
It’s All about the Zone: Spider Assemblages in Different Ecological Zones of Levantine Caves

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cave Surveys
2.2. Statistical Analysis
3. Results
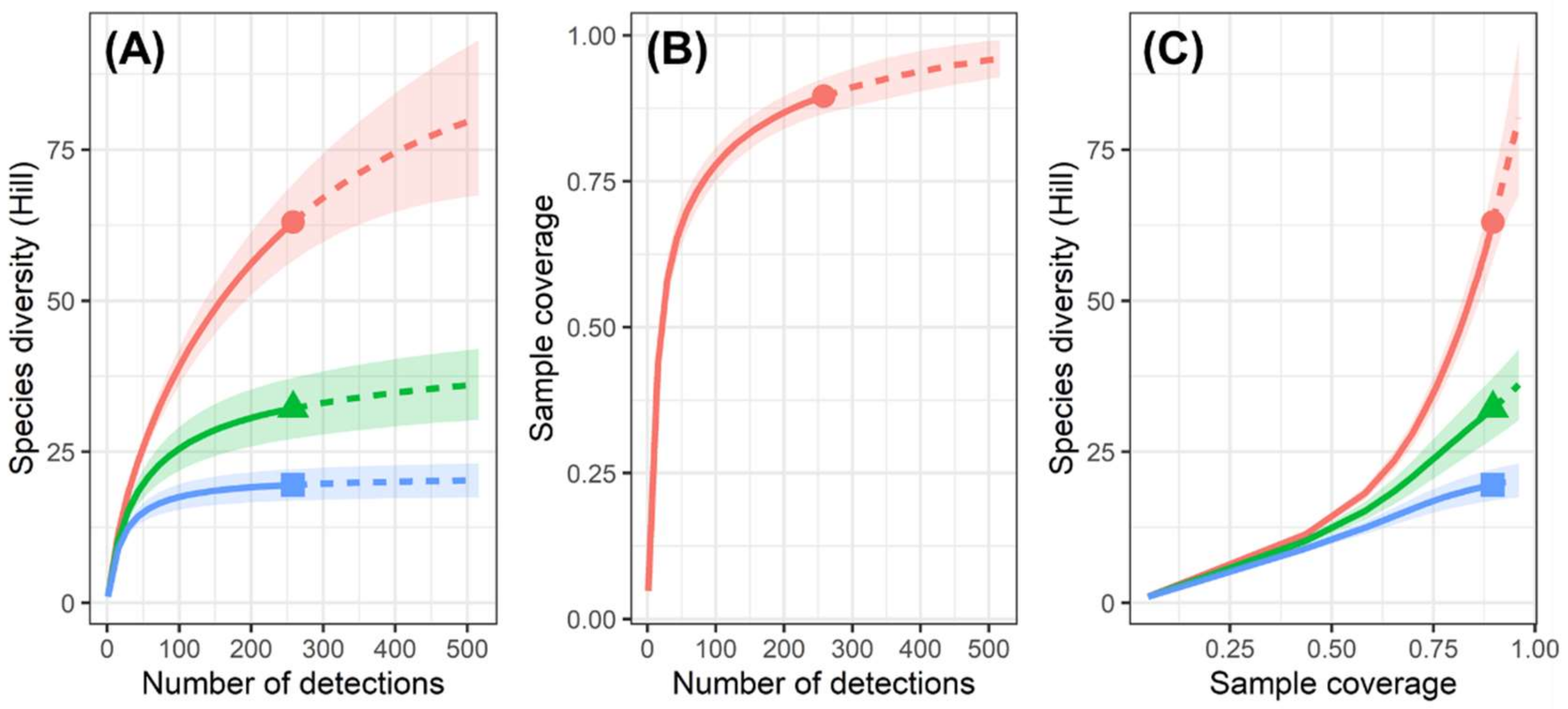
3.1. Spider Assemblage Diversity in Cave Zones in the Southern Levant
3.2. Zone Assemblage Comparison
3.3. Co-Occurrence of Spiders in Cave Zones
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Materials and Methods
Appendix A.2. Results


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Category | Dominant Zone | Dev | p | Other Associations | Interactions |
|---|---|---|---|---|---|---|
| Artema nephilit | Troglophile | Entrance/Twilight | 31.996 | 0.001 | + Temperature (Dev = 31.996, p = 0.001) − Elevation (Dev = 105.579, p =0.001) | - |
| Filistata insidiatrix | Troglophile | Entrance | 35.957 | 0.001 | − Temperature (Dev = 22.451, p = 0.001) − Elevation (Dev = 52.873, p = 0.001) N Region (Dev = 69.439, p = 0.001) | Zone/Region (Dev = 11.065, p = 0.062) |
| Filistata sp. | Troglophile | Entrance | 31.16 | 0.001 | C Region (Dev = 43.446, p = 0.001) | |
| Holocnemus pluchei | Troglophile | Entrance | 49.336 | 0.001 | − Temperature (Dev = 50.909, p = 0.001) − Elevation (Dev = 50.702, p = 0.001) | Zone/Region (Dev = 12.158, p = 0.043) |
| Pholcus sp. | Troglophile | Entrance/Twilight | 11.861 | 0.066 | C Region (Dev = 31.48, p = 0.001) | - |
| Steatoda Zavoa sp. | Troglophile | Dark | 12.275 | 0.058 | + Temperature (Dev = 51.772, p = 0.001) + Elevation (Dev = 71.376, p = 0.001) | - |
| Tegenaria Galilee sp. | Troglobite | Twilight/Dark | 34.444 | 0.001 | − Temperature (Dev = 9.696, p = 0.068) + Elevation (Dev = 29.507, p = 0.001) N Region (Dev = 31.32, p = 0.001) | Zone/Temperature (Dev = 53.994, p = 0.001) |
| Tegenaria Judea sp. | Troglobite | Twilight | 17.107 | 0.006 | − Temperature (Dev = 17.107, p = 0.006) C Region (Dev = 16.456, p = 0.005) | - |
| Uloborus plumipes | Accidental | Entrance | 22.425 | 0.001 | C Region (Dev = 29.468, p = 0.001) | - |
| Tegenaria Zavoa sp. | Troglophile | Dark | 39.491 | 0.001 | + Temperature (Dev = 158.41, p = 0.001) | - |
Appendix A.3. Discussion
References
- Snowman, C.V.; Zigler, K.S.; Hedin, M. Caves as Islands: Mitochondrial Phylogeography of the Cave-Obligate Spider Species Nesticus Barri (Araneae: Nesticidae). J. Arachnol. 2010, 38, 49–56. [Google Scholar] [CrossRef]
- Poulson, T.L.; White, W.B. The Cave Environment. Science 1969, 165, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Howarth, F.G. High-Stress Subterranean Habitats and Evolutionary Change in Cave-Inhabiting Arthropods. Am. Nat. 1993, 142, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Manenti, R.; Lunghi, E.; Ficetola, G.F. The Distribution of Cave Twilight-Zone Spiders Depends on Microclimatic Features and Trophic Supply. Invertebr. Biol. 2015, 134, 242–251. [Google Scholar] [CrossRef]
- Mammola, S. Finding Answers in the Dark: Caves as Models in Ecology Fifty Years after Poulson and White. Ecography 2019, 42, 1331–1351. [Google Scholar] [CrossRef] [Green Version]
- Tobin, B.W.; Hutchins, B.T.; Schwartz, B.F. Spatial and Temporal Changes in Invertebrate Assemblage Structure from the Entrance to Deep-Cave Zone of a Temperate Marble Cave. Int. J. Speleobiol. 2013, 42, 203–221. [Google Scholar] [CrossRef] [Green Version]
- Mammola, S.; Piano, E.; Isaia, M. Step Back! Niche Dynamics in Cave-Dwelling Predators. Acta Oecologica 2016, 75, 35–42. [Google Scholar] [CrossRef]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Cave Features, Seasonality and Subterranean Distribution of Non-Obligate Cave Dwellers. PeerJ 2017, 5, e3169. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats, 2nd ed.; Oxford University Press: Oxford, UK, 2019; ISBN 978019882076-5. [Google Scholar]
- Trajano, E.; de Carvalho, M.R. Towards a Biologically Meaningful Classification of Subterranean Organisms: A Critical Analysis of the Schiner-Racovitza System from a Historical Perspective, Difficulties of Its Application and Implications for Conservation. Subterr. Biol. 2017, 22, 1–26. [Google Scholar] [CrossRef]
- Trajano, E. Ecological Classification of Subterranean Organisms. In Encyclopedia of Caves; Elsevier: Cham, Switzerland, 2012; pp. 275–277. ISBN 978-0-12-383832-2. [Google Scholar]
- Avni, Y. Tectonic and Physiographic Settings of the Levant. In Quaternary of the Levant: Environments, Climate Change, and Humans; Bar-Yosef, O., Enzel, Y., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 3–16. ISBN 9781107090460. [Google Scholar]
- Tchernov, E.; Belmaker, M. The biogeographic history of the fauna in the Southern Levant and the correlation with climatic change. In The Red Book. Vertebrates of Israel; Dolev, A., Perevolototsky, A., Tchernov, E., Belmaker, M., Eds.; Gefen Publishing: Jerusalem, Israel, 2004; pp. 21–25. [Google Scholar]
- Belmaker, M. Biogeography and Palaeoecology of the Early Pleistocene Large Mammals in the Levant. In Quaternary of the Levant: Environments, Climate Change, and Humans; Enzel, Y., Bar-Yosef, O., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 355–362. ISBN 978-1-107-09046-0. [Google Scholar]
- Por, F.D. An Outline of the Zoogeography of the Levant. Zool. Scr. 1975, 4, 5–20. [Google Scholar] [CrossRef]
- Tchernov, E. The biogeographical history of the southern Levant. In The Zoogeography of Israel. The Distribution and Abundance at a Zoogeographical Crossroad, Monographiae Biologicae 62; Yom-Tov, Y., Tchernov, E., Eds.; Dr. W. Junk Publishers: Dordrecht, The Netherlands, 1988; pp. 159–250. ISBN 9061936500. [Google Scholar]
- Zonstein, S.; Marusik, Y.M. Checklist of the Spiders (Araneae) of Israel. Zootaxa 2013, 3671, 1. [Google Scholar] [CrossRef] [PubMed]
- Mammola, S.; Cardoso, P.; Ribera, C.; Pavlek, M.; Isaia, M. A Synthesis on Cave-Dwelling Spiders in Europe. J. Zool. Syst. Evol. Res. 2017, 56, 301–316. [Google Scholar] [CrossRef]
- Gavish-Regev, E.; Aharon, S.; Steinpress, I.A.; Seifan, M.; Lubin, Y. A Primer on Spider Assemblages in Levantine Caves: The Neglected Subterranean Habitats of the Levant—A Biodiversity Mine. Diversity 2021, 13, 179. [Google Scholar] [CrossRef]
- Wang, Y.; Naumann, U.; Wright, S.T.; Warton, D.I. Mvabund—An R Package for Model-Based Analysis of Multivariate Abundance Data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2020. Available online: https://www.r-project.org/index.html (accessed on 18 February 2021).
- Chao, A.; Gotelli, N.; Hsieh, T.; Sander, E.; Ma, K.; Colwell, R.; Ellison, A. Rarefaction and Extrapolation with Hill Numbers: A Framework for Sampling and Estimation in Species Diversity Studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Roswell, M.; Dushoff, J.; Winfree, R. A Conceptual Guide to Measuring Species Diversity. Oikos 2021, 130, 321–338. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. INEXT: An R Package for Interpolation and Extrapolation of Species Diversity (Hill Numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package Version 2.2-0. Available online: http://CRAN.Rproject.org/package=vegan (accessed on 18 February 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis. Media 2016, 35, 10–1007. [Google Scholar]
- Griffith, D.M.; Veech, J.A.; Marsh, C.J. Cooccur: Probabilistic Species Co-Occurrence Analysis in R. J. Stat. Softw. 2016, 69, 1–17. [Google Scholar] [CrossRef]
- Atmar, W.; Patterson, B.D. The Measure of Order and Disorder in the Distribution of Species in Fragmented Habitat. Oecologia 1993, 96, 373–382. [Google Scholar] [CrossRef]
- Rodríguez-Gironés, M.A.; Santamaría, L. A New Algorithm to Calculate the Nestedness Temperature of Presence–Absence Matrices. J. Biogeogr. 2006, 33, 924–935. [Google Scholar] [CrossRef]
- Heino, J.; Mykrä, H.; Muotka, T. Temporal Variability of Nestedness and Idiosyncratic Species in Stream Insect Assemblages. Biodivers. Res. 2009, 15, 198–206. [Google Scholar] [CrossRef]
- Gavish-Regev, E.; Aharon, S.; Armiach, I.; Lubin, Y. Cave Survey Yields a New Spider Family Record for Israel. Arachnol. Mitt. 2016, 51, 39–42. [Google Scholar] [CrossRef]
- Miranda, G.S.; Aharon, S.; Gavish-Regev, E.; Giupponi, A.P.L.; Wizen, G. A New Species of Charinus Simon, 1892 (Arachnida: Amblypygi: Charinidae) from Israel and New Records of C. Ioanniticus (Kritscher, 1959). Eur. J. Taxon. 2016, 234, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Aharon, S.; Huber, B.A.; Gavish-Regev, E. Daddy-Long-Leg Giants: Revision of the Spider Genus Artema Walckenaer, 1837 (Araneae, Pholcidae). Eur. J. Taxon. 2017, 376. [Google Scholar] [CrossRef] [Green Version]
- Aharon, S.; Ballesteros, J.A.; Crawford, A.R.; Friske, K.; Gainett, G.; Langford, B.; Santibáñez-López, C.E.; Ya’aran, S.; Gavish-Regev, E.; Sharma, P.P. The Anatomy of an Unstable Node: A Levantine Relict Precipitates Phylogenomic Dissolution of Higher-Level Relationships of the Armoured Harvestmen (Arachnida: Opiliones: Laniatores). Invertebr. Syst. 2019, 33, 697–717. [Google Scholar] [CrossRef]
- Wilson, E.O.; O’Brien, R.D.; Susman, M.; Boggs, W.E.; Eisner, T.; Briggs, W.R.; Dickerson, R.E.; Metzenberg, R.L. The Multiplication of Species. In Life on Earth; Sinauer Associates, Inc.: Stamford, CT, USA, 1973; pp. 824–877. [Google Scholar]
- Howarth, F. The Evolution of Non-Relictual Tropical Troglobites. Int. J. Speleol. 1987, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ruzicka, V. The First Steps in Subterranean Evolution of Spiders (Araneae) in Central Europe. J. Nat. Hist. 1999, 33, 255–265. [Google Scholar] [CrossRef]
- Barr, T.; Holsinger, J.R. Speciation in Cave Faunas. Annu. Rev. Ecol. Syst. 1985, 16, 313–337. [Google Scholar] [CrossRef]
- Huber, B.A. Cave-Dwelling Pholcid Spiders (Araneae, Pholcidae): A Review. Subterr. Biol. 2018, 26, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Mammola, S.; Goodacre, S.L.; Isaia, M. Climate Change May Drive Cave Spiders to Extinction. Ecography 2018, 41, 233–243. [Google Scholar] [CrossRef] [Green Version]








| Species | Category | Dominant Zone | Dev | p | Other Associations | Interactions |
|---|---|---|---|---|---|---|
| Artema nephilit | Troglophile | Twilight/Entrance | 34.520 | 0.001 | −Elevation (Dev = 40.978, p = 0.001) | - |
| Filistata insidiatrix | Troglophile | Entrance/Twilight | 15.973 | 0.005 | −Temperature (Dev = 33.296, p = 0.001) −Elevation (Dev = 10.758, p = 0.027) N Region (Dev = 29.727, p = 0.001) | Zone/Temperature (Dev = 19.662, p = 0.002) |
| Filistata sp. | Troglophile | Entrance | 53.846 | 0.001 | C Region (Dev = 69.669, p = 0.001) | - |
| Holocnemus pluchei | Troglophile | Entrance | 36.565 | 0.001 | −Temperature (Dev = 35.308, p = 0.001) −Elevation (Dev = 30.028, p = 0.001) | - |
| Hoplopholcus cecconii | Troglophile | Dark | 24.826 | 0.001 | + Elevation (Dev = 17.750, p = 0.004) N Region (Dev = 12.346, p = 0.042) | - |
| Loxosceles rufescens | Troglophile | Entrance | 8.525 | 0.129 | N Region (Dev = 52.706, p = 0.001) | Zone/Region (Dev = 25.701, p = 0.001) Zone/Temperature (Dev = 19.662, p = 0.002) |
| Micronetinae sp. (1) | Troglophile | Entrance | 8.585 | 0.129 | - | Zone/Temperature (Dev = 12.237, p = 0.008) |
| Oecobius sp. | Accidental | Entrance | 15.224 | 0.008 | + Elevation (Dev = 21.825, p = 0.001) | - |
| Pholcus sp. | Troglophile | Twilight | 11.537 | 0.032 | C Region (Dev = 31.480, p = 0.001) | - |
| Steatoda triangulosa | Troglophile | Entrance | 21.631 | 0.001 | - | Zone/Region (Dev = 17.309, p = 0.002) Zone/Temperature (Dev = 13.875, p = 0.006) |
| Tegenaria angustipalpis | Troglophile | Entrance | 17.133 | 0.003 | + Elevation (Dev = 11.189, p = 0.027) N Region (Dev = 18.917, p = 0.001) | - |
| Tegenaria pagana | Troglophile | Entrance/Twilight | 3.359 | 0.858 | − Elevation (Dev = 19.611, p = 0.003) N/C Region (Dev = 53.838, p = 0.001) | Zone/Region (Dev = 19.935, p = 0.001) Zone/Temperature (Dev = 27.866, p = 0.001) Zone/Elevation (Dev = 37.165, p = 0.001) |
| Tegenaria sp. from Galilee caves | Troglobite | Twilight/Dark | 19.865 | 0.001 | + Elevation (Dev = 16.046, p = 0.006) N Region (Dev = 27.424, p = 0.001) | Zone/Temperature (Dev = 13.034, p = 0.006) |
| Tegenaria sp. from Zavoa cave | Troglophile | Dark | 38.368 | 0.001 | + Temperature (Dev = 81.525, p = 0.001) | - |
| Species 1 | Species 2 | Expected Co-Occurrences | Observed Co-Occurrences | Co-Occurrence Relationship | p-Value |
|---|---|---|---|---|---|
| Artema nephilit | Hoplopholcus cecconii | 2.4 | 0 | − | 0.048 |
| Artema nephilit | Steatoda triangulosa | 3.9 | 1 | − | 0.047 |
| Chaetopelma sp. | Loxosceles rufescens | 2.5 | 6 | + | 0.004 |
| Filistata insidiatrix | Holocnemus pluchei | 6 | 11 | + | 0.005 |
| Filistata insidiatrix | Loxosceles rufescens | 8.8 | 15 | + | 0.001 |
| Filistata insidiatrix | Tegenaria pagana | 8.8 | 14 | + | 0.006 |
| Holocnemus pluchei | Loxosceles rufescens | 7.9 | 12 | + | 0.026 |
| Holocnemus pluchei | Micronetinae sp. 1 | 1.7 | 4 | + | 0.05 |
| Holocnemus pluchei | Steatoda triangulosa | 3.7 | 7 | + | 0.03 |
| Holocnemus pluchei | Uloborus plumipes | 2.3 | 5 | + | 0.036 |
| Loxosceles rufescens | Micronetinae sp. 1 | 2.5 | 5 | + | 0.042 |
| Loxosceles rufescens | Tegenaria pagana | 11.7 | 18 | + | 0.002 |
| Pholcus sp. | Tegenaria pagana | 2.5 | 5 | + | 0.042 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuff, J.P.; Aharon, S.; Armiach Steinpress, I.; Seifan, M.; Lubin, Y.; Gavish-Regev, E. It’s All about the Zone: Spider Assemblages in Different Ecological Zones of Levantine Caves. Diversity 2021, 13, 576. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110576
Cuff JP, Aharon S, Armiach Steinpress I, Seifan M, Lubin Y, Gavish-Regev E. It’s All about the Zone: Spider Assemblages in Different Ecological Zones of Levantine Caves. Diversity. 2021; 13(11):576. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110576
Chicago/Turabian StyleCuff, Jordan P., Shlomi Aharon, Igor Armiach Steinpress, Merav Seifan, Yael Lubin, and Efrat Gavish-Regev. 2021. "It’s All about the Zone: Spider Assemblages in Different Ecological Zones of Levantine Caves" Diversity 13, no. 11: 576. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110576