
Effects of Caudal Autotomy on the Locomotor Performance of Micrablepharus Atticolus (Squamata, Gymnophthalmidae)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Lizard Sampling
2.3. Locomotor Performance
2.4. Statistical Analyses
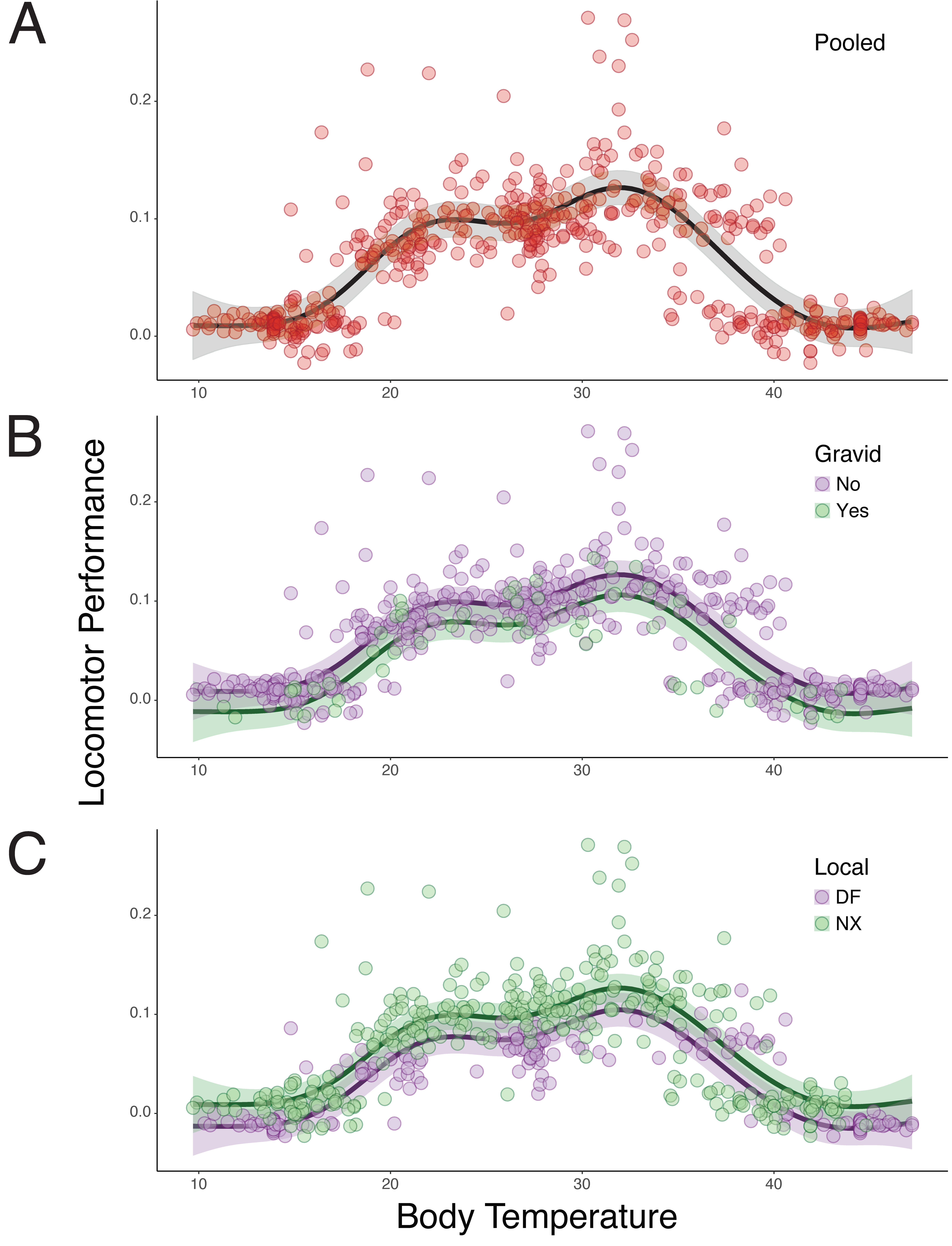
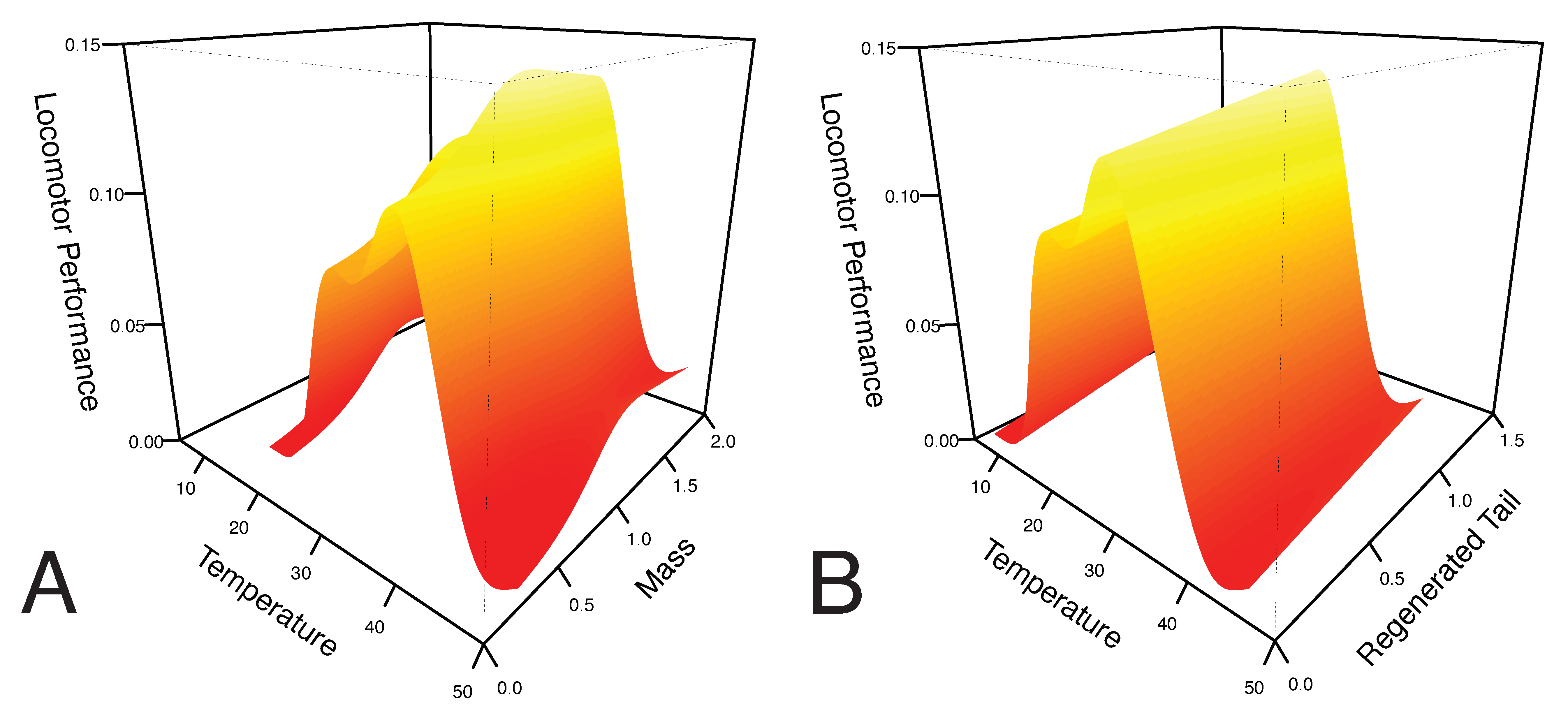
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dawkins, R.; Krebs, J.R. Arms races between and within species. Proc. R. Soc. Lond. B Biol. Sci. 1979, 205, 489–511. [Google Scholar] [PubMed]
- Emberts, Z.; Escalante, I.; Bateman, P.W. The ecology and evolution of autotomy. Biol. Rev. 2019, 94, 1881–1896. [Google Scholar] [CrossRef]
- LeBlanc, A.R.H.; MacDougall, M.J.; Haridy, Y.; Scott, D.; Reisz, R.R. Caudal autotomy as anti-predatory behaviour in Palaeozoic reptiles. Sci. Rep. 2018, 8, 3328. [Google Scholar] [CrossRef]
- Arnold, E.N. Caudal autotomy as a defense. In Biology of the Reptilia. Ecology B: Defense and Life History; Gans, C., Huey, R.B., Eds.; Alan R. Liss, Inc.: New York, NY, USA, 1988; Volume 16. [Google Scholar]
- Arnold, E.N. Evolutionary aspects of tail shedding in lizards and their relatives. J. Nat. Hist. 1984, 18, 127–169. [Google Scholar] [CrossRef]
- Clause, A.R.; Capaldi, E.A. Caudal autotomy and regeneration in lizards. J. Exp. Zool. Part A 2006, 305, 965–973. [Google Scholar] [CrossRef]
- Bellairs, A.d.A.; Bryant, S.V. Autotomy and regeneration in reptiles. In Biology of the Reptilia, Volume 15, Development B; Gans, C., Billet, F., Eds.; John Wiley & Sons: New York, NY, USA, 1985; pp. 301–410. [Google Scholar]
- Gilbert, E.A.B.; Payne, S.L.; Vickaryous, M.K. The anatomy and histology of caudal autotomy and regeneration in lizards. Physiol. Biochem. Zool. 2013, 86, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Ritzman, T.B.; Stroik, L.K.; Julik, E.; Hutchins, E.D.; Lasku, E.; Denardo, D.F.; Wilson-Rawls, J.; Rawls, J.A.; Kusumi, K.; Fisher, R.E. The gross anatomy of the original and regenerated tail in the green anole (Anolis carolinensis). Anat. Rec. 2012, 295, 1596–1608. [Google Scholar] [CrossRef]
- Alibardi, L. Development of the axial cartilaginous skeleton in the regenerating tail of lizards. Bull. Assoc. Anat. 1995, 79, 3–9. [Google Scholar]
- Fisher, R.E.; Geiger, L.A.; Stroik, L.K.; Hutchins, E.D.; George, R.M.; Denardo, D.F.; Kusumi, K.; Rawls, J.A.; Wilson-Rawls, J. A histological comparison of the original and regenerated tail in the green anole, Anolis carolinensis. Anat. Rec. 2012, 295, 1609–1619. [Google Scholar] [CrossRef] [Green Version]
- Maginnis, T.L. The costs of autotomy and regeneration in animals: A review and framework for future research. Behav. Ecol. 2006, 17, 857–872. [Google Scholar] [CrossRef] [Green Version]
- Lynn, S.E.; Borkovic, B.P.; Russell, A.P. Relative apportioning of resources to the body and regenerating tail in juvenile leopard geckos (Eublepharis macularius) maintained on different dietary rations. Physiol. Biochem. Zool. 2013, 86, 659–668. [Google Scholar] [CrossRef]
- Bateman, P.W.; Fleming, P.A. To cut a long tail short: A review of lizard caudal autotomy studies carried out over the last 20 years. J. Zool. 2009, 277, 1–14. [Google Scholar] [CrossRef]
- Cooper, W.E., Jr.; Smith, C.S. Costs and economy of autotomy for tail movement and running speed in the skink Trachylepis maculilabris. Can. J. Zool. 2009, 87, 400–406. [Google Scholar] [CrossRef]
- Cromie, G.L.; Chapple, D.G. Impact of tail loss on the behaviour and locomotor performance of two sympatric Lampropholis skink species. PLoS ONE 2012, 7, e34732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillis, G.B.; Bonvini, L.A.; Irschick, D.J. Losing stability: Tail loss and jumping in the arboreal lizard Anolis carolinensis. J. Exp. Biol. 2009, 212, 604–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.L.; Cavalieri, C.N.; Rodríguez-Romero, F.; Fox, S.F. The differential effect of tail autotomy on sprint performance between the sexes in the lizard Uta stansburiana. J. Herpetol. 2012, 46, 648–652. [Google Scholar] [CrossRef]
- Huey, R.B.; Dunham, A.E.; Overall, K.L.; Newman, R.A. Variation in locomotor performance in demographically known populations of the lizard Sceloporus merriami. Physiol. Zool. 1990, 63, 845–872. [Google Scholar] [CrossRef] [Green Version]
- McElroy, E.J.; Bergmann, P.J. Tail autotomy, tail size, and locomotor performance in lizards. Physiol. Biochem. Zool. 2013, 86, 669–679. [Google Scholar] [CrossRef] [Green Version]
- Daniels, C.B. Running: An escape strategy enhanced by autotomy. Herpetologica 1983, 39, 162–165. [Google Scholar]
- Naidenov, L.A.; Allen, W.L. Tail autotomy works as a pre-capture defense by deflecting attacks. Ecol. Evol. 2021, 11, 3058–3064. [Google Scholar] [CrossRef] [PubMed]
- Vitt, L.J.; Congdon, J.D.; Dickson, N.A. Adaptive strategies and energetics of tail autotomy in lizards. Ecology 1977, 58, 326–337. [Google Scholar] [CrossRef]
- Rodrigues, M. A new species of lizard, genus Micrablepharus (Squamata: Gymnophthalmidae), from Brazil. Herpetologica 1996, 52, 535–541. [Google Scholar]
- Colli, G.R.; Bastos, R.P.; Araujo, A.F.B. The character and dynamics of the Cerrado herpetofauna. In The Cerrados of Brazil: Ecology and Natural History of a Neotropical Savannah; Oliveira, P.S., Marquis, R.J., Eds.; Columbia University Press: New York, NY, USA, 2002; Volume 1, pp. 223–241. [Google Scholar]
- Santos, M.G.; Nogueira, C.; Giugliano, L.G.; Colli, G.R. Landscape evolution and phylogeography of Micrablepharus atticolus (Squamata, Gymnophthalmidae), an endemic lizard of the Brazilian Cerrado. J. Biogeogr. 2014, 41, 1506–1519. [Google Scholar] [CrossRef]
- de Moura, M.R.; Dayrell, J.S.; de Avelar São-Pedro, V. Reptilia, Gymnophthalmidae, Micrablepharus maximiliani (Reinhardt and Lutken, 1861): Distribution extension, new state record and geographic distribution map. Check List 2010, 6, 419–426. [Google Scholar] [CrossRef] [Green Version]
- Gainsbury, A.M.; Colli, G.R. Lizard assemblages from natural Cerrado enclaves in southwestern Amazonia: The role of stochastic extinctions and isolation. Biotropica 2003, 35, 503–519. [Google Scholar] [CrossRef]
- Vieira, G.H.C.; Mesquita, D.O.; Péres, A.K., Jr.; Kitayama, K.; Colli, G.R. Lacertilia: Micrablepharus atticolus (NCN). Natural history. Herpetol. Rev. 2000, 31, 241–242. [Google Scholar]
- Vitt, L.J. An introduction to the ecology of Cerrado lizards. J. Herpetol. 1991, 25, 79–90. [Google Scholar] [CrossRef]
- Vitt, L.J.; Caldwell, J.P. Ecological observations on Cerrado lizards in Rondônia, Brazil. J. Herpetol. 1993, 27, 46–52. [Google Scholar] [CrossRef]
- de Sousa, H.C.; Soares, A.H.S.B.; Costa, B.M.; Pantoja, D.L.; Caetano, G.H.; de Queiroz, T.A.; Colli, G.R. Fire regimes and the demography of the lizard Micrablepharus atticolus (Squamata, Gymnophthalmidae) in a biodiversity hotspot. S. Am. J. Herpetol. 2015, 10, 143–156. [Google Scholar] [CrossRef]
- Dal Vechio, F.; Recoder, R.; Zaher, H.; Rodrigues, M.T. Natural history of Micrablepharus maximiliani (Squamata: Gymnophthalmidae) in a Cerrado region of northeastern Brazil. Zoologia 2014, 31, 114–118. [Google Scholar] [CrossRef] [Green Version]
- Roscito, J.G.; Nunes, P.M.S.; Rodrigues, M.T. Digit evolution in gymnophthalmid lizards. Int. J. Dev. Biol. 2014, 58, 895–908. [Google Scholar] [CrossRef] [Green Version]
- Grizante, M.B.; Brandt, R.; Kohlsdorf, T. Evolution of body elongation in gymnophthalmid lizards: Relationships with climate. PLoS ONE 2012, 7, e49772. [Google Scholar] [CrossRef] [Green Version]
- Renous, S.; Hofling, E.; Gasc, J.P. Respective role of the axial and appendicular systems in relation to the transition to limblessness. Acta Biotheor. 1998, 46, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Renous, S.; Hofling, E.; Gasc, J.P. On the rhythmical coupling of the axial and appendicular systems in small terrestrial lizards (Sauria: Gymnophthalmidae). Zool.-Anal. Complex Syst. 1999, 102, 31–49. [Google Scholar]
- Höfling, E.; Renous, S. High frequency of pauses during intermittent locomotion of small South American gymnophthalmid lizards (Squamata, Gymnophthalmidae). Phyllomedusa 2004, 3, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Sousa, H.C.; Costa, B.M.; Morais, C.J.S.; Pantoja, D.L.; de Queiroz, T.A.; Vieira, C.R.; Colli, G.R. Blue tales of a blue-tailed lizard: Ecological correlates of tail autotomy in Micrablepharus atticolus (Squamata, Gymnophthalmidae) in a Neotropical savannah. J. Zool. 2016, 299, 202–212. [Google Scholar] [CrossRef]
- Gillis, G.; Higham, T.E. Consequences of lost endings: Caudal autotomy as a lens for focusing attention on tail function during locomotion. J. Exp. Biol. 2016, 219, 2416–2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christian, K.A.; Tracy, C.R. The effect of the thermal environment on the ability of hatchling Galapagos land iguanas to avoid predation during dispersal. Oecologia 1981, 49, 218–223. [Google Scholar] [CrossRef]
- Jayne, B.C.; Bennett, A.F. Selection on locomotor performance capacity in a natural population of garter snakes. Evolution 1990, 44, 1204–1229. [Google Scholar] [CrossRef]
- Greenwald, O.E. Thermal dependence of striking and prey capture by gopher snakes. Copeia 1974, 1974, 141–148. [Google Scholar] [CrossRef]
- Webb, W.P. Body form, locomotion and foraging in aquatic vertebrates. Am. Zool. 1984, 24, 107–120. [Google Scholar] [CrossRef]
- Garland, T.; Hankins, E.; Huey, R.B. Locomotor capacity and social dominance in male lizards. Funct. Ecol. 1990, 4, 243–250. [Google Scholar] [CrossRef]
- Chapple, D.G.; Swain, R. Effect of caudal autotomy on locomotor performance in a viviparous skink, Niveoscincus metallicus. Funct. Ecol. 2002, 16, 817–825. [Google Scholar] [CrossRef]
- Dayananda, B.; Ibarguengoytia, N.; Whiting, M.J.; Webb, J.K. Effects of pregnancy on body temperature and locomotor performance of velvet geckos. J. Therm. Biol. 2017, 65, 64–68. [Google Scholar] [CrossRef]
- Cloyed, C.S.; Grady, J.M.; Savage, V.M.; Uyeda, J.C.; Dell, A.I. The allometry of locomotion. Ecology 2021, 102, e03369. [Google Scholar] [CrossRef]
- Pironon, S.; Papuga, G.; Villellas, J.; Angert, A.L.; Garcia, M.B.; Thompson, J.D. Geographic variation in genetic and demographic performance: New insights from an old biogeographical paradigm. Biol. Rev. Camb. Philos. Soc. 2017, 92, 1877–1909. [Google Scholar] [CrossRef]
- Lin, Z.H.; Ji, X. Partial tail loss has no severe effects on energy stores and locomotor performance in a lacertid lizard, Takydromus septentrionalis. J. Comp. Physiol. B-Biochem. Syst. Environ. Physiol. 2005, 175, 567–573. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; De Moraes Goncalves, J.L.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Angilletta, M.J., Jr. Thermal Adaptation: A Theoretical and Empirical Synthesis; Oxford University Press: Oxford, UK, 2009; pp. 1–302. [Google Scholar] [CrossRef]
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2017; p. 476. [Google Scholar]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. Ser. B-Stat. Methodol. 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with R; CRC Press: Boca Raton, FL, USA, 2006; Volume 16, p. 391. [Google Scholar]
- Bartón, K. MuMIn: Multi-Model Inference, R package version 1.40.4; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Leeb, H.; Potscher, B.M. Model selection and inference: Facts and fiction. Econom. Theory 2005, 21, 21–59. [Google Scholar] [CrossRef] [Green Version]
- Hjort, N.L.; Claeskens, G. Frequentist model average estimators. J. Am. Stat. Assoc. 2003, 98, 879–899. [Google Scholar] [CrossRef] [Green Version]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multi-Model Inference, 2nd ed.; Springer: New York, NY, USA, 2002; p. 496. [Google Scholar]
- Liang, H.; Zou, G.H.; Wan, A.T.K.; Zhang, X.Y. Optimal weight choice for frequentist model average estimators. J. Am. Stat. Assoc. 2011, 106, 1053–1066. [Google Scholar] [CrossRef]
- Galipaud, M.; Gillingham, M.A.F.; David, M.; Dechaume-Moncharmont, F.-X. Ecologists overestimate the importance of predictor variables in model averaging: A plea for cautious interpretations. Methods Ecol. Evol. 2014, 5, 983–991. [Google Scholar] [CrossRef] [Green Version]
- Galipaud, M.; Gillingham, M.A.F.; Dechaume-Moncharmont, F.-X. A farewell to the sum of Akaike weights: The benefits of alternative metrics for variable importance estimations in model selection. Methods Ecol. Evol. 2017, 8, 1668–1678. [Google Scholar] [CrossRef] [Green Version]
- Lukacs, P.M.; Burnham, K.P.; Anderson, D.R. Model selection bias and Freedman’s paradox. Ann. Inst. Stat. Math. 2010, 62, 117–125. [Google Scholar] [CrossRef]
- Bauwens, D.; Garland, T.; Castilla, A.M.; Vandamme, R. Evolution of sprint speed in lacertid lizards: Morphological, physiological, and behavioral covariation. Evolution 1995, 49, 848–863. [Google Scholar]
- Sinclair, B.J.; Marshall, K.E.; Sewell, M.A.; Levesque, D.L.; Willett, C.S.; Slotsbo, S.; Dong, Y.W.; Harley, C.D.G.; Marshall, D.J.; Helmuth, B.S.; et al. Can we predict ectotherm responses to climate change using thermal performance curves and body temperatures? Ecol. Lett. 2016, 19, 1372–1385. [Google Scholar] [CrossRef] [Green Version]
- Angilletta, M.J., Jr.; Bennett, A.F.; Guderley, H.; Navas, C.A.; Seebacher, F.; Wilson, R.S. Coadaptation: A unifying principle in evolutionary thermal biology. Physiol. Biochem. Zool. 2006, 79, 282–294. [Google Scholar] [CrossRef] [Green Version]
- Higham, T.E.; Russell, A.P.; Zani, P.A. Integrative biology of tail autotomy in lizards. Physiol. Biochem. Zool. 2013, 86, 603–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huey, R.B.; Stevenson, R.D. Integrating thermal physiology and ecology of ectotherms: A discussion of approaches. Am. Zool. 1979, 19, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Huey, R.B. Temperature, physiology, and the ecology of reptiles. In Biology of the Reptilia; Gans, C., Pough, F.H., Eds.; Academic Press: London, UK, 1982; Volume 12, pp. 25–91. [Google Scholar]
- Angilletta, M.J., Jr.; Niewiarowski, P.H.; Navas, C.A. The evolution of thermal physiology in ectotherms. J. Therm. Biol. 2002, 27, 249–268. [Google Scholar] [CrossRef]
- Strangas, M.L.; Navas, C.A.; Rodrigues, M.T.; Carnaval, A.C. Thermophysiology, microclimates, and species distributions of lizards in the mountains of the Brazilian Atlantic Forest. Ecography 2019, 42, 354–364. [Google Scholar] [CrossRef]
- Colli, G.R.; Hoogmoed, M.S.; Cannatella, D.C.; Cassimiro, J.; Gomes, J.O.; Ghellere, J.M.; Nunes, P.M.S.; Pellegrino, K.C.M.; Salerno, P.; De Souza, S.M.; et al. Description and phylogenetic relationships of a new genus and two new species of lizards from Brazilian Amazonia, with nomenclatural comments on the taxonomy of Gymnophthalmidae (Reptilia: Squamata). Zootaxa 2015, 4000, 401–427. [Google Scholar] [CrossRef] [Green Version]
- Clusella-Trullas, S.; Chown, S.L. Lizard thermal trait variation at multiple scales: A review. J. Comp. Physiol. B-Biochem. Syst. Environ. Physiol. 2014, 184, 5–21. [Google Scholar] [CrossRef]
- Huey, R.B.; Kingsolver, J.G. Evolution of thermal sensitivity of ectotherm performance. Trends Ecol. Evol. 1989, 4, 131–135. [Google Scholar] [CrossRef]
- Van Damme, R.; Vanhooydonck, B. Origins of interspecific variation in lizard sprint capacity. Funct. Ecol. 2001, 15, 186–202. [Google Scholar] [CrossRef]
- Pontes-da-Silva, E.; Magnusson, W.E.; Sinervo, B.; Caetano, G.H.; Miles, D.B.; Colli, G.R.; Diele-Viegas, L.M.; Fenker, J.; Santos, J.C.; Werneck, F.P. Extinction risks forced by climatic change and intraspecific variation in the thermal physiology of a tropical lizard. J. Therm. Biol. 2018, 73, 50–60. [Google Scholar] [CrossRef]
- Mendez-Galeano, M.A.; Paternina-Cruz, R.F.; Calderon-Espinosa, M.L. The highest kingdom of Anolis: Thermal biology of the Andean lizard Anolis heterodermus (Squamata: Dactyloidae) over an elevational gradient in the Eastern Cordillera of Colombia. J. Therm. Biol. 2020, 89, 102498. [Google Scholar] [CrossRef]
- Marques, E.Q.; Marimon-Junior, B.H.; Marimon, B.S.; Matricardi, E.A.T.; Mews, H.A.; Colli, G.R. Redefining the Cerrado-Amazonia transition: Implications for conservation. Biodivers. Conserv. 2020, 29, 1501–1517. [Google Scholar] [CrossRef]
- Gaston, K.J. Geographic range limits: Achieving synthesis. Proc. R. Soc. B-Biol. Sci. 2009, 276, 1395–1406. [Google Scholar] [CrossRef] [Green Version]
- Garner, T.W.J.; Pearman, P.B.; Angelone, S. Genetic diversity across a vertebrate species’ range: A test of the central-peripheral hypothesis. Mol. Ecol. 2004, 13, 1047–1053. [Google Scholar] [CrossRef]
- Pironon, S.; Villellas, J.; Morris, W.F.; Doak, D.F.; Garcia, M.B. Do geographic, climatic or historical ranges differentiate the performance of central versus peripheral populations? Glob. Ecol. Biogeogr. 2015, 24, 611–620. [Google Scholar] [CrossRef]
- Valladares, F.; Matesanz, S.; Guilhaumon, F.; Araujo, M.B.; Balaguer, L.; Benito-Garzon, M.; Cornwell, W.; Gianoli, E.; van Kleunen, M.; Naya, D.E.; et al. The effects of phenotypic plasticity and local adaptation on forecasts of species range shifts under climate change. Ecol. Lett. 2014, 17, 1351–1364. [Google Scholar] [CrossRef] [Green Version]
- Shine, R. Effects of pregnancy on locomotor performance: An experimental study on lizards. Oecologia 2003, 136, 450–456. [Google Scholar] [CrossRef]
- Olsson, M.; Shine, R.; Bak-Olsson, E. Locomotor impairment of gravid lizards: Is the burden physical or physiological? J. Evol. Biol. 2000, 13, 263–268. [Google Scholar] [CrossRef]
- Bauwens, D.; Thoen, C. Escape tactics and vulnerability to predation associated with reproduction in the lizard Lacerta vivipara. J. Anim. Ecol. 1981, 50, 733–743. [Google Scholar] [CrossRef]
- Itonaga, K.; Jones, S.M.; Wapstra, E. Do gravid females become selfish? Female allocation of energy during gestation. Physiol. Biochem. Zool. 2012, 85, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Sinervo, B.; Hedges, R.; Adolph, S.C. Decreased sprint speed as a cost of reproduction in the lizard Sceloporus Occidentalis: Variation among populations. J. Exp. Biol. 1991, 155, 323–336. [Google Scholar] [CrossRef]
- Seigel, R.A.; Huggins, M.M.; Ford, N.B. Reduction in locomotor ability as a cost of reproduction in gravid snakes. Oecologia 1987, 73, 481–485. [Google Scholar] [CrossRef]
- Shine, R. “Costs” of reproduction in reptiles. Oecologia 1980, 46, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, R.; Bauwens, D.; Verheyen, R.F. Effect of relative clutch mass on sprint speed in the lizard Lacerta vivipara. J. Herpetol. 1989, 23, 459–461. [Google Scholar] [CrossRef]
- Cooper, W.E., Jr.; Wilson, D.S.; Smith, G.R. Sex, reproductive status, and cost of tail autotomy via decreased running speed in lizards. Ethology 2009, 115, 7–13. [Google Scholar] [CrossRef]
- Shine, R. Locomotor speeds of gravid lizards: Placing ‘costs of reproduction’ within an ecological context. Funct. Ecol. 2003, 17, 526–533. [Google Scholar] [CrossRef]
- Clemente, C.J.; Withers, P.C.; Thompson, G. Optimal body size with respect to maximal speed for the yellow-spotted monitor lizard (Varanus panoptes; Varanidae). Physiol. Biochem. Zool. 2012, 85, 265–273. [Google Scholar] [CrossRef]
- Jones, J.H.; Lindstedt, S.L. Limits to maximal performance. Annu. Rev. Physiol. 1993, 55, 547–569. [Google Scholar] [CrossRef]
- Heglund, N.C.; Taylor, C.R.; McMahon, T.A. Scaling stride frequency and gait to animal size: Mice to horses. Science 1974, 186, 1112–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt-Nielsen, K. Locomotion: Energy cost of swimming, flying, and running. Science 1972, 177, 222. [Google Scholar] [CrossRef]
- Zamora-Camacho, F.J.; Reguera, S.; Moreno-Rueda, G. Does tail autotomy affect thermoregulation in an accurately thermoregulating lizard? Lessons from a 2200-m elevational gradient. J. Zool. 2015, 297, 204–210. [Google Scholar] [CrossRef]
- Downes, S.J.; Shine, R. Why does tail loss increase a lizard’s later vulnerability to snake predators? Ecology 2001, 82, 1293–1303. [Google Scholar] [CrossRef]
- Jagnandan, K.; Russell, A.P.; Higham, T.E. Tail autotomy and subsequent regeneration alter the mechanics of locomotion in lizards. J. Exp. Biol. 2014, 217, 3891–3897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamley, T. Functions of the tail in bipedal locomotion of lizards dinosaurs and pterosaurs. Mem. Qld. Mus. 1990, 28, 153–158. [Google Scholar]
- Höfling, E.; Renous, S.; Curcio, F.F.; Eterovic, A.; de Souza Santos Filho, P. Effects of surface roughness on the locomotion of a long-tailed lizard, Colobodactylus taunayi Amaral, 1933 (Gymnophthalmidae: Heterodactylini). Int. J. Zool. 2012, 2012, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Vitt, L.J.; Caldwell, J.P. Herpetology. An Introductory Biology of Amphibians and Reptiles, 4th ed.; Elsevier, Inc.: London, UK, 2014. [Google Scholar]
- Gans, C. Tetrapod limblessness: Evolution and functional corollaries. Am. Zool. 1975, 15, 455–467. [Google Scholar] [CrossRef]
- da Silva, F.O.; Fabre, A.C.; Savriama, Y.; Ollonen, J.; Mahlow, K.; Herrel, A.; Muller, J.; Di-Poi, N. The ecological origins of snakes as revealed by skull evolution. Nat. Commun. 2018, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- McElroy, E.J. The effect of tail autotomy on locomotor performance in the long tailed grass lizard, Takydromus sexlineatus. Integr. Comp. Biol. 2011, 51, E89. [Google Scholar]
- Wiens, J.J.; Slingluff, J.L. How lizards turn into snakes: A phylogenetic analysis of body-form evolution in anguid lizards. Evolution 2001, 55, 2303–2318. [Google Scholar] [CrossRef]
- Wiens, J.J.; Brandley, M.C.; Reeder, T.W. Why does a trait evolve multiple times within a clade? Repeated evolution of snakelike body form in squamate reptiles. Evolution 2006, 60, 123–141. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameter | Brasília | Nova Xavantina | Total |
|---|---|---|---|
| Sample size | 39 | 64 | 103 |
| Mass (g) | 1.49 ± 0.35 | 0.82 ± 0.19 | 1.06 ± 0.41 |
| Snout-vent length (mm) | 38.22 ± 3.06 | 34.98 ± 2.76 | 36.12 ± 3.26 |
| Tail length (mm) | 48.62 ± 13.74 | 46.37 ± 17.64 | 47.17 ± 16.39 |
| Relative tail length | 1.27 ± 0.34 | 1.32 ± 0.50 | 1.31 ± 0.45 |
| Length of regenerated portion of tail (mm) | 6.38 ± 10.30 | 9.99 ± 11.95 | 8.71 ± 11.51 |
| Relative length of regenerated portion of tail | 0.16 ± 0.26 | 0.28 ± 0.34 | 0.24 ± 0.32 |
| Body temperature during runs (°C) | 27.94 ± 6.72 | 27.53 ± 5.43 | 27.67 ± 5.88 |
| Critical thermal minimum (°C) | 13.90 ± 1.78 | 15.20 ± 3.29 | 14.70 ± 2.08 |
| Critical thermal maximum (°C) | 44.41 ± 1.84 | 40.00 ± 3.09 | 41.69 ± 3.43 |
| Sprint speed (maximum) | 0.07 ± 0.02 | 0.10 ± 0.04 | 0.09 ± 0.04 |
| Parametric Terms | ||||
| Term | Estimate | Std. Error | t | p |
| (Intercept) | 0.0411 | 0.0065 | 6.3080 | <0.0001 |
| LocalNX | 0.0220 | 0.0069 | 3.1920 | 0.0015 |
| SexMale | 0.0015 | 0.0045 | 0.3230 | 0.7467 |
| AUYes | 0.0000 | 0.0070 | −0.0040 | 0.9970 |
| Pregnancy | −0.0203 | 0.0068 | −2.9700 | 0.0031 |
| Smooth Terms | ||||
| Term | edf | Ref. df | F | p |
| s(Temperature) | 7.976 | 7.976 | 95.569 | <0.00001 |
| s(CCr) | 1.000 | 1.000 | 0.036 | 0.84979 |
| s(RCr) | 1.000 | 1.000 | 3.037 | 0.08203 |
| s(SVL) | 1.000 | 1.000 | 0.365 | 0.54589 |
| s(Mass) | 3.119 | 3.119 | 4.983 | 0.00259 |
| Model Selection | |||||||||
| Model | df | logLik | AICc | ΔAICc | wAICc | ||||
| Pregnancy + Site + s(Mass) + s(RCr) + s(Temperature) | 11 | 934.81 | −1847.06 | 0.00 | 0.25 | ||||
| AU + Pregnancy + Site + s(Mass) + s(RCr) + s(Temperature) | 12 | 934.84 | −1845.01 | 2.05 | 0.09 | ||||
| Pregnancy + Site + s(Mass) + s(RCr) + s(Temperature)+ Sex | 12 | 934.82 | −1844.97 | 2.10 | 0.09 | ||||
| Pregnancy + Site + s(Mass) + s(Temperature) | 9 | 931.35 | −1844.32 | 2.75 | 0.06 | ||||
| AU + Pregnancy + Site + s(Mass) + s(Temperature) | 10 | 932.22 | −1843.96 | 3.10 | 0.05 | ||||
| Pregnancy + Site + s(Mass) + s(RCr) + s(SVL) + s(Temperature) | 13 | 934.93 | −1843.07 | 3.99 | 0.03 | ||||
| Model Averaging | |||||||||
| Importance | s(Temperature) | Pregnancy | s(Mass) | Site | s(RCr) | AU | Sex | s(CCr) | s(SVL) |
| Sum of model weights | 1.00 | 0.93 | 0.90 | 0.89 | 0.73 | 0.33 | 0.27 | 0.15 | 0.14 |
| Number of containing models | 255 | 253 | 253 | 254 | 254 | 255 | 255 | 255 | 254 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, N.A.; Caetano, G.H.d.O.; Campelo, P.H.; Cavalcante, V.H.G.L.; Godinho, L.B.; Miles, D.B.; Paulino, H.M.; da Silva, J.M.A.; de Souza, B.A.; da Silva, H.B.F.; et al. Effects of Caudal Autotomy on the Locomotor Performance of Micrablepharus Atticolus (Squamata, Gymnophthalmidae). Diversity 2021, 13, 562. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110562
Silva NA, Caetano GHdO, Campelo PH, Cavalcante VHGL, Godinho LB, Miles DB, Paulino HM, da Silva JMA, de Souza BA, da Silva HBF, et al. Effects of Caudal Autotomy on the Locomotor Performance of Micrablepharus Atticolus (Squamata, Gymnophthalmidae). Diversity. 2021; 13(11):562. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110562
Chicago/Turabian StyleSilva, Naiane Arantes, Gabriel Henrique de Oliveira Caetano, Pedro Henrique Campelo, Vitor Hugo Gomes Lacerda Cavalcante, Leandro Braga Godinho, Donald Bailey Miles, Henrique Monteiro Paulino, Júlio Miguel Alvarenga da Silva, Bruno Araújo de Souza, Hosmano Batista Ferreira da Silva, and et al. 2021. "Effects of Caudal Autotomy on the Locomotor Performance of Micrablepharus Atticolus (Squamata, Gymnophthalmidae)" Diversity 13, no. 11: 562. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110562