
Novelties in Fuscosporellaceae (Fuscosporellales): Two New Parafuscosporella from Thailand Revealed by Morphology and Phylogenetic Analyses

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Isolation and Morphological Data
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Sequence Alignment and Phylogenetic Analyses
3. Results
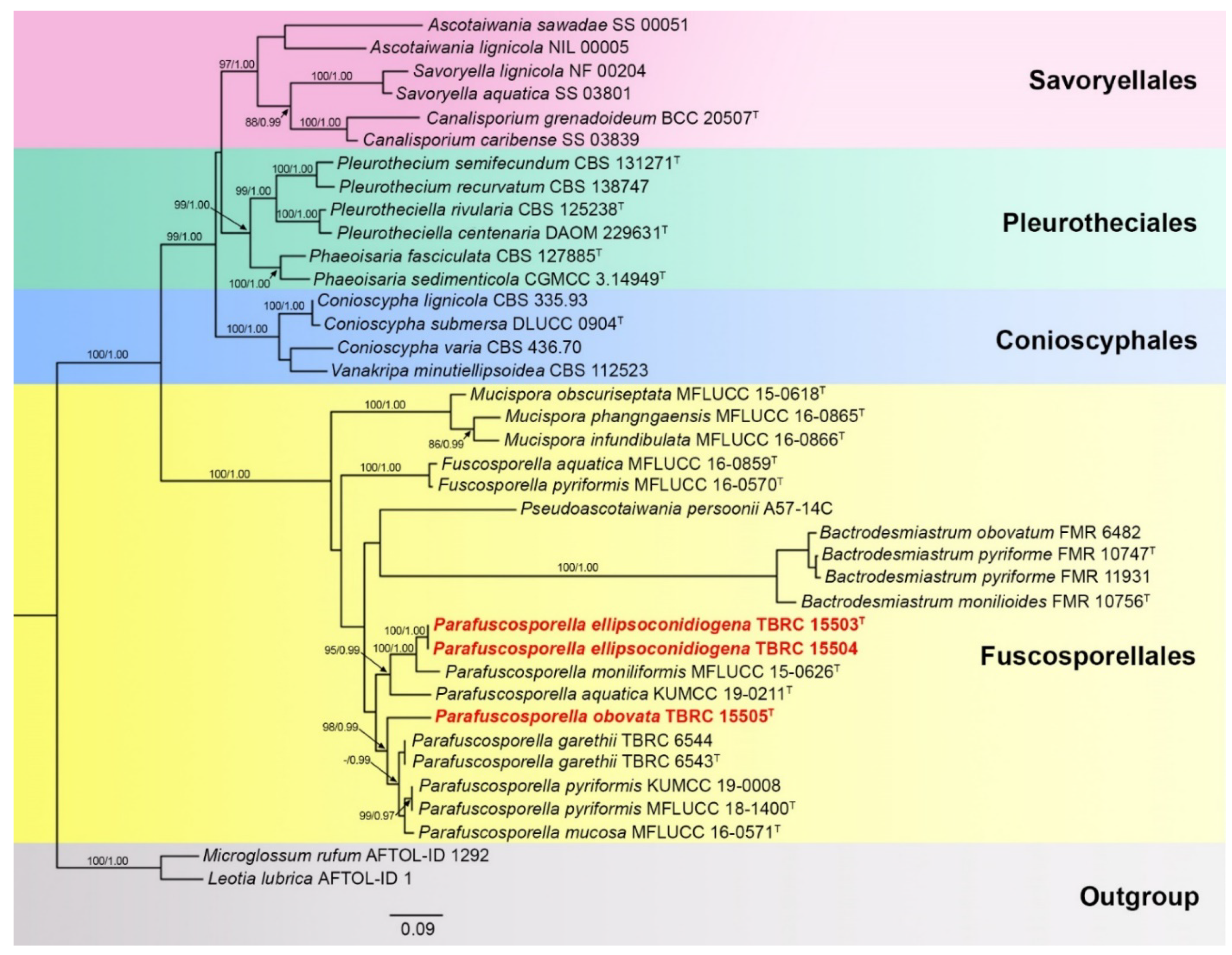
3.1. Molecular Phylogeny
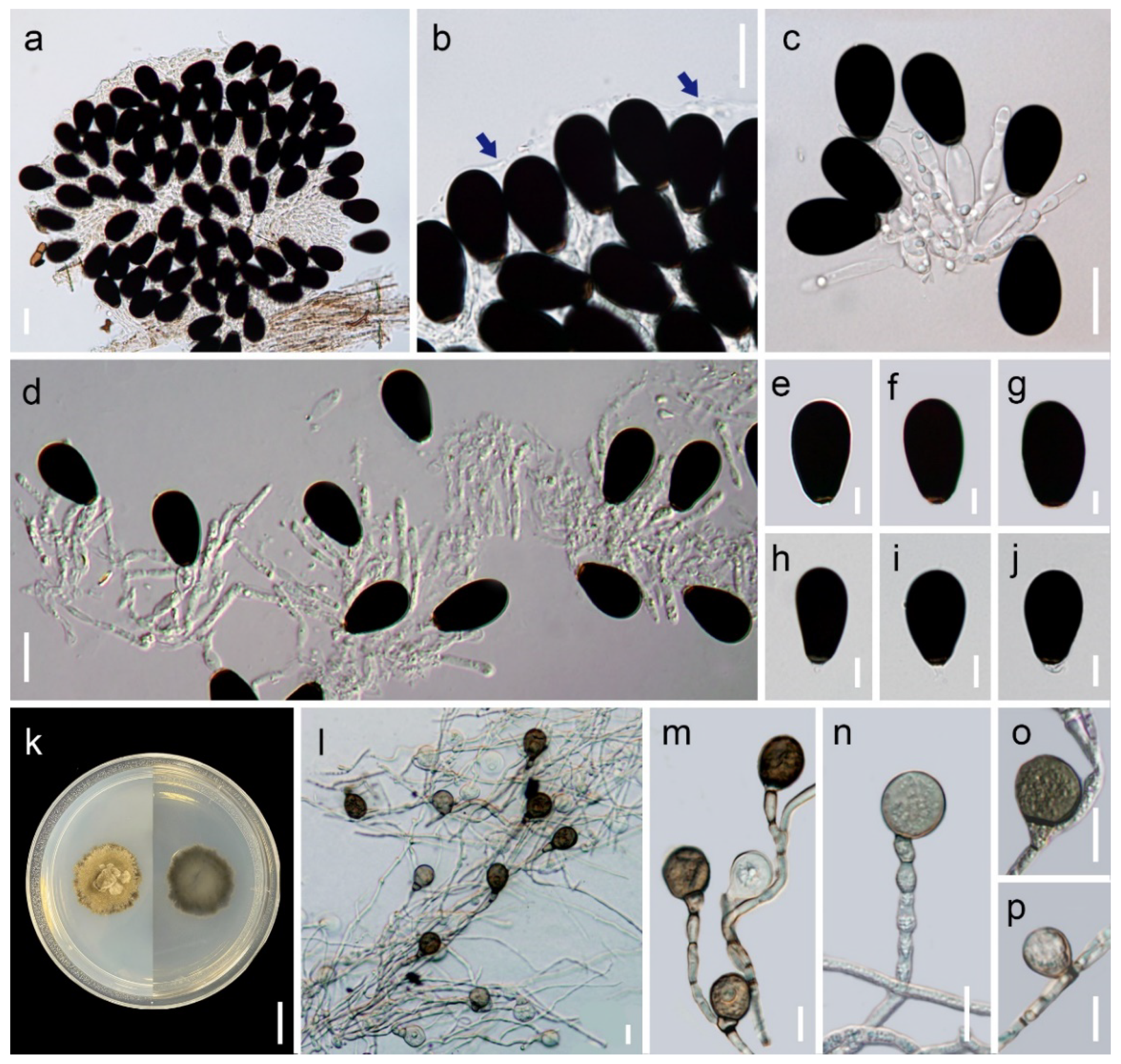
3.2. Taxonomy
3.3. Key to the species of Parafuscosporella
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, J.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Al-Sadi, A.M.; Lumyong, S.J. Fuscosporellales, a new order of aquatic and terrestrial Hypocreomycetidae (Sordariomycetes). Cryptogam. Mycol. 2016, 37, 449–475. [Google Scholar] [CrossRef]
- Boonyuen, N.; Chuaseeharonnachai, C.; Suetrong, S.; Sujinda, S.; Somrithipol, S. Parafuscosporella garethii sp. nov. (Fuscosporellales) from a rivulet in a community-based northern forest, in Thailand. Mycosphere 2016, 7, 1265–1272. [Google Scholar] [CrossRef]
- Yang, J.; Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Liu, Z.Y. Two new species in Fuscosporellaceae from freshwater habitats in Thailand. Mycosphere 2017, 8, 1893–1903. [Google Scholar] [CrossRef]
- Yang, H.; Dong, W.; Yu, X.D.; Bhat, D.J.; Boonmee, S.; Zhang, H. Four freshwater dematiaceous hyphomycetes in Sordariomycetes with two new species of Parafuscosporella. Phytotaxa 2020, 441, 19–34. [Google Scholar] [CrossRef]
- Chuaseeharonnachai, C.; Somrithipol, S.; Boonyuen, N. A new species of Fusticeps from Thailand. Mycosphere 2014, 5, 313–317. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, D.J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The faces of fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Sri-indrasutdhi, V.; Boonyuen, N.; Suetrong, S.; Chuaseeharonnachai, C.; Sivichai, S.; Gareth Jones, E.B. Wood-inhabiting freshwater fungi from Thailand: Ascothailandia grenadoidia gen. et sp. nov., Canalisporium grenadoidia sp. nov. with a key to Canalisporium species (Sordariomycetes, Ascomycota). Mycoscience 2010, 51, 411–420. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, R. Practical Methods of Optimization; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Le, S.Q.; Dang, C.C.; Gascuel, O. Modeling protein evolution with several amino acid replacement matrices depending on site rates. Mol. Biol. Evol. 2012, 29, 2921–2936. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Mark, P.; Ayres, D.L.; Höhna, A.D.S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Sukumaran, J.; Holder, M.T. DendroPy: A Python library for phylogenetic computing. Bioinformatics 2010, 26, 1569–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonyuen, N.; Chuaseeharonnachai, C.; Suetrong, S.; Sri-indrasutdhi, V.; Sivichai, S.; Gareth Jones, E.B.; Pang, K.L. Savoryellales (Hypocreomycetidae, Sordariomycetes): A novel lineage of aquatic ascomycetes inferred from multiple-gene phylogenies of the genera Ascotaiwania, Ascothailandia, and Savoryella. Mycologia 2011, 103, 1351–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Restrepo, M.; Gené, J.; Castañeda-Ruiz, R.F.; Mena-Portales, J.; Guarro, J. Emendation of the genus Bactrodesmiastrum (Sordariomycetes) and description of Bactrodesmiastrum monilioides sp. nov. from plant debris in Spain. Mycol. Prog. 2015, 14, 48. [Google Scholar] [CrossRef]
- Hernández-Restrepo, M.; Mena-Portales, J.; Gené, J.; Cano, J.; Guarro, J. New Bactrodesmiastrum and Bactrodesmium from decaying wood in Spain. Mycologia 2013, 105, 172–180. [Google Scholar] [CrossRef]
- Réblová, M.; Seifert, K. Conioscyphascus, a new ascomycetous genus for holomorphs with Conioscypha anamorphs. Stud. Mycol. 2004, 50, 95–108. [Google Scholar]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.F.; Bhat, D.J.; Lin, C.G.; Li, W.L.; Jing, Y. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef] [Green Version]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, D.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Lutzoni, F.; Kauff, F.; Cox, C.J.; McLaughlin, D.; Celio, G.; Dentinger, B.; Padamsee, M.; Hibbett, D.; James, T.J.; Baloch, E.; et al. Assembling the fungal tree of life: Progress, classification, and evolution of subcellular traits. Am. J. Bot. 2004, 91, 1446–1480. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Sung, G.H.; Johnson, D.; Hesse, C.; O’Rourke, B.; Serdani, M.; Spotts, R.; Lutzoni, F.; Hofstetter, V.; Miadlikowska, J.; et al. A five-gene phylogeny of Pezizomycotina. Mycologia 2006, 98, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Réblová, M.; Seifert, K.A.; Fournier, J.; Štepánek, V. Newly recognised lineages of perithecial ascomycetes: The new orders Conioscyphales and Pleurotheciales. Pers. Mol. Phylogeny Evol. Fungi 2016, 37, 57–81. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.L.; Li, W.; Zhang, T.Y. A new species of Phaeoisaria from intertidal marine sediment collected in Weihai, China. Mycotaxon 2014, 127, 17–24. [Google Scholar] [CrossRef]
- Réblová, M.; Seifert, K.A.; Fournier, J.; Štěpánek, V. Phylogenetic classification of Pleurothecium and Pleurotheciella gen. nov. and its dactylaria-like anamorph (Sordariomycetes) based on nuclear ribosomal and protein-coding genes. Mycologia 2012, 104, 1299–1314. [Google Scholar] [CrossRef]
- Campbell, J.; Shearer, C.A. Annulusmagnus and Ascitendus, two new genera in the Annulatascaceae. Mycologia 2004, 96, 822–833. [Google Scholar] [CrossRef]
- Bhat, D.J.; Kendrick, B. Twenty-five new conidial fungi from the Western Ghats and the Andaman Islands (India). Mycotaxon 1993, 49, 19–90. [Google Scholar]
- Bussaban, B.; Lumyong, S.; Lumyong, P.; Hyde, K.D.; McKenzie, E.H.C. Three new species of Pyricularia are isolated as zingiberaceous endophytes from Thailand. Mycologia 2003, 95, 519–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Jones, G.E.B.; Zhou, D.; Bahkali, A.H.; Hyde, K.D. Checklist of freshwater fungi in Thailand. Cryptogam. Mycol. 2011, 32, 199–217. [Google Scholar] [CrossRef]
- Perera, R. New species of Thozetella and Chaetosphaeria and new records of Chaetosphaeria and Tainosphaeria from Thailand. Mycosphere 2016, 7, 1201–1321. [Google Scholar] [CrossRef]
- Calabon, M.S.; Jones, E.B.G.; Boonmee, S.; Doilom, M.; Lumyong, S.; Hyde, K.D. Five novel freshwater Ascomycetes indicate high undiscovered diversity in lotic habitats in Thailand. J. Fungi. 2021, 7, 117. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Taxon Name | Strain Number | GenBank Accession Number | References | |
|---|---|---|---|---|
| ITS | LSU | |||
| Ascotaiwania lignicola | NIL 00005 | HQ446341 | HQ446364 | [16] |
| Ascotaiwania sawadae | SS 00051 | HQ446340 | HQ446363 | [16] |
| Bactrodesmiastrum monilioides | FMR 10756 T | NR_152539 | KF771879 | [17] |
| Bactrodesmiastrum obovatum | FMR 6482 | NR_152537 | FR870266 | [18] |
| Bactrodesmiastrum pyriforme | FMR 10747 T | NR_152536 | FR870265 | [18] |
| Bactrodesmiastrum pyriforme | FMR 11931 | HE646636 | HE646637 | [18] |
| Canalisporium caribense | SS 03839 | GQ390283 | GQ390268 | [7] |
| Canalisporium grenadoideum | BCC 20507 T | NR_111442 | GQ390267 | [7] |
| Conioscypha lignicola | CBS 335.93 | - | AY484513 | [19] |
| Conioscypha submersa | DLUCC 0904 T | NR_168820 | MK835856 | [20] |
| Conioscypha varia | CBS 436.70 | MH859785 | MH871548 | [21] |
| Fuscosporella aquatica | MFLUCC 16-0859 T | NR_156398 | NG_059853 | [3] |
| Fuscosporella pyriformis | MFLUCC 16-0570 T | NR_152555 | NG_059711 | [1] |
| Leotia lubrica | AFTOL-ID 1 | DQ491484 | AY544644 | [22] |
| Microglossum rufum | AFTOL-ID 1292 | - | DQ470981 | [23] |
| Mucispora infundibulata | MFLUCC 16-0866 T | NR_171733 | NG_073625 | [24] |
| Mucispora obscuriseptata | MFLUCC 15-0618 T | NR_152556 | NG_059709 | [1] |
| Mucispora phangngaensis | MFLUCC 16-0865 T | NR_156399 | NG_059854 | [3] |
| Parafuscosporella aquatica | KUMCC 19-0211 T | MN513034 | MN512343 | [4] |
| Parafuscosporella ellipsoconidiogena | TBRC 15503 T | OK044749 | OK044741 | This study |
| Parafuscosporella ellipsoconidiogena | TBRC 15504 | OK044750 | OK044742 | This study |
| Parafuscosporella garethii | TBRC 6543 T | OK135602 | KX958430 | [2] |
| Parafuscosporella garethii | TBRC 6544 | OK135603 | KX958431 | [2] |
| Parafuscosporella moniliformis | MFLUCC 15-0626 T | NR_152557 | NG_059710 | [1] |
| Parafuscosporella mucosa | MFLUCC 16-0571 T | NR_152554 | NG_059855 | [1] |
| Parafuscosporella obovata | TBRC 15505 T | OK044751 | OK044743 | This study |
| Parafuscosporella pyriformis | MFLUCC 18-1400 T | MN513031 | MN512340 | [4] |
| Parafuscosporella pyriformis | KUMCC 19-0008 | MN513030 | MN512339 | [4] |
| Phaeoisaria fasciculata | CBS 127885 T | NR_145395 | NG_064241 | [25] |
| Phaeoisaria sedimenticola | CGMCC 3.14949 T | JQ074237 | JQ031561 | [26] |
| Pleurotheciella centenaria | DAOM 229631 T | NR_111709 | NG_060098 | [27] |
| Pleurotheciella rivularia | CBS 125238 T | JQ429160 | JQ429232 | [27] |
| Pleurothecium recurvatum | CBS 138747 | KT278728 | KT278714 | [25] |
| Pleurothecium semifecundum | CBS 131271 T | JQ429159 | JQ429240 | [27] |
| Pseudoascotaiwania persoonii | A57-14C | - | AY590295 | [28] |
| Savoryella aquatica | SS 03801 | - | HQ446372 | [16] |
| Savoryella lignicola | NF 00204 | - | HQ446378 | [16] |
| Vanakripa minutiellipsoidea | CBS 112523 | MH862895 | MH874467 | [21] |
| Species | Conidioma | Conidiophore | Conidiogenous Cell | Conidium | Habitat and Geographical Distribution | Reference |
|---|---|---|---|---|---|---|
| P. aquatica | Sporodochial without jelly-like covering | Mostly globose to subglobose in moniliform | Globose to subglobose, 7–14 × 8–11 μm | Ellipsoidal to obovoid, 1-septate, apical cell dark brown to black, basal cell paler, 20–29 × 13–19 μm | Decaying submerged wood, Mukdahan, Thailand | [4] |
| P. ellipsoconidiogena | Sporodochial with jelly-like covering | Mostly doliiform, ellipsoidal, fusiform in moniliform, with each | Doliiform, ellipsoidal, fusiform, 7.9 – 24.3 × 5.1 – 9.6 μm | Ellipsoidal to obovoid, 1-septate, apical cell dark brown to black, basal cell light brown, 27.5 – 33 × 15 – 20 μm | Submerged twigs, Chiang Mai, Thailand | This study |
| P. garethii | Sporodochial with jelly-like covering | Cylindrical in single or mostly globose to subglobose in moniliform | Cylindrical, 1.25–2.5 μm wide, mostly globose to subglobose, 8–12.5 μm diam., or ellipsoidal, 10–15 × 7.5–8 μm | Obpyramidal, coronate apex with 4–9 conical projections, 5 –7.5 × 5 μm, 1–2-septate, distal cell black, lower cells light brown, 37.5–47.5 × 25–42.5 µm | Decaying submerged wood, Chiang Mai, Thailand | [2] |
| P. moniliformis | Sporodochial without jelly-like covering | Mostly globose to subglobose, ellipsoidal or clavate in moniliform | Globose, subglobose, ellipsoidal or clavate, 5.5–36 × 5–21 μm | Ellipsoidal to broadly obpyriform, 1-septate, dark brown to black, basal cell pale brown, 28–37 × 14–21 μm | Decaying submerged wood, Prachuap Khiri Khan, Thailand | [1] |
| P. mucosa | Sporodochial with jelly-like covering | Cylindrical in single | Globose, subglobose, ellipsoidal or clavate, 7–17 × 4–12 μm | Obovoid to obpyriform, 1-septate, brown to dark brown, basal cell paler, 26.5–36 × 12–26 μm | Decaying submerged wood, Prachuap Khiri Khan, Thailand | [1] |
| P. obovata | Sporodochial without jelly-like covering | Mostly globose to subglobose or ellipsoidal in moniliform | Globose to subglobose, 9.5–11.2 μm diam., or obovoid, 9.6–10.1 × 7.1–7.8 μm | Obovoid, broadly obovoid to subglobose, 1-septate, apical cell dark brown to black, basal cell light brown, 22.5 – 36.3 × 13 – 32.5 μm | Submerged twigs, Chiang Mai, Thailand | This study |
| P. pyriformis | Sporodochial with jelly-like covering | Cylindrical in single or globose to subglobose or ellipsoidal in moniliform | Cylindrical to clavate, 2–3 × 0.5–1 μm, or globose to subglobose, 8–13 μm diam. | Obovoid to obpyriform, 1–2 septate, dark brown to black, basal cells brown, 23–30 × 16–26 μm | Decaying submerged wood, Nakhon Si Thammarat, Thailand (Holotype); Yunnan, China (Paratype) | [4] |
| Species | Colony | Conidiogenous Cell | Conidium | Reference | |
|---|---|---|---|---|---|
| Shape and Colour | Size | ||||
| P.aquatica | Brown, dense and tight mycelia, sparse margin | Integrated | Obovoid to obpyriform, mostly 1-septate, brown to dark brown | 16–24 × 9–16 μm | [4] |
| P. ellipsoconidiogena | Brown with beige-brown patches, with a dark brown outer zone, flat, circular, velvety, serrate margin | Integrated, cylindrical, ellipsoidal, 3.75–12.5 × 3.75–6.4 μm | Broadly obpyriform, ellipsoidal, obovoid, 0 – 1-septate, medium brown to dark brown | 15–27.5 × 10.5–17.5 μm | This study |
| P. garethii | Rounded, floccose, grey to dark grey | Integrated, cylindrical | Obovoid to obpyriform, 1‒2-septate, upper cell(s) brown to dark brown, basal cell light brown | 22.5–30 × 15–25 μm | [2] |
| P. moniliformis | Dark brown, irregularly layered (on MEA) | Integrated or cylindrical, subglobose or dumbbell-shaped, 5–15 × 2–10 μm | Globose to subglobose, 0–1-septate, medium brown to dark brown | 15.5–24.5 × 13–18.5 μm | [1] |
| P. mucosa | Dark brown, irregular, sparse aerial hyphae, undulate margin, producing chlamydospores | Integrated, doliiform or obovoid, 4–9.5 × 2–5 μm | Globose to subglobose, 0–1-septate, olivaceous to pale brown | 16.5–29 × 13–19 μm | [1] |
| P. obovata | Olivaceous brown with a beige-brown outer zone, raised, circular, lanose, floccose, entire margin | Integrated, cylindrical | Ellipsoidal, obovoid to broadly obovoid, obpyriform, 0 – 1-septate, brown to dark brown | 11–17.5 × 7.5–13.8 μm | This study |
| P. pyriformis | Grey to dark grey, rounded, floccose, undulate margin | Integrated or cylindrical, 1.5–3 μm wide | Globose to subglobose, sometimes moniliform, aseptate, light brown to brown | 8–12 × 7–12 μm | [4] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boonyuen, N.; Chuaseeharonnachai, C.; Nuankaew, S.; Kwantong, P.; Pornputtapong, N.; Suwannarach, N.; Jones, E.B.G.; Somrithipol, S. Novelties in Fuscosporellaceae (Fuscosporellales): Two New Parafuscosporella from Thailand Revealed by Morphology and Phylogenetic Analyses. Diversity 2021, 13, 517. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110517
Boonyuen N, Chuaseeharonnachai C, Nuankaew S, Kwantong P, Pornputtapong N, Suwannarach N, Jones EBG, Somrithipol S. Novelties in Fuscosporellaceae (Fuscosporellales): Two New Parafuscosporella from Thailand Revealed by Morphology and Phylogenetic Analyses. Diversity. 2021; 13(11):517. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110517
Chicago/Turabian StyleBoonyuen, Nattawut, Charuwan Chuaseeharonnachai, Salilaporn Nuankaew, Papichaya Kwantong, Natapol Pornputtapong, Nakarin Suwannarach, E. B. Gareth Jones, and Sayanh Somrithipol. 2021. "Novelties in Fuscosporellaceae (Fuscosporellales): Two New Parafuscosporella from Thailand Revealed by Morphology and Phylogenetic Analyses" Diversity 13, no. 11: 517. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110517