha (South America), and Alchemilla pinnata (Guatemala and South America).
The vast majority of the species that inhabit the alpine vegetation of central Mexico have broad ranges that extend outside of alpine vegetation and/or the country (Table A1 and Table 2). Nearly 80% of the species are found below 3000 m in central Mexico. With regard to distribution, 41 species are shared between Mexico and Guatemala, and another 15 extend farther into Central America. An additional 29 species reach South America, and nine species are disjunct between Mexico and South America. There are considerably fewer species shared with the United States and/or Canada. Only nine species are restricted to Mexico and Canada and/or the United States, whereas 20 more range from Canada and/or the United States through Mexico to Central or South America. Finally, it is worth noting that fifteen species also occur natively in the Old World.
3.4. Growth Forms
In the Mexican alpine vegetation, perennial herbs are the dominant life form (85%), shrubs are uncommon (5%), and annuals (8.7%) and trees (1.3%) are rare (Table A1). The majority of the plants are autotrophs, but there are six root hemiparasites from the Orobanchaceae family, and the single saprophyte is the locally rare Monotropa hypopitys.
3.5. Conservation Status
Only 19 of the 225 native species have been evaluated for the IUCN Red List, and all of these are listed as being of Least Concern (LC). With regard to the NOM-059-SEMARNAT-2010, eight species are included, five as protected (Pr) and three as threatened (A): Arenaria bryoides (Pr), Commarostaphylis discolor (Pr), Draba nivicola (A), Juniperus monticola (Pr), Monotropa hypopitys (Pr), Eryngium proteiflorum (Pr), Castilleja tolucensis (A), and Calamagrostis eriantha (A). Threatened species are those considered to be in short- or medium-term risk of extinction if current adverse factors continue, and protected species are those that could become threatened and for which conservation or recuperation strategies are needed.
4. Discussion
4.1. Diversity
The native alpine diversity is relatively low in comparison with the overall diversity of Mexican vascular plants, 225 species of an estimated 23,314 [49], and in comparison with the diversity encountered in floristic studies of other ecosystems of Mexico. Several contributing factors should be considered, such as the miniscule area (0.0001%) of alpine vegetation within Mexico, its current geographic isolation, young geological history, and extreme environmental conditions. Furthermore, according to Körner [3], the floristic diversity of alpine areas extending up to about 100 km2 includes around 300 species, and the number does not increase substantially until complete mountain ranges are considered. Poaceae, Asteraceae, and Caryophyllaceae account for 45% of the species, and these families also dominate in terms of relative abundance in the field. All of these are dominant in alpine areas throughout the world [50,51,52]. Of the remaining families, none are represented by more than 10 species, and 20 have a single species.
The composition of these alpine islands is atypical considering the floristic composition of Mexico. Highly speciose families such as the Orchidaceae, Euphorbiaceae, and Cactaceae are not represented, an expected result considering that these are primarily warm tropical in distribution. However, it is worth noting that all of these families are present in the alpine vegetation of South America. The case of Cactaceae is particularly noteworthy because the family occurs as high as 4830 m in Peru [53], but in Mexico, which is the center of diversity of the family, no species surpasses 3200 m. Other diverse families of the Mexican flora but little represented in the alpine vegetation include Rubiaceae (1 sp.) and Fabaceae (5 spp.). Again, these are families that reach their greatest diversity in the warm tropics but do have various taxa in the alpine vegetation of South America [51]. Lamiaceae, which is primarily a northern hemisphere temperate family, is also not diverse in the area, and only two native species are present. One of these, Salvia prunelloides, is known from a single collection on El Ajusco, whereas the other, Stachys eriantha, is widespread and abundant.
At the rank of genus, the alpine vegetation is also atypical for Mexico. Of the ten most diverse genera in the country, only Muhlenbergia is found in our alpine flora, and it ties for the most diverse genus in the area, with eight species. While present and sometimes dominant in the alpine vegetation of tropical America, Muhlenbergia is not particularly common in alpine areas of other parts of the world. Additional diverse central Mexican alpine genera include Cerastium (8 spp.), Arenaria (7), Senecio (7), Agrostis (6), and Carex (6), and most of these are well represented in alpine areas of both the Old and New World. Other characteristic alpine genera include Alchemilla (5), Festuca (5), and Ranunculus (4). Interestingly, 89 genera (74%) have a single species in the area.
Within the study area, complex and still ambiguous factors affect the number of species present on the different peaks, and Pleistocene expansions and retractions certainly have influenced distributions. Diversity is often closely correlated with size, but that is not so for the peaks studied here. For example, the largest and highest alpine island on the Pico de Orizaba surprisingly has less species than the nearby Cofre de Perote, which is nearly 30 times smaller and considerably lower. The alpine areas of Iztaccíhuatl and Popocatépetl are about the same size, same elevation, and adjacent to one another, but Iztaccíhuatl has twice as many species as Popocatépetl. Instead, an important factor affecting alpine diversity appears to be the time since and extent of the last volcanic activity. For example, the last eruption of the Pico de Orizaba was in 1846, whereas the Cofre de Perote has lain dormant for nearly 900 years. Likewise, before its recent and violent eruptions, Popocatépetl experienced moderate activity during the 20th century, whereas the last major eruptions on Iztaccíhuatl go back almost 9000 years.
The least diverse peak is the Nevado de Colima with only 31 species. Although small, its area is still larger than or comparable to Tláloc and Telapón, and both of these peaks have about twice as many species. It is worth noting that the alpine moss flora of the Nevado de Colima is similarly depauperate in comparison with other alpine regions of central Mexico [54], and explanations for this discrepancy also apply to the vascular flora and include that the northern slope of the Nevado de Colima has generally sandy, unstable substrates, whereas the southern and western slopes have been affected by recent eruptions of the adjacent Volcán de Fuego. Weather patterns are distinctly drier, and it is isolated by more than 400 km from the closest alpine peak, the Nevado de Toluca.
The trend is for diversity to decrease from lower to higher alpine elevations [55], and that tendency also occurs in central Mexico. Almost all species are present between 3900–4200 m, but they gradually decrease with elevation to the point that only 10% occur at approximately 4500 m. Vascular plant growth in other areas of the Americas can exceed 5000 m [53], and in the Himalayas, it can reach more than 6000 m [56,57]. However, the upper limit of vascular plant growth occurs at 4750 m in central Mexico, and only about six species grow at this elevation.
4.2. Endemism
Only 37% of the central Mexican alpine plants are endemic to Mexico, and this percentage is considerably lower than the overall level of endemism for the Mexican flora, which is calculated at 50% [49]. Alpine regions are generally characterized by high levels of endemism [3,51,58,59]. In central Mexico, it is not so. No single peak has an alpine endemic, and only four species (2%) are endemic to the vegetation as a whole. This number ascends to 12 (5.5%) when considering restricted species that occur slightly outside the area in the adjacent subalpine forest. The presence of few endemic species may be related to the relatively young age of the high elevations in Central Mexico, and Ferrari [60] states that high elevation stratovolcanoes have only formed during the last one million years. Most of the volcanoes have been active during the last 30,000 years, and catastrophic eruptions at the high elevations may limit the evolution of an endemic component.
4.3. Introduced Taxa
The twelve introduced species are similarly concentrated in the three most diverse families: Poaceae (4 spp.), Caryophyllaceae (3), and Asteraceae (2). Geraniaceae, Lamiaceae, and Polygonaceae each have a single species. All of these species are “weedy” Old World taxa that have become widely naturalized in the New World, with the exception of Poa pratensis, a complex taxon distributed natively in both the Old and New World. Our plants belong to var. pratensis, which is introduced in Mexico [61]. The most widespread naturalized species in the area is Taraxacum officinale, which has been found at five of the alpine sites. Nine of the naturalized species have been documented from a single alpine peak. At present, none of the taxa are particularly invasive nor pose a threat to the native taxa, and most occur in disturbed areas adjacent to roads or trails. However, we expect that in the future, as temperatures rise, the presence and abundance of introduced species will increase.
4.4. Growth Forms
The dominance of perennial herbs reported here follows a well-known tendency in alpine floras [56,62,63]. It is also worth noting that epiphytes and climbers are lacking. Parasitism occurs only in the six species of the root hemiparasitic family Orobanchaceae. The stem parasite Arceuthobium globosum Hawksw. & Wiens (Santalaceae) has been documented on Pinus hartwegii in adjacent subalpine forest just below timberline on the Nevado de Toluca, but it has yet to be encountered in true alpine vegetation. Our single species of saprophyte, Monotropa hypopitys, is rare in the area and known from a single collection made on Iztaccíhuatl in 1965. Giant rosette plants are characteristic of tropical alpine vegetation throughout the world [64], but these are absent from central Mexican alpine areas despite their common occurrence at lower elevations in the region. Another characteristic of alpine regions, both temperate and tropical, is the prevalence of cushion plants, which are compact, low-growing, and mat-forming [65]. In our region, this habit is rare, and the only true cushion species is Arenaria bryoides (Figure 2G). However, it is interesting to note that whereas in other alpine islands in central Mexico, A. oresbia and A. reptans form loose mats, on the sandbanks of the Nevado de Colima, they form cushions similar to those of A. bryoides. The cushion growth, which represents an efficient trap for heat and water, is associated with extreme environmental conditions [65], such as those that occur in the sandbanks of the Nevado de Colima, where the soil is poor in nutrients and there is a high exposure to ultraviolet radiation, strong winds, and desiccation.
4.5. Phytogeography
Mexico is of special biographical interest due to its unique position between the Nearctic and Neotropical kingdoms, and there are strong affinities with both of these regions [66,67]. With regard to the generic alpine flora of central Mexico, 106 genera (82%) occur in both the Old and New Worlds, and 24 genera (18%) are restricted to the Americas (Table 3). Only one genus is endemic to Mexico, Mexerion. Two occur from the United States to Mexico, and 12 occur from Mexico to Central or South America. Nine are found from the United States and/or Canada through Mexico to Central and/or South America.
The species-level phytogeography is described best by a single word: disjunction. Although some species do occur in nearly continuous extensions across central Mexico, many have interesting disjunctions, and there is a complex mosaic of disjunct species of different origin. All of the narrowly endemic and near-endemic species are isolated by the extensive expanses of coniferous forests that separate the peaks. There are also interesting disjunctions from far-off alpine regions, both to the north and south. Especially noteworthy is Colobanthus quitensis, one of only two species of vascular plants native to the Antarctic. There are isolated populations on the Pico de Orizaba, Cofre de Perote, and Iztaccíhuatl, with the nearest occurrences nearly 3000 km away in northern South America. Other notable alpine central Mexico–South American disjunctions include Calamagrostis rigescens, Luzula vulcanica, Poa calycina, P. chamaeclinos, and P. gymnantha. In Mexico, the latter two are known only from small areas on Iztaccíhuatl and Telapón respectively; however, they are frequent in the high Andes of South America [61]. Plantago tubulosa and Achemilla pinnata have similar disjunct distributions but also occur in alpine vegetation of Guatemala. The central Mexican alpine vegetation is also the southern limit for some circumboreal, alpine or Artic species including Stellaria umbellata and Sibbaldia procumbens, which are known in Mexico from only the study area. Probably the most peculiar of all the disjunctions is that of Cardamine obliqua, a species shared between the high-elevation mountains of central Mexico and those of eastern Africa. Our plants have been treated as a separate variety, and it remains to be confirmed by modern phylogenetic studies if these incredibly disjunct plants indeed form single lineage or represent a strange case of morphological convergence. Finally, it is worth mentioning some species that are broadly disjunct throughout the globe, such as Cystopteris fragilis and Trisetum spicatum. Much work remains to evaluate the origin and phylogeography of the alpine plants, but most of the widely spaced disjunctions have arrived by long distance dispersal, especially those species that are widespread in the Andes.
Sarmiento [68] mentioned that the affinities of the central Mexican highlands are with the north, whereas Rzedowski [69] concluded that the alpine grasslands of Mexico have strong affinities with the South American flora. Our results support the latter. Both in terms of genera and species, there are clearly more taxa shared with Central and South America than with the United States and Canada. In fact, more central Mexican alpine species occur in Peru and Bolivia than in the western United States, despite the latter countries being considerably farther away. These finding mirror those of Delgadillo [54] who likewise determined that the alpine moss flora of the Nevado de Colima, although composed of a broad assortment of phytogeographic elements, had its strongest affinities with Meso and South America.
4.6. Conservation
Alpine ecosystems in general are considered threatened by global warming [22,70], and those of central Mexico also are subject to additional anthropogenic pressures such as tourism, livestock grazing, induced burning, aforestation, and the construction of water-retaining ditches [26]. There is no comprehensive assessment of the conservation status of the alpine plants of central Mexico. In fact, only 19 of the 237 species have been evaluated for inclusion in the IUCN Red List. Most of these are relatively widespread taxa, and all of them were determined to be of Least Concern (LC). The alternative “Norma Oficial Mexicana NOM-059” [46] is the only official document used by Mexican governmental and environmental agencies in matters concerning rare and endangered species. It lists three species as threatened (A) and five species as protected (Pr). However, these lists certainly are not an accurate representation of the endangered taxa, and many more species are at risk than indicated, at least regionally. For example, the South American disjunct alpine endemics are all restricted, some being known from one or two areas and most being represented by less than 10 collections. The circumboreal Sibbaldia procumbens and Stellaria umbellata are similarly rare, and the latter is known in central Mexico from a single collection made in 1958. The strict endemics are projected to have much narrower distributions towards the end of the 21st century [25]. Additionally, some rare taxa previously documented from the area were not encountered during our surveys and are likely at risk, at least locally. These are Gentiana perpusilla, Stellaria umbellata, Sedum clavifolium, Agrostis calderoniae, Poa chamaechilnos, P. gymnantha, and Viola hemsleyana.
4.7. Upward Vegetational Shifts
Various recent upward migrations of alpine plants have been documented [57,58,71,72], and this phenomenon is both evidence for and the result of global warming. A study from 130 years ago [73] provides a detailed description of the vegetation occurring on five of the alpine peaks in central Mexico. The highest limit of vascular plants was given as 4572 m, and six species were reported to reach this elevation: Arenaria bryoides, Chionolaena lavandulacea, Senecio procumbens, Robinsonecio gerberifolius, Castilleja tolucensis, and Draba jorullenis. The perpetual snow line was reported to occur near this elevation on the Pico de Orizaba. The author also documents the upper limit of Pinus hartwegii on the Pico de Orizaba to be at 4267 m and slightly higher on the adjacent Sierra Negra, 4358 m. In contrast, our studies document 21 species occurring above 4500 m, with six species of vascular plants occurring at 4750 m. The perpetual snow line is also considerably higher now, at above 5000 m. With regard to Pinus hartwegii, it occurs as high as 4550 m on the Pico de Orizaba. In fact, scattered, sporadic individuals are frequent up to 4400 m on many of the peaks. The colonization of pines to higher elevations appears to be recent because the upper limit populations are composed of healthy, juvenile individuals with never any older or deceased plants. Our report of Abies religiosa from 4271 m also represents an elevational extension for this common tree which forms dense forests below the alpine zone, and Heilprin [73] listed its maximum elevation at 3650 m. There is also evidence that one of the alpine endemics, Castilleja tolucensis, is no longer present at lower elevations in Iztaccíhuatl where it was previously documented to be abundant. It was found in 1965 at 3800 m (Rzedowski 20139, ENCB) on Iztaccíhuatl. However, searches at the collection site failed to find it below 4000 m, and it is scarce at this elevation. All the aforementioned examples suggest that, like on other high mountains of the world, upwards vegetation shifts have already occurred on the alpine peaks of central Mexico during the past century.
5. Conclusions
The alpine flora of central Mexico is a unique assemblage of taxa with distinct biogeographic and evolutionary histories. Knowing species composition is just the first step, and our intention is to stimulate research on and conservation of the central Mexican alpine vegetation. There is still much to be accomplished before a thorough understanding of this ecosystem is attained. In particular, it is critical to conduct a formal conservation assessment of all species and, if necessary, implement programs for their survival. The highly specialized tropical alpine flora provides excellent opportunities for monitoring the effects of climate change, and long-term studies—in particular, the establishment of GLORIA plots [72]—are necessary to document population dynamics and taxa migrations and displacements. Phylogeographic studies are also important to understand the evolutionary histories and genetic diversity of species and populations inhabiting different volcanoes.
Supplementary Materials
The following is available online at https://0-www-mdpi-com.brum.beds.ac.uk/1424-2818/13/1/31/s1, Supplementary material: List of species, including growth form, overall distribution, occurrence on the different volcanoes, voucher information (including herbarium), and in some cases ecological preference and/or conservation status.
Author Contributions
Conceptualization, methodology, and funding acquisition, V.W.S. and Y.R.-A.; fieldwork, R.A.H.-C., L.A.-A., Y.R.-A. and V.W.S.; plant determination, R.A.H.-C., L.A.-A., Y.R.-A., and V.W.S.; data curation, R.A.H.-C. and L.A.-A.; estimation of sizes of different alpine areas, R.A.H.-C.; preparation of the manuscript, V.W.S., Y.R.-A., R.A.H.-C. and L.A.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), project JF076 “Flora Alpina del Centro de México”.
Data Availability Statement
Data is contained within the article and the supplementary material.
Acknowledgments
We thank the Natural Protected Areas of Pico de Orizaba, Iztaccíhuatl-Popocatépetl, La Malinche and Nevado de Colima, as well as the Nevado de Toluca Special Protection Area for permitting us to collect; the herbaria ASU, ENCB, FCME, IBUG, MEXU, MSC, NY, RSA-POM, QMEX, UAMIZ, and US kindly allowed us to consult their collections; Abraham Ornelas elaborated the map of the volcanoes, and Damián Piña Bedolla prepared Figure 2 and Figure 3; Adolfo López and Carlos A. Gutiérrez-Chávez graciously assisted with fieldwork and photography; Jerzy Rzedowski helped obtain important literature and offered meaningful discussions about the project; Socorro González and Anton Reznicek provided determinations of Carex (Cyperaceae); Armando Rincón and Gustavo Medrano collected material on El Ajusco and the Pico de Orizaba, respectively; Harald Pauli provided the photo of Luzula racemosa; and four anonymous reviewers gave many useful comments and corrections. Finally, we thank the special issue editors Juan Lorite and Manuela Winkler.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Species list of the central Mexican alpine flora. * = Endemic to the study area; + = introduced. Occurence on the different volcanoes: AJ = El Ajusco (63), CP = Cofre de Perote (93), IZ = Iztaccíhuatl (170), ML = La Malinche (62), NC = Nevado de Colima (31), NT = Nevado de Toluca (102), PO = Pico de Orizaba (78), PP = Popocatépetl (81), SN = Sierra Negra (46), TE = Telapón (57), TL = Tláloc (72). Habit is as follows: an = annual herb, ph = perennial herb, sh = shrub, sub = subshrub, tr = tree. Overall distribution: TMVB = Trans-Mexican Volcanic Belt. Voucher information can be consulted in the Supplementary material.
Table A1.
Species list of the central Mexican alpine flora. * = Endemic to the study area; + = introduced. Occurence on the different volcanoes: AJ = El Ajusco (63), CP = Cofre de Perote (93), IZ = Iztaccíhuatl (170), ML = La Malinche (62), NC = Nevado de Colima (31), NT = Nevado de Toluca (102), PO = Pico de Orizaba (78), PP = Popocatépetl (81), SN = Sierra Negra (46), TE = Telapón (57), TL = Tláloc (72). Habit is as follows: an = annual herb, ph = perennial herb, sh = shrub, sub = subshrub, tr = tree. Overall distribution: TMVB = Trans-Mexican Volcanic Belt. Voucher information can be consulted in the Supplementary material.
| Taxon | NC | NT | AJ | TL | TE | IZ | PP | ML | PO | SN | CP | Habit | Overall Distribution |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PTERIDOPHYTES | |||||||||||||
| ASPLENIACEAE | |||||||||||||
| Asplenium castaneum Schltdl. & Cham. | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Bolivia and Peru | |
| Asplenium fibrillosum Pringle & Davenp. | x | ph | endemic to central Mexico | ||||||||||
| DRYOPTERIDACEAE | |||||||||||||
| Elaphoglossum mathewsii (Fée) T. Moore | x | x | ph | Mexico to Bolivia and Chile | |||||||||
| Polystichum speciosissimum (A. Braun ex Kunze) Copel. | x | x | x | x | x | x | x | x | ph | central Mexico to Panama | |||
| POLYPODIACEAE | |||||||||||||
| Alansmia spathulata (A.R. Sm.) Moguel & M. Kessler | x | x | ph | Mexico to Guatemala | |||||||||
| Melpomene peruviana (Desv.) A.R. Sm. & R.C. Moran | x | x | x | ph | disjunct between Mexico and South America | ||||||||
| Pleopeltis polylepis (Roem. ex Kunze) T. Moore var. polylepis | x | ph | USA to Guatemala | ||||||||||
| Polypodium calirhiza S.A. Whitmore & A.R. Sm. | x | ph | western USA to central Mexico | ||||||||||
| PTERIDACEAE | |||||||||||||
| Gaga marginata (H.B.K.) F.W. Li & Windham | x | ph | Mexico to Bolivia and Argentina | ||||||||||
| Myriopteris lendigera (Cav.) J. Sm. | x | ph | southwestern USA to Ecuador and the Antilles | ||||||||||
| Pellaea ternifolia (Cav.) Link | x | ph | USA to Argentina, Chile, and the Antilles | ||||||||||
| WOODSIACEAE | |||||||||||||
| Cystopteris fragilis (L.) Bernh. | x | x | x | x | x | x | x | x | x | x | ph | Canada to Chile and Argentina, also native to the Old World | |
| GYMNOSPERMS | |||||||||||||
| CUPRESSACEAE | |||||||||||||
| Juniperus monticola Martínez fo. compacta Martínez | x | x | x | x | x | x | x | x | x | x | x | sh | endemic to Mexico |
| PINACEAE | |||||||||||||
| Abies religiosa (H.B.K.) Schltdl. & Cham. | x | tr | Mexico to Guatemala | ||||||||||
| Pinus hartwegii Lindl. | x | x | x | x | x | x | x | x | x | x | x | tr | Mexico to Honduras |
| ANGIOSPERMS | |||||||||||||
| APIACEAE | |||||||||||||
| Chaerophyllum orizabae (I.M. Johnst.) K.F. Chung | x | x | x | x | ph | endemic to the TMVB | |||||||
| Chaerophyllum tolucanum (I.M. Johnst.) K.F. Chung | x | x | x | x | x | ph | endemic to the TMVB | ||||||
| Eryngium carlinae F. Delaroche | x | ph | Mexico to Panama | ||||||||||
| Eryngium proteiflorum F. Delaroche | x | x | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||
| Lilaeopsis schaffneriana (Schltdl.) J.M. Coult. & Rose | x | ph | Mexico to Ecuador | ||||||||||
| Ottoa oenanthoides H.B.K. | x | ph | Mexico to Ecuador | ||||||||||
| Tauschia alpina (J.M. Coult. & Rose) Mathias | x | x | x | x | ph | endemic to the TMVB | |||||||
| ASTERACEAE | |||||||||||||
| Achillea millefolium L. | x | x | x | ph | nearly cosmopolitan in temperate regions | ||||||||
| Ageratina prunellifolia (H.B.K.) R.M. King & H. Rob. | x | x | x | x | ph | Mexico to Guatemala | |||||||
| Ageratina vernicosa (Sch. Bip. ex Greenm.) R.M. King & H. Rob. | x | sh | endemic to Mexico | ||||||||||
| Baccharis conferta H.B.K. | x | x | x | sh | endemic to Mexico | ||||||||
| Barkleyanthus salicifolius (H.B.K.) H. Rob & Brettell | x | x | sh | southwestern USA to Honduras | |||||||||
| Bidens anthemoides (DC.) Sherff | x | x | x | ph | endemic to Mexico | ||||||||
| Bidens triplinervia H.B.K. | x | x | x | x | x | x | x | x | x | sh | Mexico to Argentina | ||
| Chionolaena lavandulifolia (H.B.K.) Benth. & Hook. f. ex B.D. Jacks. | x | x | x | x | x | x | x | x | x | sub | Mexico to Central America | ||
| Chionolaena salicifolia (Bertol.) G.L. Nesom | x | sub | Mexico to Guatemala | ||||||||||
| Cirsium ehrenbergii Sch. Bip. | x | x | x | x | x | ph | endemic to Mexico | ||||||
| Cirsium jorullense (H.B.K.) Spreng. | x | ph | endemic to Mexico | ||||||||||
| Cirsium nivale (H.B.K.) Sch. Bip. | x | x | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||
| Conyza coronopifolia H.B.K. | x | an | Mexico to South America | ||||||||||
| Cotula mexicana (DC.) Cabrera | x | ph | Mexico to South America | ||||||||||
| Dahlia merckii Lehm. | x | ph | endemic to Mexico | ||||||||||
| Hymenoxys integrifolia (H.B.K.) Bierner | x | ph | Mexico to Guatemala | ||||||||||
| Erigeron galeottii (A. Gray) Greene | x | x | x | x | x | x | ph | endemic to Mexico | |||||
| Gamochaeta americana (Mill.) Wedd. | x | x | x | ph | Canada to South America and also the Antilles | ||||||||
| Gamochaeta purpurea (L.) Cabrera | x | ph | Canada to South America | ||||||||||
| Gamochaeta standleyi (Steyerm.) G.L. Nesom | x | x | x | x | ph | Mexico to Guatemala | |||||||
| Hieracium mexicanum Less. | x | x | x | x | x | x | ph | Mexico to Guatemala | |||||
| Laennecia schiedeana (Less.) G.L. Nesom | x | x | x | x | x | x | x | x | x | an/ph | USA to Panama | ||
| Mexerion sarmentosum (Klatt) G.L. Nesom | x | x | x | x | x | ph | endemic to the TMVB | ||||||
| Osbertia stolonifera (DC.) Greene | x | x | ph | Mexico to Guatemala | |||||||||
| Oxylobus arbutifolius (H.B.K.) A. Gray | x | x | x | x | x | x | x | x | x | ph/sub | Mexico to Guatemala | ||
| Packera bellidifolia (H.B.K.) W.A. Weber & Á. Löve | x | x | x | x | x | x | x | ph | endemic to Mexico | ||||
| Packera toluccana (DC.) W.A. Weber & Á. Löve | x | x | x | x | x | x | ph | endemic to Mexico | |||||
| Pseudognaphalium inornatum (DC.) Anderb. | x | ph | endemic to Mexico | ||||||||||
| Pseudognaphalium liebmannii (Sch. Bip. ex Klatt) Anderb. | x | x | x | x | x | x | x | x | x | x | x | an/ph | endemic to Mexico |
| Pseudognaphalium nubicola (I.M. Johnst.) Anderb. | x | x | ph | endemic to Mexico | |||||||||
| Pseudognaphalium oxyphyllum (DC.) Kirp. | x | x | x | ph | Mexico to Guatemala | ||||||||
| Robinsonecio gerberifolius (Sch. Bip. ex Hemsl.) T.M. Barkley & Janovec | x | x | x | x | x | x | ph | Mexico to Guatemala | |||||
| Roldana angulifolia (DC.) H. Rob. & Brettell | x | x | x | sh | endemic to Mexico | ||||||||
| Roldana barba-johannis (DC.) H. Rob. & Brettell | x | x | sh | Mexico to Guatemala | |||||||||
| Roldana candicans (Née) Villaseñor, S. Valencia & Coombes | x | x | ph | endemic to Mexico | |||||||||
| Roldana reticulata (DC.) H. Rob. & Brettell | x | ph | endemic to Mexico | ||||||||||
| Selloa plantaginea H.B.K. | x | x | ph | endemic to the TMVB | |||||||||
| Senecio callosus Sch. Bip. | x | x | x | ph | Mexico to Guatemala | ||||||||
| Senecio cinerarioides H.B.K. | x | x | x | x | sh | endemic to Mexico | |||||||
| Senecio mairetianus DC. | x | x | x | x | x | x | x | x | x | x | sh | Mexico to Guatemala | |
| Senecio procumbens H.B.K. | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||||
| Senecio roseus Sch. Bip. | x | x | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||
| +Senecio vulgaris L. | x | an | introduced from the Old World | ||||||||||
| +Taraxacum officinale F.H. Wigg. | x | x | x | x | x | ph | introduced from the Old World | ||||||
| BERBERIDACEAE | |||||||||||||
| Berberis alpina Zamudio | x | x | x | x | x | x | x | sh | endemic to central Mexico | ||||
| BORAGINACEAE | |||||||||||||
| Hackelia mexicana (Schltdl. & Cham.) I.M. Johnst. | x | ph | Mexico to Bolivia and Peru | ||||||||||
| Lithospermum distichum Ortega | x | x | ph | Mexico to Guatemala | |||||||||
| Phacelia heterophylla Pursh | x | x | ph | USA to central Mexico | |||||||||
| Phacelia platycarpa (Cav.) Spreng. | x | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala | ||
| BRASSICACEAE | |||||||||||||
| Barbarea orthoceras Ledeb. | x | ph | Canada to Mexico | ||||||||||
| Cardamine obliqua Hochst. ex A. Rich. var. stylosa Rollins | x | ph | disjunct between Mexico and east Africa | ||||||||||
| Descurainia impatiens (Schltd. & Cham.) O.E. Schulz | x | x | x | x | x | x | x | ph | Mexico to Guatemala | ||||
| Draba jorullensis H.B.K. | x | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala |
| *Draba nivicola Rose | x | x | x | x | x | x | x | x | ph | endemic to the alpine TMVB | |||
| Erysimum capitatum (Douglas ex Hook.) Greene | x | x | ph | USA to Mexico | |||||||||
| Lepidium schaffneri Thell. | x | ph | endemic to Mexico | ||||||||||
| Romanschulzia arabiformis (DC.) Rollins | x | ph | endemic to Mexico | ||||||||||
| CAMPANULACEAE | |||||||||||||
| Lobelia nana H.B.K. | x | ph | Mexico to Guatemala | ||||||||||
| CARYOPHYLLACEAE | |||||||||||||
| Arenaria bourgaei Hemsl. | x | ph | Mexico to Ecuador | ||||||||||
| Arenaria bryoides Willd. ex Schltdl. | x | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala | ||
| Arenaria lanuginosa (Michx.) Rohrb. | x | x | ph | USA to South America and the Antilles | |||||||||
| Arenaria lycopodioides Willd. ex Schltdl. | x | ph | Mexico to Guatemala | ||||||||||
| Arenaria oresbia Greenm. | x | x | x | x | x | x | x | ph | endemic to Mexico | ||||
| Arenaria parvifolia Benth. | x | x | x | x | x | x | x | x | ph | disjunct between Mexico and South America | |||
| Arenaria reptans Hemsl. | x | x | x | x | x | x | x | x | x | ph | Mexico, Central America, and South America | ||
| Cerastium brachypodum (Engelm. ex A. Gray) B.L. Rob. | x | ph | Canada to central Mexico | ||||||||||
| +Cerastium glomeratum Thuill. | x | ph | introduced from the Old World | ||||||||||
| Cerastium nutans Raf. | x | x | ph | Canada to Guatemala | |||||||||
| Cerastium orithales Schltdl. | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||||
| *Cerastium purpusii Greenm. | x | x | ph | endemic to the alpine TMVB | |||||||||
| Cerastium ramigerum Bartl. | x | x | x | x | x | x | x | x | ph | endemic to the TMVB | |||
| Cerastium tolucense D.A. Good | x | x | x | x | x | ph | endemic to the TMVB | ||||||
| Cerastium vulcanicum Schltdl. | x | x | x | x | x | x | x | ph | Mexico to Guatemala | ||||
| Colobanthus quitensis (H.B.K.) Bartl. | x | x | x | ph | disjunct between Mexico and South America, also Antarctica | ||||||||
| Drymaria effusa A. Gray var. depressa (Greene) J.A. Duke | x | x | x | x | x | an | USA to Mexico | ||||||
| Sagina saginoides (L.) H. Karst. | x | x | ph | circumboreal, in the Americas from Canada and Greenland to central Mexico | |||||||||
| +Scleranthus annuus L. | x | an | introduced from the Old World | ||||||||||
| +Stellaria media (L.) Vill. | x | an/ph | introduced from the Old World | ||||||||||
| Stellaria umbellata Turzc. | x | ph | disjunct between central Mexico and the western USA and Canada, also in the Old World | ||||||||||
| COMMELINACEAE | |||||||||||||
| Weldenia candida Schult. f. | x | ph | Mexico to Guatemala | ||||||||||
| CRASSULACEAE | |||||||||||||
| Echeveria mucronata Schltdl. | x | ph | endemic to Mexico | ||||||||||
| Echeveria secunda Booth ex Lindl. | x | x | x | x | x | x | x | ph | endemic to central Mexico | ||||
| Echeveria subalpina Rose & Purpus | x | x | ph | endemic to the TMVB | |||||||||
| Sedum clavifolium Rose | x | ph | endemic to the TMVB | ||||||||||
| Sedum goldmanii (Rose) Moran | x | x | x | x | x | x | x | ph | endemic to central Mexico | ||||
| Sedum minimum Rose | x | x | x | x | x | x | ph | endemic to the TMVB | |||||
| Sedum obcordatum R.T. Clausen | x | x | x | ph | endemic to the TMVB | ||||||||
| Tillaea saginoides Maxim. | x | x | x | an | Alaska to southern Mexico, also native to the Old World | ||||||||
| CYPERACEAE | |||||||||||||
| Carex curviculmis Reznicek | x | x | x | ph | endemic to Mexico | ||||||||
| Carex echinata Murray | x | ph | USA to Guatemala | ||||||||||
| Carex geophila Mack. | x | ph | USA to Guatemala | ||||||||||
| Carex hermannii Cochrane | x | ph | endemic to the TMVB | ||||||||||
| Carex orizabae Liebm. | x | x | x | ph | Mexico to Guatemala | ||||||||
| Carex peucophila Holm. | x | x | x | x | ph | Mexico to Guatemala | |||||||
| Eleocharis acicularis (L.) Roem. & Schult. | x | ph | Greenland and Canada to Ecuador, also native to the Old World | ||||||||||
| ELATINACEAE | |||||||||||||
| Elatine brachysperma A. Gray | x | an/ph | USA to Mexico | ||||||||||
| ERICACEAE | |||||||||||||
| Comarostaphylis discolor (Hook.) Diggs | x | sh | Mexico to Guatemala | ||||||||||
| Monotropa hypopitys L. | x | ph | Alaska to Guatemala, also native to the Old World | ||||||||||
| Gaultheria myrsinoides H.B.K. | x | x | x | x | x | x | x | sh/sub | Mexico to South America | ||||
| Vaccinium cespitosum Michx. | x | x | x | x | x | x | x | x | x | sh/sub | Alaska and Canada to Guatemala | ||
| FABACEAE | |||||||||||||
| Astragalus hintonii Barneby | x | ph | endemic to the TMVB | ||||||||||
| Astragalus tolucanus B.L. Rob. & Seaton | x | ph | Mexico to Guatemala | ||||||||||
| Lupinus aschenbornii S. Schauer | x | x | x | x | x | x | ph | endemic to the TMVB | |||||
| Lupinus montanus H.B.K. | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala | |
| Trifolium amabile H.B.K. | x | ph | Mexico to Costa Rica | ||||||||||
| GENTIANACEAE | |||||||||||||
| Gentiana ovatiloba Kusn. | x | x | ph | Mexico to Guatemala | |||||||||
| Gentiana perpusilla Brandegee | x | x | an | endemic to the TMVB | |||||||||
| Halenia brevicornis (H.B.K.) G. Don | x | an | Mexico to South America | ||||||||||
| Halenia plantaginea (H.B.K.) G. Don | x | x | x | x | ph | endemic to Mexico | |||||||
| Halenia pringlei B.L. Rob. & Seaton | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||||
| GERANIACEAE | |||||||||||||
| +Erodium cicutarium (L.) L’Hér. ex Aiton | x | ph | introduced from the Old World | ||||||||||
| Geranium cruceroense R. Knuth | x | ph | endemic to central Mexico | ||||||||||
| Geranium latum Small | x | x | ph | endemic to central Mexico | |||||||||
| Geranium potentillifolium DC. | x | x | ph | endemic to Mexico | |||||||||
| Geranium seemannii Peyr. | x | ph | Mexico to South America | ||||||||||
| GROSSULARIACEAE | |||||||||||||
| Ribes ciliatum Humb. & Bonpl. ex Roem. & Schult. | x | x | x | x | x | x | x | x | sh | Mexico to Costa Rica | |||
| IRIDACEAE | |||||||||||||
| Sisyrinchium quadrangulatum Klatt | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||||
| Sisyrinchium tenuifolium Humb. & Bonpl. ex Willd. | x | x | ph | endemic to Mexico | |||||||||
| JUNCACEAE | |||||||||||||
| Juncus balticus Willd. var. mexicanus (Willd. ex Schult. & Schult. f.) Snogerup | x | ph | Canada to Argentina, also native to the Old World | ||||||||||
| Luzula caricina E. Mey. | x | ph | Mexico to Guatemala | ||||||||||
| Luzula denticulata Liebm. | x | ph | Mexico to Panama | ||||||||||
| Luzula racemosa Desv. | x | x | x | x | x | x | x | x | x | x | x | ph | Mexico to South America |
| Luzula vulcanica Liebm. | x | x | x | ph | Mexico, Central America, and South America | ||||||||
| LAMIACEAE | |||||||||||||
| +Prunella vulgaris L. | x | ph/bi | introduced from the Old World | ||||||||||
| Salvia prunelloides H.B.K. | x | ph | endemic to Mexico | ||||||||||
| Stachys eriantha Benth. | x | x | x | x | x | x | x | ph | disjunct between Mexico and South America | ||||
| MELANTHIACEAE | |||||||||||||
| Anticlea frigida (Schltdl. & Cham.) Zomlefer & Jodd. | x | x | ph | endemic to Mexico | |||||||||
| MONTIACEAE | |||||||||||||
| Calandrinia acaulis H.B.K. | x | x | x | x | x | x | x | x | x | ph | Mexico to South America | ||
| Claytonia perfoliata Donn ex Willd. | x | ph | USA to Guatemala | ||||||||||
| Montia chamissoi (Ledeb. ex Spreng.) Greene | x | an/ph | Alaska and Canada to Mexico | ||||||||||
| Montia fontana L | x | an | temperate zones of the world | ||||||||||
| ONAGRACEAE | |||||||||||||
| Epilobium ciliatum Raf. | x | x | ph | Canada to South America | |||||||||
| OROBANCHACEAE | |||||||||||||
| Castilleja moranensis H.B.K. | x | x | ph | endemic to the TMVB | |||||||||
| Castilleja pectinata M. Martens & Galeotti | x | x | ph | Mexico to Guatemala | |||||||||
| Castilleja scorzonerifolia H.B.K. | x | x | x | x | x | x | x | ph | endemic to Mexico | ||||
| Castilleja tenuifolia M. Martens & Galeotti | x | ph | endemic to Mexico | ||||||||||
| *Castilleja tolucensis H.B.K. | x | x | x | x | x | x | x | endemic to the alpine TMVB | |||||
| Pedicularis orizabae Schltdl. & Cham. | x | ph | central Mexico to Guatemala | ||||||||||
| OXALIDACEAE | |||||||||||||
| Oxalis alpina (Rose) Rose ex R. Knuth | x | x | x | ph | USA to Guatemala | ||||||||
| PHRYMACEAE | |||||||||||||
| Erythranthe glabrata (H.B.K.) G.L. Nesom | x | ph | Canada to Argentina and Chile | ||||||||||
| PIPERACEAE | |||||||||||||
| Peperomia basiradicans G. Mathieu | x | x | ph | endemic to the TMVB | |||||||||
| PLANTAGINACEAE | |||||||||||||
| Callitriche heterophylla Pursh | x | x | ph | Greenland and Canada to South America | |||||||||
| Penstemon gentianoides (H.B.K.) Poir. | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala | |
| Penstemon roseus (Cerv. ex Sweet) G. Don | x | x | x | ph | endemic to Mexico | ||||||||
| Plantago australis Lam. | x | ph | USA to South America | ||||||||||
| Plantago nivea H.B.K. | x | ph | Mexico to Guatemala | ||||||||||
| *Plantago tolucensis Pilg. | x | x | x | x | x | x | ph | endemic to the alpine TMVB | |||||
| Plantago tubulosa Decne. | x | ph | Mexico, Guatemala, and South America | ||||||||||
| Veronica serpyllifolia L. | x | x | ph | widespread in the northern hemisphere and the Americas | |||||||||
| POACEAE | |||||||||||||
| Agrostis bourgaei E. Fourn. | x | x | x | x | ph | endemic to the TMVB | |||||||
| Agrostis calderoniae Acosta Cast. | x | ph | endemic to the TMVB | ||||||||||
| Agrostis perennans (Walter) Tuck. | x | ph | Canada to Guatemala | ||||||||||
| Agrostis subpatens Hitchc. | x | x | x | x | ph | Mexico to Costa Rica | |||||||
| Agrostis tolucensis H.B.K. | x | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Argentina and Chile |
| Bromus carinatus Hook. & Arn. | x | x | ph | USA to Costa Rica | |||||||||
| +Bromus diandrus Roth. | x | ph | introduced from the Old World | ||||||||||
| Bromus exaltatus Bernh. | x | x | ph | central Mexico to Panama | |||||||||
| Calamagrostis eriantha (H.B.K.) Steud. | x | x | x | ph | endemic to Mexico | ||||||||
| Calamagrostis orizabae (Rupr. ex Fourn.) Beal | x | x | x | x | ph | endemic to Mexico | |||||||
| Calamagrostis rigescens (J. Presl) Scribn. | x | x | ph | disjunct between Mexico and South America | |||||||||
| Calamagrostis tolucensis (H.B.K.) Trin. ex Steud. | x | x | x | x | x | x | x | x | ph | central Mexico to Honduras | |||
| Cinna poiformis (H.B.K.) Scribn. & Merr. | x | x | x | x | ph | central Mexico to Bolivia and Peru | |||||||
| Deschampsia liebmanniana (E. Fourn.) Hitchc. | x | x | x | x | ph | endemic to the TMVB | |||||||
| Festuca hephaestophila Nees | x | x | x | x | x | x | x | x | x | ph | central Mexico to Guatemala | ||
| Festuca jaliscana E.B. Alexeev. | x | ph | endemic to the TMVB | ||||||||||
| Festuca livida (H.B.K.) Willd. ex Spreng. | x | x | x | x | x | x | x | ph | central Mexico and Guatemala | ||||
| Festuca orizabensis E.B. Alexeev. | x | ph | endemic to Mexico | ||||||||||
| Festuca tolucensis H.B.K. | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Colombia and Venezuela | |
| Muhlenbergia macroura (H.B.K.) Hitch. | x | ph | northern Mexico to Costa Rica | ||||||||||
| Muhlenbergia nigra Hitchc. | x | x | x | x | x | ph | central Mexico and Guatemala | ||||||
| Muhlenbergia orophila Swallen | x | ph | central Mexico and Guatemala | ||||||||||
| Muhlenbergia peruviana (P. Beauv.) Steud. | x | x | x | ph | southwestern USA to Guatemala; South America | ||||||||
| Muhlenbergia quadridentata (H.B.K.) Trin. | x | x | x | x | ph | northern Mexico to Guatemala | |||||||
| Muhlenbergia ramulosa (H.B.K.) Swallen | x | x | ph | southwestern USA to Costa Rica; Argentina | |||||||||
| Muhlenbergia tricholepis (Torr.) Columbus | x | x | ph | western USA to central Mexico | |||||||||
| Muhlenbergia utilis (Torr.) Hitchc. | x | ph | southwestern USA to southern Mexico | ||||||||||
| Nassella mexicana (Hitchc.) R.W. Pohl | x | x | ph | central Mexico to Argentina | |||||||||
| Peyritschia koelerioides (Peyr.) E. Fourn. | x | x | x | ph | Mexico to Guatemala | ||||||||
| Phleum alpinum L. | x | x | ph | circumboreal, in the Americas from Canada and Greenland to Chile and Argentina | |||||||||
| +Poa annua L. | x | x | x | x | x | x | x | ph | introduced from the Old World | ||||
| Poa calycina (Ball) Refulio var. mathewsii (Ball) Refulio | x | x | x | x | x | x | ph | disjunct between Mexico and South America | |||||
| Poa chamaeclinos Pilg. | x | ph | disjunct between Mexico and South America | ||||||||||
| Poa gymnantha Pilg. | x | ph | disjunct between Mexico and South America | ||||||||||
| Poa orizabensis Hitchc. | x | x | x | ph | Mexico to Guatemala | ||||||||
| +Poa pratensis L. var. pratensis | x | ph | introduced | ||||||||||
| Poa scaberula Hook. f. | x | x | x | x | x | ph | Mexico to South America | ||||||
| Stipa ichu (Ruiz & Pav.) Kunth | x | x | x | ph | Mexico to South America | ||||||||
| Trisetum ligulatum Finot & Zuloaga | x | ph | endemic to Mexico | ||||||||||
| Trisetum rosei Scribn. & Merr. | x | x | x | ph | Mexico to Guatemala | ||||||||
| Trisetum spicatum (L.) K. Richt. | x | x | x | x | x | x | x | x | x | x | x | ph | Canada and Greenland to Chile and Argentina, also native to the Old World |
| +Vulpia myuros (L.) C.C. Gmel. | x | ph | introduced from the Old World | ||||||||||
| POLEMONIACEAE | |||||||||||||
| Polemonium grandiflorum Benth. | x | ph | endemic to Mexico | ||||||||||
| POLYGONACEAE | |||||||||||||
| +Rumex acetosella L. | x | x | ph | introduced from the Old World | |||||||||
| RANUNCULACEAE | |||||||||||||
| Ranunculus donianus Pritz. ex Walp. | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala | |||
| Ranunculus multicaulis D. Don ex G. Don | x | x | x | x | x | ph | Mexico to Central America | ||||||
| Ranunculus peruvianus Pers. | x | ph | Mexico to South America | ||||||||||
| Ranunculus praemorsus H.B.K. ex DC. | x | ph | Mexico to South America | ||||||||||
| ROSACEAE | |||||||||||||
| Acaena elongata L. | x | x | x | sh | Mexico to South America | ||||||||
| Alchemilla aphanoides Mutis ex L. f. | x | x | ph | Mexico to Central America | |||||||||
| Alchemilla pinnata Ruiz & Pav. | x | x | ph | Mexico to Guatemala; disjunct in South America | |||||||||
| Alchemilla pringlei (Rydb.) Fedde | x | x | x | x | ph | Mexico to Guatemala | |||||||
| Alchemilla procumbens Rose | x | x | x | x | x | x | x | ph | Mexico to South America | ||||
| Alchemilla vulcanica Schltdl. & Cham. | x | x | x | x | x | x | x | x | x | x | x | ph | Mexico to South America |
| Potentilla candicans Humb. & Bonpl. ex Nestl. | x | ph | endemic to the TMVB | ||||||||||
| Potentilla ranunculoides H.B.K. | x | x | x | x | x | x | x | ph | endemic to Mexico | ||||
| Potentilla richardii Lehm. | x | x | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||
| Sibbaldia procumbens L. | x | x | x | x | ph | circumboreal, in the Americas from Canada and Greenland to Mexico | |||||||
| RUBIACEAE | |||||||||||||
| Crusea longiflora (Roem. & Schult.) W.R. Anderson | x | an | Mexico to Central America | ||||||||||
| SALICACEAE | |||||||||||||
| Salix paradoxa H.B.K. | x | tr/sh | endemic to Mexico | ||||||||||
| SCROPHULARIACEAE | |||||||||||||
| Limosella aquatica L. | x | x | x | an | also native to the Old World | ||||||||
| SOLANACEAE | |||||||||||||
| Solanum demissum Lindl. | x | x | x | ph | central Mexico to Guatemala | ||||||||
| URTICACEAE | |||||||||||||
| Urtica orizabae Liebm. | x | ph | endemic to the TMVB | ||||||||||
| VERBENACEAE | |||||||||||||
| Glandularia teucriifolia (M. Martens & Galeotti) Umber. | x | x | ph | Mexico to Guatemala | |||||||||
| VIOLACEAE | |||||||||||||
| Viola beamanii Calderón | x | x | ph | endemic to the TMVB | |||||||||
| Viola hemsleyana Calderón | x | ph | endemic to Mexico | ||||||||||
Appendix B
Excluded species. The names Berberis schiedeana Schltdl. (Berberidaceae), Juncus articus Willd. (Juncaceae), and Poa villaroelii Phil. (Poaceae) have been misapplied to Berberis alpina, Juncus balticus var. mexicanus, and Poa chamaechlinos, respectively. We have been unable to find specimens of the following species to verify their prescence: Asteraceae: Ageratina oreithales (Greenm.) B.L. Turner, A. pazcuarensis (H.B.K.) R.M. King & H. Rob., Heterosperma pinnatum Cav., Pseudognaphalium canescens Anderb.; Cyperaceae: Carex longicaulis Boeck., C. volcanica F.J. Herm., Eleocharis montana (H.B.K.) Roem. & Schult.; Fabaceae: Lupinus mexicanus Cerv; Gentianaceae: Halenia crassiuscula B.L. Rob. & Seat.; Geraniaceae: Geranium aristisepalum H.E. Moore; Onagraceae: Oenothera purpusii Munz; Pinaceae: Pinus montezumae Lamb.; Plantaginaceae: Plantago major L., Plantago sericea Benth.; Poaceae: Agrostis exarata Trin., A. hyemalis (Walter) Britton, Sterns & Poggenb., A. vinosa Swallen, Muhlenbergia montana (Nutt.) Hitchc., Trisetum altijugum (E. Fourn.) Scribn., T. irazuense (Kuntze) Hitchc., T. viride (H.B.K.) Kunth; Violaceae: Viola painteri Rose & House.
References
- Rzedowski, J. Vegetación de México, 1st ed.; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: México City, Mexico, 2006; p. 504. [Google Scholar]
- Beaman, J.H. The timberlines of Iztaccihuatl and Popocatepetl. Ecology 1962, 43, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Körner, C.H. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems, 2nd ed.; Springer Science & Business Media: New York, NY, USA, 2003; p. 349. [Google Scholar]
- Billings, W.D. Adaptations and origins of alpine plants. Arct. Alp. Res. 1974, 6, 129–142. [Google Scholar] [CrossRef]
- Miranda, F. La Vegetación de Chiapas; Gobierno del estado de Chiapas: Tuxtla Gutiérrez, Mexico, 1952; p. 324. [Google Scholar]
- Beaman, J.H.; Andersen, J.W. The vegetation, floristics and phytogeography of the summit of Cerro Potosí, Mexico. Am. Midl. Naturalist. 1966, 75, 1–33. [Google Scholar] [CrossRef]
- McDonald, J. The alpine-subalpine flora of northeastern Mexico. SIDA 1990, 14, 21–28. [Google Scholar]
- García-Arévalo, A.; González-Elizondo, S. Flora y vegetación de la cima del Cerro Potosí, Nuevo León, México. Acta Bot. Mex. 1991, 13, 53–74. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Tuena, A.; Orozco-Esquivel, M.T.; Ferrari, L. Igneous petrogenesis of the Trans-Mexican volcanic belt. Geo. Soc. Amer. Spec. Pap. 2007, 422, 129–181. [Google Scholar]
- Almeida-Leñero, L.; Escamilla, M.; De Azcárate, J.G.; González-Trápaga, A.; Cleef, A.M. Vegetación alpina de los volcanes Popocatépetl, Iztaccíhuatl y Nevado de Toluca. In Biodiversidad de la Faja Volcánica Transmexicana; Luna, I., Morrone, J.J., Espinosa, D., Eds.; FES-Zaragoza-CONABIO: México City, Mexico, 2007; pp. 179–198. [Google Scholar]
- García, E. Modificaciones al Sistema de Clasificación de Köppen, 2nd ed.; Instituto de Geofísica, Universidad Nacional Autónoma de México: México City, Mexico, 1973; p. 246. [Google Scholar]
- Lauer, W.; Klaus, D. Geoecological investigations on the timberline of Pico de Orizaba, Mexico. Arct. Alp. Res. 1975, 7, 315–330. [Google Scholar] [CrossRef]
- Smith, A.P.; Young, T.P. Tropical alpine plant ecology. Annu. Rev. Ecol. Syst. 1987, 18, 137–158. [Google Scholar] [CrossRef]
- Hedberg, O. Features of afroalpine plant ecology. Acta Phytogeogr. Suecica 1964, 49, 1–44. [Google Scholar]
- Rundel, P.W. Tropical alpine climates. In Tropical Alpine Environments: Plant Form and Function; Rundel, P.W., Smith, A.P., Meinzer, F.C., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 21–44. [Google Scholar]
- Emiliani, C. Paleotemperature analysis of Caribbean cores P6304-8 and P6304-9 and a generalized temperature curve for the past 425,000 years. J. Geol. 1996, 74, 109–124. [Google Scholar] [CrossRef]
- Mastretta-Yanes, A.; Moreno-Letelier, A.; Piñero, D.; Jorgensen, T.H.; Emerson, B.C. Biodiversity in the Mexican highlands and the interaction of geology, geography and climate within the Trans-Mexican Volcanic Belt. J. Biogeogr. 2015, 42, 1586–1600. [Google Scholar] [CrossRef]
- McDonald, J.A. Fitogeografía e historia de la flora alpina-subalpina del noreste de México: Orígenes y Distribución. In Diversidad Biológica de México; Ramamoorthy, T., Bye, R., Lot, A., Fa, A., Eds.; Instituto de Biología, Universidad Nacional de México: México City, Mexico, 1998; pp. 665–686. [Google Scholar]
- Buytaert, W.; Cuesta-Camacho, F.; Tobón, C. Potential impacts of climate change on the environmental services of humid tropical alpine regions. Glob. Ecol. Biogeogr. 2011, 20, 19–33. [Google Scholar] [CrossRef]
- Ernakovich, J.G.; Hopping, K.A.; Berdanier, A.B.; Simpson, R.T.; Kachergis, E.J.; Steltzer, H.; Wallenstein, M.D. Predicted responses of arctic and alpine ecosystems to altered seasonality under climate change. Glob. Chang. Biol. 2014, 20, 3256–3269. [Google Scholar] [CrossRef] [PubMed]
- Pauli, H.; Gottfried, M.; Reiter, K.; Klettner, C.; Grabherr, G. Signals of range expansions and contractions of vascular plants in the high Alps: Observations (1994–2004) at the GLORIA* master site Schrankogel, Tyrol, Austria. Glob. Chang. Biol. 2007, 13, 147–156. [Google Scholar] [CrossRef]
- Rehfeldt, G.; Crookston, N.; Sáenz, C.; Campbell, E. North American vegetation model for land-use planning in a changing climate: A solution to large classification problems. Ecol. Appl. 2012, 22, 119–141. [Google Scholar] [CrossRef]
- Seimon, T.A.; Seimon, A.; Daszak, P.; Halloy, S.R.; Schloegel, L.M.; Aguilar, C.A.; Simmons, E. Upward range extension of Andean anurans and chytridiomycosis to extreme elevations in response to tropical deglaciation. Glob. Change Biol. 2007, 13, 288–299. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA. 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Amezcua, Y.; Steinmann, V.W.; Ruiz-Sanchez, E.; Rojas-Soto, O.R. Mexican alpine plants in the face of global warming: Potential extinction within a specialized assemblage of narrow endemics. Biodivers. Conserv. 2016, 25, 865–888. [Google Scholar] [CrossRef]
- Maza-Villalobos, S.; Cotler, H.; Almeida-Leñero, L.; Hoth, J.; Steinmann, V.W.; Mastretta, A.; Hernández-Cárdenas, R.A. Conservando el pastizal alpino mexicano. Conocimientos, amenazas y esperanzas. Biodiversitas 2019, 142, 12–16. [Google Scholar]
- Scherrer, P.; Pickering, C.M. Effects of grazing, tourism and climate change on the alpine vegetation of Kosciuszko National Park. Vic. Naturalist. 2001, 118, 93. [Google Scholar]
- Fernández, F.J.A.; López-Domínguez, J.C. Biodiversidad del Parque Nacional La Malinche; Gobierno del Estado de Tlaxcala: Tlaxcala, Mexico, 2005; p. 224. [Google Scholar]
- Narave, H. La vegetación del cofre de Perote, Veracruz, Mexico. Biótica 1985, 10, 35–64. [Google Scholar]
- Hernández-Cárdenas, R.A.; Mendoza-Ruiz, A.; Arredondo-Amezcua, L.; Steinmann, V.W. The Alpine Ferns of the Trans-Mexican Volcanic Belt. Am. Fern J. 2019, 109, 11–25. [Google Scholar] [CrossRef]
- Neyra, J.A. Guía de Las Altas Montañas de México y Una de Guatemala, 1st ed.; CONABIO y Gobierno Federal: México City, Mexico, 2012; p. 413. [Google Scholar]
- McVaugh, J. Gramineae. In Flora Novo-Galiciana: A Descriptive Account of the Vascular Plants of Western Mexico; Anderson, J.R., Ed.; Ann Arbor University of Michigan Press: Ann Arbor, MI, USA, 1983; Volume 14, pp. 1–436. [Google Scholar]
- De Rzedowski, G.C.; Rzedowski y colaboradores, J. Flora Fanerogámica del Valle de México, 2nd ed.; 1a reimp.; @Instituto de Ecología, A.C. y Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Pátzcuaro, Mexico, 2005; p. 1406. [Google Scholar]
- Durán-Espinosa, C. Grossulariaceae. In Flora de Veracruz; Sosa, V., Ed.; Instituto de Ecología, A.C. y Centro de Investigaciones Tropicales: Xalapa, Mexico, 2001; Volume 122, pp. 1–15. [Google Scholar]
- Villareal, J.A. Gentianaceae. In Flora de Veracruz; Sosa, V., Ed.; Instituto de Ecología, A.C. y Centro de Investigaciones Tropicales: Xalapa, Mexico, 2001; Volume 121, pp. 1–67. [Google Scholar]
- Thiers, B. Index Herbariorum: A global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/science/ih (accessed on 8 November 2020).
- Lot, A.; Chiang, F. Manual de Herbario: Administración y Manejo de Colecciones, Técnicas de Recolección y Preparación de Ejemplares Botánicos; Consejo Nacional de la Flora de México: Ciudad de México, Mexico, 1986. [Google Scholar]
- Mickel, J.T.; Smith, A.R. The Pteridophytes of Mexico (Memoirs of the New York Botanical Garden 88); The New York Botanical Garden: New York, NY, USA, 2004; p. 1054. [Google Scholar]
- Sigel, E.M.; Windham, M.D.; Haufler, C.H.; Pryer, K.M. Phylogeny, divergence time estimates, and phylogeography of the diploid species of the Polypodium vulgare complex (Polypodiaceae). Syst. Bot. 2014, 39, 1042–1055. [Google Scholar] [CrossRef]
- Ppg, I.; Shmakov, A. A community-based classification for extant ferns and lycophytes. J. Syst. Evol. 2016, 6, 563–603. [Google Scholar] [CrossRef]
- Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; Stevens, P.F. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. 2016, 181, 1–20. [Google Scholar]
- GBIF: The Global Biodiversity Information Facility. Available online: https://www.gbif.org (accessed on 15 June 2017).
- Tropicos.org. Missouri Botanical Garden. Available online: http://www.tropicos.org (accessed on 10 January 2015).
- iNaturalist Application. Available online: https://www.inaturalist.org/ (accessed on 1 November 2020).
- IUCN 2020. The IUCN Red List of Threatened Species. Version 2020-3. Available online: www.iucnredlist.org (accessed on 10 February 2020).
- Secretaría de Medio Ambiente y Recursos Naturales. Norma Oficial Mexicana NOM-059-SEMARNAT-2010. Protección Ambiental-Especies nativas de México de flora y fauna silvestres-Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio. Lista de especies en riesgo. D. Of. De La Fed. De México 2010, 30, 1–78. [Google Scholar]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environm. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Zamudio, S. Notas sobre el género Berberis (Berberidaceae) en México. Acta Bot. Mex. 2009, 87, 31–70. [Google Scholar] [CrossRef] [Green Version]
- Villaseñor, J.L. Checklist of the native vascular plants of Mexico. Rev. Mex. Biodivers. 2016, 87, 559–902. [Google Scholar] [CrossRef] [Green Version]
- Körner, C.H. Alpine plant diversity: A global survey and functional interpretations. In Arctic and Alpine Biodiversity: Patterns, Causes and Ecosystem Consequences; Chapin, F.S., Körner, C., Eds.; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- Luteyn, J.L. Páramos, A Checklist of Plant Diversity, Geographical Distribution, and Botanical Literature; New York Botanical Garden Press: New York, NY, USA, 1999; p. 278. [Google Scholar]
- Rai, I.D.; Adhikari, B.S.; Rawa, G.S. Floral diversity along subalpine and alpine ecosystems in Tungnath area of Kedarnath wildlife sanctuary, Uttarakhand. Indian Forester. 2012, 138, 927–940. [Google Scholar]
- Hoxey, P. Notes on the cacti of the Sandia valley, Puno, Peru and a description of a new species of Echinopsis (Cactaceae). Bradleya 2016, 34, 187–199. [Google Scholar] [CrossRef]
- Delgadillo, C. Musgos alpinos del Nevado de Colima, México. Bot. Sci. 1987, 47, 7–11. [Google Scholar] [CrossRef]
- Sklenář, P.; Balslev, H. Geographic flora elements in the Ecuadorian superpáramo. Flora-Morphol. Distrib. Funct. Ecol. Plants 2007, 202, 50–61. [Google Scholar] [CrossRef]
- Klimes, L. Life-forms and clonality of vascular plants along an altitudinal gradient in E Ladakh (NW Himalayas). Basic Appl. Ecol. 2003, 4, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Ángel, R.; Conrad, R.; Dvorsky, M.; Kopecky, M.; Kotilínek, M.; Hilesalu, I.; Schweingruber, F.; Dolezal, J. The root-associated microbial community of the world’s highest growing vascular plants. Microb. Ecol. 2016, 72, 394–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuesta, F.; Muriel, P.; Beck, S.; Meneses, R.I.; Halloy, S.; Salgado, S.; Ortiz, E.; Becerra, M.T. (Eds.) Biodiversidad y Cambio Climático en los Andes Tropicales—Conformación de Una Red de Investigación Para Monitorear sus Impactos y Delinear Acciones de Adaptación; Red Gloria-Andes: Lima, Ecuador, 2012; p. 180. [Google Scholar]
- Vargas, G.; Sánchez, J.J. Plantas con flores de los páramos de Costa Rica y Panamá: El páramo ístmico. In Páramos de Costa Rica; Kappelle, M., Horn, S.P., Eds.; INBio: San José, Costa Rica, 2005; pp. 397–435. [Google Scholar]
- Ferrari, L.; Orozco-Esquivel, T.; Manea, V.; Manea, M. The dynamic history of the Trans-Mexican Volcanic Belt and the Mexico subduction zone. Tectonophysics 2012, 522, 122–149. [Google Scholar] [CrossRef]
- Soreng, R.J.; Peterson, P.M. Revision of Poa, L. (Poaceae, Pooideae, Poeae, Poinae) in Mexico: New records, re-evaluation of P. ruprechtii, and two new species, P. palmeri and P. wendtii. PhytoKeys 2012, 15, 1–104. [Google Scholar] [CrossRef] [Green Version]
- Dehshiri, M.M.; Safikhani, K.; Mostafavi, H. Alpine flora of some part of Alvand Mountain, Hamadan province. J. Pl. Biol. 2016, 30, 89–104. [Google Scholar]
- Mark, A.F.; Dickinson, K.J.M.; Hofstede, R.G.M. Alpine vegetation, plant distribution, life forms, and environments in a perhumid New Zealand Region: Oceanic and tropical high mountain affinities. Arct. Antarc. Alp. Res. 2000, 32, 240–254. [Google Scholar] [CrossRef]
- Smith, A.P. Introduction to tropical vegetation. In Tropical Alpine Environments: Plant Form and Function; Rundel, P.W., Smith, A.P., Meinzer, F.C., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 1–19. [Google Scholar]
- Aubert, S.; Boucher, F.; Lavergne, F.; Renaud, J.; Choler, P. 1914–2014: A revised worldwide catalogue of cushion plants 100 years after Hauri and Schröter. Alp. Bot. 2014, 124, 59–70. [Google Scholar] [CrossRef]
- Morrone, J.J. Fundamental biogeographic patterns across the Mexican Transition Zone: An evolutionary approach. Ecography 2010, 33, 335–361. [Google Scholar] [CrossRef]
- Gámez, N.; Escalante, T.; Rodríguez, G.; Linaje, M.; Morrone, J.J. Caracterización biogeográfica de la Faja Volcánica Transmexicana y análisis de los patrones de distribución de su mastofauna. Rev. Mex. Biodivers. 2012, 81, 258–272. [Google Scholar] [CrossRef] [Green Version]
- Sarmiento, G. Ecological Features of Climate. In High Tropical Mountains. High Altitude Tropical Biogeography; Vuilleumer, F., Monasterio, M., Eds.; Oxford University Press: New York, NY, USA, 1986; pp. 11–45. [Google Scholar]
- Rzedowski, J. An Ecological and Phytogeographical Analysis of the Grasslands of Mexico. Taxon 1975, 24, 67–80. [Google Scholar] [CrossRef]
- Ramírez-Villegas, J.; Cuesta, F.; Devenish, C.; Peralvo, M.; Jarvis, A.; Arnillas, C.A. Using species distributions models for designing conservation strategies of Tropical Andean biodiversity under climate change. J. Nat. Conserv. (Jena) 2014, 22, 391–404. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Ghosn, D. Recent plant diversity changes on Europe’s mountain summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [Green Version]
- Heilprin, A. The Temperate and Alpine Floras of the Giant Volcanoes of Mexico. Proc. Am. Philos. Soc. 1892, 30, 4–22. [Google Scholar]
Figure 1.
Map of the Trans-Mexican Volcanic Belt of central Mexico showing the location of the volcanoes with alpine vegetation at their summits.
Figure 1.
Map of the Trans-Mexican Volcanic Belt of central Mexico showing the location of the volcanoes with alpine vegetation at their summits.

Figure 2.
Alpine vegetation of central Mexico. (A) North side of the Pico de Orizaba; (B) La Malinche from a distance; (C) Nevado de Toluca; (D) north side of Popocátepetl; (E,F) Nevado de Toluca; (G) rocky outcrop covered with Echeveria secunda on Tláloc; (H) seep on Iztaccíhuatl. Credits: (A,E,F,H) Victor Steinmann; (B,D,G) Adolfo López; (C) Libertad Arredondo.
Figure 2.
Alpine vegetation of central Mexico. (A) North side of the Pico de Orizaba; (B) La Malinche from a distance; (C) Nevado de Toluca; (D) north side of Popocátepetl; (E,F) Nevado de Toluca; (G) rocky outcrop covered with Echeveria secunda on Tláloc; (H) seep on Iztaccíhuatl. Credits: (A,E,F,H) Victor Steinmann; (B,D,G) Adolfo López; (C) Libertad Arredondo.

Figure 3.
Characteristic plants of the central Mexican alpine flora. (A) Senecio procumbens; (B) Potentilla richardii; (C) Phacelia platycarpa; (D) Calandrinia acaulis; (E) Eryngium proteiflorum; (F) Vaccinium cespitosum; (G) Cerastium purpusii; (H) Castilleja tolucensis growing in a cushion of Arenaria bryoides; (I) Ranunculus multicaulis; (J) Erigeron galeottii; (K) Festuca livida; (L) Carex curviculmis; (M) Cerastium tolucense; (N) Berberis alpina; (O) Arenaria bryoides; (P) Lupinus montanus; (Q) Chaerophyllum tolucanum; (R) Luzula racemosa. Credits: (A–C,E–N,P,Q) Victor Steinmann; (D) Libertad Arredondo; (O) Adolfo López; (R) Harald Pauli.
Figure 3.
Characteristic plants of the central Mexican alpine flora. (A) Senecio procumbens; (B) Potentilla richardii; (C) Phacelia platycarpa; (D) Calandrinia acaulis; (E) Eryngium proteiflorum; (F) Vaccinium cespitosum; (G) Cerastium purpusii; (H) Castilleja tolucensis growing in a cushion of Arenaria bryoides; (I) Ranunculus multicaulis; (J) Erigeron galeottii; (K) Festuca livida; (L) Carex curviculmis; (M) Cerastium tolucense; (N) Berberis alpina; (O) Arenaria bryoides; (P) Lupinus montanus; (Q) Chaerophyllum tolucanum; (R) Luzula racemosa. Credits: (A–C,E–N,P,Q) Victor Steinmann; (D) Libertad Arredondo; (O) Adolfo López; (R) Harald Pauli.

Figure 4.
Family-level composition of the vascular flora of central Mexico.

Table 1.
General information about location, diversity, and volcanic activity of the study sites. Source: * Global Volcanism Program; n Neyra, 2012.
Table 1.
General information about location, diversity, and volcanic activity of the study sites. Source: * Global Volcanism Program; n Neyra, 2012.
| Volcano | Location | Elevation (m) | Summit Coordinates | Alpine Area (km2) | Number of Species | Last Eruption (ya) |
|---|---|---|---|---|---|---|
| El Ajusco | Ciudad de México | 3940 | 19°12′25″ N | 2 | 63 | 1620 * |
| 99°15′35″ W | ||||||
| Telapón | Estado de México and Puebla | 4080 | 19°22′15.42″ N | 0.03 | 57 | 30,000 n |
| 98°43′12.42″ W | ||||||
| Tláloc | Estado de México and Puebla | 4140 | 19°24′41.7″ N | 0.97 | 72 | 30–31,000 n |
| 98°42′44.8″ W | ||||||
| Cofre de Perote | Veracruz | 4230 | 19°29′39.24″ N | 1.57 | 93 | 900 * |
| 97°08′51.12″ W | ||||||
| Nevado de Colima | Jalisco | 4280 | 19°33′46.92″ N | 0.91 | 31 | 30–35,000 n |
| 103°36′30.66″ W | ||||||
| Malinche | Puebla and Tlaxcala | 4440 | 19°13′51.54″ N | 3.55 | 62 | 3190 * |
| 98°01′55.26″ W | ||||||
| Sierra Negra | Puebla | 4580 | 18°59′07.68″ N | 5.23 | 46 | unknown |
| 97°18′51.24″ W | ||||||
| Nevado de Toluca | Estado de México | 4690 | 19°06′06.72″ N | 15.27 | 102 | 3370 * |
| 99°46′03.36″ W | ||||||
| Iztaccíhuatl | Puebla and Estado de México | 5220 | 19°10′48.4″ N | 38.58 | 170 | 20,000 (fissurelike eruption at 9000) n |
| 98°38′29.3″ W | ||||||
| Popocatépetl | Estado de México, Puebla and Morelos | 5460 | 19°01′18.4″ N | 32.79 | 81 | currently active * |
| 98°37′39.6″ W | ||||||
| Pico de Orizaba | Veracruz y Puebla | 5640 | 19°01′48″ N | 48.83 | 78 | 174 * |
| 97°16′10″ W |
Table 2.
Phytogeographic summary of the species.
| Distribution | Number of Species (Percentage of Flora) |
|---|---|
| Endemic to the study area | 4 (1.7%) |
| Endemic to the study area and adjacent subalpine forest | 8 (3.4%) |
| Endemic to the Trans-Mexican Volcanic Belt | 36 (14.8%) |
| Endemic to Mexico | 76 (32.1%) |
| Mexico to Guatemala | 41 (17.3%) |
| Mexico to Central America (beyond Guatemala) | 15 (6.3%) |
| Mexico, Central America, and South America | 29 (12.2%) |
| Disjunct between Mexico and South America | 9 (3.8%) |
| Canada and/or USA to Mexico | 9 (3.8%) |
| Canada and/or USA to Central/South America | 20 (8.4%) |
| Also native to the Old World | 15 (6.3%) |
| Introduced from the Old World | 12 (5.1%) |
Table 3.
Distribution of central Mexican alpine genera occurring only in the Americas.
| Region | Genera |
|---|---|
| Endemic to Mexico | Mexerion |
| USA/Canada to Mexico | Hymenoxys, Tauschia |
| USA/Canada to Central America | Comarostaphylis, Laennecia, Roldana |
| Mexico to Central America | Dahlia, Osbertia, Oxylobus, Romanschulzia, Weldenia |
| USA/Canada to South America | Barkleyanthus, Crusea, Gaga, Glandularia, Nassela, Phacelia |
| Mexico to South America | Alansmia, Chionolaena, Echeveria, Melpomene, Ottoa, Peyritschia, Selloa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Steinmann, V.W.; Arredondo-Amezcua, L.; Hernández-Cárdenas, R.A.; Ramírez-Amezcua, Y. Diversity and Origin of the Central Mexican Alpine Flora. Diversity 2021, 13, 31. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010031
AMA Style
Steinmann VW, Arredondo-Amezcua L, Hernández-Cárdenas RA, Ramírez-Amezcua Y. Diversity and Origin of the Central Mexican Alpine Flora. Diversity. 2021; 13(1):31. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010031
Chicago/Turabian StyleSteinmann, Victor W., Libertad Arredondo-Amezcua, Rodrigo Alejandro Hernández-Cárdenas, and Yocupitzia Ramírez-Amezcua. 2021. "Diversity and Origin of the Central Mexican Alpine Flora" Diversity 13, no. 1: 31. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010031
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.