
Local Actions to Tackle a Global Problem: A Multidimensional Assessment of the Pollination Crisis in Chile

 , , , ,
, , , , 

Abstract
:1. Introduction
2. Pollinators in Agricultural Landscapes
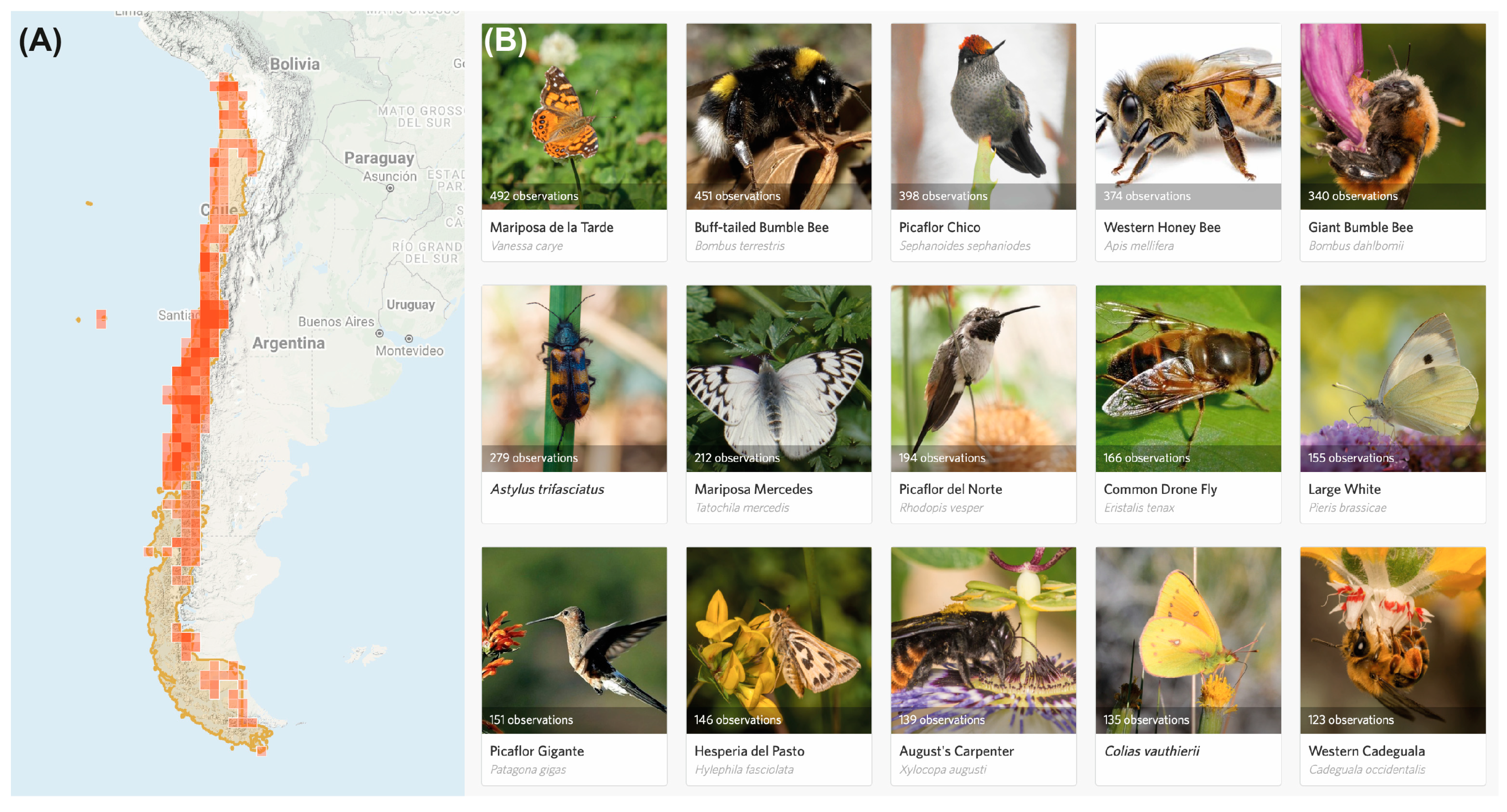
2.1. Native Pollinators
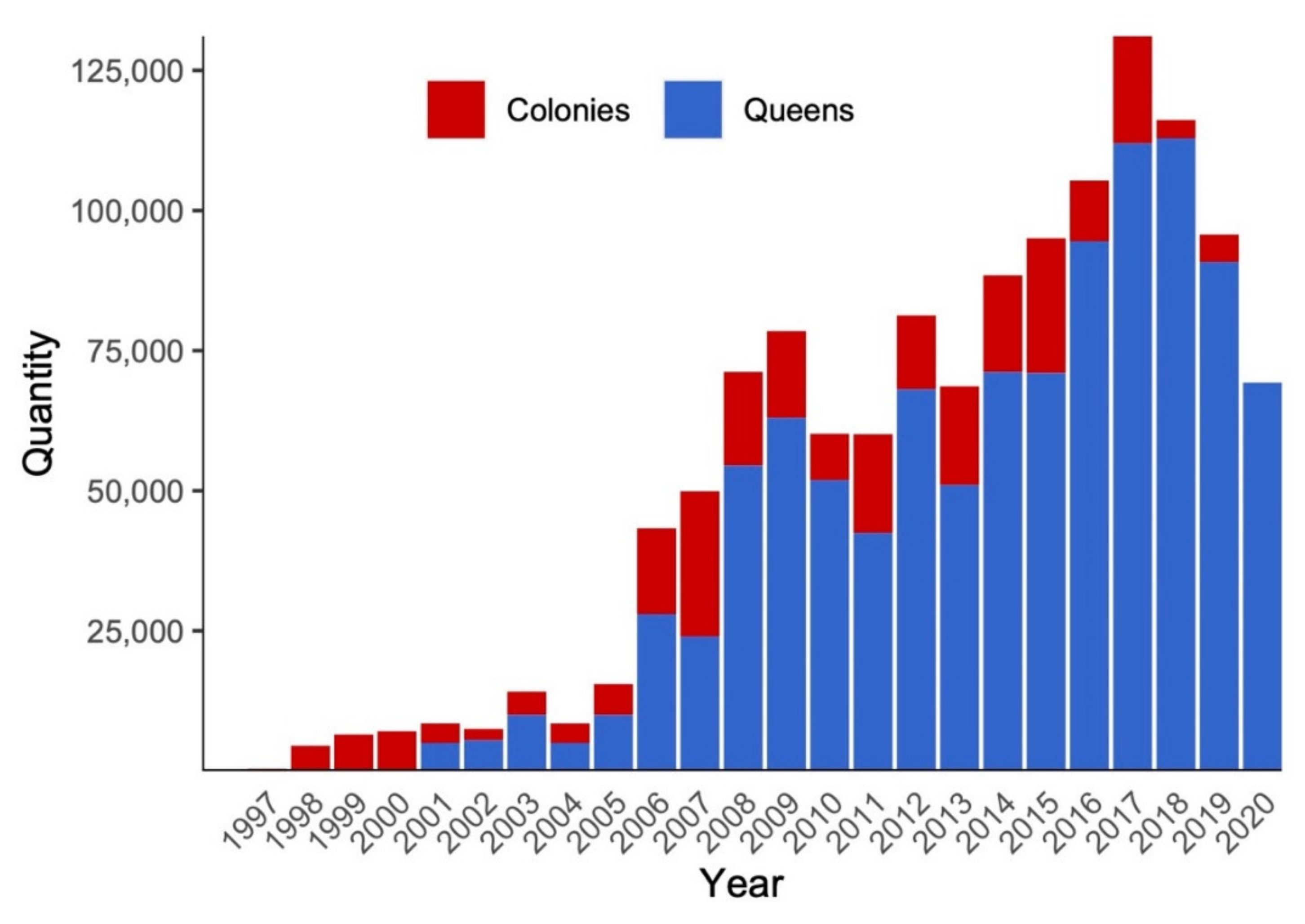
2.2. Managed Pollinators
2.3. Crop Dependence on Pollination
3. Drivers of Pollination Decline
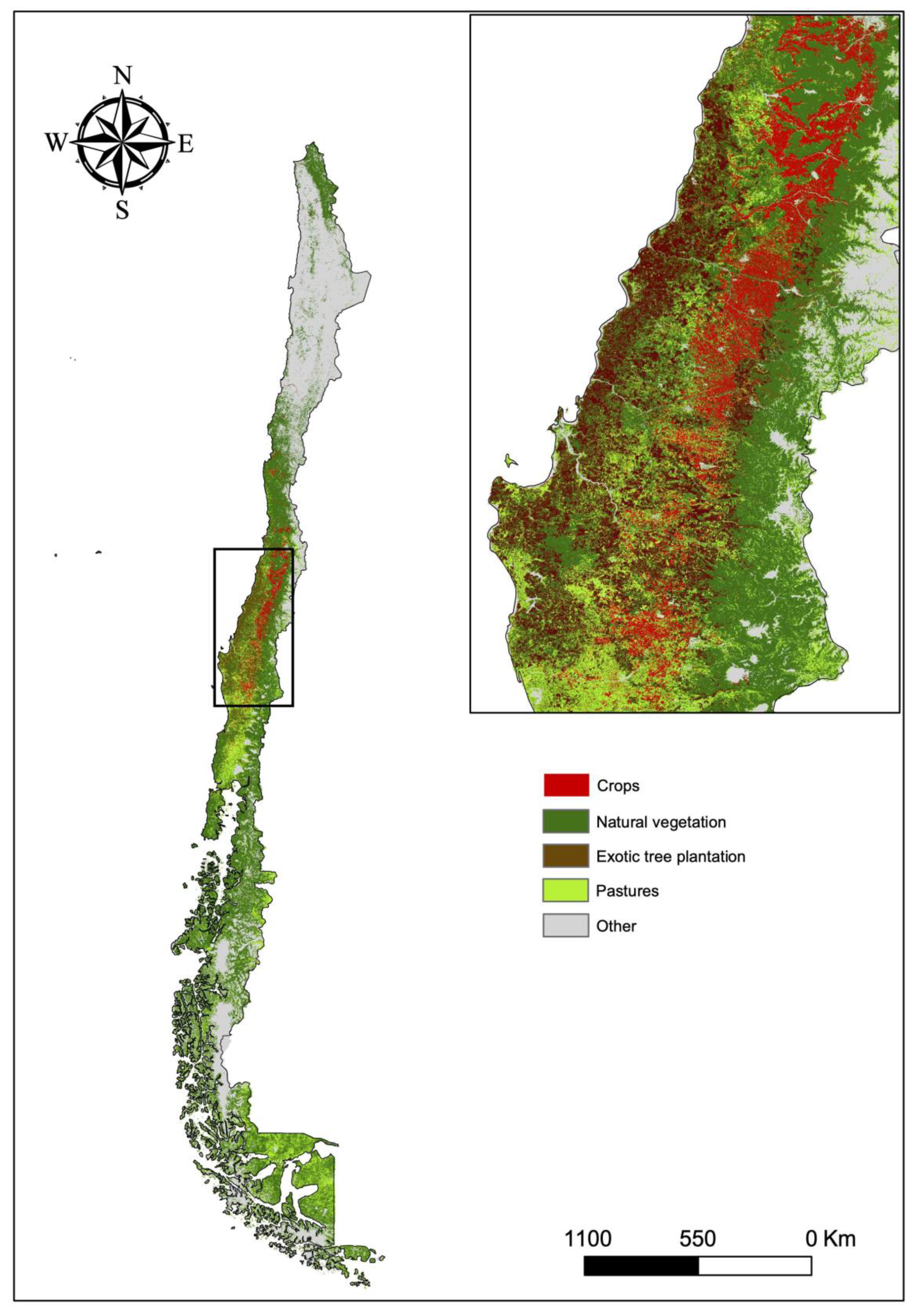
3.1. Habitat Loss
3.2. Pesticides
3.3. Invasive Species
3.4. Climate Change
4. Gaps and Future Challenges
4.1. Knowledge Gaps on Pollinators and Pollination in Chile
4.2. Future Challenges
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Biodiversity—Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- IPBES. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES secretariat: Bonn, Germany, 2019. [Google Scholar]
- Valiente-Banuet, A.; Aizen, M.A.; Alcantara, J.M.; Arroyo, J.; Cocucci, A.; Galetti, M.; Garcia, M.B.; Garcia, D.; Gomez, J.M.; Jordano, P.; et al. Beyond species loss: The extinction of ecological interactions in a changing world. Funct. Ecol. 2015, 29, 299–307. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. P Roy. Soc. B-Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Perfecto, I.; Vandermeer, J.; Wright, A. Nature’s matrix. Linking agriculture, biodiversity conservation and food sovereignty; Routledge: London, UK, 2019. [Google Scholar]
- Altieri, M.A.; Nicholls, C. Agroecology: Challenges and opportunities for farming in the Anthropocene. Int. J. Agric. Nat. Resour. 2020, 47, 204–215. [Google Scholar]
- Altieri, M.A. Agroecology: The Science of Sustainable Agriculture, 2nd ed.; Westview Press: Boulder, CO, USA, 2015. [Google Scholar]
- Gliessman, S.R. Agroecology: The Ecology of Sustainable Food Systems, 3rd ed.; CRC Press Taylor & Francis: Boca Raton, FL, USA, 2015. [Google Scholar]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef]
- Schutz, J. Creating an integrated protected area network in Chile: A GIS assessment of ecoregion representation and the role of private protected areas. Environ. Conserv. 2018, 45, 269–277. [Google Scholar] [CrossRef]
- Pérez, R.; Valdés, A. Análisis de la Evolución y Desempeño Agroexportador de Chile, 2000–2017; Oficina de estudios y Políticas Agrarias—ODEPA, 2019. Available online: https://www.odepa.gob.cl/wp-content/uploads/2019/04/EvolucionAgroExp.pdf (accessed on 28 April 2021).
- Pefaur, J. Evolución de la Fruticultura Chilena en los Últimos 20 años; Oficina de estudios y Políticas Agrarias—ODEPA, 2020. Available online: https://bibliotecadigital.odepa.gob.cl/bitstream/handle/20.500.12650/70234/evolucionFruticulturachilena.pdf (accessed on 28 April 2021).
- Schulz, J.J.; Cayuela, L.; Echeverria, C.; Salas, J.; Benayas, J.M.R. Monitoring land cover change of the dryland forest landscape of Central Chile (1975–2008). Appl. Geogr. 2010, 30, 436–447. [Google Scholar] [CrossRef] [Green Version]
- FAO. FAOSTAT Statistics Database (updated March 2021); Food and Agriculture Organization of the United Nations: Rome, Italy, 2021. [Google Scholar]
- Isbell, F.; Adler, P.R.; Eisenhauer, N.; Fornara, D.; Kimmel, K.; Kremen, C.; Letourneau, D.K.; Liebman, M.; Polley, H.W.; Quijas, S. Benefits of increasing plant diversity in sustainable agroecosystems. J. Ecol. 2017, 105, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Firbank, L.G.; Petit, S.; Smart, S.; Blain, A.; Fuller, R.J. Assessing the impacts of agricultural intensification on biodiversity: A British perspective. Philos. T. R. Soc. B 2008, 363, 777–787. [Google Scholar] [CrossRef]
- Lonsdorf, E.; Kremen, C.; Ricketts, T.; Winfree, R.; Williams, N.; Greenleaf, S. Modelling pollination services across agricultural landscapes. Ann. Bot. 2009, 103, 1589–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef] [PubMed]
- López-Aliste, M.; Flores-Prado, L.; Ruz, L.; Sepúlveda, Y.; Rodríguez, S.; Saraiva, A.M.; Fontúrbel, F.E. Wild bees of Chile: A database on taxonomy, sociality, and ecology. Ecology 2021, 102, e03377. [Google Scholar] [CrossRef] [PubMed]
- Smith-Ramirez, C.; Vieli, L.; Barahona-Segovia, R.M.; Montalva, J.; Cianferoni, F.; Ruz, L.; Fontúrbel, F.E.; Valdivia, C.E.; Medel, R.; Pauchard, A.; et al. The reasons why Chile should stop importing commercial bumblebee Bombus terrestris (Linnaeus) and to start controlling it. Gayana 2018, 82, 118–127. [Google Scholar] [CrossRef]
- Mogren, C.L.; Rand, T.A.; Fausti, S.W.; Lundgren, J.G. The effects of crop intensification on the diversity of native pollinator communities. Environ. Entomol. 2016, 45, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Williams, N.M.; Aizen, M.A.; Gemmill-Herren, B.; LeBuhn, G.; Minckley, R.; Packer, L.; Potts, S.G.; Roulston, T.a.; Steffan-Dewenter, I. Pollination and other ecosystem services produced by mobile organisms: A conceptual framework for the effects of land-use change. Ecol. Lett. 2007, 10, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Nabhan, G.P.; Buchmann, S.L. Services provided by pollinators. In Nature’s Services: Societal Dependence on Natural Ecosystems; Daily, G.C., Ed.; Island Press: Washington, DC, USA, 1997; pp. 133–150. [Google Scholar]
- Eeraerts, M.; Smagghe, G.; Meeus, I. Pollinator diversity, floral resources and semi-natural habitat, instead of honey bees and intensive agriculture, enhance pollination service to sweet cherry. Agr. Ecosyst. Environ. 2019, 284, 106586. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Garratt, M.P.D.; Truslove, C.; Coston, D.; Evans, R.; Moss, E.; Dodson, C.; Jenner, N.; Biesmeijer, J.; Potts, S. Pollination deficits in UK apple orchards. J. Pollinat. Ecol. 2014, 12, 9–14. [Google Scholar] [CrossRef]
- Miñarro, M.; Somoano, A.; Ventura, J. Intra-annual continuous reproduction of the apple pest Microtus lusitanicus: Implications for management. Crop. Prot. 2017, 96, 164–172. [Google Scholar] [CrossRef]
- Estay, P.; Wagner, A.; Escaff, M. Evaluation of Bombus dahlbomii (Guér.) as a pollinating agent for tomato flowers under greenhouse conditions. Agricultura Técnica 2001, 61, 113–119. [Google Scholar] [CrossRef]
- Monzón, V.H.; Avendano-Soto, P.; Araujo, R.O.; Garrido, R.; Mesquita-Neto, J.N. Avocado crops as a floral resource for native bees of Chile. Rev. Chil. Hist. Nat. 2020, 93, art5. [Google Scholar] [CrossRef]
- Muñoz, A.E.; Plantegenest, M.; Amouroux, P.; Zaviezo, T. Native flower strips increase visitation by non-bee insects to avocado flowers and promote yield. Basic Appl. Ecol. 2021, 56, 369–378. [Google Scholar] [CrossRef]
- Smith-Ramirez, C.; Rendón-Funes, A.; Barahona-Segovia, R.; Moya, W. Consequences of the high abundance of Bombus terrestris on the pollination of Vicia faba. J. Pollinat. Ecol. 2021, 29, 258–272. [Google Scholar] [CrossRef]
- Lander, T.A.; Harris, S.A.; Boshier, D.H. Flower and fruit production and insect pollination of the endangered Chilean tree, Gomortega keule in native forest, exotic pine plantation and agricultural environments. Rev. Chil. Hist. Nat. 2009, 82, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Murúa, M.; Cisterna, J.; Rosende, B. Pollination ecology and breeding system of two Calceolaria species in Chile. Rev. Chil. Hist. Nat. 2014, 87, art7. [Google Scholar] [CrossRef] [Green Version]
- Rego, J.O.; Schlindwein, C.; Garrido, R.; Monzón, V.H. Low fruit set in an endangered tree: Pollination by exotic bumblebees and pollen resource for relictual native bees. Arthropod-Plant. Interact. 2021, 15, 491–501. [Google Scholar] [CrossRef]
- Murúa, M.; Espindola, A.; González, A.; Medel, R. Pollinators and crossability as reproductive isolation barriers in two sympatric oil-rewarding Calceolaria (Calceolariaceae) species. Evol. Ecol. 2017, 31, 421–434. [Google Scholar] [CrossRef]
- Montalva, J.; Ruz, L. Actualización de la lista sistemática de las abejas chilenas (Hymenoptera: Apoidea). Revista Chilena Entomología 2017, 35, 15–52. [Google Scholar]
- Barahona-Segovia, R.M.; Riera, P.; Paninao-Monsalvez, L.; Guzmán, V.V.; Henriquez-Piskulich, P. Updating the knowledge of the flower flies (Diptera: Syrphidae) from Chile: Illustrated catalog, extinction risk and biological notes. Zootaxa 2021, 4959, 1–178. [Google Scholar] [CrossRef]
- Medel, R.; González-Browne, C.; Fontúrbel, F.E. Pollination in the Chilean Mediterranean-type ecosystem: A review of current advances and pending tasks. Plant Biol. 2018, 20, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith-Ramirez, C.; Martinez, P.; Nuñez, M.; González, C.; Armesto, J.J. Diversity, flower visitation frequency and generalism of pollinators in temperate rain forests of Chiloe Island, Chile. Bot. J. Linn. Soc. 2005, 147, 399–416. [Google Scholar] [CrossRef] [Green Version]
- Aizen, M.A.; Morales, C.L.; Morales, J.M. Invasive mutualists erode native pollination webs. PLoS Biol. 2008, 6, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, N.; Sánchez, P.; Pauchard, A.; Urrutia, J.; Cavieres, L.; Marticorena, A. Plantas Invasoras del Centro-sur de Chile; Instituto de Ecología y Biodiversidad (IEB): Santiago de Chile, Chile, 2014. [Google Scholar]
- Morales, C.L.; Arbetman, M.P.; Cameron, S.A.; Aizen, M.A. Rapid ecological replacement of a native bumble bee by invasive species. Front. Ecol. Environ. 2013, 11, 529–534. [Google Scholar] [CrossRef]
- Schmid-Hempel, R.; Eckhardt, M.; Goulson, D.; Heinzmann, D.; Lange, C.; Plischuk, S.; Escudero, L.R.; Salathé, R.; Scriven, J.J.; Schmid-Hempel, P. The invasion of southern South America by imported bumblebees and associated parasites. J. Anim. Ecol. 2014, 83, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Aebi, A.; Neumann, P. Endosymbionts and honey bee colony losses? Trends Ecol. Evol. 2011, 26, 494. [Google Scholar] [CrossRef] [Green Version]
- Committee on the Status of Pollinators in North America, N.R.C. Status of Pollinators in North. Ame; The National Academies Press: Washington, DC, USA, 2007. [Google Scholar]
- Williams, I.H. Insect Pollination and Crop Production: A European Perspective. In Pollinating Bees—The Conservation Link Between Agriculture and Nature; Kevan, P., Imperatriz-Fonseca, V.L., Eds.; Ministry of Environment: Brasília, Brazil, 2002; pp. 59–65. [Google Scholar]
- Rader, R.; Howlett, B.G.; Cunningham, S.A.; Westcott, D.A.; Newstrom-Lloyd, L.E.; Walker, M.K.; Teulon, D.A.; Edwards, W. Alternative pollinator taxa are equally efficient but not as effective as the honeybee in a mass flowering crop. J. Appl. Ecol. 2009, 46, 1080–1087. [Google Scholar] [CrossRef]
- Aizen, M.A.; Harder, L.D. The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 2009, 19, 915–918. [Google Scholar] [CrossRef] [Green Version]
- Requier, F.; Antúnez, K.; Morales, C.L.; Aldea Sánchez, P.; Castilhos, D.; Garrido, P.M.; Giacobino, A.; Reynaldi, F.J.; Rosso Londoño, J.M.; Santos, E. Trends in beekeeping and honey bee colony losses in Latin America. J. Apic Res. 2018, 57, 657–662. [Google Scholar] [CrossRef]
- SAG. Response Letter Nº 3462 (May 26th 2021) by Request Under the Law 20.285 of Access to Public Information; Chilean Ministry of Agriculture: Santiago de Chile, Brazil, 2021.
- Fontúrbel, F.E.; Murúa, M.M.; Vieli, L. Invasion dynamics of the European bumblebee Bombus terrestris in the southern part of South America. Sci. Rep. 2021, 11, 1–7. [Google Scholar]
- Zhao, Y.Y.; Feng, D.L.; Yu, L.; Wang, X.Y.; Chen, Y.L.; Bai, Y.Q.; Hernandez, H.J.; Galleguillos, M.; Estades, C.; Biging, G.S.; et al. Detailed dynamic land cover mapping of Chile: Accuracy improvement by integrating multi-temporal data. Remote Sens. Environ. 2016, 183, 170–185. [Google Scholar] [CrossRef]
- ODEPA-CIREN. Estadísticas Productivas. 2021. Available online: https://www.odepa.gob.cl/estadisticas-del-sector/estadisticas-productivas (accessed on 26 October 2021).
- Eguillor, P.M. Agricultura Orgánica, ODS y Cambio Climático: Mercado Internacional y Nacional. 2020. Available online: https://bibliotecadigital.odepa.gob.cl/bitstream/handle/20.500.12650/70558/Articulo%20AgriculturaOrganica.pdf (accessed on 26 October 2021).
- ODEPA-INE. VII Nacional Census of Agriculture and Forestry. 2007. Available online: https://www.odepa.gob.cl/estadisticas-del-sector/ficha-nacional-y-regionales (accessed on 26 October 2021).
- Stokstad, E. The case of the empty hives. Science 2007, 316, 970–972. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- VanEngelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y. Colony collapse disorder: A descriptive study. PLoS ONE 2009, 4, e6481. [Google Scholar] [CrossRef]
- Flores, J.M.; Gámiz, V.; Jiménez-Marín, Á.; Flores-Cortés, A.; Gil-Lebrero, S.; Garrido, J.J.; Hernando, M.D. Impact of Varroa destructor and associated pathologies on the colony collapse disorder affecting honey bees. Res. Vet. Sci. 2021, 135, 85–95. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemuller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Murray, T.E.; Kuhlmann, M.; Potts, S.G. Conservation ecology of bees: Populations, species and communities. Apidologie 2009, 40, 211–236. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.J.; Paxton, R.J. The conservation of bees: A global perspective. Apidologie 2009, 40, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Phillips, B.B.; Shaw, R.F.; Holland, M.J.; Fry, E.L.; Bardgett, R.D.; Bullock, J.M.; Osborne, J.L. Drought reduces floral resources for pollinators. Global Change Biol. 2018, 24, 3226–3235. [Google Scholar] [CrossRef]
- Aizen, M.A.; Smith-Ramirez, C.; Morales, C.L.; Vieli, L.; Saez, A.; Barahona-Segovia, R.M.; Arbetman, M.P.; Montalva, J.; Garibaldi, L.A.; Inouye, D.W.; et al. Coordinated species importation policies are needed to reduce serious invasions globally: The case of alien bumblebees in South America. J. Appl. Ecol. 2019, 56, 100–106. [Google Scholar] [CrossRef] [Green Version]
- De Palma, A.; Abrahamczyk, S.; Aizen, M.A.; Albrecht, M.; Basset, Y.; Bates, A.; Blake, R.J.; Boutin, C.; Bugter, R.; Connop, S. Predicting bee community responses to land-use changes: Effects of geographic and taxonomic biases. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, A.; Altamirano, A.; Cayuela, L.; Lara, A.; González, M. Native forest loss in the Chilean biodiversity hotspot: Revealing the evidence. Reg. Environ. Change 2017, 17, 285–297. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Fischer, J. Habitat Fragmentation and Landscape Change: An Ecological and Conservation Synthesis; Island Press: Washington, DC, USA, 2013. [Google Scholar]
- Ewers, R.M.; Didham, R.K. Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. 2006, 81, 117–142. [Google Scholar] [CrossRef] [PubMed]
- Miñarro, M.; García, D.; Martínez, R. Impact of insect pollinators in agriculture: Importance and management of their biodiversity. Ecosistemas 2018, 27, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Karp, D.S.; Mendenhall, C.D.; Sandi, R.F.; Chaumont, N.; Ehrlich, P.R.; Hadly, E.A.; Daily, G.C. Forest bolsters bird abundance, pest control and coffee yield. Ecol. Lett. 2013, 16, 1339–1347. [Google Scholar] [CrossRef]
- Rodriguez-San Pedro, A.; Chaperon, P.N.; Beitran, C.A.; Allendes, J.L.; Avila, F.I.; Grez, A.A. Influence of agricultural management on bat activity and species richness in vineyards of central Chile. J. Mammal. 2018, 99, 1495–1502. [Google Scholar] [CrossRef]
- García, D.; Miñarro, M.; Martínez-Sastre, R. Enhancing ecosystem services in apple orchards: Nest boxes increase pest control by insectivorous birds. J. Appl. Ecol. 2021, 58, 465–475. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Gemmill-Herren, B.; D’Annolfo, R.; Graeub, B.E.; Cunningham, S.A.; Breeze, T.D. Farming approaches for greater biodiversity, livelihoods, and food security. Trends Ecol. Evol. 2017, 32, 68–80. [Google Scholar] [CrossRef]
- Manosathiyadevan, M.; Bhuvaneshwari, V.; Latha, R. Impact of insects and pests in loss of crop production: A review. In Sustainable Agriculture towards Food Security; Dhanarajan, A., Ed.; Springer: Singapore, 2017; pp. 57–67. [Google Scholar]
- Casallanovo, F.; Simone, D.M.; Santos, G.S.; de Oliveira Kaminski, T.S.; Cione, A.P.; Peranginangin, N. Estimating pesticide environmental concentrations in Latin America: The importance of developing local scenarios. Integr. Environ. Assess. Manage. 2021, 17, 901–904. [Google Scholar] [CrossRef]
- Blacquiere, T.; Smagghe, G.; Van Gestel, C.A.M.; Mommaerts, V. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment (vol 21, pg 973, 2012). Ecotoxicology 2012, 21, 1581. [Google Scholar] [CrossRef] [Green Version]
- Woodcock, B.A.; Bullock, J.M.; Shore, R.F.; Heard, M.S.; Pereira, M.G.; Redhead, J.; Ridding, L.; Dean, H.; Sleep, D.; Henrys, P.; et al. Country-specific effects of neonicotinoid pesticides on honey bees and wild bees. Science 2017, 356, 1393–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergara-Amado, J.; Manzi, C.; Franco, L.M.; Contecha, S.C.; Marquez, S.J.; Solano-Iguaran, J.J.; Haro, R.E.; Silva, A.X. Effects of residual doses of neonicotinoid (imidacloprid) on metabolic rate of queen honey bees Apis mellifera (Hymenoptera: Apidae). Apidologie 2020, 51, 1091–1099. [Google Scholar] [CrossRef]
- Bass, C.; Field, L.M. Neonicotinoids. Curr. Biol. 2018, 28, R772–R773. [Google Scholar] [CrossRef] [Green Version]
- Tasei, J.N.; Lerin, J.; Ripault, G. Sub-lethal effects of imidacloprid on bumblebees, Bombus terrestris (Hymenoptera: Apidae), during a laboratory feeding test. Pest. Manag. Sci. 2000, 56, 784–788. [Google Scholar] [CrossRef]
- Moncharmont, F.X.D.; DeCourtye, A.; Hennequet-Hantier, C.; Pons, O.; Pham-Delegue, M.H. Statistical analysis of honeybee survival after chronic exposure to insecticides. Environ. Toxicol. Chem. 2003, 22, 3088–3094. [Google Scholar] [CrossRef] [Green Version]
- Diaz, T.; del-Val, E.; Ayala, R.; Larsen, J. Alterations in honey bee gut microorganisms caused by Nosema spp. and pest control methods. Pest. Manag. Sci. 2019, 75, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Syromyatnikov, M.Y.; Isuwa, M.M.; Savinkova, O.V.; Derevshchikova, M.I.; Popov, V.N. The effect of pesticides on the microbiome of animals. Agriculture 2020, 10, 79. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Qi, S.; Xue, X.; Niu, X.; Wu, L. Nitenpyram disturbs gut microbiota and influences metabolic homeostasis and immunity in honey bee (Apis mellifera L.). Environ. Pollut. 2020, 258, 113671. [Google Scholar] [CrossRef]
- Henríquez-Piskulich, P.A.; Schapheer, C.; Vereecken, N.J.; Villagra, C. Agroecological strategies to safeguard insect pollinators in biodiversity hotspots: Chile as a case study. Sustainability 2021, 13, 6728. [Google Scholar] [CrossRef]
- Rodríguez, S.S.; Pérez-Giraldo, L.C.; Vergara, P.M.; Carvajal, M.A.; Alaniz, A.J. Native bees in Mediterranean semi-arid agroecosystems: Unravelling the effects of biophysical habitat, floral resource, and honeybees. Agr. Ecosyst. Environ. 2021, 307, 107188. [Google Scholar] [CrossRef]
- Smith-Ramirez, C.; Ramos-Jiliberto, R.; Valdovinos, F.S.; Martinez, P.; Castillo, J.A.; Armesto, J.J. Decadal trends in the pollinator assemblage of Eucryphia cordifolia in Chilean rainforests. Oecologia 2014, 176, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Jiliberto, R.; Albornoz, A.A.; Valdovinos, F.S.; Smith-Ramírez, C.; Arim, M.; Armesto, J.J.; Marquet, P.A. A network analysis of plant–pollinator interactions in temperate rain forests of Chiloé Island, Chile. Oecologia 2009, 160, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Medel, R.; González-Browne, C.; Salazar, D.A.; Ferrer, P.; Ehrenfeld, M. The most effective pollinator principle applies to new invasive pollinators. Biol. Lett. 2018, 30, art14. [Google Scholar] [CrossRef] [PubMed]
- Arbetman, M.P.; Meeus, I.; Morales, C.L.; Aizen, M.A.; Smagghe, G. Alien parasite hitchhikes to Patagonia on invasive bumblebee. Biol. Invasions 2013, 15, 489–494. [Google Scholar] [CrossRef]
- Arismendi, N.; Riveros, G.; Zapata, N.; Smagghe, G.; Gonzalez, C.; Vargas, M. Occurrence of bee viruses and pathogens associated with emerging infectious diseases in native and non-native bumble bees in southern Chile. Biol. Invasions 2021, 23, 1175–1189. [Google Scholar] [CrossRef]
- González-Varo, J.P.; Biesmeijer, J.C.; Bommarco, R.; Potts, S.G.; Schweiger, O.; Smith, H.G.; Steffan-Dewenter, I.; Szentgyorgyi, H.; Woyciechowski, M.; Vilà, M. Combined effects of global change pressures on animal-mediated pollination. Trends Ecol. Evol. 2013, 28, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Magrach, A.; González-Varo, J.P.; Boiffier, M.; Vilà, M.; Bartomeus, I. Honeybee spillover reshuffles pollinator diets and affects plant reproductive success. Nat. Ecol. Evol. 2017, 1, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Morales, C.L.; Aizen, M.A. Invasive mutualisms and the structure of plant-pollinator interactions in the temperate forests of north-west Patagonia, Argentina. J. Ecol. 2006, 94, 171–180. [Google Scholar] [CrossRef]
- Ruz, L.; Herrera, R. Preliminary observations on foraging activities of Bombus dahlbomii and Bombus terrestris (Hym: Apidae) on native and non-native vegetation in Chile. In Proceedings of the VIII International Symposium on Pollination-Pollination: Integrator of Crops and Native Plant Systems, Mosonmagyaróvár, Hungary, 10–14 July 2000; Volume 561, pp. 165–169. [Google Scholar]
- Valdivia, C.E.; Carroza, J.P.; Orellana, J.I. Geographic distribution and trait-mediated causes of nectar robbing by the European bumblebee Bombus terrestris on the Patagonian shrub Fuchsia magellanica. Flora 2016, 225, 30–36. [Google Scholar] [CrossRef]
- Esterio, G.; Cares-Suarez, R.; Gonzalez-Browne, C.; Salinas, P.; Carvallo, G.; Medel, R. Assessing the impact of the invasive buff-tailed bumblebee (Bombus terrestris) on the pollination of the native Chilean herb Mimulus luteus. Arthropod-Plant. Interact. 2013, 7, 467–474. [Google Scholar] [CrossRef]
- Montalva, J.; Arroyo, M.T.K.; Ruz, L. Bombus terrestris Linnaeus (Hymenoptera: Apidae: Bombini) en Chile: Causas y consecuencias de su introducción. Rev. Jardín Botánico Chagual 2008, 6, 13–20. [Google Scholar]
- Carvallo, G.O.; Medel, R.; Navarro, L. Assessing the effects of native plants on the pollination of an exotic herb, the blueweed Echium vulgare (Boraginaceae). Arthropod-Plant. Interact. 2013, 7, 475–484. [Google Scholar] [CrossRef]
- Aldunce, P.; Araya, D.; Sapiain, R.; Ramos, I.; Lillo, G.; Urquiza, A.; Garreaud, R. Local perception of drought impacts in a changing climate: The mega-drought in central Chile. Sustainability 2017, 9, 2053. [Google Scholar] [CrossRef] [Green Version]
- Boisier, J.P.; Rondanelli, R.; Garreaud, R.D.; Muñoz, F. Anthropogenic and natural contributions to the Southeast Pacific precipitation decline and recent megadrought in central Chile. Geophys Res. Lett 2016, 43, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Garreaud, R.D.; Alvarez-Garreton, C.; Barichivich, J.; Boisier, J.P.; Christie, D.; Galleguillos, M.; LeQuesne, C.; McPhee, J.; Zambrano-Bigiarini, M. The 2010-2015 megadrought in central Chile: Impacts on regional hydroclimate and vegetation. Hydrol. Earth Syst. Sc. 2017, 21, 6307–6327. [Google Scholar] [CrossRef] [Green Version]
- Fontúrbel, F.E.; Lara, A.; Lobos, D.; Little, C. The cascade impacts of climate change could threaten key ecological interactions. Ecosphere 2018, 9, e02485. [Google Scholar] [CrossRef] [Green Version]
- Novoa, V.; Ahumada-Rudolph, R.; Rojas, O.; Saez, K.; de la Barrera, F.; Arumi, J.L. Understanding agricultural water footprint variability to improve water management in Chile. Sci. Total Environ. 2019, 670, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Barria, P.; Rojas, M.; Moraga, P.; Murnoz, A.; Bozkurt, D.; Alvarez-Garreton, C. Anthropocene and streamflow: Long-term perspective of streamflow variability and water rights. Elementa-Sci. Anthrop. 2019, 7, 1–15. [Google Scholar] [CrossRef]
- Arroyo, M.T.K.; Robles, V.; Tamburrino, I.; Martinez-Harms, J.; Garreaud, R.D.; Jara-Arancio, P.; Pliscoff, P.; Copier, A.; Arenas, J.; Keymer, J.; et al. Extreme drought affects visitation and seed set in a plant species in the central Chilean Andes heavily dependent on hummingbird pollination. Plants 2020, 9, 1553. [Google Scholar] [CrossRef] [PubMed]
- Rais, A.; van de Kuilen, J.W.G.; Pretzsch, H. Growth reaction patterns of tree height, diameter, and volume of Douglas-fir (Pseudotsuga menziesii [Mirb.] Franco) under acute drought stress in Southern Germany. Eur. J. Forest Res. 2014, 133, 1043–1056. [Google Scholar] [CrossRef]
- Bell, D.M.; Pabst, R.J.; Shaw, D.C. Tree growth declines and mortality were associated with a parasitic plant during warm and dry climatic conditions in a temperate coniferous forest ecosystem. Glob. Chang. Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Miranda, A.; Lara, A.; Altamirano, A.; Di Bella, C.; Gonzalez, M.E.; Camarero, J.J. Forest browning trends in response to drought in a highly threatened mediterranean landscape of South America. Ecol. Indic. 2020, 115, 106401. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C.; Carrasco, B.; Orchard, C. Combining niche modelling, land-use change, and genetic information to assess the conservation status of Pouteria splendens populations in central Chile. Int. J. Ecol. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepulveda, H.H.; Veloso-Aguila, D. The Central Chile Mega Drought (2010-2018): A climate dynamics perspective. Int. J. Climatol. 2020, 40, 421–439. [Google Scholar] [CrossRef]
- Memmott, J.; Craze, P.G.; Waser, N.M.; Price, M.V. Global warming and the disruption of plant-pollinator interactions. Ecol. Lett. 2007, 10, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Blüthgen, N.; Klein, A.M. Functional complementarity and specialisation: The role of biodiversity in plant-pollinator interactions. Basic Appl. Ecol. 2011, 12, 282–291. [Google Scholar] [CrossRef]
- Pitts-Singer, T.L.; Artz, D.R.; Peterson, S.S.; Boyle, N.K.; Wardell, G.I. Examination of a managed pollinator strategy for almond production using Apis mellifera (Hymenoptera: Apidae) and Osmia lignaria (Hymenoptera: Megachilidae). Environ. Entomol. 2018, 47, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Aizen, M.A.; Vásquez, D.P.; Smith-Ramírez, C. Historial natural y conservación de los mutualismos planta-animal del bosque templado de Sudamérica austral. Rev. Chil. Hist. Nat. 2002, 75, 79–97. [Google Scholar] [CrossRef]
- Smith-Ramírez, C. Los picaflores y su recurso floral en el bosque templado de la isla de Chiloé, Chile [Hummingbirds and their floral resources in temperate forests of Chiloé island, Chile]. Rev. Chil. Hist. Nat. 1993, 66, 65–73. [Google Scholar]
- Hornung-Leoni, C.T.; Gonzalez-Gomez, P.L.; Troncoso, A.J. Morphology, nectar characteristics and avian pollinators in five Andean Puya species (Bromeliaceae). Acta Oecol. 2013, 51, 54–61. [Google Scholar] [CrossRef]
- Guerrero, P.C.; Antinao, C.A.; Vergara-Merino, B.; Villagra, C.A.; Carvallo, G.O. Bees may drive the reproduction of four sympatric cacti in a vanishing coastal mediterranean-type ecosystem. PeerJ 2019, 7, e7865. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.S.; Ceríaco, L.M.; Daniel, G.M.; Dellapé, P.M.; Löbl, I.; Marinov, M.; Reis, R.E.; Young, M.T.; Dubois, A.; Agarwal, I. The taxonomic impediment: A shortage of taxonomists, not the lack of technical approaches. Zool. J. Linn. Soc. 2021, 193, 381–387. [Google Scholar] [CrossRef]
- Knight, T.M.; Ashman, T.L.; Bennett, J.M.; Burns, J.H.; Passonneau, S.; Steets, J.A. Reflections on, and visions for, the changing field of pollination ecology. Ecol. Lett. 2018, 21, 1282–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuadra-Valdés, J.; Vizentin-Bugoni, J.; Fontúrbel, F.E. An exotic magnet plant alters pollinator abundance and behavior: A field test with a native mistletoe. Biol. Invasions 2021. [Google Scholar] [CrossRef]
- Johnson, S.D.; Peter, C.I.; Nilsson, L.A.; Agren, J. Pollination success in a deceptive orchid is enhanced by co-occurring rewarding magnet plants. Ecology 2003, 84, 2919–2927. [Google Scholar] [CrossRef] [Green Version]
- Kovács-Hostyánszki, A.; Espíndola, A.; Vanbergen, A.J.; Settele, J.; Kremen, C.; Dicks, L.V. Ecological intensification to mitigate impacts of conventional intensive land use on pollinators and pollination. Ecol. Lett. 2017, 20, 673–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polit-Granja, E. Situación de las Viñas y los Vinos Orgánicos Chilenos; Oficina de Estudios y Políticas Agrarias (ODEPA): Santiago de Chile, Chile, 2005.
- Barbosa, O.; Villagra, P. Socio-ecological studies in urban and rural ecosystems in Chile. In Earth Stewardship: Linking Ecology and Ethics in Theory and Practice; Rozzi, R., Chapin, F.S., Callicott, J.B., Pickett, S.T.A., Power, M.E., Armesto, J.J., May, R.H., Eds.; Springer: Cham, Switzerland, 2015; pp. 297–311. [Google Scholar]
- Pandya, R.E. A framework for engaging diverse communities in citizen science in the US. Front. Ecol. Environ. 2012, 10, 314–317. [Google Scholar] [CrossRef]
- Giovanetti, M.; Albertazzi, S.; Flaminio, S.; Ranalli, R.; Bortolotti, L.; Quaranta, M. Pollination in agroecosystems: A review of the conceptual framework with a view to sound monitoring. Land 2021, 10, 540. [Google Scholar] [CrossRef]
- Decourtye, A.; Armengaud, C.; Renou, M.; Devillers, J.; Cluzeau, S.; Gauthier, M.; Pham-Delegue, M.H. Imidacloprid impairs memory and brain metabolism in the honeybee (Apis mellifera L.). Pestic Biochem. Phys. 2004, 78, 83–92. [Google Scholar] [CrossRef]
- Guez, D.; Suchail, S.; Gauthier, M.; Maleszka, R.; Belzunces, L.P. Contrasting effects of imidacloprid on habituation in 7- and 8-day-old honeybees (Apis mellifera). Neurobiol. Learn. Mem. 2001, 76, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Siviter, H.; Muth, F. Do novel insecticides pose a threat to beneficial insects? P. Roy. Soc. B-Biol. Sci. 2020, 287, 20201265. [Google Scholar] [CrossRef] [PubMed]
- SAG. Listado de Plaguicidas de Uso Agrícola Prohibidos en Chile. Última actualización 24 de febrero de 2020. Código: F-RIS-RAI-PA-013; Servicio Agrícola y Ganadero: Santiago de Chile, Chile, 2014.
- Cordero, S.; Castano-Villa, G.J.; Fontúrbel, F.E. The best bang for the bucks: Rethinking global investment in biodiversity conservation. Sustainability 2020, 12, 9252. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gap | Description | Opportunity |
|---|---|---|
| Lack of local data | Systematized databases and the low number of pollination and pollinator studies in the country are a barrier to promote evidence-based decision-making processes and regulation towards the conservation of pollinators. | Promote local and country-wide studies to assess pollinator diversity across temporal and spatial scales. Depositing local data on GBIF and local/national institutions. |
| High proportion of exotic species | Plant–pollinator networks are dominated by exotic plants and pollinator species in agricultural landscapes. This affects native pollinators by displacing them from these areas. | Use scientific evidence and citizen science to monitor, reinforce the exotic species control, and influence legislation to stop new importations. Promote transboundary agreements to prevent undesired species introductions and invasions. |
| Lack of regulation | In Chile, the protection of biodiversity is still challenging due to scarce regulation and weak institutional governance. Therefore, the conservation of native pollinator species is largely dependent on local voluntary actions, which might increase when crop pollination services are visible. | Create public awareness and establish dialog spaces between scientists and decisionmakers to influence legislation. Develop a national pollinator conservation strategy. Improve governance empowering a state agency for biodiversity protection. |
| Lack of monitoring schemes | There are no formal monitoring schemes on pollinators in the country which contributes to the lack of local information on the conservation status of pollinator species in the country. | Implement citizen science endeavors and standardized monitoring schemes. Red list the native pollinators following IUCN criteria and use this to set conservation priorities. |
| Unsustainable agricultural management | Agricultural landscapes are detrimental for pollinators when managed under unsustainable practices, such as high chemical inputs, large extensions of monocultures, and the suppression of non-crop habitats. | Provide evidence of the value of wild pollinators to farmers, and management practices that promote pollinators in agricultural landscapes (e.g., pollinator-friendly areas, nesting/refugee sites, natural vegetation corridors). |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieli, L.; Murúa, M.M.; Flores-Prado, L.; Carvallo, G.O.; Valdivia, C.E.; Muschett, G.; López-Aliste, M.; Andía, C.; Jofré-Pérez, C.; Fontúrbel, F.E. Local Actions to Tackle a Global Problem: A Multidimensional Assessment of the Pollination Crisis in Chile. Diversity 2021, 13, 571. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110571
Vieli L, Murúa MM, Flores-Prado L, Carvallo GO, Valdivia CE, Muschett G, López-Aliste M, Andía C, Jofré-Pérez C, Fontúrbel FE. Local Actions to Tackle a Global Problem: A Multidimensional Assessment of the Pollination Crisis in Chile. Diversity. 2021; 13(11):571. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110571
Chicago/Turabian StyleVieli, Lorena, Maureen M. Murúa, Luis Flores-Prado, Gastón O. Carvallo, Carlos E. Valdivia, Giselle Muschett, Manuel López-Aliste, Constanza Andía, Christian Jofré-Pérez, and Francisco E. Fontúrbel. 2021. "Local Actions to Tackle a Global Problem: A Multidimensional Assessment of the Pollination Crisis in Chile" Diversity 13, no. 11: 571. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110571