
Linking Habitat and Associated Abiotic Conditions to Predict Fish Hotspots Distribution Areas within La Paz Bay: Evaluating Marine Conservation Areas

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
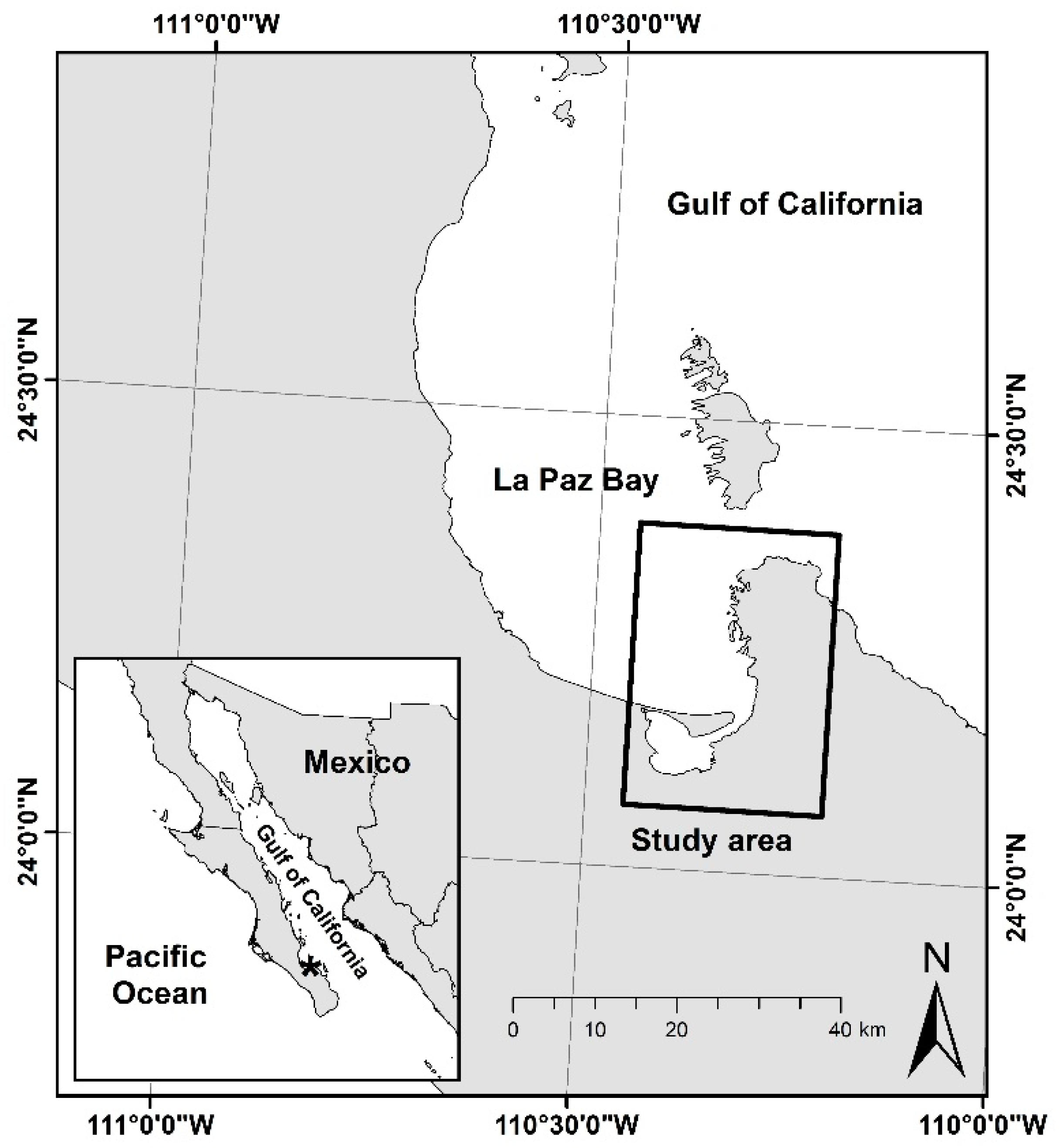
2.1. Study Area
2.2. Methods
2.2.1. Occurrence Data
2.2.2. Environmental Layers and Calibration Area
2.2.3. Model Calibration and Evaluation
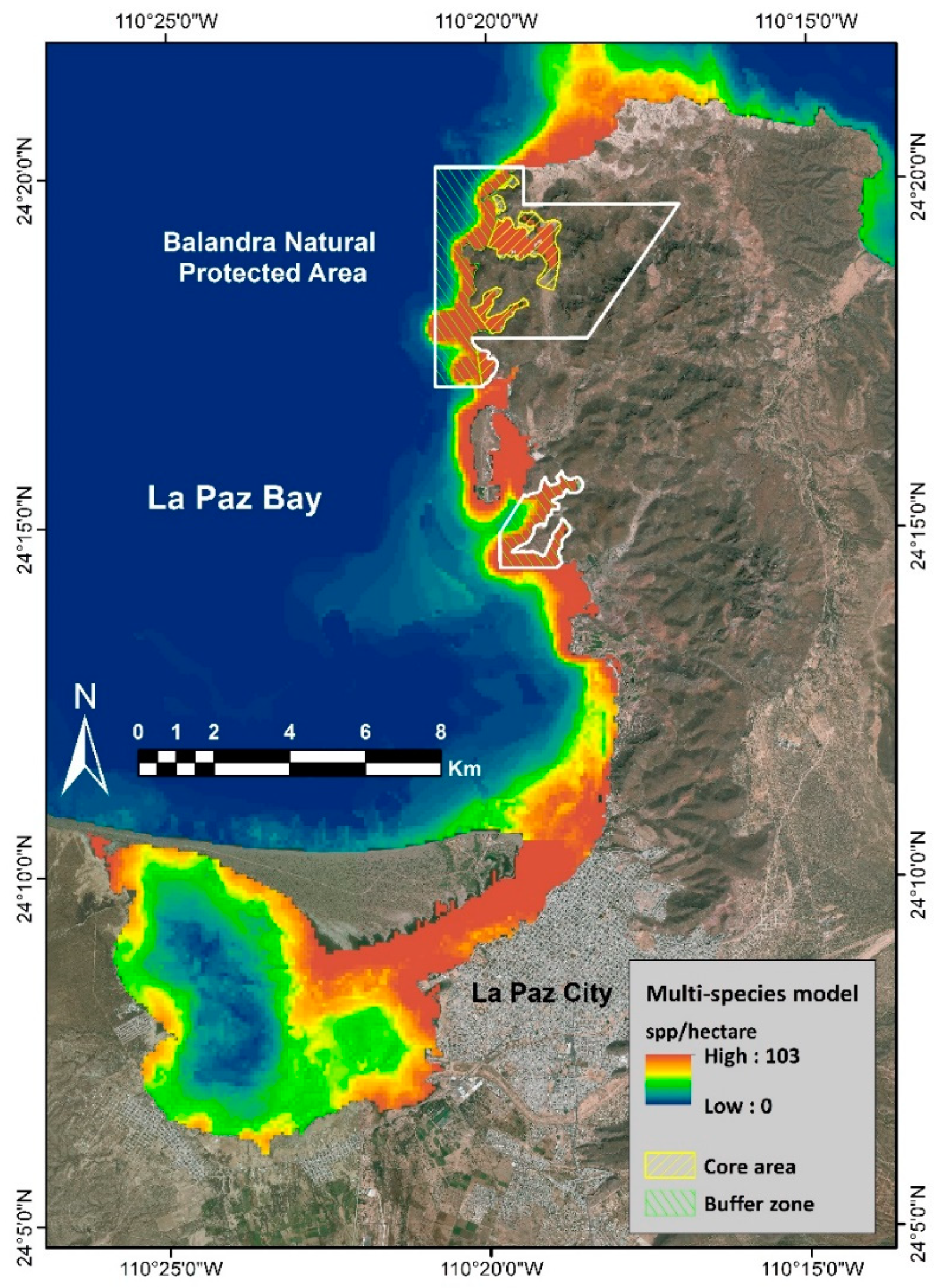
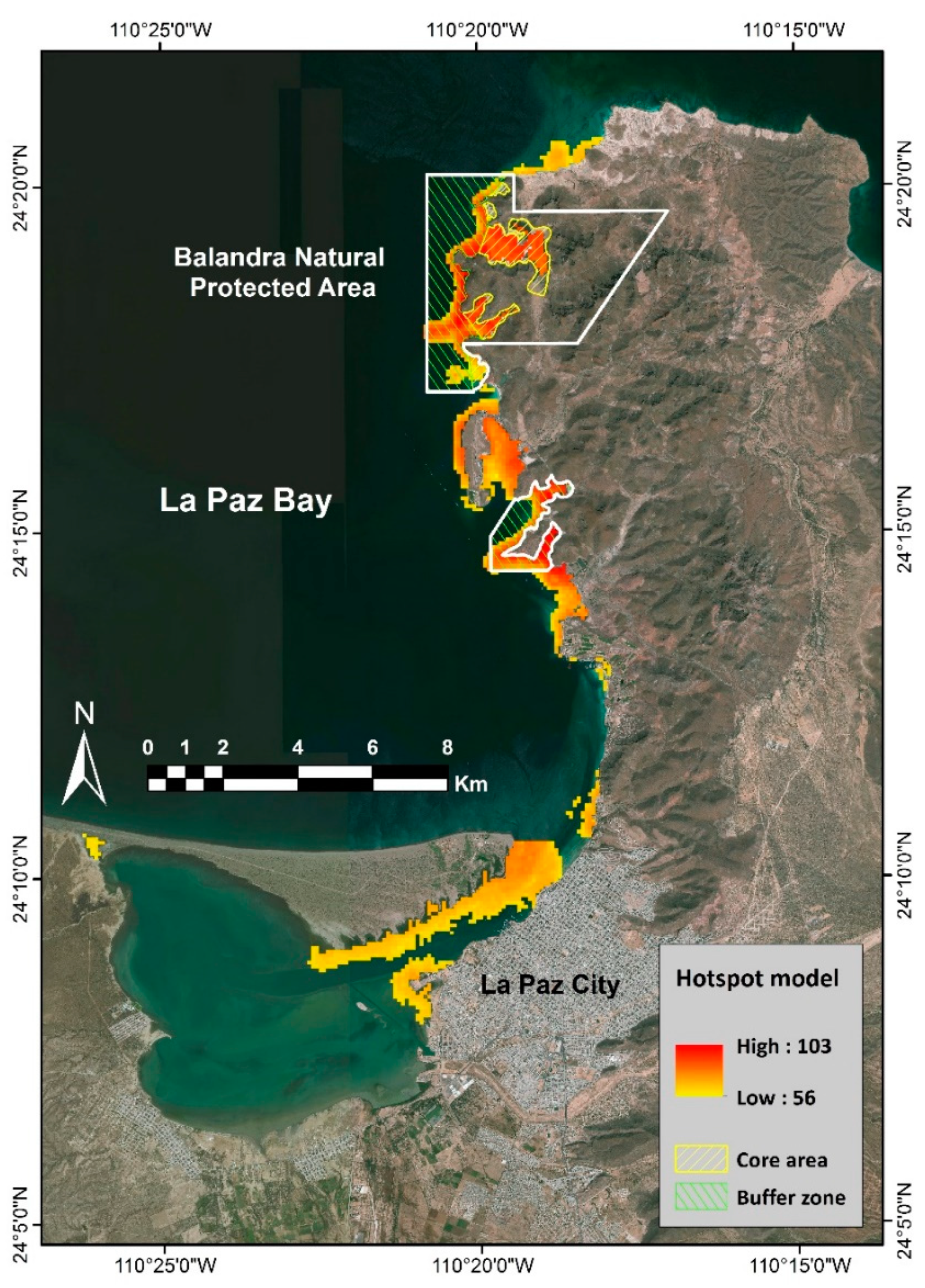
2.2.4. Multi-Species and Hotspot Models
3. Results
4. Discussion
4.1. Hotspot vs. Marine Conservation Areas
4.2. What Do the Models Indicate?
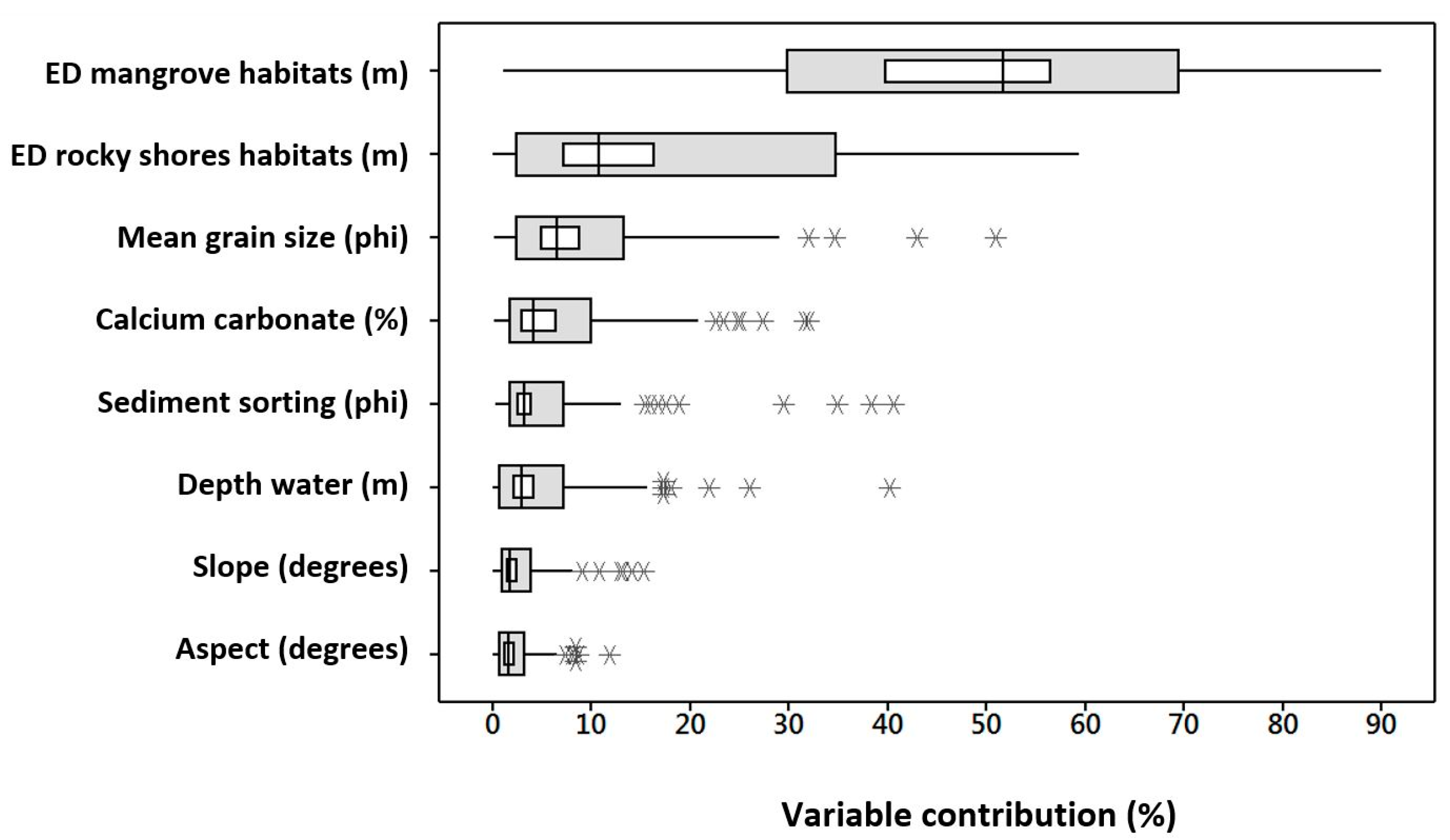
4.3. Variables and Scales
4.4. Impact and Applications
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Girardello, M.; Santangeli, A.; Mori, E.; Chapman, A.; Fattorini, S.; Naidoo, R.; Bertolino, S.; Svenning, J.-C. Global synergies and trade-off between multiple dimensions of biodiversity and ecosystem services. Sci. Rep. 2019, 9, 5636. [Google Scholar] [CrossRef]
- Brooks, T.M.; Mittermeier, R.A.; da Fonseca, G.A.; Gerlach, J.; Hoffmann, M.; Lamoreux, J.F.; Mittermeier, C.G.; Pilgrim, J.D.; Rodrigues, A.S. Global biodiversity conservation priorities. Science 2006, 313, 58–61. [Google Scholar] [CrossRef] [Green Version]
- Selig, E.R.; Turner, W.R.; Troeng, S.; Wallace, B.P.; Halpern, B.S.; Kaschner, K.; Lascelles, B.G.; Carpenter, K.E.; Mitter-meir, R.A. Global priorities for marine biodiversity conservation. PLoS ONE 2014, 9, e82898. [Google Scholar] [CrossRef]
- Mora, C.; Titensor, D.P.; Adl, S.; Simspn, A.G.B.; Worm, B. How many species are there on Earth and in the ocean? PLoS Biol. 2011, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, J.E.; Folke, C.; Halpern, B.S.; Jackson, J.B.C.; Lotze, H.K.; Micheli, F.; Pa-Lumbi, S.R.; et al. Impacts of biodiversity loss on ocean ecosystem services. Science 2006, 314, 787–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoegh-Guldberg, O.; Bruno, J.F. The impact of climate change on the world’s marine ecosystems. Science 2010, 328, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Kano, Y.; Dudgeon, D.; Nam, S.; Samejima, H.; Watanabe, K.; Grudpan, C.; Magtoon, W.; Musikasinthorn, P.; Thanh Nguyen, P.; Praxaysonbath, B.; et al. Impacts of dams and global warming on fish biodiversity in the Indo-Burma hotspot. PLoS ONE 2016, 11, e0160151. [Google Scholar] [CrossRef] [Green Version]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global biodiversity conservation: The critical role of hotspots. In Biodiversity Hotspots; Zachos, F.E., Habel, J.C., Eds.; Springer Publishers: London, UK, 2011; pp. 3–22. [Google Scholar]
- Cañadas, E.M.; Fenu, G.; Peñas, J.; Lorite, J.; Mattana, E.; Bacchetta, G. Hotspots within hotspots: Endemic plant richness, environmental drivers, and implications for conservation. Biol. Conserv. 2014, 170, 282–291. [Google Scholar] [CrossRef]
- Tittensor, D.P.; Mora, C.; Walter, J.; Lotze, W.; Ricard, D.; Berghe, E.V.; Worm, B. Global patterns and predictors of marine biodiversity across taxa. Nature 2010, 466, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Stuart-Smith, R.D.; Bates, A.E.; Lefcheck, J.S.; Duffy, J.E.; Baker, S.C.; Thomson, R.J.; Stuart-Smith, J.F.; Hill, N.A.; Kinin-month, S.J.; Airoldi, L.; et al. Integrating abundance and functional traits reveals new global hotspots of fish diversity. Nature 2013, 501, 539–542. [Google Scholar] [CrossRef]
- McMahan, D.; Fuentes-Montejo, C.E.; Ginger, L.; Carrasco, J.C.; Chakrabarty, P.; Matamoros, W.A. Climate change models predict decreases in the range of a microendemic freshwater fish in Honduras. Sci. Rep. 2020, 10, 12693. [Google Scholar] [CrossRef]
- Jetz, W.; McPherson, J.M.; Guralnick, R.P. Integrating biodiversity distribution knowledge: Toward a global map of life. Trends Ecol. Evol. 2012, 27, 151–159. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Bastos Araújo, M. Ecological Niche and Geographic Distribution; Princeton University Press: Princeton, NJ, USA, 2011; 316 p. [Google Scholar]
- Teixeira, H.; Berg, T.; Uusitalo, L.; Fürhaupter, K.; Heiskanen, A.-S.; Mazik, K.; Lynam, C.P.; Neville, S.; Rodriguez, J.G.; Papadopolou, N.; et al. A catalogue of marine biodiversity indicators. Front. Mar. Sci. 2016, 207, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Newbold, T.; Reader, T.; El-Gabbas, A.; Berg, W.; Shohdi, W.M.; Zalat, S.; Baha El Din, S.; Gilbert, F. Testing the accuracy of species distribution models using species records from a new field survey. Oikos 2010, 119, 1326–1334. [Google Scholar] [CrossRef]
- Baltensperger, A.P.; Huettmann, F. Predictive spatial niche and biodiversity hotspot models for small mammal communities in Alaska: Applying machine-learning to conservation planning. Landsc. Ecol. 2015, 30, 681–697. [Google Scholar] [CrossRef]
- Cooper-Bohannon, R.; Rebelo, H.; Jones, G.; Cotterill, F.; Monadjem, A.; Schoeman, M.C.; Taylor, P.; Park, K. Predicting bat distributions and diversity hotspots in southern Africa. Hystrix Ital. J. Mamm. 2016, 27, e11722. [Google Scholar] [CrossRef]
- Smith, J.N.; Kelly, N.; Renner, I.W. Validation of presence-only models for conservation planning and the application to whales in a multiple-use marine park. Ecol. Appl. 2020, 31, e02214. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, H. Study on rare and endangered plants under climate: Maxent modeling for identifying hot spots in northwest China. CERNE 2021, 27, e-102667. [Google Scholar] [CrossRef]
- Robinson, N.M.; Nelson, W.A.; Cosello, M.; Sutherland, J.; Lundquist, C. A systematic review of marine-based species distribution models (SDMs) with recommendations for best practice. Front. Mar. Sci. 2017, 421, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.E.; McInerny, G.; O’Hara, K.; Harper, R.; Salido, L.; Emmott, S.; Joppa, L.N. Scientists and software–surveying the species distribution modelling community. Divers. Distrib. 2015, 21, 258–267. [Google Scholar] [CrossRef]
- González-Acosta, A.F.; Balart, E.F.; Ruiz-Campos, G.; Espinosa Pérez, H.; Cruz-Escalona, V.H.; Hernández-López, A. Diversidad y conservación de los peces de la bahía de La Paz, Baja California Sur, México. Rev. Mex. Biodiver. 2018, 89, 705–740. [Google Scholar] [CrossRef]
- Galván-Piña, V.H.; Galván-Magaña, F.; Abitia-Cárdenas, L.A.; Gutiérrez-Sánchez, F.J.; Rodríguez-Romero, J. Seasonal structure of fish assemblages in rocky and sandy habitats in Bahía de La Paz, Mexico. Bull. Mar. Sci. 2003, 72, 19–35. [Google Scholar]
- Diario Oficial de la Federación. Decreto por el que se Declara Área Natural Protegida, con el Carácter de Área de Protección de Flora y Fauna, la Región Conocida como Balandra, Localizada en el Municipio de La Paz, en el Estado de Baja California Sur; Secretaría de Medio Ambiente y Recursos Naturales: Ciudad de México, Mexico, 2012. [Google Scholar]
- Diario Oficial de la Federación. Resumen del Programa de Manejo del Área de Protección de Flora y Fauna Balandra; Secretaría de Medio Ambiente y Recursos Naturales: Ciudad de Mexico, Mexico, 2015. [Google Scholar]
- González-Acosta, A.F.; De-la-Cruz-Agüero, G.; De-la-Cruz-Agüero, J.; Ruiz-Campos, G. Ictiofauna asociada al manglar del estero El Conchalito, Ensenada de La Paz, B.C.S., México. CICIMAR Oceán. 1999, 14, 121–131. [Google Scholar]
- Choumiline, K.; Godínez-Orta, L.; Nikolaeva, A.; Derkachev, A.; Shumilin, E. Evaluation of contribution sources for the sediments of the La Paz Lagoon based on statistical treatment of the mineralogy of their heavy fraction and surrounding rock and drainage basin characteristics. Bol. Soc. Geol. Mex. 2009, 61, 97–109. [Google Scholar] [CrossRef]
- Nava-Sánchez, E.H.; Gorsline, D.S.; Molina-Cruz, A. The Baja California peninsula borderland: Structural and sedimentological characteristics. Sediment. Geol. 2001, 144, 63–82. [Google Scholar] [CrossRef]
- Halfar, J.; Ingle, J., Jr.; Godínez-Orta, L. Modern non-tropical mixed carbonate-siliciclastic sediments and environments of the southwestern Gulf of California, Mexico. Sediment. Geol. 2004, 165, 93–115. [Google Scholar] [CrossRef]
- Silverberg, N.; Aguirre-Bahena, F.; Mucci, A. Time-series measurements of settling particulate matter in Alfonso Basin, La Paz Bay, southwestern Gulf of California. Cont. Shelf Res. 2014, 84, 169–187. [Google Scholar] [CrossRef]
- Urcádiz-Cázares, F.J.; Cruz-Escalona, V.H.; Nava-Sánchez, E.; Ortega-Rubio, A. Clasificación de unidades del fondo marino a partir de la distribución espacial de los sedimentos superficiales de la Bahía de La Paz, Golfo de California. Hidrobiológica 2017, 27, 399–409. [Google Scholar] [CrossRef]
- Steller, D.L.; Riosmena-Rodríguez, R.; Foster, M.S.; Roberts, C.A. Rhodolith bed diversity in the Gulf of California: The importance of rhodoliths structure and consequences of disturbance. Aquat. Conserv. 2003, 13, 5–20. [Google Scholar] [CrossRef]
- Abitia-Cárdenas, L.A.; Rodríguez-Romero, J.; Galván-Magaña, F.; De-la-Cruz-Agüero, J.; Chávez-Ramos, H. Lista sistemática de la ictiofauna de Bahía de La Paz, Baja California Sur, México. Cienc. Mar. 1994, 20, 159–181. [Google Scholar] [CrossRef] [Green Version]
- Urcádiz-Cázares, F.J.; Cruz-Escalona, V.H.; Peterson, M.; Marín-Enríquez, M.; González-Acosta, A.F.; Martínez-Flores, G.; Hernández-Carmona, G.H.; Aguilar Medrano, R.; Del Pino-Machado, A.; Ortega-Rubio, A. Ecological niche modelling of endemic fish within La Paz Bay: Implications for conservation. J. Nat. Conserv. 2021, 60, 125981. [Google Scholar] [CrossRef]
- Arriaga Cabrera, L.; Aguilar, V.; Espinoza, J.M. Regiones Prioritarias y Planeación para la Conservación de la Biodiversidad. In: Capital Natural de México. Vol. II: Estado de Conservación y Tendencias de Cambio; CONABIO: Ciudad de México, Mexico, 2009; pp. 433–457. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modelling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.R. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Pearson, G.R.; Raxworthy, C.J.; Nakamura, M.; Peterson, T.A. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Monk, J.; Ierodiaconou, D.; Versace, V.L.; Bellgrove, A.; Harvey, E.; Rattray, A.; Laurenson, L.; Quinn, G.P. Habitat suit-ability for marine fishes using presence-only modelling and multibeam sonar. Mar. Ecol. Prog. Ser. 2010, 420, 157–174. [Google Scholar] [CrossRef] [Green Version]
- Wisz, M.S.; Hijmans, R.J.; Peterson, A.T.; Graham, C.H.; Guisan, A. Effect of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C.; Baca-González, V. MaxEnt’s parameter configuration and small samples: Are we paying attention to recommendations? A systematic review. PeerJ 2017, 5, e3093. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of Maxent for ecologist. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Zayas-Álvarez, J.A. Análisis Temporal de la Estructura Comunitaria de los Peces Crípticos Asociados A un Arrecife Artificial en Punta Diablo, Bahía de La Paz, B.C.S., México. Master’s Thesis, Centro de Investigaciones Biológicas del Noroeste S.C., La Paz, Mexico, 2015. [Google Scholar]
- Balart, E.F.; González-Cabello, A.; Romero-Ponce, C.; Zayas-Alvarez, A.; Calderón-Parra, M.; Campos-Dávila, L.; Find-Ley, L.T. Length-weight relationships of cryptic reef fishes from the southwestern Gulf of California, México. J. Appl. Ichthyol. 2006, 22, 316–318. [Google Scholar] [CrossRef]
- López-Rasgado, F.J.; Herzka, S.Z.; Del-Monte-Luna, P.; Serviere-Zaragoza, E.; Balart, E.F.; Lluch-Cota, S.E. Fish assemblages in three arid mangrove systems of the Gulf of California: Comparing observations from 1980 and 2010. Bull. Mar. Sci. 2012, 88, 919–945. [Google Scholar] [CrossRef]
- GBIF.org. GBIF Occurrence. Available online: https://10.15468/dl.k5mc7e (accessed on 25 February 2021).
- Sillero, N.; Barbosa, M. Common mistakes in ecological niche models. Int. J. Geogr. Inf. Syst. 2021, 35, 213–226. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction (Ecology, Biodiversity and Conservation); Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Schmiing, M.; Diogo, H.; Serrão Santos, R.; Alfonso, P. Assessing hotspots within hotspots to conserve biodiversity and support fisheries management. Mar. Ecol. Prog. Ser. 2014, 513, 187–199. [Google Scholar] [CrossRef]
- Austin, M.P.L. Spatial prediction of species distribution: An interface between ecological theory and statistical modelling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef] [Green Version]
- Pittman, S.; Brow, K.A. Multi-scale approach for predicting fish species distributions across coral reef seascapes. PLoS ONE 2011, 6, e20583. [Google Scholar] [CrossRef] [PubMed]
- Ŝiaulys, A. Empirical Modelling of Macrozoobenthos Species Distribution and Benthic Habitat Quality Assessment. Ph.D. Thesis, Coastal Research and Planning Institute, Klaipèda University, Klaipèda, Lithuania, 2013. [Google Scholar]
- Snickarsa, M.; Sundblad, G.; Sandström, A.; Ljunggren, L.; Bergström, U.; Johansson, G.; Mattila, J. Habitat selectivity of a substrate spawning fish: Modelling requirements for the Eurasian perch Perca fluviatilis. Mar. Ecol. Prog. Ser. 2010, 398, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Froese, R.; Pauly, D. (Eds.) FishBase. World Wide Web Electronic Publication. 2020. Available online: www.fishbase.org (accessed on 15 January 2020).
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.A.; Peterson, A.T.; Soberon, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Peterson, A.T.; Pepes, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modelling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- O’Brien, R.M. A caution regarding rules of thumb for variance inflation factors. Qual. Quant. 2007, 41, 673–690. [Google Scholar] [CrossRef]
- Warren, D.L.; Matzke, N.; Cardillo, M.; Baumgartner, J.; Beaumont, L.; Huron, N.; Simões, M.; Iglesias, T.L.; Dinnage, R. ENMTools (Software Package). 2019. Available online: https://github.com/danlwarren/ENMTools (accessed on 8 March 2020).
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why input settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2007, 17, 145–151. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A. Insights into the area under the receiver operating characteristic curve (AUC) as a discrimination measure in species distribution modelling. Glob. Ecol. Biogeogr. 2012, 21, 498–507. [Google Scholar] [CrossRef]
- Leroy, B.; Delsol, R.; Hugueny, B.; Meynard, C.N.; Barhoumi, C.; Barbet-Massin, M.; Bellard, C. Witho ut quality pres-ence–absence data, discrimination metrics such as TSS can be misleading measures of model performance. J. Biogeogr. 2018, 45, 1–9. [Google Scholar] [CrossRef]
- Anderson, R.P.; Lew, D.; Peterson, A.T. Evaluating predictive models of species’ distributions: Criteria for selecting optimal models. Ecol. Model. 2003, 162, 211–232. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Le Lay, G.; Helfer, V.; Randin, C.; Guisan, A. Evaluating the ability of habitat suitability models to predict species presence. Ecol. Model. 2006, 199, 142–152. [Google Scholar] [CrossRef]
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Myers, N. Threatened biotas: ‘Hotspots’ in tropical forests. Environmentalist 1988, 8, 187–208. [Google Scholar] [CrossRef]
- Cushman, S.A.; Elliot, N.B.; Bauer, D.; Kesch, K.; Bahaa-el-din, L.; Bothwell, H.; Flyman, M.; Mtare, G.; Macdonald, D.W.; Loveridge, A.J. Prioritizing core areas, corridors and conflict hotspots for lion conservation in southern Africa. PLoS ONE 2018, 13, e0196213. [Google Scholar] [CrossRef]
- Trujillo, A.P.; Thurman, H.V. Essentials of Oceanography, 12th ed.; Pearson Education, Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Wang, L.; Kerr, L.A.; Record, N.R.; Bridger, E.; Tupper, B.; Mills, K.E.; Armstrong, E.M.; Pershing, A.J. Modeling marine pelagic fish species spatiotemporal distributions utilizing a maximum entropy approach. Fish. Oceanogr. 2018, 27, 571–586. [Google Scholar] [CrossRef]
- Leathwick, J.R.; Elith, J.; Francis, M.P.; Hastie, T.; Taylor, P. Variation in demersal fish species richness in the oceans surrounding New Zealand: An analysis using boosted regression trees. Mar. Ecol. Prog. Ser. 2006, 321, 267–281. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.; Drazen, J.C.; Radford, B.T.; Kelley, C.; Newman, S.J. Improving essential fish habitat designation to support sustainable ecosystem-based fisheries management. Mar. Policy 2016, 69, 32–41. [Google Scholar] [CrossRef]
- Hogarth, P.J. The Biology of Mangroves and Seagrasses; Oxford University Press: Oxford, UK, 2015; p. 389. [Google Scholar]
- Little, C.; Williams, G.A.; Trowbridge, C.D. The Biology of Rocky Shores; Oxford University Press: Oxford, UK, 2009; p. 356. [Google Scholar]
- Verfaillie, E.; Van-Lancker, V.; Van-Meirvenne, M. Multivariate geostatistics for the predictive modelling of the surficial sand distribution in shelf seas. Cont. Shelf Res. 2006, 26, 2454–2468. [Google Scholar] [CrossRef]
- Haris, K.; Chakraborthy, B.; Ingole, B.; Menezes, A.; Srivastava, R. Seabed habitat mapping employing single and multi-beam backscatter data: A case study from the western continental shelf of India. Cont. Shelf Res. 2012, 48, 40–49. [Google Scholar] [CrossRef]
- Mora, C. Ecology of Fishes on Coral Reef; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marine Conservation Areas | Official Decree | Management Plan | Hotspot (Hectares) |
|---|---|---|---|
| BNPA(BZ) | Yes | Yes | 408 |
| BNPA(CA) | Yes | Yes | 309 |
| HBRS | No * | No | 402 |
| HEM-ELP-RS | No * | No | 1249 |
| BCIAELP | No | No | 1219 |
| PMRICBCS | No | No | 1778 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urcádiz-Cázares, F.J.; Cruz-Escalona, V.H.; Peterson, M.S.; Aguilar-Medrano, R.; Marín-Enríquez, E.; González-Peláez, S.S.; Del Pino-Machado, A.; Enríquez-García, A.B.; Borges-Souza, J.M.; Ortega-Rubio, A. Linking Habitat and Associated Abiotic Conditions to Predict Fish Hotspots Distribution Areas within La Paz Bay: Evaluating Marine Conservation Areas. Diversity 2021, 13, 212. https://0-doi-org.brum.beds.ac.uk/10.3390/d13050212
Urcádiz-Cázares FJ, Cruz-Escalona VH, Peterson MS, Aguilar-Medrano R, Marín-Enríquez E, González-Peláez SS, Del Pino-Machado A, Enríquez-García AB, Borges-Souza JM, Ortega-Rubio A. Linking Habitat and Associated Abiotic Conditions to Predict Fish Hotspots Distribution Areas within La Paz Bay: Evaluating Marine Conservation Areas. Diversity. 2021; 13(5):212. https://0-doi-org.brum.beds.ac.uk/10.3390/d13050212
Chicago/Turabian StyleUrcádiz-Cázares, Francisco Javier, Víctor Hugo Cruz-Escalona, Mark S. Peterson, Rosalía Aguilar-Medrano, Emigdio Marín-Enríquez, Sergio Scarry González-Peláez, Arturo Del Pino-Machado, Arturo Bell Enríquez-García, José Manuel Borges-Souza, and Alfredo Ortega-Rubio. 2021. "Linking Habitat and Associated Abiotic Conditions to Predict Fish Hotspots Distribution Areas within La Paz Bay: Evaluating Marine Conservation Areas" Diversity 13, no. 5: 212. https://0-doi-org.brum.beds.ac.uk/10.3390/d13050212