
Ecological, Historical, Economic, and Political Factors Shaping the Human–Gorilla Interface in the Mone-Oku Forest, Cameroon


Anthropology, School of Social Sciences, The University of Auckland, Auckland 1142, New Zealand
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(4), 175; https://0-doi-org.brum.beds.ac.uk/10.3390/d13040175
Submission received: 20 January 2021
/
Revised: 9 March 2021
/
Accepted: 16 April 2021
/
Published: 19 April 2021
(This article belongs to the Special Issue Humans and Wild Animals: Interactions in Deep Time, Recent History, and Now)
Abstract
:Cross River gorillas are the least numerous of the African ape taxa. Since their rediscovery, several organisations have sought to conserve these critically endangered apes, resulting in a “crisis conservation narrative” focused on the threats posed by local human activities. However, landscapes are not just ecological, they are also social, shaped by political and economic processes. This study examines the interconnections between humans and gorillas by approaching the Mone-Oku forest as a combination of ecological, social, and political systems. Through a combined use of botanical surveys, analyses of nesting sites, participant observation, and semistructured interviews, we obtained nuanced ecological and ethnographic insights into the human–gorilla interface. The results illustrate a history of alterations within the Mone-Oku forest, some of which are human-induced. These alterations have had both positive and negative outcomes for the gorillas and continue to the present day, where political history has shaped limited livelihood alternatives, increasing the reliance on a forest that has remained a constant in the more recent history of “developmental” neglect and isolation. However, this situation is not static, with future alterations to the forest also subject to regional and international political and economic influences, such as the increased worldwide demand for cacao.
1. Introduction
Humans have a long history of sympatry with nonhuman primates in Africa, with evidence suggesting that Cross River gorillas (Gorilla gorilla diehli) and humans have coexisted for at least 40,000 to 50,000 years in the Nigeria–Cameroon borderlands [1]. How humans have interacted with the gorillas and the forest over time, shaping the landscape we see today, however, remains largely unexplored. Rather, research agendas have focused on what some have labelled “the perilous state of the Cross River gorilla” [2] (p. 27) since their rediscovery and subspecies designation resulted in a critically endangered classification based on the low population estimates of 250–300 mature individuals combined with the ongoing threat of habitat loss [3,4]. This classification has contributed to a “crisis narrative”, whereby there is an urgent need to conserve these elusive apes, focusing research programs on the threats posed by local human communities who share the wider landscape [5]. While the anthropogenic nature of threats to the survival of the Cross River gorilla is not debated, the situation is complex, inherently challenging, and requires the integration of multiple approaches to understand the potential causes and consequences of anthropogenic actions over an extended period [6].
This study aims to step away from the crisis narrative by applying a dual ethnoprimatological and political ecology framework. Ethnoprimatology abandons the view that primates in contact with humans represents an “unnatural” situation; rather, it recognises that in many instances of ethnoprimatological enquiry, humans and primates have a long history of sympatry, whereby these entanglements have shaped the ecological and social landscapes we come to see today [7,8,9,10,11,12]. More recently, there has been a push to extend ethnoprimatological investigations to include multiple temporal and spatial axes through further integration with other disciplines [7,8,13,14,15]. As a large amount of ethnoprimatological research is concerned with the conservation of primates and areas of biodiversity, it is necessary to recognise and reflect on the ways that conserving nature is often based on Western scientific knowledge that privileges one way of knowing [7,16,17]. The use of a dual ethnoprimatological and political ecology framework offers the potential to regain the reflexivity required from a truly ethnographic ethnoprimatology [7].
Political ecologists have examined the politics of knowledge acquisition, highlighting that scientific research, such as the research driving the “biodiversity crisis”, is rooted in assumptions about the relationships between nature and humans [18,19,20,21,22,23]. Fairhead and Leach [24] stress the importance of considering the anthropogenic nature of ecosystems otherwise, people only see what they are seeking—human-caused degradation. This approach also recognises that a human community is never an isolated entity. Local systems link with national and international programs, policies, and economic opportunities allowing for an exploration of alternative causes of environmental degradation as opposed to what is usually proposed in the dominant narrative [23,25,26,27]. By investigating all aspects of anthropogenic influence on wider biodiversity, researchers are better placed to do more than document and monitor species decline [28]. This is due to the fact that people are actors within ecosystems and the links between ecology, policy, and economy are multidimensional and dialectical [29].
Cross River gorillas are found across a geographical range of 12,000 km2, of which they are found to occupy 700 km2 [30]. The majority of this population occurs in a landscape of mostly continuous forest in the Afi Mountain Wildlife Sanctuary, Nigeria, to the Kagwene Gorilla Sanctuary, Cameroon, with a small and likely isolated population in the Lebialem-Mone Forest Landscape [30]. This region has a markedly seasonal climate, with a longer and more intense dry season and higher elevations than the lowland tropical forests where western lowland gorillas range [31,32]. At each key locality, estimates of gorilla numbers range between 6 and 30 individuals [30]. Genetically, there are three subpopulations with the Western (Afi) and Eastern (Kagwene) subpopulations numbering between 20 and 30 individuals and a larger more genetically diverse Central subpopulation numbering between 160 and 230 individuals [33,34]. Recent migration by both sexes has been found to occur between all the subpopulations, although at a low rate [33]. However, there is evidence of a population bottleneck, suggesting a recent rapid reduction in population size, rather than structure, within the last 100–200 years, placing the smaller subpopulations at increased risk of diversity loss through genetic drift and inbreeding [33]. The dominant explanation for this population reduction is the increased availability of firearms throughout the 19th century making the hunting of larger mammals more accessible [30,33,34], although alternative contributing factors such as the introduction of novel diseases at the time remains unexplored [5].
The behaviour of the Cross River gorilla has largely been inferred through indirect ecological methods, such as nest site analysis and feeding signs, based on the ethical decision not to habituate study groups to a researcher’s presence as this increases their vulnerability to poaching when population numbers are already critical [31,35,36,37,38,39,40]. Cross River gorillas have demonstrated flexible responses to the seasonality of their environment through their diverse diet, ranging behaviours, and variable grouping patterns that also highlight the uniqueness of each locality [31,35,36,40]. The preference for nest sites to be located in areas of mid-elevation and montane forest, on steep and rocky slopes, that is often a fair distance from the nearest village has generated the assumption that Cross River gorillas avoid areas of human activity when selecting nest sites [38,41,42]; the relationship between human activities, food availability and gorilla presence is not straightforward and may be site-specific. Sawyer and Brashares [43] found the Mone-Oku gorillas were more likely to occupy areas of mid-elevation with reduced slope and suggest this may represent a compromise in behaviours between preferred food species and the avoidance of human activities.
Here, we begin to examine the history of interconnections between the people of Nga village and Cross River gorillas within the unprotected Mone-Oku Forest, Cameroon. Through a combined use of botanical surveys, analyses of nesting sites, participant observation, semistructured interviews, and archival research, we approach the forest as a combination of ecological, social and political systems. In particular, we aim to identify: (1) whether the gorillas had a preferred habitat type(s) for nest sites; (2) whether the gorillas are selective in the plant species used for the construction of night nests; (3) the location of night nests in relation to anthropogenic activities throughout the forest; and (4) how historical, political and economic forces manifest within Nga village.
2. Materials and Methods
2.1. Study Site
AW conducted this research in a 50 km2 area of the Mone-Oku Forest complex, Akwaya Subdivision, Southwest Cameroon (Figure 1). Specifically, the area is an unprotected forest corridor located between Mone Forest Reserve and the Mbulu Forest that provides tenuous links in the north to the protected Takamanda National Park and the Kagwene Gorilla Sanctuary. The forest is topographically diverse as elevations range between 130 and 1000 m a.s.l. and vegetation falls into the Guinea Congolian type [44]. It is a mosaic landscape of villages, farmland, secondary and primary forests. The climate is tropical with a protracted wet season from April to October and a shorter dry season from November to March [45]. As part of the Pleistocene refugium, there are high levels of species diversity, richness, and endemism across flora and fauna [32]. During this study, seven primate species were observed including the endangered Nigeria–Cameroon chimpanzee (Pan troglodytes ellioti) and the drill (Mandrillus leucophaeus). Previous surveys estimated within the forest complex there is a population of 6–15 unhabituated Cross River gorillas [30].
There are seven villages within the immediate area that access the forest and its resources. AW approached Nga village in May 2014 to take part in the ethnographic component of this research as they are traditional owners over part of the study site. With the permission of the village chief, meetings were held to describe the research and invite those who were interesting in participating to “opt-in”. The Nga people have two neighbouring village sites: Nga and Tito, approximately 5.5 km apart with a total population of around 500 people. The locate dialect is Manta; however, Pidgin English is widely spoken.
The area is fertile, and most individuals gain an income through small landholdings producing cocoa, oil palm, coconut, maize, vegetables, and fruits. Hunting, including the use of wire snares or locally made shotguns, is largely for subsistence. However, it is an immediate source of income when cash is required. The area suffers from limited vehicle transportation links to Mamfe, the nearest town (ca. 48 km), and the roads are only passable by motorbike during the rainy season. The Wildlife Conservation Society (WCS) has been consistently monitoring the gorillas in the region since 2009, with the initiation of the Gorilla Guardian Program (GGP). This program promotes the conservation of the Cross River gorilla and the Nigeria–Cameroon chimpanzee in villages outside of protected areas and provides individuals nominated within each village with tracking and ecological training [46].
2.2. Ethnographic Data Collection and Analysis
Twenty-eight semistructured interviews were conducted between September and October 2014 following a quota sampling approach for those who expressed an interest in participating, firstly by village site, then gender and age class [5]. AW administered interviews with the aid of a local Cameroonian field assistant in Pidgin English. When participants were unable to understand or express their ideas, translations were made from the dialect to English. All participants were residents; however, two had migrated with no formal marital ties to the region. The interviews were designed to discuss a range of topics including forest use, traditional conservation practices, farming practices, economic pathways, and village history, in addition to discussing perceptions of the Mone-Oku Forest and the Cross River gorillas.
AW used participant observation to expand upon the semistructured interviews and obtain ethnographic insight into the complexity of activities that make up social life in the villages of Eshobi and Nga [47]. Ethnographic fieldwork was carried out between September 2014 and March 2015 interspaced with periods of research in the forest. In total, AW spent 27 days within the village and a further 96 days in the forest with local field assistants. AW took a moderate level of participation with a focus on engaging in activities relating to farming, conservation activities, and forest use [47,48]. This included observing village meetings held by WCS when they were present for their GGP and working with them in their forest activities recording ape nest sites. AW developed strong relationships with her local field assistants who were happy to show her how they navigated and tracked animals in the forest when hunting and how to prepare bushmeat for sale. Through participant observation, AW was more readily positioned to explore what resources people use and how they use them [9,12]. Discussions with key informants clarified meanings and interpretations of major themes, which were then used to inform future observations [48]. AW’s field notes included both objective observations and her subjective feelings, recognizing these notes were also a product she constructed [49]. Reflexivity was a continual element throughout this research and analysis with consideration being given to AW’s position, as her understanding of the people who became involved in this research developed [50]. This involved reflecting and re-evaluating relationships with informants and how their participation may have caused themselves or others to change, through to acknowledging how interview surroundings may have affected the nature of interview data [50].
Interviews and field notes were entered in Excel documents and thematically coded. To preserve participants’ anonymity, where requested, we refer to participants by their occupation (e.g., farmer or hunter). AW also conducted research at the National Archives of Cameroon in Buea. The archives store British colonial documents including data regarding the economic and political policies of the British administration in Akwaya Subdivision. Archival and historical sources were used throughout our analysis to supplement, where possible, the oral histories provided by participants or to place these histories in a wider context.
2.3. Ecological Data Collection and Analysis
2.3.1. Surveying
AW conducted reconnaissance surveys between May 2014 and March 2015 to record human presence within the forest and to locate gorilla nest sites. Surveys covered 272 km within the study area. Transects were deemed unpractical due to both the difficult nature of the terrain and the potential costs involved in opening up new areas of the forest [51]. Surveys were conducted during daylight hours with the direction of travel usually determined by the location of the randomly selected botanical plot coordinates. Nest sites were additionally located from hunter observations and reports as part of the GGP or after opportunistically sighting the gorillas. Along each reconnaissance route, AW recorded all human signs as GPS waypoints (e.g., spent bullet cartridges, traps, bush huts and forest clearings). Bullet cartridges were collected when encountered to avoid a repeat count and were later disposed of. As the study site is an unclassified forest, any traps encountered were left intact except for those located on known chimpanzee and gorilla trails. This decision was made together with the village after a discussion regarding the legal ramifications if a hunter was caught by authorities having accidently trapped a protected animal.
2.3.2. Botanical Sampling
Stratified random sampling was used to sample the forest vegetation. The study area was divided into 500 × 500 m quadrants and coordinates for each plot were randomly selected with 121 plots sampled [52]. Plots were 20 × 50 m (1000 m2) and all tree species with a diameter at breast height (DBH) (1.3 m above ground) greater than 10 cm were identified with the aid of a locally trained botanist. Local names were also recorded where possible. For each tree, DBH was recorded to the nearest 0.1 cm using a diameter tape and the height was measured with a clinometer. Within the centre of each plot, a 4 × 4 m subplot was established to record the understory vegetation. Here, the stem frequencies for each plant species were counted and recorded. Name changes, authorities, and synonymy was checked using the database The Plant List [53]. Vegetation was then classified following a modified version of Sunderland et al. [54] according to the presence of key indicator plants (Table 1) and habitat characteristics were also recorded for each plot (Table 2).
2.3.3. Nest Sites
When a nest or a group of nests was observed, the area within an approximate radius of 100 m was carefully searched to locate other nests. As both chimpanzees and gorillas are known to nest in trees or on the ground, the presence of lobed versus scattered dung, odours and hairs were used to distinguish a gorilla nest from a chimpanzee nest where the sites were fresh [55]. The distance between nests was also measured, as the spacing between individual nests has also been found to be a reliable indicator for species identification [55]. A nest site was defined as a cluster of nests built concurrently, where the nearest nest is less than 50 m away [56]. Day nest sites were also distinguished from night nest sites as not every individual within a group will construct a day nest [52,57]. While structural differences are well-established in the literature, elaborately constructed day nests may be included in this sample, as the classification of day and night nests is based on indirect evidence. Nest sites unable to be identified as gorilla or chimpanzee were recorded as an ape nest site.
Nest sites were assigned approximate ages based on the visual appearance of the vegetation and the presence of dung, odours, and other signs of ape activity (Table 3) [58,59]. GPS coordinates were taken at the centre of each nest site, with the sites then classified as arboreal, ground, or mixed sites [42]. The choice of tree in which a nest is constructed can be highly selective. Previous research has found that tree selection can be influenced by the quality, flexibility and strength of a tree, the size of a tree’s leaves and their phytochemical properties, as well as the seasonal availability of the building material [60,61]. Therefore, for each arboreal nest, we recorded the height of the nest and the nesting tree, the DBH of the nesting tree, and the tree species. For ground nests, the number of plant genera used within the nest and the number of plant genera within a two-metre radius of the nest were counted [62]. Habitat characteristics were also recorded at the centre of each nest site using the same definitions as for the botanical plots (Table 2).
2.3.4. Nest Site Analysis
For each vegetation type, an importance value index (IVI) was calculated at both the family and species levels to identify the most dominant tree species and families with a DHB greater than 10 cm [5,54,63]. Descriptions of subplot species are based on the most abundant species. A high importance value indicates a tree species is well-represented due to a combination of (a) a large number of individuals of that species relative to other species within the community, or (b) a smaller number of individuals for a species that are large relative to other species within the community [63]. To determine whether vegetation differences occurred in gorilla nesting areas, we then repeated the calculation of IVIs for plots located within the immediate nesting zone. To identify statistically significant differences in floristic characteristics (e.g., stem density, tree height, tree DBH, and basal area) across vegetation classes, a nonparametric Kruskal–Wallis H test was run in SPSS 20.
To determine whether the gorillas showed a preference for particular habitat types within the study area, chi-square tests were initially run in SPSS 20. The observed values were taken from nest sites and compared to expected frequencies from the botanical plot data. Due to the low frequencies for some gorilla nesting habitat categories, some categories were collapsed or removed from the chi-square analysis. As the results were statistically significant except for understory density, a multinomial logistic regression was then run in SYSTAT 13. The dependent variables were chimpanzee sites, gorilla sites, and botanical sites that represented the total forest. This was to determine which of the examined habitat characteristics—vegetation class, elevation, degree of slope, and the amount of canopy cover—had a greater influence on the selection of nest sites.
2.3.5. Nesting Plant Preferences
Manly’s alpha was used to identify whether there were preferences for arboreal nesting tree species. This index measures resource preference based on a selection ratio comparing the proportion of a nesting species used to the proportion of available trees of that species [52]. The resulting value was then compared to a ratio of neutral preference based on the number of all species available. Neutral preference was calculated by a Manly’s alpha value of 1/m. Tree species were considered preferred if Manly’s alpha was >1/m and not preferred if values were <1/m. To calculate m, the number of tree species available for nesting, only those botanical plots within the direct nesting area of the gorillas (n = 25) were included. To determine the statistical significance of these nesting preferences, a binomial test of the difference between observed and expected numbers of trees used for nesting was then run in SPSS 20.
To determine plant preferences in ground nest construction, the proportion of each genus used within the nest and the proportion of that genus within a two-metre radius were calculated. A one-sample Wilcoxon signed rank test was then run for each plant genus to determine whether the medians differed [62]. The significance level was set at 0.05.
2.3.6. Human Forest Use
All GPS tracklogs and waypoints were imported into QGIS for analysis. Within QGIS, the measurement tool was used to calculate distances between nest sites and hunting roads, along with calculations of the total survey distances. The heatmap tool was used for both nesting sites and human signs to allow for visualisation of waypoint clusters as a means to identify high concentrations of activity (both human and ape). To determine if human activity varied throughout the study area, we compared elevations below 500 m with elevations above 500 m. Encounter rates (ERs) for each zone were calculated by dividing the number of human signs observed for each zone by the total distance surveyed in that zone, in addition to the ER within the gorilla nesting range. However, the ER is likely to underrepresent signs of human activity within the study site for several reasons. Firstly, the collection of bullet cartridges is also the procedure of WCS who conducted three surveys during the study period. Secondly, most of the trapping occurs during the height of the rainy season, and minimal surveys were conducted during this period.
2.4. Ethical Note
This research was approved by the University of Auckland Human Participants Ethics Committee (Reference 011245), and the Cameroonian Ministry of Scientific Research and Innovation (MINRESI) in collaboration with the Wildlife Conservation Society (Research Permit #65, 2014). The University of Auckland Animal Ethics Committee did not think that observational field research required ethical approval. Despite this, we strove to consider all the ethical implications of this research to form an “ethically engaged primatology” balancing the considerations given to both the human participants and the gorillas [5,9,64,65,66,67,68].
3. Results and Discussion
3.1. Characteristics of the Gorilla Nesting Environment
Cross River gorillas occupy a habitat that is somewhat intermediate between the high altitude, predominantly folivorous mountain gorillas and the frugivorous western lowland gorillas. Although they are phylogenetically and geographically most closely related to the western lowland gorilla, most Cross River gorilla groups range in ridge forests or at higher elevations (above 500 m a.s.l.) [31,35,41]. The Mone-Oku gorillas were found to nest over an area of 4.5 km2 south of Nga village, both within the Mone Forest Reserve and just outside of its borders (n = 30 nest sites; 151 nests) (Figure 2). At times, nest sites were found within 140 m from the farm–forest boundary and as close as one kilometre to the village. As a result of this proximity, there is a frequent human presence within the gorillas nesting range in the form of forest clearance, collection of nontimber forest products, and hunting. The ER reflects these levels of human activity. Within the gorilla nesting range, the ER of 1.99 signs/km was higher than that recorded across the entire study site (1.70 signs/km).
However, the gorillas appear to be reducing the chance of direct encounters with humans by concealing their night nests within the surrounding environment. An initial multinomial logistic regression of nesting habitat characteristics found vegetation class was not a significant predictor for the location of a gorilla nest site, Z = 0.638, p = 0.523, so it was removed, and the analysis was rerun. The second analysis found that elevation and degree of the slope were statistically significant predictors of gorilla nest sites, though canopy cover was not (Table 4). It is necessary to highlight that the model only accounted for a small amount of the variance (30%), likely a reflection of the small sample size. However, these results suggest the gorillas tend to select nesting locations at lowland elevations (300–450 m a.s.l.) with steep slopes. The Mone-Oku gorillas’ preference to locate nest sites on steep slopes supports the hypothesis that this is a universal behaviour among Cross River gorillas, whereby the relative inaccessibility of the slopes offers the gorillas protection from many human activities, thereby creating safe sleeping sites [37,38,39,42,54].
The lowland elevations in the gorillas’ nesting range are largely comprised of old secondary and lowland forest. This area has a rich diversity of tree species dominated by the Leguminosae family (Figure 3), but a low stand basal area (BA) corresponding to lower levels of canopy cover and an increased density of the forest understory (Table 5). The understory density was high, but variable, with the most abundant plants being herbs from the Rubiaceae, Marantaceae, and Araceae families. Although the degree of canopy cover was not found to be a significant predictor of nest sites within this sample, the high frequency of nest sites located in areas of very open forest canopy has also been observed in many populations of western gorillas [42,59,69,70]. Often, these areas of open canopy allow for the growth of preferred gorilla nesting materials [59,62,71]. Significant preferences for some plant species were observed in the construction of both ground (n = 82) (Table 6 and Table 7) and arboreal nests (n = 58) (Table 8) by the Mone-Oku gorillas. Structurally, these plants aid in the construction of a comfortable nest such as the densely packed large leaves of the herb Aframomum spp. or the combination of large leaves and drooping branches in Pycnanthus angolensis [5].
The location of nest sites may also be influenced by diet and the potential need to compete successfully with other frugivorous animals. Western lowland gorillas are known to nest close to favoured food sources rather than travel to an ideal nesting site [56,59,71]. In this study, the Mone-Oku gorillas frequently incorporated plants that are known to be a part of their diet into their nests and/or nested in areas that were surrounded by plants known to be consumed (c.f., Mawambi, Cameroon) [39,72]. Drawing from an earlier study of the Mone-Oku gorillas’ diet, the gorillas relied heavily on Palisota spp., Aframomum spp., and Cercestis camerunensis that decrease in abundance as slope and elevation increase, and species within the Marantaceae family were important fall-back foods [36]. When floristic comparisons were made across the Mone-Oku Forest, the gorilla nesting range has the highest abundance of Marantaceae species, and important food species such as P. angolensis and Guarea thompsonii are dominant tree species [31,36]. Further, fruiting trees such as Parkia bicolor are deemed an important food species at other Cross River gorilla sites [31,39] and are known to comprise part of the Mone-Oku gorillas’ diet were only recording within the gorilla nesting range. If the Mone-Oku gorillas prefer to nest close to favoured food sources, then the distribution of these plant taxa appear to be influencing the gorillas’ nesting range away from areas of a general conservation focus.
The nesting range of the Mone-Oku gorillas is reflective of their historical widespread distribution in lowland forests [73]. However, it is unusual compared to what has been observed at other Cross River gorilla sites. Typically, the Cross River gorilla is said to be found in small suboptimal refuge areas, despite the availability of suitable but unoccupied habitat throughout their range [31,38,41,74]. Alternatively, as Sawyer and Brashares suggest, previous macroscale analyses have failed to incorporate the compromises between resource abundance and potential risks [43]. The Mone-Oku gorillas appear to be currently benefiting from past anthropogenic habitat alteration that has resulted in large areas of regenerating secondary forest that is also absent of chimpanzee activity. Although the gorillas are wary of humans, the abundance of preferred nesting and food species appears to outweigh the risks posed by higher levels of human activity within their nesting range.
3.2. How Humans Have Contributed to Shaping the Current Gorilla Nesting Range
The forest landscapes we observe today have been shaped by the history of ecological, economic, and political processes [75,76,77,78]. There is now recognition that most landscapes have been altered by repeated episodes of human activity over multiple millennia and those previous ideals of “pristine” landscapes may have never existed in many instances [75,79,80,81]. Specific to West African rainforests, Connah acknowledged:
… the conviction remained that much of the rainforest consisted of dense natural vegetation that was unaffected by human settlement and which had few resources to offer. Nothing could be further from the truth and it is now realized that most of the forest has been subject to shifting agricultural exploitation at one time or another, and in some areas many times over [1] (pp. 112–113).
The archaeological record for the Cross River region is patchy. There is evidence to suggest that modern humans and nonhuman primates have coexisted in the region for at least 40,000 to 50,000 years with hunter-gatherers exploiting the rainforest and its margins around 12,000 years ago [1]. Remains from Cross River gorillas, Nigeria–Cameroon chimpanzees and drills have been dated to around 5000 BP from the Shum Laka rock shelter near Bamenda [82]. Here, primates comprised 14% of the total faunal assemblage illustrating that people were hunting in heavily wooded areas through to the late Holocene [82,83]. The Western Bantu expansion, believed to have begun with the contraction of the rainforests around the Nigeria–Cameroon borderland, has received more attention across Central Africa [84,85]. Coinciding with the expansion is evidence for more sedentary ways of life with slash-and-burn agriculture for the cultivation of yams and oil palms and iron smelting [1,84,85]. However, interpretations of anthropogenic versus ecological dynamics are confounded by the lack of ecological records focusing on the scope and scale of impacts associated with prehistoric agricultural practices [81].
The oral histories of Nga village did not extend back far enough to provide detail of how people lived in the forest in the distant past. People recount their ancestors living in the forest and moving from place to place during times of warfare, with people coming together to establish the village more recently. Röschenthaler [86] found that in the precolonial era the Cross River people were highly mobile, moving to new areas due to fighting (including slave-trading), water shortages, depleted soils, or to increase their economic prospects closer to trade routes that were “diffuse and multidirectional” [87] (p. 64) [88]. Similar movements involving forest clearance for slash-and-burn agriculture and the subsequent abandonment of farms as people migrate was found to have had a positive influence on forest ecosystems across Central Africa. This is due to the fact that these practices create gaps in the forest canopy suitable for the germination of light-demanding trees such as Lophira alata (Redwood), which are widely sought after in the current timber industry [89,90,91]. From the 1850s onwards, coinciding with colonial regimes and the widespread forced movement of people out of forests and closer to rivers and roads, there is a noticeable reduction in the regeneration of these tree species believed to be a result of reduced human disturbance in the forest [89,90,91]. These light-demanding timber species were also identified within the Mone-Oku Forest with several species used by the gorillas [5]. Diospyros spp. (ebony) is perhaps the most significant genera of these trees as it is both a food and nesting source for the gorillas. While Milicia excelsa (African teak or Iroko) and Pterocarpus soyauxii (African padauck) offer a wide range of food resources, the Cross River gorilla is known to feed on its fruits, flowers, seeds, leaves, and bark [31,36,39]. These results suggest that a degree of disturbance greater than what occurs naturally is required to maintain certain habitats and tree species within the rainforest. They also demonstrate that a degree of human disturbance within the forest can have positive benefits for the Cross River gorilla, promoting the growth of some tree species used for nesting and feeding.
Cameroon’s Cross River region has a colonial history of everchanging boundaries resulting in “developmental” neglect and isolation, which has played an important role in shaping people’s reliance on the Mone-Oku Forest [5]. It is from this history and the “reopening” of the Akwaya region, both physically and metaphorically, that we see a clear example of the social nature of the forest landscape [78], and its impact on the nesting range of the Mone-Oku gorillas. Before the 1980s, the people of Nga lived at one site, approximately one kilometre southeast of Nga’s current location. The village is believed to have been established between 100 and 300 years ago but is likely to have emerged as a result of the German administration’s forced amalgamation policies following the 1904 Mpawmanku War [87]. By 1924, British ethnographic surveys reveal Nga as a small village (ca. 77 adults and children) involved in farming and collecting oil palm kernels (Elaeis guineensis) to supply the African and Eastern Trade Corporation based in Mamfe [92]. Sharwood-Smith and Cantle reported on farming within the region:
As everywhere the bulk of agricultural work is done by the women. The men clear new bush and set their Plantains after which they do little else. The area is cultivated by one woman, including her husband’s plantain patch, is approximately one acre, of which two fifths is plantains, two fifths is cocoyams, and one fifth luxury crops (corn, beans, pepper, groundnuts, pumpkins and gourds or tobacco) [92] (p. 40).
The elder men of Nga support these views of past farming practices. They said the first farms were small, no larger than one hectare, and close to the village. This was due to the fact that people were only planting what they needed for food and a small income, such as plantains, cocoyams, kola nuts, and oil palm, which self-germinated.
Although mostly described as farmers, the Nga people also hunted for subsistence throughout the Mone-Oku Forest, using a combination of trapping pits, snares, and den guns where available. Information about the historical hunting of the gorillas mostly came from the elder men within Nga. They explained that in the past, people did not fear the repercussions of hunting the apes, but this was also tempered by the role gorillas played as totems within their traditional practices [5]. Hunters killed the gorillas to feed their families, for medicine, and to gain prestige. One elder animatedly said he was a great hunter in his time, but hunting was mostly for food as there was no market to sell the meat. He recalled:
Ku [gorilla] sleep for ground, broken sticks put them for ground. As you get close, they call and you stay quiet. I kill gorilla when eating chop [food], then carried them to use for country juju [medicine]. When die, put Ku for seat and tie up place where shot … After tied to chair, carry Ku to that back place where women would dance and dance before butchering.
The celebrations that occurred after the death of a gorilla were due to the gorillas’ designation as “village beef”; therefore, they are for all to share and could not be sold even if a market were available. Accounts indicate that before the 1920s hunting of the gorillas at Nga was limited with one killed approximately every four to five years [5].
As Nga’s population grew (est. 321 people in 1967) [93], it became increasingly difficult to locate suitable sites to construct new homes as the old village site was stony. Ojong Solomon remembered, “… The stones were plenty and some people died falling from the stones, so they decided to move further down to Tito”. The site of Tito appealed for several reasons; the first was that it allowed people to be closer to the proposed road and thereby closer to “development”. One Mammi from Nga recalls they moved “because they were too far in the bush, wanted to move closer to the road. I decided to stay as I have all bush, all things for here”. The second reason for the move was political, to ensure the people of neighbouring villages, Akwa and Bantaco, would no longer encroach on their forest to hunt or clear farmland. Establishing the Tito site appeared fraught with difficulty as one elder described:
… we came to here to clear black bush [primary forest], burn all the bush. For night elephant comes for church house, tigers [leopards] came, carry goats to bush, I was the greatest hunter, I go shoot dem tigers through the door with dem powder guns. Corn was fine for here, as was mushroom from trees that had been felled. That time no palm, so we went bush to cut those tall ones, aye, we suffered those times… Some go back, as said the Tito place is bad. People shooting others with guns, me shot here [ribs] sutee!! After death where one shot, they scattered. Some decided for here and some for up.
While buffalos, elephants, and chimpanzees were remembered as causing a lot of damage as Tito was established, but they are no longer seen to be an issue. One farmer said that he “sees with the opening of farms, animals are going further into the bush. Animals reduced a bit due to the increase of population and hunters—elephants, buffalo, water beef [chevrotain], sleeping deer [Bay duiker], giant pangolin”. The impact the move and resulting forest clearance around the Tito site had on the gorillas remained curiously absent from our discussions. AW’s roles as a researcher, a “white man”, “Wildlife”, a New Zealander, a woman, and her interests in the welfare of both the human participants and the apes influenced participants’ perceptions and the information they choose to provide. In the village, AW was of unusual standing and did not fit into the typical preconceived categories of village life. She was unable to disassociate herself from being perceived as “Wildlife” and part of this perception was likely to have arisen from the collaboration with, and introduction, from the WCS. These perceptions are likely to have prevented discussion around topics such as bushmeat hunting with some participants. As AW gained rapport with several key informants, she was able to have discussions about any bushmeat they had collected and what they had seen in the forest throughout their travels. Official reports state the last Mone-Oku gorilla hunted was in 1996, an approximately 20-year reprieve at the time of the research [94]. Tito has since become a well-established site for the majority of the Nga people. Farms are now mature and contain valued nontimber forest products (e.g., njansan, bush mango) that were transplanted to reduce the need to travel into the forest. Although, large tracks of lowland forest immediately surrounding Tito are also being cleared as people seek to capitalise on the cacao trade (see below).
The majority of the original Nga farms are now regenerating secondary forest that are favoured nesting and feeding environments for Mone-Oku gorillas. Although this human habitat alteration is unlikely to result in a long-term abundance of herbaceous vegetation [95], it will increase the germination frequency of light-demanding tree species, including those preferred by the gorillas [89,90,91]. Similarly, shifting settlement systems in Mali have been demonstrated to modify habitats in such a way that they are both attractive and available to chimpanzees, supplementing natural habitats both spatially and temporally [76]. People’s recollections about the move from old Nga to Tito also highlights our lack of understanding about community ecology in the region. For example, how do gorillas interact with other mammals and what are the effects of feeding competition on their ranging behaviour? The Mone-Oku gorillas and chimpanzees have distinct nesting ranges and appear to occupy different habitats as a likely response to feeding competition [5]. However, has the decrease in forest buffalo and elephants in the Mone-Oku Forest due to human hunting reduced feeding competition, further allowing the gorillas to occupy open areas with greater herb and fruit availability? Limited studies on forest buffalo suggest there is dietary overlap in both staple and fallback herb species that may affect the ranging behaviour of both taxa, especially in times of low fruit availability [31,36,96,97]. In Gabon, elephants were found to competitively exclude western lowland gorillas in times of low herb availability, irrespective of fruit abundance, where both taxa were competing for fruit [98]. The large areas of “unoccupied” but suitable Cross River gorilla habitats may be a result of competitive exclusion in areas of sympatry and remain in need of further investigation.
Human land-use patterns and hunting pressures have played a significant role in shaping the wider Cross River landscape. Still, there has been a bias towards focusing on negative anthropogenic interactions (e.g., hunting and habitat loss) without consideration given to how anthropogenic habitat alteration has shaped the forest over time with the potential to create favourable new microhabitats even if they are temporary. The focus on the anthropogenic in Cross River gorilla conservation means that interspecies interactions and the effects of competition on the gorillas’ ranging and feeding behaviours have been neglected.
3.3. Cacao and Wider Connections
The villages around the Mone Forest Reserve, including Nga, have been labelled as threats to the conservation of the Cross River gorillas due to the recent rapid expansion of their cacao farms [30]. However, the situation is complex. Through the reconfirmed presence of the Cross River gorilla during the early 1990s, Akwaya Subdivision became repopulated in the eyes of the international scientific community [35,54,99,100,101,102]. At the same time, Akwaya was also establishing a network of connections to the remainder of Cameroon through roading projects and economic developments. This has resulted in a landscape where local practices have been shaped by external forces but only the local outcomes are realised as key threats to the conservation of the Cross River gorilla.
The lack of access to Akwaya Subdivision became highlighted in the 1960s when the Southern Cameroons voted to be reunited with the Republic of Cameroon. Upon independence, Mamfe (approx. 48 km from Nga) became the main commercial and administrative town for the region, yet the subdivision could only be reached via foot (approximately 2–3 days walk) within Cameroon. Early attempts to construct a road were made by the Western Cameroonian Ministry in the late 1960s, but the project was abandoned when the then Federal Government refused to contribute funds [101]. It was the belief that this road was going to be constructed that played an instrumental role in the people of Nga’s decision to relocate to the Tito site. It was not until 1991 that the first motorable bridge was constructed across the Cross River by a Lebanese timber company which received a logging concession in the region [102]. This bridge and the seasonal earth road that slowly followed began to fully connect the subdivision to Mamfe and Cameroon. As the road was established many sought to take advantage of the benefits that arose from increased market access with increases in hunting, harvesting of nontimber forest products, and forest conversion to farmland recorded [101]. Contributing to the forest clearance was a series of political and economic changes that turned cacao into a profitable cash crop.
Cacao has a long history within Cameroon. The German Administration introduced it to the Southwest in 1886 and rapidly established plantations that resulted in cacao becoming the main agricultural export by 1913 [103,104], but cacao only became an important cash crop within Nga village and the wider subdivision in the 1990s. There appear to be two key reasons for the shift to farming cacao. The first relates to the dangerous nature of harvesting oil palm kernels, the main cash crop prior to this period. As one elder from Nga commented, “it was dangerous to go up the trees to find money. However, now cocoa is a new thing and it is safer because it is on the ground”. The second reason given behind the shift to cacao was the increase in the purchase price that arose from the severe economic crisis Cameroon suffered in the early 1990s. This crisis was partly a result of the falling prices of raw materials on the international market that saw Cameroon reform local markets following the implementation of structural adjustment loans from the World Bank [105]. While international cacao prices gradually increased across the 1990s, Cameroon’s economic reforms removed the fixed purchase price, opening the competitive world markets to Cameroonian farmers [103,106].
Cacao has since become an integral crop to the people of Nga and it is quickly becoming interwoven into the fabric of society. The size of a man’s cacao farm appears to dictate his standing among his peers, with those who dedicate their time and energy into their farm accumulating great wealth and power within the community. In contrast, there was a stigma attached to the few men who did not have a cacao farm and often struggled to provide for their family and perhaps, more importantly, their children’s education. Most men within the village have cleared forest specifically for cacao farms, with many expressing a desire to increase their farmland. As Tuku wished: “If I was feeling strong, I would clear as I go and never look back”. Despite these desires, the village has certain rules surrounding forest clearance that practically limits the size of most farms to two or three hectares as a farmer must be able to maintain the land that is cleared.
However, the desires to increase cacao production are not only local. In 2009, President Bia announced his intention to transform Cameroon from a developing to an emerging nation by 2035. As part of the 2035 emergence drive, the Cameroon Government sought to capitalize on the expected increase in demand for cacao and triple its cacao production by 2020 (an increase of 600,000 tonnes per year), to become one of the world’s top three producers [107]. To facilitate these goals, President Bia announced at Mamfe in 2015 that the Akwaya Road would be tarred, finally fully connecting Akwaya to the rest of Cameroon, although the project has not commenced in 2021 [108].
The combination of local, political, and international economic processes favouring cacao farming within Akwaya certainly has the potential to cause widespread deforestation. With limited access to education on or support for farming cacao, the general practice within Akwaya is to clear the nonuseful trees within a farms’ boundaries, as this allows more sunlight and greater harvests. Nutrient absorbing food crops (e.g., cassava and cocoyams) are also grown with cacao increasing the potential for soil depletion and barren land [109]. However, it is not necessary to grow cacao this way. Across Africa and Brazil are long-established complex agroforests where cacao is cultivated with a mixture of fruit and forest tree species [110,111,112,113,114]. These agroforests have demonstrated greater long-term viability of cacao farms while retaining the soil nutrients, allowing for the potential regeneration of the forest should the farms be abandoned for social or economic reasons.
The rapid forest clearance in Indonesia for oil palm plantations and the precarious plight of orangutans demonstrates how powerful world market forces can be in small locales [115]. With cacao production playing a large role in Nga, initiatives seeking to conserve the Cross River gorilla are likely to gain greater community support if they can find mutually beneficial ways to support cacao farming. Positive relationships and guidance on making cacao farms more productive and long-lasting has the potential to preserve negotiated forest corridors throughout the gorillas’ range. Ultimately, how far cacao farms expand throughout the Mone-Oku Forest and the wider region depends as much on the development of local infrastructure and global markets as it does on the local farmers.
It is also necessary to remember that the situation is not static. Since the completion of this fieldwork in 2015, Cameroon has experienced internal hostilities between the Anglophone regions and the largely Francophone Government that appear to be once again reshaping people’s relationships with the forest and the animals within. Since 2016, it is estimated that over 2000 people have died and over half a million people are displaced, with both the Cameroon Government and separatist forces committing serious abuses against the civilian population [116]. Large numbers of people are taking refuge in the forests along the Nigeria–Cameroon border as villages are set alight, with many reporting to be living off what they can hunt and gather in the forest [117]. In January 2021, there remains little intervention from the international community or negotiation between the Anglophone and Francophone factions. The situation may be further compounded as Cameroon experiences its second wave of the COVID-19 virus, with humanitarian organisations struggling to gain access to the area to provide healthcare and education [118]. The fate of the people, wildlife, and forests across Southwest Cameroon remains unknown; however, lessons are available from regions that have experienced previous armed conflicts. Where wildlife populations have declined during a period of armed conflict, population recovery has been possible when food insecurity and supply chains are addressed at local levels and support is provided to local and governmental institutions that manage conservation-related activities [119,120,121].
4. Conclusions
Cross River gorilla conservation has largely focused on creating and managing protected spaces for these critically endangered apes, with some calling for these protected spaces to increase [30,74]. This research illustrates the ways in which an ethnoprimatological approach in a defined place can enhance our understanding of the interactions that exist between primates and people. Understanding how primates may change their behaviour in anthropogenic environments can provide valuable insights for the conservation of a species and further understanding of great ape cognition and evolution [122]. To contribute to this understanding, McLennan et al. [123] highlight the importance of comparisons among populations exposed to varying forms and degrees of anthropogenic influence.
The typically unusual ranging behaviour of the Mone-Oku gorillas, with preferences for lowland areas in proximity to humans, illustrates their capacity for behavioural flexibility while also highlighting the complexities of anthropogenic activities within a landscape. The Mone-Oku gorillas avoid nesting in areas with the highest density of human activity and use characteristics of the nesting environment to conceal their nest sites. Within this nesting environment, significant preferences towards a particular set of plant species were also observed in the construction of both ground and arboreal nests, suggesting the availability of these preferred species also influences nest site selection. In addition, many of the plants observed within the nesting environment are also known food sources for the Mone-Oku population and these correlations are worthy of future investigation. These preferred nesting and feeding plant species are useful indicators of suitable gorilla habitats that have the potential to aid conversations surrounding land management strategies for future conservation plans for the Cross River gorilla.
The Mone-Oku Forest has a long history of alterations, some of which are human-induced. These alterations continue to the present day, where political history has shaped limited livelihood alternatives, thereby increasing the reliance on the forest that has remained a constant. Past village movements, as illustrated by the move to the Tito site, have the potential to positively influence gorilla ranging patterns through the regeneration of secondary forest and may also inadvertently reduce feeding competition with previously sympatric mammals. This research also questions the assumption in traditional conservation narratives that the main threats to species survival are at the local level. When analysis is applied at different scales, such as the nation-state or international markets, the impact of wider political and economic processes on regions perceived as marginal is illustrated.
Author Contributions
Conceptualisation, A.H.W.; methodology, A.H.W. and N.M.; formal analysis, A.H.W.; investigation, A.H.W.; resources, A.H.W. and N.M.; data curation, A.H.W.; writing—original draft preparation, A.H.W.; writing—review and editing, A.H.W. and N.M.; visualisation, A.H.W.; supervision, N.M.; project administration, A.H.W.; funding acquisition, A.H.W. and N.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by the American Society of Primatologists Conservation Small Grant, the University of Auckland Arts Doctoral Research Fund, and the University of Auckland Doctoral Scholarship.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Human Participants Ethics Committee of The University of Auckland (Reference 011245, 13 March 2014).
Informed Consent Statement
Informed consent was obtained from all participants involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We are grateful to Nga for welcoming A.H.W. into their village and sharing their lives with her. To A.H.W.’s field team, Okon Felix, Nkeh Michael Obi, Atina William, Nkunyu Paius, Tiger Wood and Takie Simon Ndo, thank you for helping to make this project possible. The Wildlife Conservation Society and their staff at the Takamanda-Mone Landscape Project provided invaluable friendships, logistic support, and introductions. To Judith Littleton and Bruce Floyd, thank you for your patience, support, and invaluable feedback throughout this process. We would also like to thank the two reviewers for their constructive comments and insight.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Connah, G. Forgotten Africa: An Introduction to Its Archaeology; Taylor and Francis: Oxfordshire, UK, 2004. [Google Scholar]
- Oates, J.F. Workshop on the conservation of Cross River Gorillas. Gorilla J. 2001, 22, 26–27. [Google Scholar]
- Sarmiento, E.E.; Oates, J.F. The Cross River gorillas: A distinct subspecies Gorilla gorilla diehli Matschie 1904. Am. Mus. Novit. 2000, 3304, 1–55. [Google Scholar] [CrossRef] [Green Version]
- Gorilla gorilla ssp. diehli. The IUCN Red List of Threatened Species 2016. Available online: https://www.iucnredlist.org/species/39998/102326240 (accessed on 3 January 2020).
- Wade, A.H. Shared Landscapes: The Human-Ape Interface within the Mone-Oku Forest, Cameroon. Ph.D. Thesis, The University of Auckland, Auckland, New Zealand, 2020. [Google Scholar]
- Setchell, J.M.; Fairet, E.; Shutt, K.; Waters, S.; Bell, S. Biosocial conservation: Integrating biological and ethnographic methods to study human–primate interactions. Int. J. Primatol. 2017, 38, 401–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dore, K.M. Ethnoprimatology without conservation: The political ecology of farmer–green monkey (Chlorocebus sabaeus) relations in St. Kitts, West Indies. Int. J. Primatol. 2018, 39, 918–944. [Google Scholar] [CrossRef]
- Riley, E.P.; Fuentes, A.; Dore, K.M. Introduction: Doing Ethnoprimatology in the Anthropocene. In Ethnoprimatology: A Practical Guide to Research at the Human-Nonhuman Primate Interface; Dore, K.M., Riley, E.P., Fuentes, A., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 1–6. [Google Scholar]
- Malone, N.; Palmer, A.; Wade, A.H. Incorporating the ethnographic perspective: The value, process, and responsibility of working with human participants. In Ethnoprimatology: A Practical Guide to Research at the Human-Nonhuman Primate Interface; Dore, K.M., Riley, E.P., Fuentes, A., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 176–189. [Google Scholar]
- Parathian, H.E.; McLennan, M.R.; Hill, C.M.; Frazão-Moreira, A.; Hockings, K.J. Breaking through disciplinary barriers: Human–wildlife interactions and multispecies ethnography. Int. J. Primatol. 2018, 39, 749–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, E.P. The Maturation of ethnoprimatology: Theoretical and methodological pluralism. Int. J. Primatol. 2018, 39, 705–729. [Google Scholar] [CrossRef]
- Riley, E.P.; Ellwanger, A.L. Methods in ethnoprimatology: Exploring the human-nonhuman primate interface. In Primate Ecology and Conservation: A Handbook of Techniques; Sterling, E., Bynum, N., Blair, M., Eds.; Oxford University Press: Oxford, UK, 2013; pp. 128–150. [Google Scholar]
- Ellwanger, A.L.; Lambert, J.E. Investigating niche construction in dynamic human-animal landscapes: Bridging ecological and evolutionary timescales. Int. J. Primatol. 2018, 39, 797–816. [Google Scholar] [CrossRef]
- Jost Robinson, C.A.; Remis, M.J. Engaging holism: Exploring multispecies approaches in ethnoprimatology. Int. J. Primatol. 2018, 39, 776–796. [Google Scholar] [CrossRef]
- Malone, N.; Wade, A.H.; Fuentes, A.; Riley, E.P.; Remis, M.; Robinson, C.J. Ethnoprimatology: Critical interdisciplinarity and multispecies approaches in anthropology. Crit. Anthropol. 2014, 34, 8–29. [Google Scholar] [CrossRef]
- Escobar, A. Whose knowledge, whose nature? Biodiversity, conservation, and the political ecology of social movements. J. Political Ecol. 1998, 5, 53–82. [Google Scholar] [CrossRef]
- Sousa, J.; Hill, C.M.; Ainslie, A. Chimpanzees, sorcery and contestation in a protected area in Guinea-Bissau. Soc. Anthropol. 2017, 25, 364–379. [Google Scholar] [CrossRef]
- Escobar, A. After nature: Steps to an antiessentialist political ecology. Curr. Anthropol. 1999, 40, 1–30. [Google Scholar] [CrossRef]
- Blaikie, P.; Brookfield, H. Land Degradation and Society; Methuen: London, UK, 1987. [Google Scholar]
- Forsyth, T. Critical Political Ecology: The Politics of Environmental Science; Routledge: London, UK, 2004. [Google Scholar]
- Nygren, A.; Rikoon, S. Political ecology revisited: Integration of politics and ecology does matter. Soc. Nat. Resour. 2008, 21, 767–782. [Google Scholar] [CrossRef]
- Stott, P.; Sullivan, S. Political Ecology: Science, Myth and Power; Arnold Publishers: London, UK, 2000. [Google Scholar]
- Wolf, E. Ownership and political ecology. Anthropol. Q. 1972, 45, 201–205. [Google Scholar] [CrossRef]
- Fairhead, J.; Leach, M. Reframing Deforestation: Global Analyses and Local Realities: Studies in West Africa; Routledge: London, UK, 1998. [Google Scholar]
- Jones, S. A political ecology of wildlife conservation in Africa. Rev. Afr. Political Econ. 2006, 33, 483–495. [Google Scholar] [CrossRef]
- Mulder, M.B.; Coppolillo, P. Conservation: Linking Ecology, Economics, and Culture; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- Neumann, R.P. Making Political Ecology; Oxford University Press: New York, NY, USA, 2005. [Google Scholar]
- Nelson, J.G.; Serafin, R. Assessing biodiversity: A human ecological approach. Ambio 1992, 21, 212–218. [Google Scholar]
- Gezon, L.L. Global Visions, Local Landscapes: A Political Ecology of Conservation, Conflict, and Control in Northern Madagascar; Altamira Press: Lanham, MA, USA, 2006. [Google Scholar]
- Dunn, A.; Bergl, R.; Byler, D.; Eben-Ebai, S.; Etiendem, D.N.; Fotso, R.; Ikfuingei, R.; Imong, I.; Jameson, C.; Macfie, E.J.; et al. Revised Regional Action Plan for the Conservation of the Cross River Gorilla (Gorilla gorilla diehli) 2014–2019; IUCN/SSC Primate Specialist Group and Wildlife Conservation Society: New York, NY, USA, 2014. [Google Scholar]
- McFarland, K.L. Ecology of Cross River gorillas (Gorilla gorilla diehli) on Afi Mountain, Cross River State, Nigeria. Ph.D. Thesis, City University of New York, New York, NY, USA, 2007. [Google Scholar]
- Oates, J.F.; Bergl, R.A.; Linder, J.M. Africa’s Gulf of Guinea Forests: Biodiversity Patterns and Conservation Priorities; Conservation International Center for Applied Biodiversity Science: Washington, DC, USA, 2004. [Google Scholar]
- Bergl, R.A.; Vigilant, L. Genetic analysis reveals population structure and recent migration within the highly fragmented range of the Cross River gorilla (Gorilla gorilla diehli). Mol. Ecol. 2007, 16, 501–516. [Google Scholar] [CrossRef]
- Bergl, R.A.; Bradley, B.J.; Nsubuga, A.; Vigilant, L. Effects of habitat fragmentation, population size and demographic history on genetic diversity: The Cross River gorilla in a comparative context. Int. J. Primatol. 2008, 70, 848–859. [Google Scholar] [CrossRef]
- Oates, J.F.; McFarland, K.L.; Groves, J.L.; Bergl, R.A.; Linder, J.M.; Disotell, T.R. The Cross River gorilla: The natural history and status of a neglected and critically endangered subspecies. In Gorilla Biology: A Multidisciplinary Perspective; Taylor, A., Goldsmith, M., Eds.; Cambridge University Press: Cambridge, UK, 2003; pp. 472–497. [Google Scholar]
- Sawyer, S.C. The Ecology and Conservation of the Critically Endangered Cross River Gorilla in Cameroon. Ph.D. Thesis, University of California, Oakland, CA, USA, 2012. [Google Scholar]
- Sunderland-Groves, J.L.; Ekinde, A.; Mboh, H. Cross River gorilla (Gorilla gorilla diehli) nesting behaviour at Kagwene Mountain, Cameroon: Implications for assessing group size and density. Int. J. Primatol. 2009, 30, 253–266. [Google Scholar] [CrossRef]
- Imong, I.; Robbins, M.M.; Mundry, R.; Bergl, R.; Kühl, H.S. Distinguishing ecological constraints from human activity in species range fragmentation: The case of Cross River gorillas. Anim. Conserv. 2014, 17, 323–331. [Google Scholar] [CrossRef]
- Etiendem, D.N. Ecological and Anthropogenic Factors of Cross River Gorilla (Gorilla gorilla diehli) Survival at Mawambi Hills, Southwest Cameroon. Ph.D. Thesis, Vrije Universiteit, Brussels, Belgium, 2013. [Google Scholar]
- Etiendem, D.N.; Tagg, N. Feeding ecology of Cross River gorillas (Gorilla gorilla diehli) at Mawambi Hills: The influence of resource seasonality. Int. J. Primatol. 2013, 34, 1261–1280. [Google Scholar] [CrossRef]
- Etiendem, D.N.; Funwi-Gabga, N.; Tagg, N.; Hens, L.; Indah, E.K. The Cross River gorillas (Gorilla gorilla diehli) at Mawambi Hills, south-west Cameroon: Habitat suitability and vulnerability to anthropogenic disturbance. Folia Primatol. 2013, 84, 18–31. [Google Scholar] [CrossRef]
- De Vere, R.A.; Warren, Y.; Nicholas, A.; Mackenzie, M.E.; Higham, J.P. Nest site ecology of the Cross River gorilla at the Kagwene Gorilla Sanctuary, Cameroon, with special reference to anthropogenic influence. Am. J. Primatol. 2011, 73, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, S.C.; Brashares, J.S. Applying resource selection functions at multiple scales to prioritize habitat use by the endangered Cross River gorilla. Divers. Distrib. 2013, 19, 943–954. [Google Scholar] [CrossRef]
- White, F. The Vegetation of Africa, A Descriptive Memoir to Accompany the UNESCO/AETFAT/UNSO Vegetation Map of Africa (3 Plates, Northwestern African, Northeastern Africa, and Southern Africa, 1:5,000,000); UNESCO: Paris, France, 1983. [Google Scholar]
- Mboh, H.; Warren, Y. Large Mammal Survey of the Proposed Takamanda National Park; Unpublished Report; Wildlife Conservation Society: Limbe, Cameroon, 2007. [Google Scholar]
- Jameson, C. Gorilla Guardian update: Expansion of the community-based monitoring network. Gorilla J. 2012, 45, 13–15. [Google Scholar]
- Blommaert, J.; Jie, D. Ethnographic Fieldwork: A Beginner’s Guide; Multilingual Matters: Bristol, UK, 2010. [Google Scholar]
- Dewalt, K.M.; Dewalt, B.R. Participant Observation: A Guide for Fieldworkers, 1st ed.; AltaMira Press: Walnut Creek, CA, USA, 2002. [Google Scholar]
- Spradley, J.P. Participant Observation; Holt, Rinehart and Winston: New York, NY, USA, 1980. [Google Scholar]
- Davies, C.A. Reflexive Ethnography: A Guide to Researching Selves and Others; Routledge: London, UK, 1999. [Google Scholar]
- Kühl, H.; Maisels, F.; Ancrenaz, M.; Williamson, E.A. Best Practice Guidelines for Surveys and Monitoring of Great Ape Populations; IUCN SSC Primate Specialist Group: Gland, Switzerland, 2008. [Google Scholar]
- Brownlow, A.R.; Plumptre, A.J.; Reynolds, V.; Ward, R. Sources of variation in the nesting behavior of chimpanzees (Pan troglodytes schweinfurthii) in the Budongo Forest, Uganda. Am. J. Primatol. 2001, 55, 49–55. [Google Scholar] [CrossRef]
- The Plant List: A Working List of all Plant Species. Version 1.1. 2013. Available online: http://www.theplantlist.org/ (accessed on 14 July 2015).
- Sunderland-Groves, J.L.; Maisels, F.; Ekinde, A. Surveys of the Cross River Gorilla and chimpanzee populations in Takamanda Forest Reserve, Cameroon. In Takamanda—The Biodiversity of an African Rainforest; Comiskey, J.A., Sunderland, T.C.H., Sunderland-Groves, J.L., Eds.; Smithsonian Institution: Washington, DC, USA, 2003; pp. 129–140. [Google Scholar]
- White, L.J.T.; Edwards, A. Conservation Research in the African Rain Forest: A Technical Handbook; Wildlife Conservation Society: New York, NY, USA, 2000. [Google Scholar]
- Fruth, B.; Hohmann, G. Nest building behavior in the great apes: The great leap forward? In Great Ape Societies; McGrew, W.C., Merchant, L.F., Nishida, T., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 225–240. [Google Scholar]
- Stewart, F.A.; Pruetz, J.D.; Hansell, M.H. Do chimpanzees build comfortable nests? Am. J. Primatol. 2007, 69, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Koops, K.; McGrew, W.C.; de Vries, H.; Matsuzawa, T. Nest-building by chimpanzees (Pan troglodytes verus) at Seringbara, Nimba Mountains: Antipredation, thermoregulation, and antivector hypotheses. Int. J. Primatol. 2012, 33, 356–380. [Google Scholar] [CrossRef]
- Tutin, C.E.; Parnell, R.J.; White, L.J.; Fernandez, M. Nest building by lowland gorillas in the Lopé Reserve, Gabon: Environmental influences and implications for censusing. Int. J. Primatol. 1995, 16, 53–76. [Google Scholar] [CrossRef]
- Samson, D.R.; Hunt, K.D. A thermodynamic comparison of arboreal and terrestrial sleeping sites for dry-habitat chimpanzees (Pan troglodytes schweinfurthii) at the Toro-Semliki Wildlife Reserve, Uganda. Am. J. Primatol. 2012, 74, 811–818. [Google Scholar] [CrossRef]
- Van Casteren, A.; Sellers, W.I.; Thorpe, S.K.; Coward, S.; Crompton, R.H.; Myatt, J.P.; Ennos, A.R. Nest-building orangutans demonstrate engineering know-how to produce safe, comfortable beds. Proc. Natl. Acad. Sci. USA 2012, 109, 6873–6877. [Google Scholar] [CrossRef] [Green Version]
- Rothman, J.M.; Pell, A.N.; Dierenfeld, E.S.; Mccann, C.M. Plant choice in the construction of night nests by gorillas in the Bwindi Impenetrable National Park, Uganda. Am. J. Primatol. 2006, 68, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley & Sons: New York, NY, USA, 1974. [Google Scholar]
- Fedigan, L.M. Ethical issues faced by field primatologists: Asking the relevant questions. Am. J. Primatol. 2010, 72, 754–771. [Google Scholar] [CrossRef] [PubMed]
- Bezanson, M.; McNamara, A. The what and where of primate field research may be failing primate conservation. Evol. Anthropol. Issues News Rev. 2019, 28, 166–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacKinnon, K.C.; Riley, E.P. Contemporary ethical issues in field primatology. In Ethics in the Field: Contemporary Challenges; MacClancy, J., Fuentes, A., Eds.; Berghahn Books: New York, NY, USA, 2013; pp. 98–107. [Google Scholar]
- Riley, E.P.; Bezanson, M. Ethics of primate fieldwork: Toward an ethically engaged primatology. Annu. Rev. Anthropol. 2018, 47, 493–512. [Google Scholar] [CrossRef]
- Strier, K.B. Are observational field studies of wild primates really noninvasive? In Ethics in the Field: Contemporary Challenges; MacClancy, J., Fuentes, A., Eds.; Berghahn Books: New York, NY, USA, 2013; pp. 67–83. [Google Scholar]
- Mehlman, P.T.; Doran, D.M. Influencing western gorilla nest construction at Mondika Research Center. Int. J. Primatol. 2002, 23, 1257–1285. [Google Scholar] [CrossRef]
- Remis, M. The positional behavior of lowland gorillas (Gorilla gorilla gorilla) in the Dzanga-Sangha Reserve, Central African Republic. Am. J. Primatol. 1993, 30, 344. [Google Scholar]
- Willie, J.; Petre, C.A.; Tagg, N.; Lens, L. Density of herbaceous plants and distribution of western gorillas in different habitat types in south-east Cameroon. Afr. J. Ecol. 2013, 51, 111–121. [Google Scholar] [CrossRef]
- Etiendem, D.N. Tree nesting in Mawambi Hills gorillas. Gorilla J. 2013, 46, 10–12. [Google Scholar]
- Bergl, R.A.; Warren, Y.; Nicholas, A.; Dunn, A.; Imong, I.; Sunderland-Groves, J.L.; Oates, J.F. Remote sensing analysis reveals habitat, dispersal corridors and expanded distribution for the Critically Endangered Cross River gorilla Gorilla gorilla diehli. Oryx 2012, 46, 278–289. [Google Scholar] [CrossRef] [Green Version]
- Imong, I.; Robbins, M.M.; Mundry, R.; Bergl, R.; Kühl, H.S. Informing conservation management about structural versus functional connectivity: A case study of Cross River gorillas. Am. J. Primatol. 2014, 76, 978–988. [Google Scholar] [CrossRef]
- Boivin, N.L.; Zeder, M.A.; Fuller, D.Q.; Crowther, A.; Larson, G.; Erlandson, J.M.; Denham, T.; Petraglia, M.D. Ecological consequences of human niche construction: Examining long-term anthropogenic shaping of global species distributions. Proc. Natl. Acad. Sci. USA 2016, 113, 6388–6396. [Google Scholar] [CrossRef] [Green Version]
- Duvall, C.S. Human settlement ecology and chimpanzee habitat selection in Mali. Landsc. Ecol. 2008, 23, 699–716. [Google Scholar] [CrossRef]
- Malone, N.; Selby, M.; Longo, S. Political-ecological dimensions of silvery gibbon conservation efforts: An endangered ape in (and on) the verge. Int. J. Sociol. 2014, 44, 34–53. [Google Scholar] [CrossRef]
- Tsing, A.L. Friction; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- Ellis, E.C.; Kaplan, J.O.; Fuller, D.Q.; Vavrus, S.; Goldewijk, K.K.; Verburg, P.H. Used planet: A global history. Proc. Natl. Acad. Sci. USA 2013, 110, 7978–7985. [Google Scholar] [CrossRef] [Green Version]
- Grayson, D.K. The archaeological record of human impacts on animal populations. J. World Prehist. 2001, 15, 1–68. [Google Scholar] [CrossRef]
- Wright, D.K.; MacEachern, S.; Ambrose, S.H.; Choi, J.; Choi, J.-H.; Lang, C.; Wang, H. Iron Age landscape changes in the Benoué River Valley, Cameroon. Quat. Res. 2019, 92, 323–339. [Google Scholar] [CrossRef]
- De Maret, P.; Clist, B.; Van Neer, W. Résultats des premiéres fouilles dans les abris de Shum Laka et d’Abeke au Nord-Ouest du Cameroun. L’anthropologie 1987, 91, 559–584. [Google Scholar]
- Bostoen, K.; Clist, B.; Doumenge, C.; Grollemund, R.; Hombert, J.M.; Muluwa, J.K.; Maley, J.; Blench, R.; Di Carlo, P.; Good, J.; et al. Middle to late Holocene Paleoclimatic change and the early Bantu expansion in the rain forests of Western Central Africa. Curr. Anthropol. 2015, 56, 367–368. [Google Scholar] [CrossRef]
- Neumann, K.; Bostoen, K.; Höhn, A.; Kahlheber, S.; Ngomanda, A.; Tchiengué, B. First farmers in the Central African rainforest: A view from southern Cameroon. Quat. Int. 2012, 249, 53–62. [Google Scholar] [CrossRef]
- Giresse, P.; Maley, J.; Chepstow-Lusty, A. Understanding the 2500 yr BP rainforest crisis in West and Central Africa in the framework of the Late Holocene: Pluridisciplinary analysis and multi-archive reconstruction. Glob. Planet. Chang. 2020, 192, 103257. [Google Scholar] [CrossRef]
- Röschenthaler, U.M. Translocal cultures: The slave trade and cultural transfer in the Cross River region. Soc. Anthropol. 2006, 14, 71–91. [Google Scholar] [CrossRef]
- Michels, S. Imagined Power Contested: Germans and Africans in the Upper Cross River Area of Cameroon, c. 1887–c. 1915; ’Lit Verlag: Münster, Germany, 2004; Volume 2. [Google Scholar]
- Röschenthaler, U.M. Purchasing Culture: The Dissemination of Associations in the Cross River Region of Cameroon and Nigeria; Africa World Press: Trenton, NJ, USA, 2011. [Google Scholar]
- Biwolé, A.B.; Morin-Rivat, J.; Fayolle, A.; Bitondo, D.; Dedry, L.; Dainou, K.; Hardy, O.J.; Doucet, J.L. New data on the recent history of the littoral forests of southern Cameroon: An insight into the role of historical human disturbances on the current forest composition. Plant Ecol. Evol. 2015, 148, 19–28. [Google Scholar] [CrossRef]
- Garcin, Y.; Deschamps, P.; Ménot, G.; De Saulieu, G.; Schefuß, E.; Sebag, D.; Dupont, L.M.; Oslisly, R.; Brademann, B.; Mbusnum, K.G.; et al. Early anthropogenic impact on Western Central African rainforests 2600 y ago. Proc. Natl. Acad. Sci. USA 2018, 115, 3261–3266. [Google Scholar] [CrossRef] [Green Version]
- Morin-Rivat, J.; Fayolle, A.; Favier, C.; Bremond, L.; Gourlet-Fleury, S.; Bayol, N.; Lejeune, P.; Beeckman, H.; Doucet, J.L. Present-day central African forest is a legacy of the 19th century human history. eLife 2017, 6, e20343. [Google Scholar] [CrossRef] [Green Version]
- Sharwood-Smith, B.E.; Cantle, L.L. An Assessment Report on the Anyang and Manta Tribes of Mamfe Division, Cameroons Province; Buea National Archive AF4; Buea National Archive: Buea, Cameroon, 1924. [Google Scholar]
- Courade, G.; Ngo-Botok, V.; Ngum-Kwe, F. Village Dictionary of Manyu Division; Office de la Recherche Scientifique et Technique Outre-mer: Yaounde, Cameroun, 1973. [Google Scholar]
- Hull, G.; Cooper, J.E. Catalogue of preserved gorilla materials. In Gorilla Pathology and Health: With a Catalogue of Preserved Materials; Hull, G., Cooper, J.E., Eds.; Elsevier Inc.: London, UK, 2017; pp. 379–602. [Google Scholar]
- Blake, S. The Ecology of Forest Elephant Distribution and Its Implications for Conservation. Ph.D. Thesis, University of Edinburgh, Edinburgh, UK, 2002. [Google Scholar]
- Bekhuis, P.D.; De Jong, C.B.; Prins, H.H. Diet selection and density estimates of forest buffalo in Campo-Ma’an National Park, Cameroon. Afr. J. Ecol. 2008, 46, 668–675. [Google Scholar] [CrossRef]
- Eniang, E.A.; Ebin, C.O.; Nchor, A.A.; Agba, G.I.; Gideon, I.K.; Amori, G.; Luiselli, L. Distribution and status of the African forest buffalo Syncerus caffer nanus in south-eastern Nigeria. Oryx 2017, 51, 538–541. [Google Scholar] [CrossRef] [Green Version]
- Head, J.S.; Boesch, C.; Makaga, L.; Robbins, M.M. Sympatric chimpanzees (Pan troglodytes troglodytes) and gorillas (Gorilla gorilla gorilla) in Loango National Park, Gabon: Dietary composition, seasonality, and intersite comparisons. Int. J. Primatol. 2011, 32, 755–775. [Google Scholar] [CrossRef]
- Bergl, R.A. Conservation Biology of the Cross River gorilla (Gorilla gorilla diehli). Ph.D. Thesis, The City University of New York, New York, NY, USA, 2006. [Google Scholar]
- Nuesiri, E.O. Conservation and resource use in Cross River—Takamanda Forest Complex of Cameroon and Nigeria; Fauna and Flora International: Cambridge, UK, 2003. [Google Scholar]
- Bitondo, D.; Schmidt-Soltau, K.; Ntep, J.; Onya, H.; Wadt Zela, F.F.; Frida, N.A.; Tambe, J.B.; Ebot, R. The Mamfe—Akwaya Road Project: An Environmental Impact Assessment; German Development Cooperation: Buea, Cameroon, 1999. [Google Scholar]
- Oates, J.F. Myth and Reality in the Rain Forest: How Conservation Strategies Are Failing in West Africa; University of California Press: Berkeley, CA, USA, 1999. [Google Scholar]
- Laird, S.A.; Awung, G.L.; Lysinge, R.J. Cocoa farms in the Mount Cameroon region: Biological and cultural diversity in local livelihoods. Biodivers. Conserv. 2007, 16, 2401–2427. [Google Scholar] [CrossRef]
- Klarer, A.J. The Evolution and Expansion of Cacao Farming in South West Cameroon and Its Effects on Local Livelihoods. Ph.D. Thesis, University of Copenhagen, Copenhagen, Denmark, 2014. [Google Scholar]
- Dada, L.A. The African Export Industry: What Happened and How Can It Be Revived. Case Study on the Cameroonian Cocoa Sector; Agricultural Management, Marketing and Finance, Working Paper; Food and Agricultural Organisation (FAO): Geneva, Switzerland, 2007. [Google Scholar]
- Abei, L. An Analysis of the Competitive Performance of the Cameroonian Cocoa Industry. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2017. [Google Scholar]
- Business in Cameroon. Available online: https://www.businessincameroon.com/agriculture/0204-9006-cameroon-cutscocoa-production-forecasts-for-2020-from-600-000-to-285-000-tons (accessed on 29 April 2019).
- Cameroon Concord. Available online: http://www.cameroonconcordnews.com/cameroon-was-never-one-theagbaw-ebai-debate/ (accessed on 17 July 2019).
- Mukete, N.; Li, Z.; Beckline, M.; Patricia, B. Cocoa production in Cameroon: A socioeconomic and technical efficiency perspective. Int. J. Agric. Econ. 2018, 3, 1–8. [Google Scholar] [CrossRef]
- Asase, A.; Tetteh, D.A. The role of complex agroforestry systems in the conservation of forest tree diversity and structure in southeastern Ghana. Agrofor. Syst. 2010, 79, 355–368. [Google Scholar] [CrossRef]
- Degrande, A.; Schreckenberg, K.; Mbosso, C.; Anegbeh, P.; Okafor, V.; Kanmegne, J. Farmers’ fruit tree-growing strategies in the humid forest zone of Cameroon and Nigeria. Agrofor. Syst. 2006, 67, 159–175. [Google Scholar] [CrossRef]
- Herzog, F. Multipurpose shade trees in coffee and cocoa plantations in Côte d’Ivoire. Agrofor. Syst. 1994, 27, 259–267. [Google Scholar] [CrossRef]
- Jagoret, P.; Michel-Dounias, I.; Malézieux, E. Long-term dynamics of cocoa agroforests: A case study in central Cameroon. Agrofor. Syst. 2011, 81, 267–278. [Google Scholar] [CrossRef]
- Ruf, F.; Schroth, G. Chocolate forests and monocultures: A historical review of cocoa growing and its conflicting role in tropical deforestation and forest conservation. In Agroforestry and Biodiversity Conservation in Tropical Landscapes; Schroth, G., Da Fonseca, G.A.B., Harvey, C.A., Gascon, C., Vasconcelos, H.L., Izac, A.M.N., Eds.; Island Press: Washington, DC, USA, 1995; pp. 107–133. [Google Scholar]
- Voigt, M.; Wich, S.A.; Ancrenaz, M.; Meijaard, E.; Abram, N.; Banes, G.L.; Campbell-Smith, G.; d’Arcy, L.J.; Delgado, R.A.; Erman, A.; et al. Global demand for natural resources eliminated more than 100,000 Bornean orangutans. Curr. Biol. 2018, 28, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Human Rights Watch. Available online: https://www.hrw.org/news/2019/05/23/cameroon-promised-investigation-shouldbe-independent (accessed on 3 June 2019).
- Foreign Policy. Available online: https://foreignpolicy.com/2019/05/13/cameroons-separatistmovement-is-going-international-ambazonia-military-forces-amf-anglophonecrisis/ (accessed on 3 June 2019).
- United Nations Office for the Coordination of Humanitarian Affairs. Available online: https://www.humanitarianresponse.info/sites/www.humanitarianresponse.info/files/documents/files/sitrep_nwsw_nov_2020_vf.pdf (accessed on 14 January 2020).
- Gaynor, K.M.; Fiorella, K.J.; Gregory, G.H.; Kurz, D.J.; Seto, K.L.; Withey, L.S.; Brashares, J.S. War and wildlife: Linking armed conflict to conservation. Front Ecol Environ. 2016, 14, 533–542. [Google Scholar] [CrossRef]
- Plumptre, A.J.; Nixon, S.; Critchlow, R.; Vieilledent, G.; Kirkby, A.; Williamson, E.A.; Hall, J.; Kujirakwinja, D. Status of Grauer’s Gorilla and Chimpanzee in Eastern Democratic Republic of Congo: Historical and Current Distribution and Abundance; Wildlife Conservation Society, Fauna & Flora International and Institut Congolais pour la Conservation de la Nature: New York, NY, USA, 2015. [Google Scholar]
- Shambaugh, J.; Oglethorpe, J.; Ham, R. The Trampled Grass: Mitigating the Impacts of Armed Conflict on the Environment; Biodiversity Support Program: Washington, DC, USA, 2001. [Google Scholar]
- Hockings, K.J.; McLennan, M.R.; Carvalho, S.; Ancrenaz, M.; Bobe, R.; Byrne, R.W.; Dunbar, R.I.; Matsuzawa, T.; McGrew, W.C.; Williamson, E.A.; et al. Apes in the Anthropocene: Flexibility and survival. Trends Ecol. Evol. 2015, 30, 215–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLennan, M.R.; Spagnoletti, N.; Hockings, K.J. The implications of primate behavioral flexibility for sustainable human–primate coexistence in anthropogenic habitats. Int. J. Primatol. 2017, 38, 105–121. [Google Scholar] [CrossRef]
Figure 1.
Location of the Mone-Oku study area, Southwest Cameroon. Insert: location of the region within Central-West Africa.
Figure 1.
Location of the Mone-Oku study area, Southwest Cameroon. Insert: location of the region within Central-West Africa.

Figure 2.
A map of gorilla nest sites and botanical plot locations within the gorilla nesting zone of the Mone-Oku Forest.
Figure 2.
A map of gorilla nest sites and botanical plot locations within the gorilla nesting zone of the Mone-Oku Forest.

Figure 3.
Structure and composition of gorilla nesting area habitat by (A) the ten species with the highest importance values and (B) the ten families with the highest importance values.
Figure 3.
Structure and composition of gorilla nesting area habitat by (A) the ten species with the highest importance values and (B) the ten families with the highest importance values.



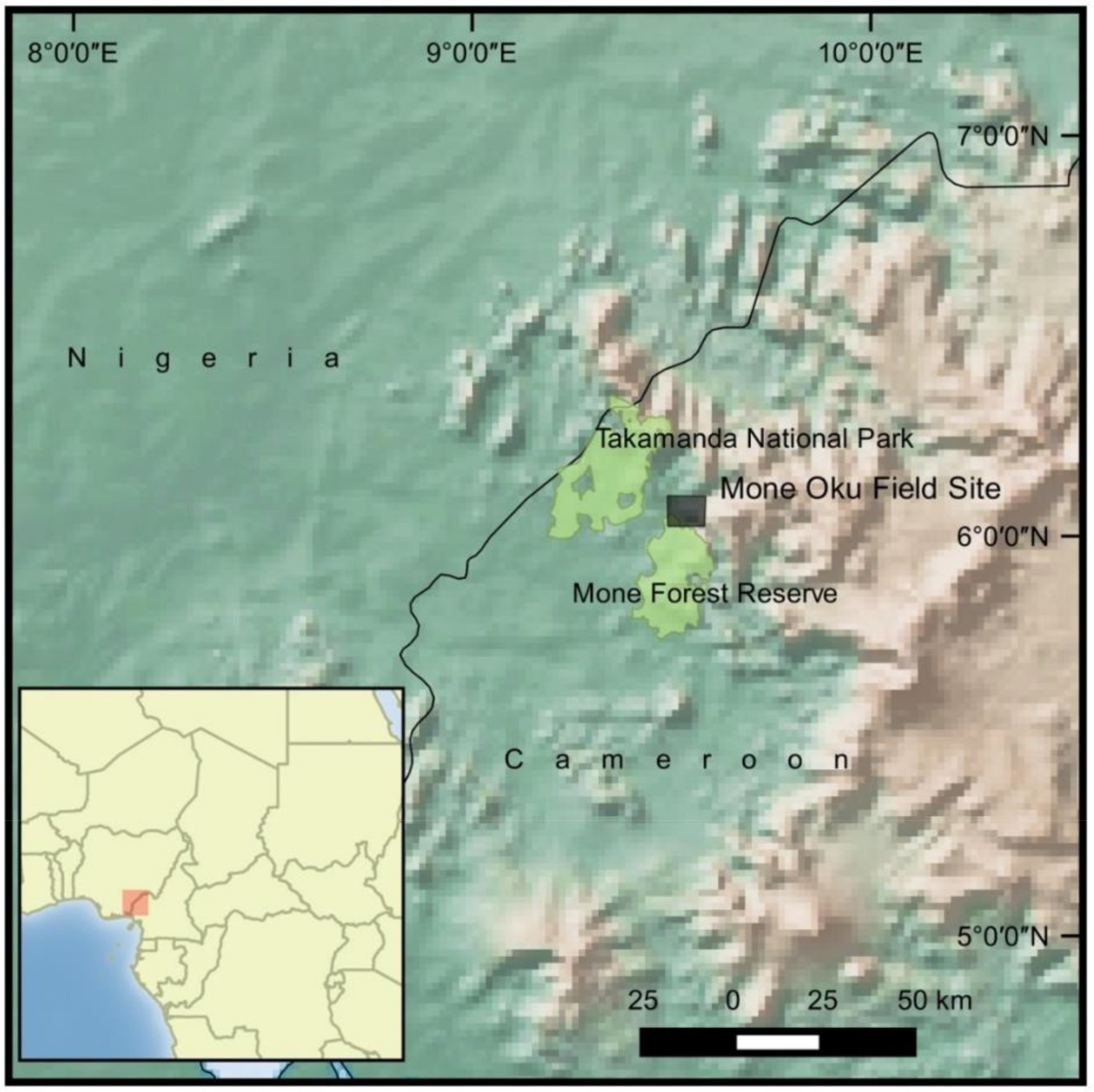{kind=link}
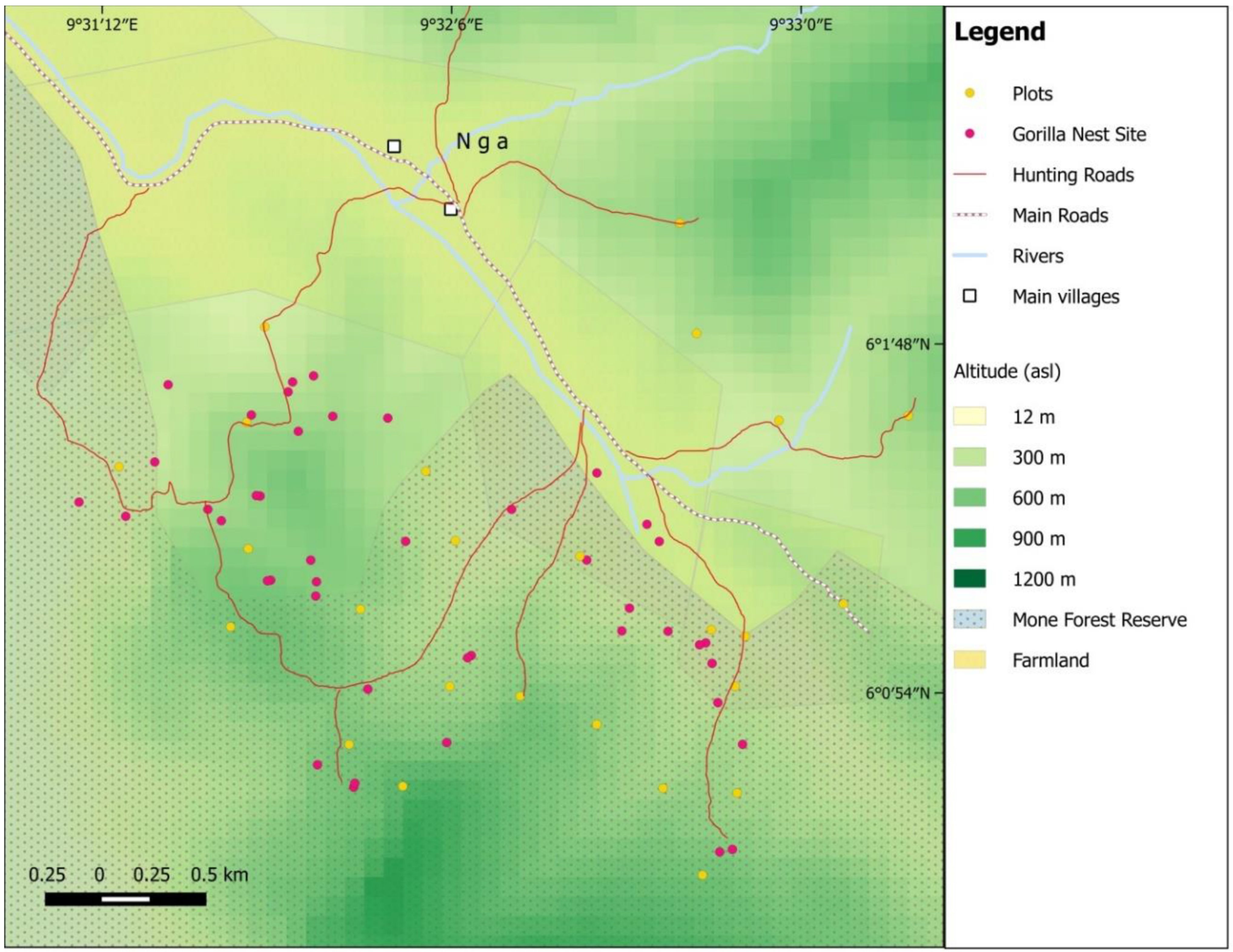{kind=link}
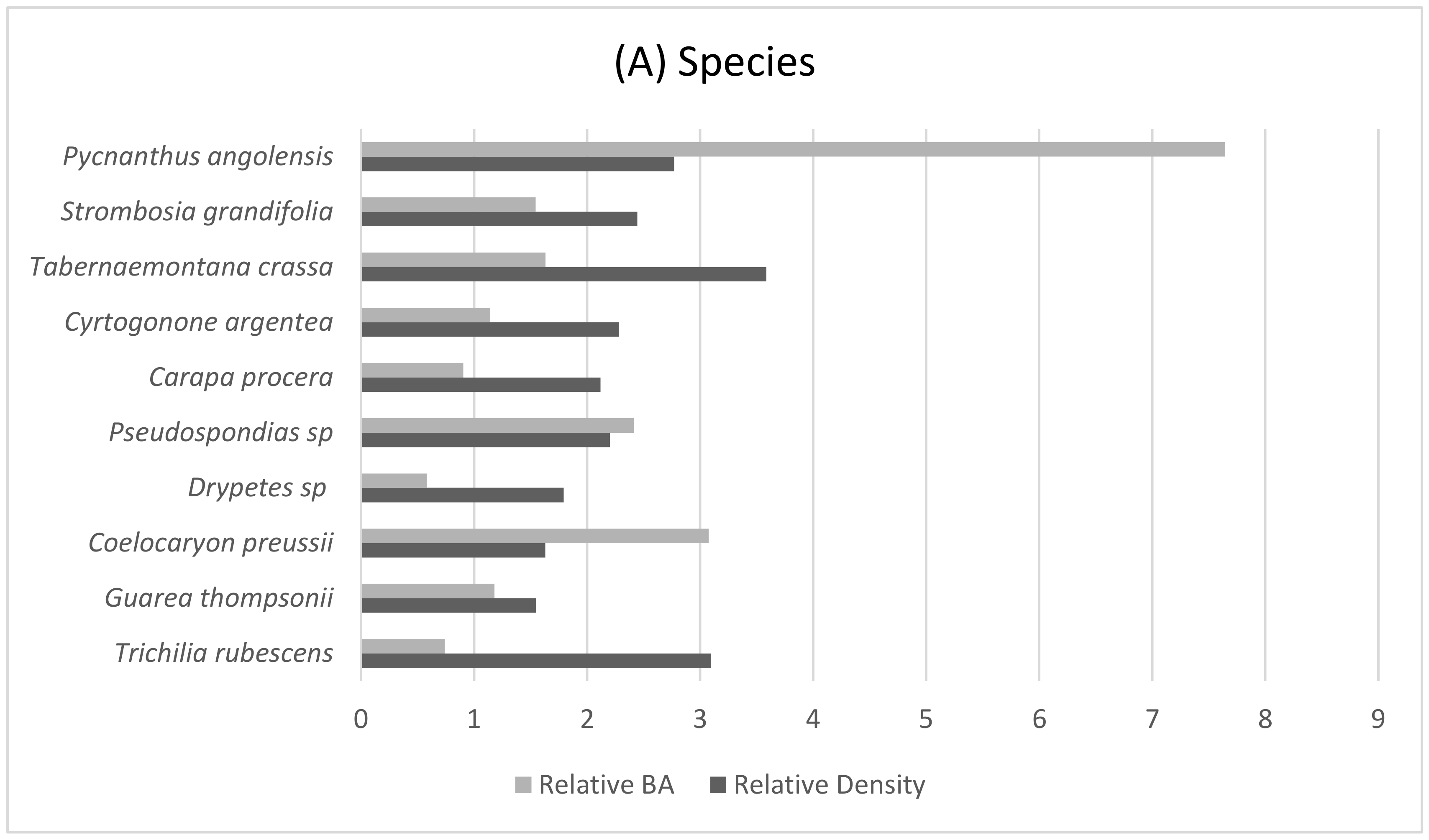{kind=link}
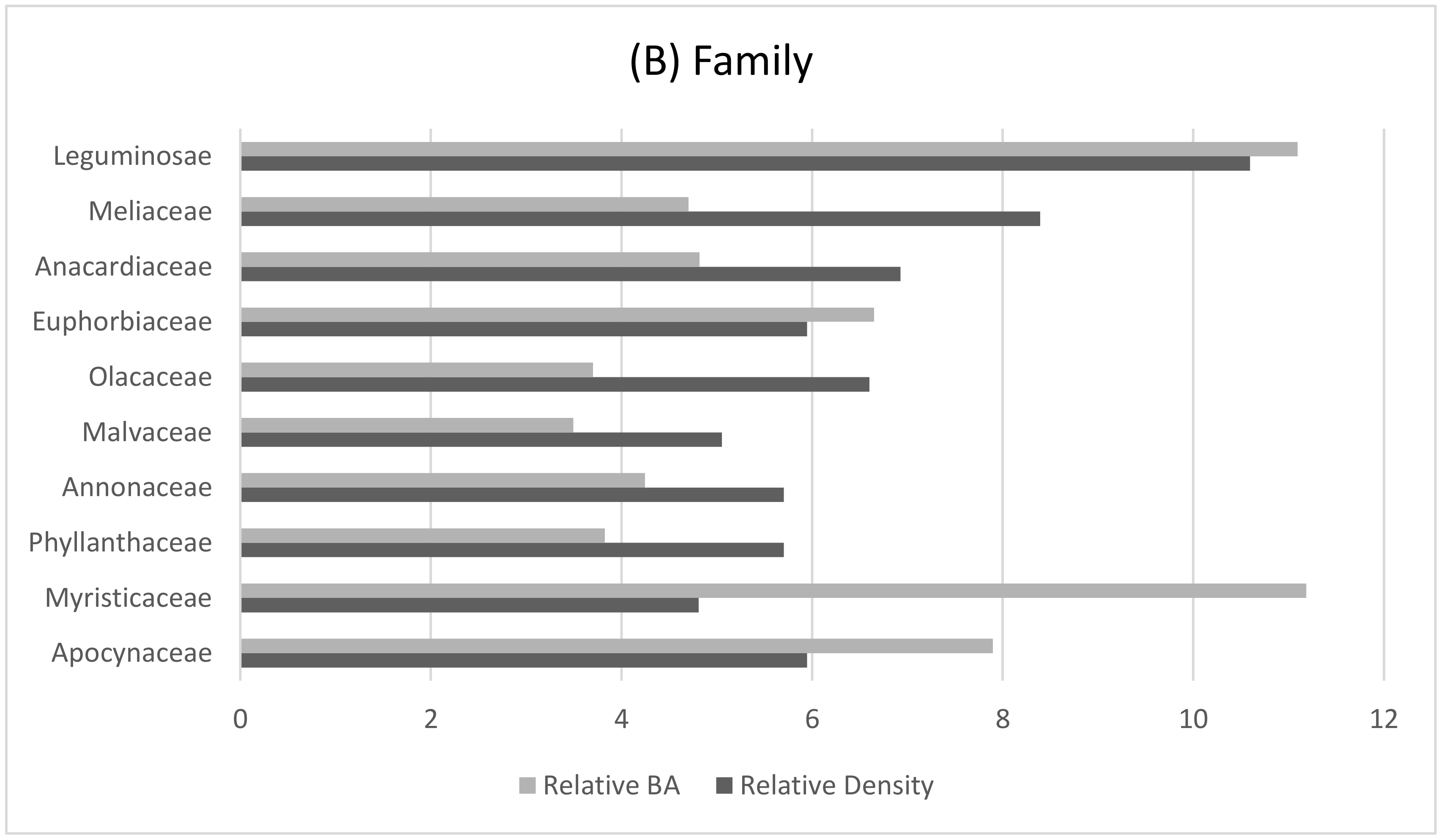{kind=link}
Table 1.
Vegetation classes and key indicator plant species.
| Vegetation Class | Description | Indicator Plants |
|---|---|---|
| Old secondary forest | Forest recovering from disturbance; trees are smaller and canopy height is lower than in primary forest, either from human modification or natural disturbances. | Elaeis guineensis, Musanga cecropioides, Piptadenianstrum africanum, Tabernaemontana crassa, Pycnanthus angolensis |
| Lowland forest < 500 m a.s.l. | Mature forest with tall and large trees; a high canopy and a distinctive middle story. | Afrostyrax kameroensis, Desbordesia spp., Carapa procera, Parkia bicolor, Strombosia grandifolia |
| Lowland ridge forest | Mature forest growing on the sides of finger-like ridges; at some places the forest is interspersed with rocky cliffs. | Allenblackia floribunda, Tapura africana, Chrysophyllum spp., Lophira alata, Pseudospondias microcarpa |
| Mid-elevation forest 500–800 m a.s.l. | Mature forest with a lower canopy and denser understory. Diversity is reduced but also contains elements of both lowland and montane forests. | Homalium spp., Syzygium spp., Xylopia staudtii, Carpolobia lutea, Rinorea dentata, Dichapetalum spp. |
| Montane forest 800–1500 m a.s.l. | Mature forest with a further reduction in canopy height which is often disjunct; a decrease in species richness. | Dactyladeina staudii, Dracenea arborea, Eugenia spp. |
Table 2.
Descriptions of habitat categories [42] (p. 256).
Table 2.
Descriptions of habitat categories [42] (p. 256).
| Category | Description | |
|---|---|---|
| Density of understory | Very dense | (Stems < 50 cm apart) |
| Dense | (Stems < 1 m apart) | |
| Sparse | (Stems > 1 m apart) | |
| Slope | Precipitous | (Over 50%, >26.6°) |
| Steep | (10–50%, 5.7–26.6°) | |
| Gentle | (0–10%, 0–5.7°) | |
| Flat | ||
| Canopy cover | Very closed | (>75%) |
| Closed | (51–75%) | |
| Open | (26–50%) | |
| Very open | (0–25%) |
| Age Category | Approximate Age | Definition |
|---|---|---|
| New | Less than two days | Nest has green unwilted leaves. Presence of dung and urine in or around the nest. |
| Recent | Less than one week | Nest has wilted green leaves. |
| Old and intact | Between one week to one month | Nest remains intact but leaves have turned brown. |
| Decomposing | More than one month | Nest consists of interwoven stems and branches that lack leaves. |
Table 4.
Multinomial logistic regression: habitat characteristics of gorilla nest sites.
| Parameter | Estimate | Standard Error | Z-Score | p-Value | 95% Confidence Interval | |
|---|---|---|---|---|---|---|
| Lower | Upper | |||||
| Canopy cover | −0.357 | 0.253 | −1.412 | 0.158 | −0.853 | 0.139 |
| Elevation | −0.679 | 0.189 | −3.589 | <0.001 | −1.049 | −0.308 |
| Degree of slope | 1.029 | 0.387 | 2.660 | 0.008 | 0.271 | 1.788 |
Table 5.
Floristic characteristics from each vegetation class and ape nesting ranges.
| Vegetation Class | Plots (n) | Trees DBH > 10 cm (n) | Tree Density (Trees/ha) | Tree Species (n) | Understory Species (n) | Stem Density (Stems/m2) | Mean Height (m) | Mean DBH (cm) | Stand Basal Area (m2/ha) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M | SD | M | SD | M | SD | |||||||
| Old secondary | 9 | 472 | 520 | 119 | 129 | 24 | 11 | 16 | 8 | 23 | 17 | 34 |
| Lowland | 42 | 2083 | 500 | 203 | 212 | 29 | 18 | 16 | 7 | 23 | 15 | 29 |
| Lowland ridge | 7 | 365 | 520 | 116 | 80 | 21 | 9 | 17 | 8 | 24 | 19 | 39 |
| Mid-elevation | 53 | 2824 | 530 | 215 | 226 | 22 | 20 | 16 | 8 | 23 | 16 | 33 |
| Montane | 9 | 517 | 570 | 120 | 113 | 26 | 20 | 14 | 7 | 22 | 15 | 32 |
| Ape nesting range | ||||||||||||
| Chimpanzee | 23 | 1303 | 570 | 164 | 143 | 15 | 6 | 16 | 10 | 24 | 17 | 39 |
| Gorilla | 25 | 1227 | 490 | 183 | 171 | 24 | 20 | 16 | 8 | 23 | 16 | 29 |
Table 6.
Mean and median number of nest stems and proportion of the 15 most commonly used plant genera within night nests constructed by gorillas, and the two-metre radius around the nest. Note: * denotes statistical significance; ** denotes species recorded in diet of Mone-Oku gorillas [36].
Table 6.
Mean and median number of nest stems and proportion of the 15 most commonly used plant genera within night nests constructed by gorillas, and the two-metre radius around the nest. Note: * denotes statistical significance; ** denotes species recorded in diet of Mone-Oku gorillas [36].
| Genera | Form | # of Times Plant Used in Nest | Mean # of Stems in Nest | Median Proportion of Stems in Nest | # of Times Plant Was Available in Environment | Mean # of Stems in Environment | Median Proportion of Stems in Environment | Median of the Difference | p-Value | r-Value | Preference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Aframomum** | Herb | 406 | 13 | 0.50 | 644 | 22 | 0.28 | −0.1146 | * 0.016 | 0.41 | Preferred |
| Acanthaceae | Herb | 104 | 5 | 0.35 | 582 | 14 | 0.24 | 0.1163 | * 0.001 | 0.49 | Avoided |
| Urera ** | Liana | 87 | 5 | 0.14 | 150 | 7 | 0.03 | −0.0402 | 0.171 | - | Neutral |
| Palisota ** | Herb | 58 | 2 | 0.03 | 172 | 9 | 0.05 | 0.0213 | 0.317 | - | Neutral |
| Piper ** | Vine | 54 | 6 | 0.02 | 60 | 3 | 0.02 | 0.0206 | 0.198 | - | Neutral |
| Marantaceae | Herb | 53 | 13 | 0.08 | 92 | 9 | 0.05 | 0.0435 | 0.600 | - | Neutral |
| Alchornea | Shrub | 46 | 4 | 0.13 | 71 | 5 | 0.07 | −0.0563 | 0.053 | - | Neutral |
| Leea ** | Shrub | 40 | 4 | 0.05 | 34 | 2 | 0.02 | −0.0215 | 0.136 | - | Neutral |
| Protomegabaria ** | Tree | 39 | 6 | 0.13 | 20 | 7 | 0.02 | −0.1339 | * 0.025 | 0.79 | Preferred |
| Penianthus ** | Shrub | 38 | 3 | 0.03 | 108 | 4 | 0.05 | 0.0339 | 0.673 | - | Neutral |
| Thaumatococcus ** | Herb | 30 | 30 | 0.75 | 37 | 37 | 0.79 | 0.0372 | 0.317 | - | Neutral |
| Landolphia ** | Liana | 28 | 2 | 0.04 | 49 | 2 | 0.02 | 0.0222 | 0.248 | - | Neutral |
| Sorindeia ** | Shrub | 19 | 1 | 0.02 | 71 | 2 | 0.03 | 0.0233 | 0.512 | - | Neutral |
| Rinorea ** | Shrub | 18 | 2 | 0.03 | 56 | 2 | 0.03 | 0.0241 | 0.347 | Neutral | |
| Pavetta ** | Shrub | 17 | 2 | 0.01 | 59 | 2 | 0.03 | 0.0270 | 0.163 | - | Neutral |
Table 7.
Mean and median number of nest stems and proportion of the preferred plant genera within night nests constructed by gorillas and the two-metre radius around the nest.
Table 7.
Mean and median number of nest stems and proportion of the preferred plant genera within night nests constructed by gorillas and the two-metre radius around the nest.
| Genera | Form | # of Times Plant Used in Nest | Mean # of Stems in Nest | Median Proportion of Stems in Nest | # of Times Plant Was Available in Environment | Mean # of Stems in Environment | Median Proportion of Stems in Environment | Median of the Difference | p-Value | r-Value |
|---|---|---|---|---|---|---|---|---|---|---|
| Aframomum | Herb | 406 | 13 | 0.50 | 644 | 22 | 0.28 | −0.1146 | 0.016 | 0.41 |
| Protomegabaria | Tree | 39 | 6 | 0.13 | 20 | 7 | 0.02 | −0.1339 | 0.025 | 0.79 |
| Tiliaceae | Vine | 13 | 2 | 0.07 | 6 | 2 | 0.01 | −0.0606 | 0.028 | 0.69 |
Table 8.
Preference of tree species for arboreal nesting in the 13 most commonly used nesting tree species. Note: Value for neutral preference for Manly’s α was 0.005. ** denotes species recorded in the diet of Mone-Oku gorillas [36].
Table 8.
Preference of tree species for arboreal nesting in the 13 most commonly used nesting tree species. Note: Value for neutral preference for Manly’s α was 0.005. ** denotes species recorded in the diet of Mone-Oku gorillas [36].
| Tree Species | Observed # of Trees Used for Nesting (1) | Cumulative % | Proportion of (1) to All Trees Used for Nesting | # of Trees Sampled in Botanical Plots (2) | Proportion of (2) to All Trees Sampled | Expected # of Trees Used for Nesting | Manly’s α | Category by Significant Preference | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| Sorindeia sp. 2 ** | 10 | 17 | 0.172 | 63 | 0.033 | 1.9 | 0.019 | Preferred | <0.001 |
| Pycnanthus angolensis ** | 6 | 28 | 0.103 | 49 | 0.026 | 1.5 | 0.015 | Preferred | 0.004 |
| Microdesmis sp. | 3 | 33 | 0.052 | 24 | 0.013 | 0.7 | 0.015 | Preferred | 0.04 |
| Angylocalyx pynaertii | 2 | 36 | 0.034 | 44 | 0.023 | 1.3 | 0.005 | Neutral | NS |
| Chrysophyllum sp. ** | 2 | 40 | 0.034 | 12 | 0.006 | 0.4 | 0.020 | Preferred | 0.048 |
| Cola lateritia | 2 | 43 | 0.034 | 8 | 0.004 | 0.2 | 0.030 | Preferred | 0.023 |
| Maesobotrya sp. | 2 | 47 | 0.034 | 18 | 0.009 | 0.5 | 0.013 | Preferred | NS |
| Napoleonaea vogelii ** | 2 | 50 | 0.034 | 24 | 0.013 | 0.7 | 0.010 | Preferred | NS |
| Protomegabaria sp. ** | 2 | 53 | 0.034 | 18 | 0.009 | 0.5 | 0.013 | Preferred | NS |
| Pseudospondias sp. ** | 2 | 57 | 0.034 | 29 | 0.015 | 0.9 | 0.008 | Preferred | NS |
| Tabernaemontana crassa | 2 | 60 | 0.034 | 49 | 0.026 | 1.5 | 0.005 | Neutral | NS |
| Trema sp. | 2 | 64 | 0.034 | 1 | 0.001 | 0.0 | 0.240 | Preferred | 0.002 |
| Trichilia rubescens ** | 2 | 67 | 0.034 | 46 | 0.024 | 1.4 | 0.005 | Neutral | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wade, A.H.; Malone, N. Ecological, Historical, Economic, and Political Factors Shaping the Human–Gorilla Interface in the Mone-Oku Forest, Cameroon. Diversity 2021, 13, 175. https://0-doi-org.brum.beds.ac.uk/10.3390/d13040175
AMA Style
Wade AH, Malone N. Ecological, Historical, Economic, and Political Factors Shaping the Human–Gorilla Interface in the Mone-Oku Forest, Cameroon. Diversity. 2021; 13(4):175. https://0-doi-org.brum.beds.ac.uk/10.3390/d13040175
Chicago/Turabian StyleWade, Alison H., and Nicholas Malone. 2021. "Ecological, Historical, Economic, and Political Factors Shaping the Human–Gorilla Interface in the Mone-Oku Forest, Cameroon" Diversity 13, no. 4: 175. https://0-doi-org.brum.beds.ac.uk/10.3390/d13040175
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.