
Aquatic Habitat Bird Occurrences at Photovoltaic Solar Energy Development in Southern California, USA


Abstract
:1. Introduction
2. Materials and Methods
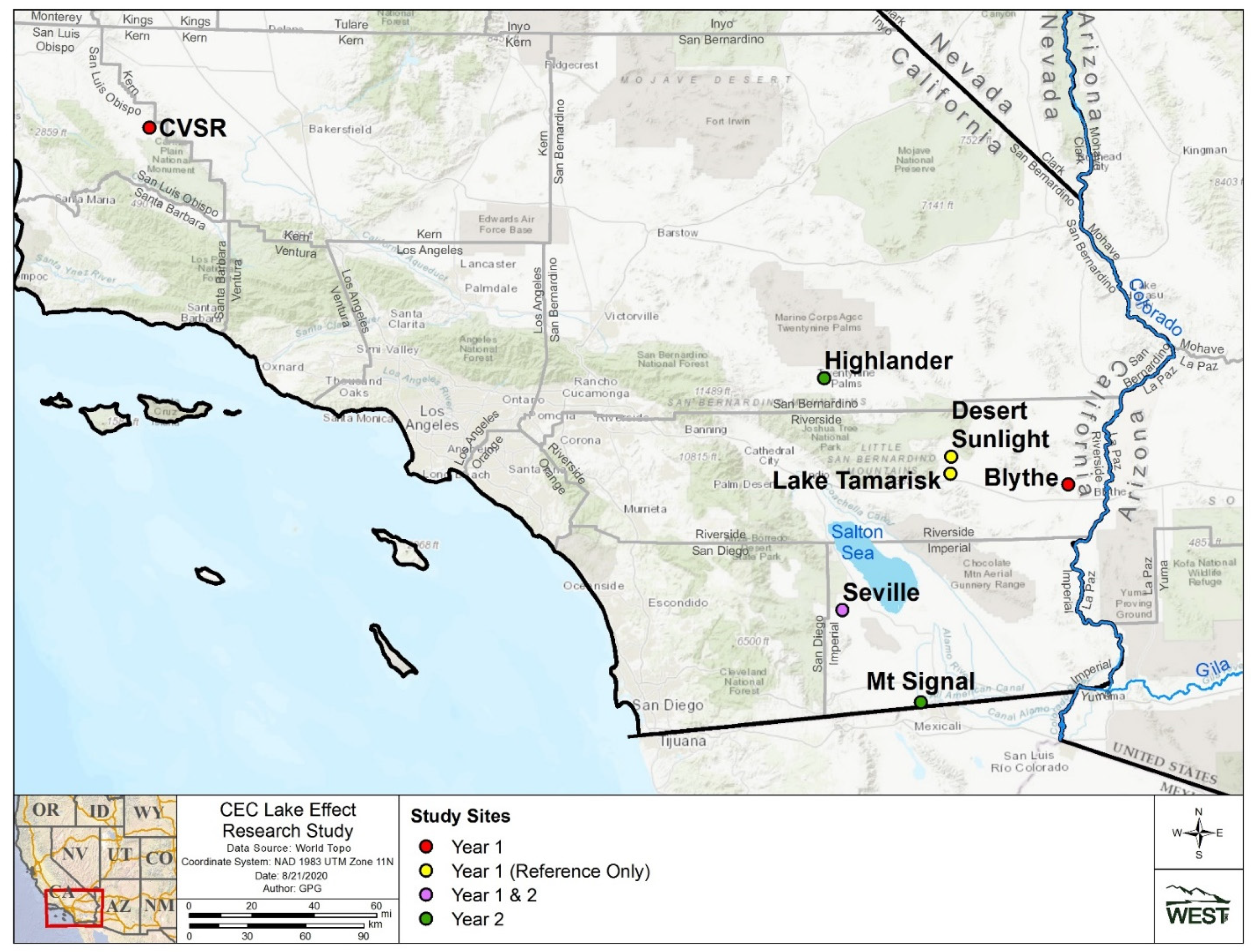
2.1. Study Sites
2.2. Fixed-Point Count Surveys
2.3. Fatality Surveys
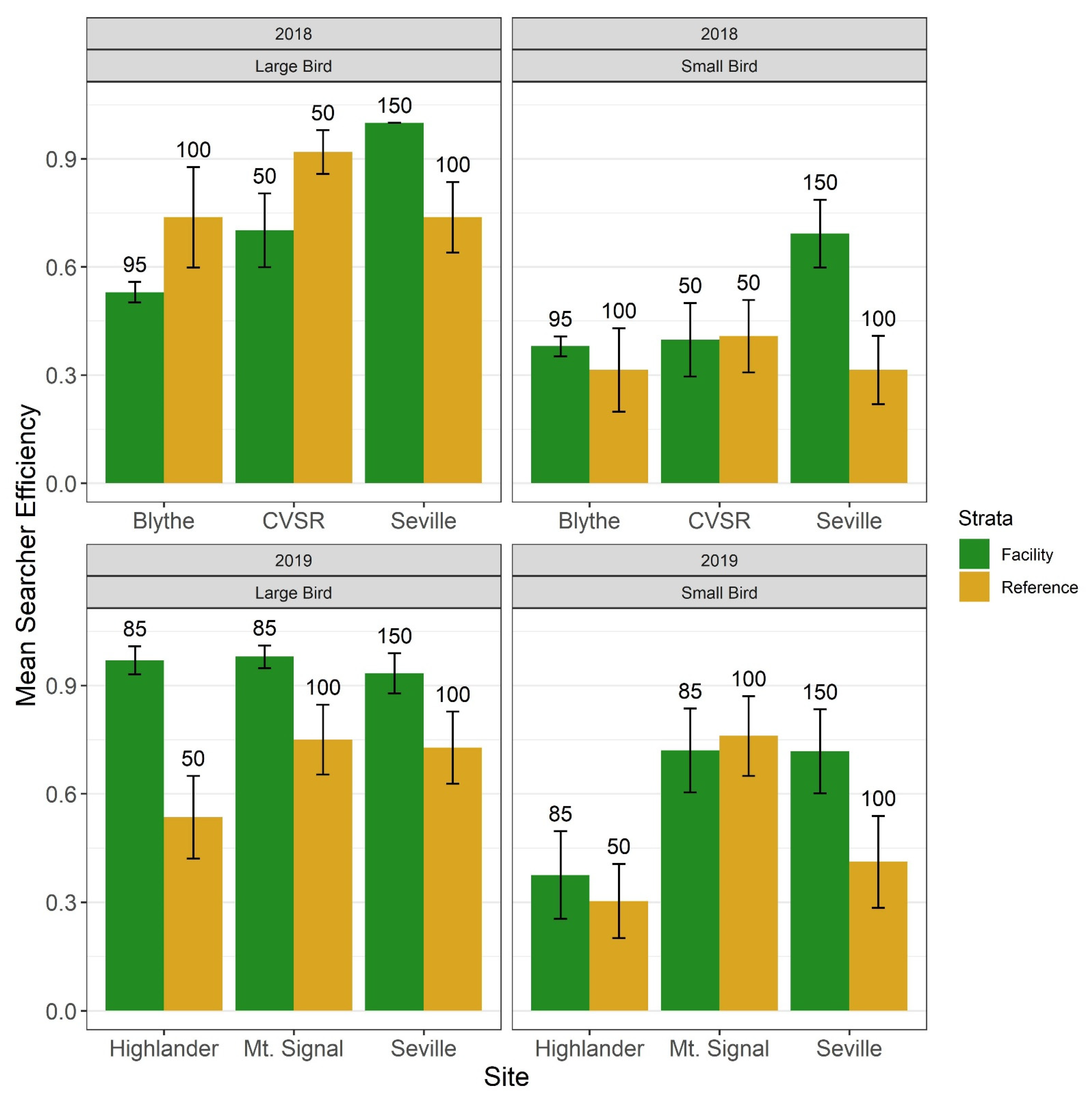
2.4. Searcher Efficiency Trials
2.5. Data Analysis
3. Results
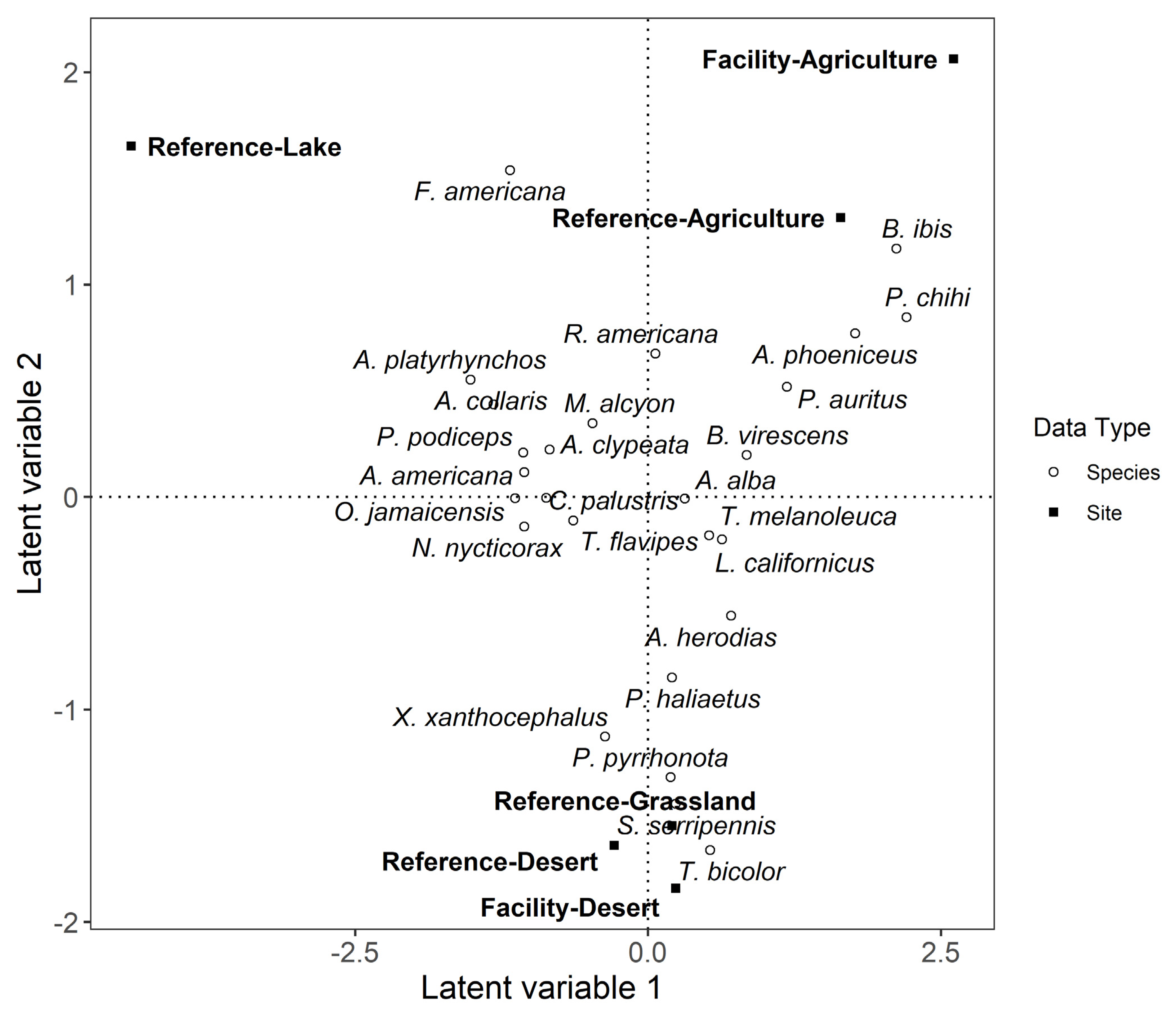
3.1. Species Richness and Community Association of Live Aquatic Habitat Birds at Photovoltaic Solar and Reference Areas
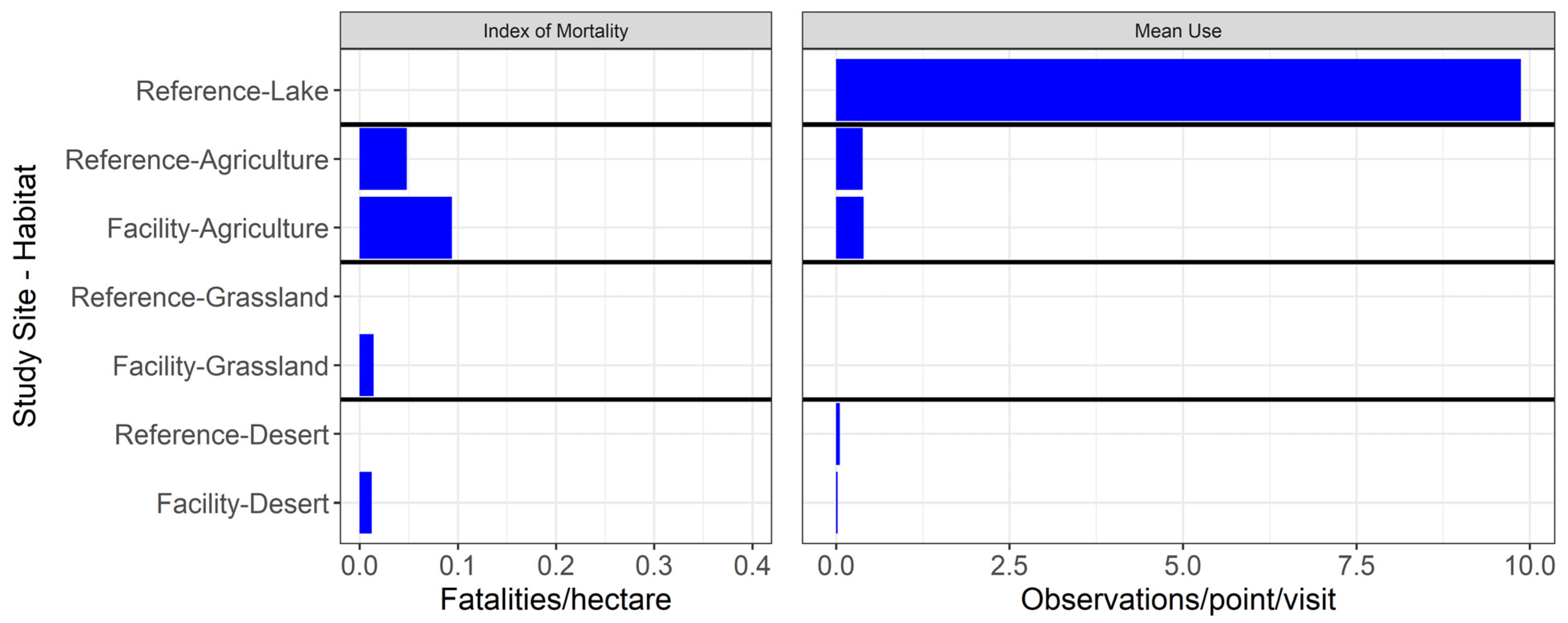
3.2. Mortality of Aquatic Habitat Birds at Photovoltaic Solar and Reference Areas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lovich, J.E.; Ennen, J.R. Wildlife conservation and solar energy development in the desert Southwest, United States. Biosci 2011, 61, 982–992. [Google Scholar] [CrossRef] [Green Version]
- Guerra, K.; Dahm, K.; Dundorf, S. Oil and Gas Produced Water Management and Beneficial Use in the Western United States; US Department of the Interior, Bureau of Reclamation Denver: Lakewood, CO, USA, 2011. Available online: https://www.usbr.gov/research/dwpr/reportpdfs/report157.pdf (accessed on 31 December 2020).
- Kagan, R.A.; Viner, T.C.; Trail, P.W.; Espinoza, E.O. Avian Mortality at Solar Energy Facilities in Southern California: A Preliminary Analysis; National Fish and Wildlife Forensics Laboratory, U.S. Fish and Wildlife Service: Ashland, OR, USA, 2014.
- Kosciuch, K.; Riser-Espinoza, D.; Gerringer, M.; Erickson, W. A summary of bird mortality at photovoltaic utility scale solar facilities in the southwestern U.S. PLoS ONE 2020, 15, e0232034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loss, S.R.; Will, T.; Loss, S.S.; Mara, P.P. Bird–building collisions in the United States: Estimates of annual mortality and species vulnerability. Condor 2014, 116, 8–23. [Google Scholar] [CrossRef] [Green Version]
- Gehring, J.; Kerlinger, P.; Manville, A.M., II. The role of tower height and guy wires on avian collisions with communication towers. J. Wildl. Manag. 2011, 75, 848–855. [Google Scholar] [CrossRef]
- Loss, S.R.; Will, T.; Marra, P.P. Estimates of Bird Collision Mortality at Wind Facilities in the Contiguous United States. Biol. Conserv. 2013, 168, 201–209. [Google Scholar] [CrossRef]
- Erickson, W.P.; Wolfe, M.M.; Bay, K.J.; Johnson, D.H.; Gehring, J.L. A comprehensive analysis of small passerine fatalities from collisions with turbines at wind energy facilities. PLoS ONE 2014, 9, 107491. [Google Scholar] [CrossRef] [PubMed]
- Solar Farms Threaten Birds. Available online: https://www.scientificamerican.com/article/solar-farms-threaten-birds/ (accessed on 27 July 2021).
- Conkling, T.J.; Loss, S.R.; Diffendorfer, J.E.; Duerr, A.E.; Katzner, T.E. Limitations, lack of standardization, and recommended best practices in studies of renewable energy effects on birds and bats. Conserv. Biol. 2020, 35, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Jin, S.; Danielson, P.; Homer, C.; Gass, L.; Bender, S.M.; Case, A.; Costello, C.; Dewitz, J.; Fry, J.; et al. A new generation of the United States National Land Cover Database: Requirements, research priorities, design, and implementation strategies. ISPRS J. Photogramm. Remote Sens. 2018, 146, 108–123. [Google Scholar] [CrossRef]
- Multi-Resolution Land Characteristics Consortium. National Land Cover Database (NLCD) 2016; Multi-Resolution Land Characteristics Consortium: Sioux Falls, SD, USA, 2019.
- Ralph, C.J.; Sauer, J.R.; Droege, S. Monitoring Bird Populations by Point Counts; Pacific Southwest Research Station, US Department of Agriculture, Forest Service: Albany, CA, USA, 1995. Available online: https://www.fs.fed.us/psw/publications/documents/psw_gtr149/ (accessed on 27 July 2021).
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L.J. An Introduction to Distance Sampling: Estimating Abundance of Biological Populations; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Advanced Distance Sampling; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Huso, M.; Dietsch, T.; Nicolai, C. Mortality Monitoring Design for Utility-Scale Solar Power Facilities; U.S. Geological Survey: Reston, VA, USA, 2016. Available online: https://pubs.er.usgs.gov/publication/ofr20161087 (accessed on 27 July 2021). [CrossRef] [Green Version]
- H.T. Harvey & Associates. Ivanpah Solar Electric Generating System Avian & Bat Monitoring Plan. 2013–2014 Annual Report (Revised): (29 October 2013–20 October 2014); H.T. Harvey & Associates: Los Gatos, CA, USA, 2015. Available online: https://efiling.energy.ca.gov/GetDocument.aspx?tn=204258&DocumentContentId=2191 (accessed on 27 July 2021).
- Martinson, L.; Gerringer, M.; Lombardi, J.; (Western EcoSystems Technology, Inc., Cheyenne, WY, USA). Post-construction monitoring at the Blythe Solar Power Project, Riverside County, California. Second annual report draft: 2017–2018. Personal communication, 2019. [Google Scholar]
- Western EcoSystems Technology, Inc. Ivanpah Solar Electric Generating System Avian & Bat Monitoring Plan: 2014–2015 Annual Report, and Two Year Comparison: 21 October 2014–20 October 2015; Western EcoSystems Technology, Inc.: Cheyenne, WY, USA, 2016. [Google Scholar]
- Western EcoSystems Technology, Inc. Ivanpah Solar Electric Generating System Avian & Bat Monitoring Plan: 2015–2016 Annual Report; Western EcoSystems Technology, Inc.: Cheyenne, WY, USA, 2017. Available online: https://efiling.energy.ca.gov/Lists/DocketLog.aspx?docketnumber=07-AFC-05C (accessed on 27 July 2021).
- Western EcoSystems Technology, Inc.; (Cheyenne, WY, USA). Post-construction monitoring at the Blythe Solar Power Project, Riverside County, California. First annual report draft: 2016–2017. Personal communication, 2018. [Google Scholar]
- U.S. Fish and Wildlife Service. Land-Based Wind Energy Guidelines; U.S. Fish and Wildlife Service: Washington, DC, USA, 2012. Available online: http://www.fws.gov/cno/pdf/Energy/2012_Wind_Energy_Guidelines_final.pdf (accessed on 27 July 2021).
- Caplat, P.; Fonderflick, J. Area mediated shifts in bird community composition: A study on a fragmented Mediterranean grassland. Biodivers. Conserv. 2009, 18, 2979–2995. [Google Scholar] [CrossRef]
- Visser, E.; Perold, V.; Ralston-Paton, S.; Cardenal, A.C.; Ryan, P.G. Assessing the impacts of a utility-scale photovoltaic solar energy facility on birds in the Northern Cape, South Africa. Renew. Energy 2019, 133, 1285–1294. [Google Scholar] [CrossRef]
- Cornell Lab of Ornithology. All about Birds. Available online: https://www.allaboutbirds.org (accessed on 31 December 2020).
- Chao, A. Estimating the population size for capture-recapture data with unequal catchability. Biometrics 1987, 43, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.H.; Wang, Y.T.; Walther, B.A.; Chao, A. Improved nonparametric lower bound of species richness via a modified Good-Turing frequency formula. Biometrics 2014, 70, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Vegan: Community Ecology Package (version 2.5-7). Available online: https://CRAN.R-project.org/package=vegan (accessed on 17 September 2021).
- Boral: Bayesian Ordination and Regression Analysis (version 2.0). Available online: https://cran.r-project.org/web/packages/boral/index.html (accessed on 17 September 2021).
- Hui, F.K.C. Boral: Bayesian ordination and regression analysis of multivariate abundance data in R. Methods Ecol. Evol. 2016, 7, 744–750. [Google Scholar] [CrossRef] [Green Version]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L. Distance Sampling: Estimating Abundance of Biological Populations; Chapman & Hall: London, UK, 1993. [Google Scholar]
- LaPorte, N.; Storer, R.W.; Nuechterlein, G.L. Western Grebe (Aechmophorus occidentalis), version 1.0. In Birds of the World; Rodewald, P.G., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Higgs, E.; Harris, J.A. Novel ecosystems: Implications for conservation and restoration. Trends Ecol. Evol. 2009, 24, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Robertson, B.A.; Blumstein, D.T. How to disarm and evolutionary trap. Conserv. Sci. Pract. 2019, 1, e116. [Google Scholar] [CrossRef] [Green Version]
- Shuford, W.D.; Warnock, N.; Molina, K.C.; Sturm, K.K. The Salton Sea as critical habitat to migratory and resident waterbirds. In The Salton Sea: Developments in Hydrobiology; Barnum, D.A., Elder, J.F., Stephens, D., Friend, M., Eds.; Springer: Dordrecht, The Netherlands, 2002; Volume 161, pp. 255–274. [Google Scholar] [CrossRef]
- Josephson, J.; Josephson, S. Abductive Inference; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Kosciuch, K.L.; Parker, T.H.; Sandercock, B.K. Nest desertion by a cowbird host: An antiparasite behavior or a response to egg loss? Behav. Ecol. 2006, 17, 917–924. [Google Scholar] [CrossRef] [Green Version]
- Robertson, B.A.; Chalfoun, A.D. Evolutionary traps as keys to understanding behavioral maladaptation. Curr. Opin. Behav. Sci. 2016, 12, 12–17. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aquatic Habitat Type (2016 NLCD) | Desert/Scrub | Grassland | Agricultural | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Blythe | Highlander II | Seville | Reference A | Lake Tamarisk | CVSR | Mt. Signal 3 | Total | |||||||||
| Area | % | Area | % | Area | % | Area | % | Area | % | Area | % | Area | % | Area | % | |
| Barren Land | 2929.50 | 16.59 | 8898.36 | 60.88 | 14,977.13 | 95.63 | 1811.56 | 10.95 | 52.43 | 0.64 | 556.29 | 3.25 | 2885.69 | 10.58 | 14,977.13 | 27.43% |
| Cultivated Crops | 117.40 | 0.66 | 0.00 | 118.28 | 0.76 | - | 0.00 | - | 0.00 | 161.68 | 0.95 | 12,124.00 | 44.46 | 118.28 | 10.70% | |
| Deciduous Forest | - | 0.00 | 0.90 | 0.01 | - | 0.00 | - | 0.00 | - | 0.00 | - | 0.00 | 0.54 | 0.00 | - | 0.00% |
| Developed, High Intensity | 17.04 | 0.10 | 25.83 | 0.18 | 0.63 | 0.00 | 27.44 | 0.17 | 3.51 | 0.04 | 0.84 | 0.00 | 91.35 | 0.33 | 0.63 | 0.14% |
| Developed, Low Intensity | 300.37 | 1.70 | 290.43 | 1.99 | 29.18 | 0.19 | 632.00 | 3.82 | 91.96 | 1.12 | 11.44 | 0.07 | 1157.39 | 4.24 | 29.18 | 2.15% |
| Developed, Medium Intensity | 71.73 | 0.41 | 80.55 | 0.55 | 82.25 | 0.53 | 198.59 | 1.20 | 32.44 | 0.40 | 1.39 | 0.01 | 540.68 | 1.98 | 82.25 | 0.86% |
| Developed, Open Space | 1107.89 | 6.27 | 1591.03 | 10.89 | 99.80 | 0.64 | 625.56 | 3.78 | 98.89 | 1.21 | 2013.04 | 11.77 | 1612.27 | 5.91 | 99.80 | 6.11% |
| Emergent Herbaceous Wetlands | - | 0.00 | - | 0.00 | 0.72 | 0.00 | - | 0.00 | - | 0.00 | 12.58 | 0.07 | 88.19 | 0.32 | 0.72 | 0.09 |
| Evergreen Forest | - | 0.00 | 0.09 | 0.00 | - | 0.00 | - | 0.00 | - | 0.00 | - | 0.00 | 2.70 | 0.01 | - | 0.00 |
| Hay/Pasture | 11.97 | 0.07 | 2.07 | 0.01 | 8.63 | 0.06 | - | 0.00 | 9.00 | 0.11 | 0.36 | 0.00 | 1398.05 | 5.13 | 8.63 | 1.22 |
| Herbaceous | 715.57 | 4.05 | 44.62 | 0.31 | 0.18 | 0.00 | 154.85 | 0.94 | 17.59 | 0.21 | 13,541.04 | 79.15 | 20.33 | 0.07 | 0.18 | 12.38 |
| Open Water | 0.09 | 0.00 | 24.91 | 0.17 | 0.63 | 0.00 | 3.15 | 0.02 | 6.30 | 0.08 | 0.09 | 0.00 | 45.46 | 0.17 | 0.63 | 0.07 |
| Shrub/Scrub | 12,389.26 | 70.15 | 3654.09 | 25.00 | 329.55 | 2.10 | 13,089.43 | 79.13 | 7894.04 | 96.20 | 808.78 | 4.73 | 1324.14 | 4.86 | 329.55 | 33.73 |
| Unclassified | - | 0.00 | - | 0.00 | - | 0.00 | - | 0.00 | - | 0.00 | 0.00 | 5506.45 | 20.19 | 4.70 | ||
| Woody Wetlands | - | 0.00 | 2.70 | 0.02 | 14.62 | 0.09 | - | 0.00 | - | 0.00 | 0.00 | 474.59 | 1.74 | 14.62 | 0.42 | |
| Total | 17,660.82 | 100.00 | 14,615.57 | 100.00 | 15,661.60 | 100.00 | 16,542.57 | 100.00 | 8206.15 | 100.00 | 17,107.52 | 100.00 | 27,271.83 | 100.00 | 15,661.60 | 100.00 |
| Species Name | Habitat and Study Site 1 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Desert/Scrub | Grassland | Agricultural | Total | ||||||||||
| B-F | B-R | H-F | H-R | S-F | S-R | RA-R | LT-R | CVSR-F | CVSR-R | Mt. S-F | Mt. S-R | ||
| American coot Fulica americana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1686 | 0 | 0 | 0 (1) | 0 (1) | 1686 (2) |
| white-faced ibis Plegadis chihi | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 (1) | 0 | 300 | 565 (1) | 865 (2) |
| cattle egret Bubulcus ibis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 315 | 14 (1) | 329 (1) |
| red-winged blackbird Agelaius phoeniceus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 80 (1) | 240 | 320 (1) |
| mallard Anas platyrhynchos | 0 | 0 | 0 | 0 | 0 (1) | 0 | 0 | 264 | 0 | 0 | 0 | 0 | 264 (1) |
| ring-necked duck Aythya collaris | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 241 | 0 | 0 | 0 | 0 | 241 |
| ruddy duck Oxyura jamaicensis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 119 | 0 | 0 | 0 | 0 | 119 |
| black-crowned night-heron Nycticorax nycticorax | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 60 | 0 | 0 | 0 | 0 | 61 |
| tree swallow Tachycineta bicolor | 4 | 6 | 0 | 0 | 23 | 13 | 0 | 0 | 0 | 5 | 0 | 3 | 54 |
| pied-billed grebe Podilymbus podiceps | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 53 | 0 | 0 | 0 | 0 | 53 |
| great egret Ardea alba | 0 | 0 | 1 | 0 | 0 | 4 | 0 | 6 | 0 | 0 | 6 (1) | 31 (1) | 48 (2) |
| northern rough-winged swallow Stelgidopteryx serripennis | 1 | 5 | 0 | 0 | 5 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 13 |
| double-crested cormorant Phalacrocorax auritus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 13 | 0 | 13 |
| American wigeon Mareca americana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 0 | 0 | 0 | 0 | 12 |
| yellow-headed blackbird Xanthocephalus xanthocephalus | 0 | 0 | 1 | 10 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 12 |
| northern shoveler Spatula clypeata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 0 | 0 | 0 | 0 | 9 |
| cliff swallow Petrochelidon pyrrhonota | 0 | 0 | 0 | 0 | 2 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 7 |
| belted kingfisher Megaceryle alcyon | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 1 | 6 |
| great blue heron Ardea herodias | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 (1) | 4 (1) | 5 (2) |
| marsh wren Cistothorus palustris | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 4 |
| American avocet Recurvirostra americana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 2 |
| lesser yellowlegs Tringa flavipes | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| osprey Pandion haliaetus | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| California gull Larus californicus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| green heron Butorides virescens | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| greater yellowlegs Tringa melanoleuca | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| blue-winged teal Spatula discors | 0 (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 (1) |
| common loon Gavia immer | 0 | 0 | 0 | 0 | 0 (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 (1) |
| western grebe Aechmophorus occidentalis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 (1) | 0 (1) |
| sora Porzana carolina | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 (1) | 0 | 0 (1) |
| Total | 5 (1) | 12 | 3 | 11 | 30 (2) | 24 | 0 | 2463 | 0 (1) | 5 | 715 (5) | 860 (6) | 4128 (15) |
| Reference-Lake | Facility-Desert | Reference-Desert | Facility-Agriculture | Reference-Agriculture | Facility-Grassland | Reference-Grassland | |
|---|---|---|---|---|---|---|---|
| Species richness | 14 | 6 | 7 | 6 | 9 | 0 | 1 |
| Chao’s estimator | 14.5 | 7.5 | 8 | 6 | 12 | NA | 1 |
| Standard deviation | 1.28 | 2.54 | 2.24 | 0.46 | 4.48 | NA | 0 |
| 90% Confidence interval | 14.05–19.20 | 6.22–16.16 | 7.11–16.04 | 6–NA | 9.51–26.81 | NA | NA |
| Facility-Desert | Reference-Desert | Facility-Agriculture | Reference-Agriculture | Facility-Grassland | Reference-Grassland | |
|---|---|---|---|---|---|---|
| Species richness | 3 | 0 | 5 | 6 | 1 | 0 |
| Chao’s estimator | 6 | NA | 15 | 21 | 1 | NA |
| Standard deviation | 4.29 | NA | 10.04 | 13.46 | 0 | NA |
| 90% Confidence interval | 3.53–20.00 | NA | 7.53–44.49 | 10.24–59.09 | NA | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosciuch, K.; Riser-Espinoza, D.; Moqtaderi, C.; Erickson, W. Aquatic Habitat Bird Occurrences at Photovoltaic Solar Energy Development in Southern California, USA. Diversity 2021, 13, 524. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110524
Kosciuch K, Riser-Espinoza D, Moqtaderi C, Erickson W. Aquatic Habitat Bird Occurrences at Photovoltaic Solar Energy Development in Southern California, USA. Diversity. 2021; 13(11):524. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110524
Chicago/Turabian StyleKosciuch, Karl, Daniel Riser-Espinoza, Cyrus Moqtaderi, and Wallace Erickson. 2021. "Aquatic Habitat Bird Occurrences at Photovoltaic Solar Energy Development in Southern California, USA" Diversity 13, no. 11: 524. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110524