
Genetic Diversity and Population Structure of Two Endangered Neotropical Parrots Inform In Situ and Ex Situ Conservation Strategies

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
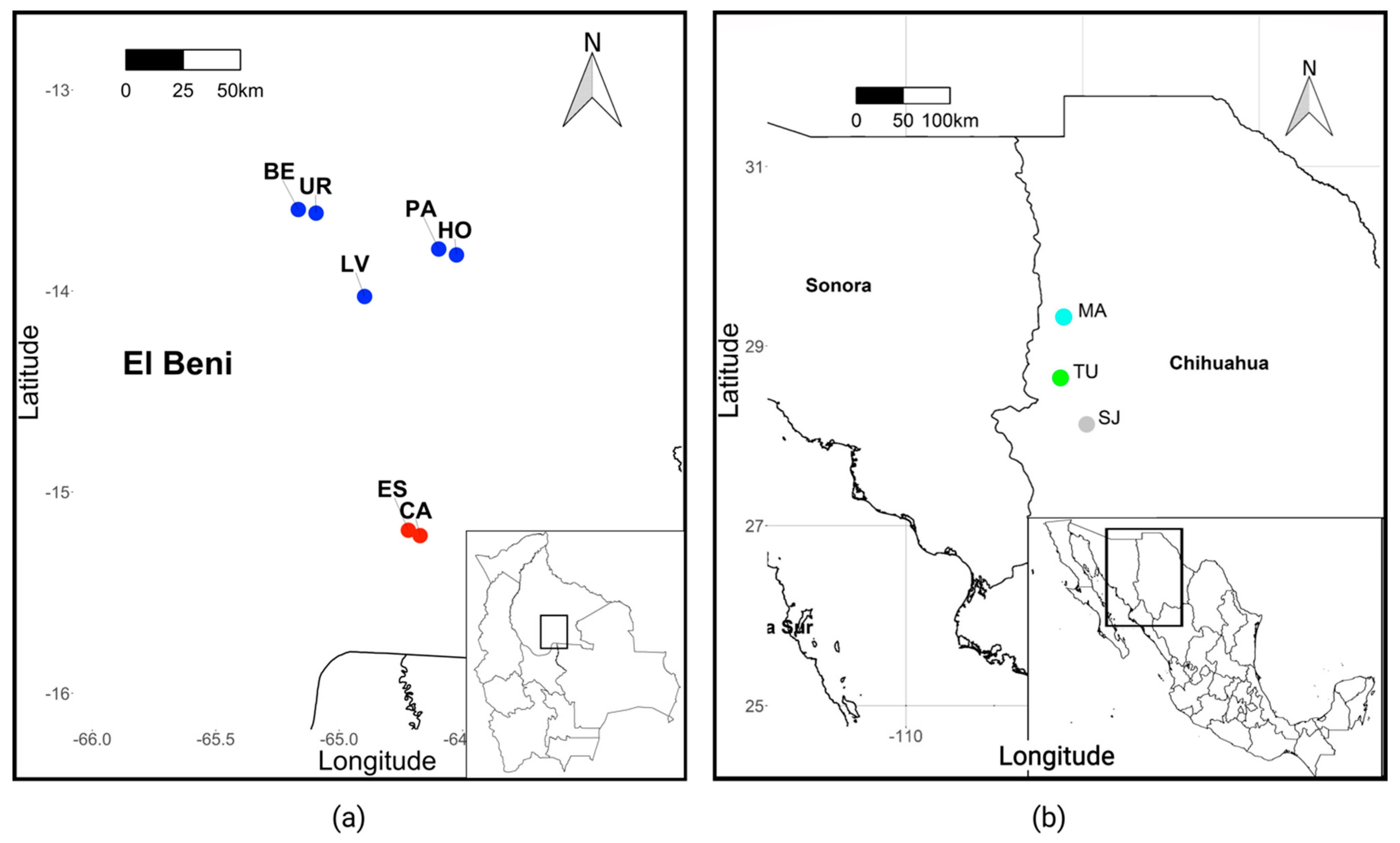
2.1. Sample Collection
2.2. DNA Extraction
2.3. Microsatellite Genotyping
2.4. Population Structure and Genetic Diversity
2.5. Demographic History
3. Results
3.1. Microsatellite Validation
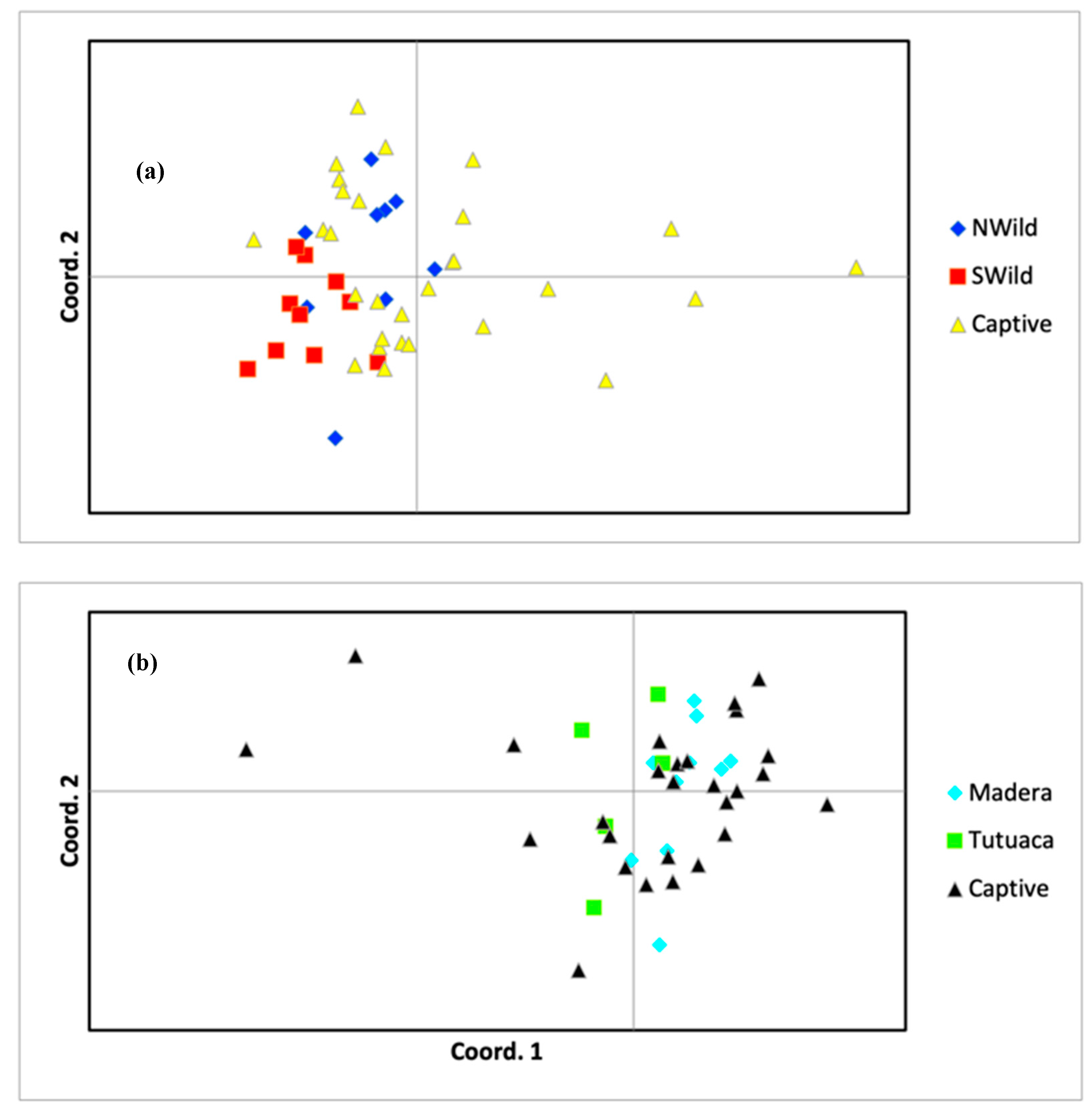
3.2. Population Structure
3.3. Genetic Diversity of Captive and Wild Populations
3.4. Demographic History
4. Discussion
4.1. Population Structure
4.2. Genetic Diversity
4.3. Demographic History
4.4. Conservation Implications
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Palstra, F.; Ruzzante, D. Genetic estimates of contemporary effective population size: What can they tell us about the importance of genetic stochasticity for wild population persistence? Mol. Ecol. 2008, 17, 3428–3447. [Google Scholar] [CrossRef]
- Witzenberger, K.A.; Hochkirch, A. Ex situ conservation genetics: A review of molecular studies on the genetic consequences of captive breeding programmes for endangered animal species. Biodiv. Conserv. 2011, 20, 1843–1861. [Google Scholar] [CrossRef]
- Haig, S.M.; Ballou, J.D.; Derrickson, S.R. Management options for preserving genetic diversity: Reintroduction of guam rails to the wild. Conserv. Biol. 1990, 4, 290–300. [Google Scholar] [CrossRef]
- Seddon, P.; Armstrong, D.; Maloney, R. Developing the science of reintroduction biology. Conserv. Biol. 2007, 21, 303–312. [Google Scholar] [CrossRef]
- Hedrick, P.W.; Garcia-Dorado, A. Understanding inbreeding depression, purging, and genetic rescue. Trends Ecol. Evol. 2016, 31, 940–952. [Google Scholar] [CrossRef]
- Briskie, J.V.; Mackintosh, M. Hatching failure increases with severity of population bottlenecks in birds. Proc. Natl. Acad. Sci. USA 2004, 101, 558–561. [Google Scholar] [CrossRef] [Green Version]
- Frankham, R.; Hendry, H.; Margan, S.; Briscoe, D. Does equalization of family sizes reduce genetic adaption to captivity? Anim. Conserv. 2006, 3, 357–363. [Google Scholar] [CrossRef]
- Milián-García, Y.; Jensen, E.L.; Madsen, J.; Álvarez Alonso, S.; Serrano Rodríguez, A.; Espinosa López, G.; Russello, M.A. Founded: Genetic reconstruction of lineage diversity and kinship informs ex situ conservation of cuban amazon parrots (Amazona leucocephala). J. Hered. 2015, 106, 573–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkunsky, I.; Daniele, G.; Kacoliris, F.P.; Díaz-Luque, J.A.; Silva Frias, C.P.; Aramburu, R.M.; Gilardi, J.D. Reproductive parameters in the critically endangered blue-throated macaw: Limits to the recovery of a parrot under intensive management. PLoS ONE 2014, 9, e99941. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; By, R. Poaching, habitat loss and the decline of neotropical parrots: A comparative spatial analysis. J. Exp. Criminol. 2013, 9, 333–353. [Google Scholar] [CrossRef]
- Collar, N.J. Globally threatened parrots: Criteria, characteristics and cures. Int. Zoo Yearb. 2000, 37, 21–35. [Google Scholar] [CrossRef]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org (accessed on 18 July 2021).
- Berkunsky, I.; Quillfeldt, P.; Brightsmith, D.J.; Abbud, M.C.; Aguilar, J.M.R.E.; Alemán-Zelaya, U.; Aramburú, R.M.; Arce-Arias, A.; Balas-McNab, R.; Balsby, T.J.S.; et al. Current threats faced by Neotropical parrot populations. Biol. Conserv. 2017, 214, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Wright, T.; Toft, C.; Enkerlin-Hoeflich, E.; Gonzalez-Elizondo, J.; Albornoz, M.; Rodríguez-Ferraro, A.; Rojas-Suárez, F.; Sanz D’Angelo, V.; Trujillo, A.; Beissinger, S.; et al. Nest poaching in neotropical parrots. Conserv. Biol. 2001, 15, 710–720. [Google Scholar] [CrossRef]
- Watson, R. International Studbook: Annual Report and Recommendations for 2011, Spix’s Macaw (Cyanopsitta spixii); Al Wabra Wildlife Preservation: Al Shahaniya, Qatar, 2011. [Google Scholar]
- BirdLife International. Cyanopsitta spixii. Available online: http://datazone.birdlife.org/species/factsheet/spixs-macaw-cyanopsitta-spixii (accessed on 18 July 2021).
- Ballou, J.D.; Lacy, R.C. Identifying genetically important individuals for management of genetic diversity in captive populations. In Population Management for Survival and Recovery; Ballou, J.D., Gilpin, M., Foose, T.J., Eds.; Columbia University Press: New York, NY, USA, 1995; pp. 76–111. [Google Scholar]
- Jamieson, I.G. Founder effects, inbreeding, and loss of genetic diversity in four avian reintroduction programs. Conserv. Biol. 2011, 25, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Lutaaya, B.E.; Misztal, I.; Bertrand, J.K.; Mabry, J.W. Inbreeding in populations with incomplete pedigrees. J. Anim. Breed. Genet. 1999, 116, 475–480. [Google Scholar] [CrossRef]
- Collar, N.J. Parrot reintroduction: Towards a synthesis of best practice. In Proceedings of the 6th International Parrot Convention, Loro Parque, Puerto de la Cruz, Tenerife, Spain, 27–30 September 2006; pp. 82–107. [Google Scholar]
- Ebenhard, T. Conservation breeding as a tool for saving animal species from extinction. Trends Ecol. Evol. 1995, 10, 438–443. [Google Scholar] [CrossRef]
- Viggers, K.L.; Lindenmayer, D.; Spratt, D. The importance of disease in reintroduction programmes. Wildl. Res. 1993, 20, 687–698. [Google Scholar] [CrossRef]
- McPhee, M. Generations in captivity increases behavioral variance: Considerations for captive breeding and reintroduction programs. Biol. Conserv. 2004, 115, 71–77. [Google Scholar] [CrossRef]
- Morrison, C.E.; Johnson, R.N.; Grueber, C.E.; Hogg, C.J. Genetic impacts of conservation management actions in a critically endangered parrot species. Conserv. Genet. 2020, 21, 869–877. [Google Scholar] [CrossRef]
- Coleman, R.; Weeks, A. Population Genetics of the Endangered Orange-Bellied Parrot (Neophema chrysogaster): Project Update; University of Melbourne: Melbourne, Australia, 2013. [Google Scholar]
- Snyder, N.F.R.; Koenig, S.E.; Koschmann, J.; Snyder, H.A.; Johnson, T.B. Thick-billed parrot releases in Arizona. Condor 1994, 96, 845–862. [Google Scholar] [CrossRef]
- White, T.H.; Collar, N.J.; Moorhouse, R.J.; Sanz, V.; Stolen, E.D.; Brightsmith, D.J. Psittacine reintroductions: Common denominators of success. Biol. Conserv. 2012, 148, 106–115. [Google Scholar] [CrossRef]
- BirdLife International. Ara Glaucogularis. Available online: https://dx.doi.org/10.2305/IUCN.UK.2018-2.RLTS.T22685542A130868462.en (accessed on 1 June 2021).
- Herzog, S.K.; Maillard, Z.O.; Boorsma, T.; Sánchez-Ávila, G.; García-Solíz, V.H.; Paca-Condori, A.C.; De Abajo, M.V.; Soria-Auza, R.W. First systematic sampling approach to estimating the global population size of the critically endangered blue-throated macaw Ara glaucogularis. Bird Conserv. Int. 2021, 31, 293–311. [Google Scholar] [CrossRef]
- Anderson, G. North American Regional Studbook for the Blue-Throated Macaw (Ara glaucogularis); St. Augustine Alligator Farm Zoological Park: St. Augustine, FL, USA, 2017. [Google Scholar]
- Maestri, M.L.; Ferrati, R.; Berkunsky, I. Evaluating management strategies in the conservation of the critically endangered Blue-throated Macaw (Ara glaucogularis). Ecol. Model. 2017, 361, 74–79. [Google Scholar] [CrossRef]
- Strem, R.; Bouzat, J. Population viability analysis of the blue-throated macaw (Ara glaucogularis) using individual-based and cohort-based PVA programs. Open Conserv. Biol. J. 2012, 6, 12–24. [Google Scholar] [CrossRef] [Green Version]
- BirdLife International. Rhynchopsitta Pachyrhyncha (Amended Version of 2016 Assessment). Available online: https://0-dx-doi-org.brum.beds.ac.uk/10.2305/IUCN.UK.2017-1.RLTS.T22685766A110475642.en (accessed on 1 June 2021).
- U.S. Fish and Wildlife Service. Thick-Billed Parrot (Rhynchopsitta pachyrhyncha) Recovery Plan Addendum; U.S. Fish and Wildlife Service, Southwest Region: Albuquerque, NM, USA, 2013.
- Putnam, A. Thick-Billed Parrot (Rhynchopsitta pachyrhyncha) AZA Species Survival Plan; San Diego Zoo Global: San Diego, CA, USA, 2014. [Google Scholar]
- Guerra, J.E.; Cruz-Nieto, J.; Ortiz-Maciel, S.G.; Wright, T.F. Limited geographic variation in the vocalizations of the endangered thick-billed parrot: Implications for conservation strategies. Condor 2008, 110, 639–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monterrubio-Rico, T.C.; Enkerlin-Hoeflich, E. Present use and characteristics of thick-billed parrot nest sites in northwestern Mexico. J. Field Ornithol. 2004, 75, 96–103. [Google Scholar] [CrossRef]
- Snyder, N.F.; Enkerlin-Hoeflich, E.C.; Cruz-Nieto, M.A.; Valdes-Peña, R.A.; Ortiz-Maciel, S.G.; Cruz-Nieto, J. Thick-billed parrot (rhynchopsitta pachyrhyncha). In Birds of the World, 1st ed.; Billerman, S.M., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Sheppard, J.K.; Rojas, J.I.G.; Cruz, J.; GonzÁLez, L.F.T.; Nieto, M.Á.C.; Lezama, S.D.J.; Juarez, E.A.; Lamberski, N. Predation of nesting thick-billed parrots Rhychopsitta pachyrhyncha by bobcats in northwestern Mexico. Bird Conserv. Int. 2020, 1–9. [Google Scholar] [CrossRef]
- Monterrubio-Rico, T.C.; Charre-Medellin, J.F.; Sáenz-Romero, C. Current and future habitat availability for thick-billed and maroon-fronted parrots in northern Mexican forests. J. Field Ornithol. 2015, 86, 1–16. [Google Scholar] [CrossRef]
- Frankham, R. Inbreeding and extinction: A threshold effect. Conserv. Biol. 1995, 9, 792–799. [Google Scholar] [CrossRef]
- GE Healthcare Life Sciences. Whatman™ FTA Elute™. Available online: https://at.vwr.com/assetsvc/asset/de_AT/id/17887357/contents (accessed on 20 September 2017).
- Olah, G.; Heinsohn, R.G.; Espinoza, J.R.; Brightsmith, D.J.; Peakall, R. An evaluation of primers for microsatellite markers in scarlet macaw (Ara macao) and their performance in a Peruvian wild population. Conserv. Genet. Resour. 2015, 7, 157–159. [Google Scholar] [CrossRef]
- Caparroz, R.; Miyaki, C.Y.; Baker, A.J. Characterization of microsatellite loci in the blue-and-gold macaw, Ara ararauna (Psittaciformes: Aves). Mol. Ecol. Notes 2003, 3, 441–443. [Google Scholar] [CrossRef]
- Russello, M.A.; Saranathan, V.; Buhrman-Deever, S.; Eberhard, J.; Caccone, A. Characterization of polymorphic microsatellite loci for the invasive monk parakeet (Myiopsitta monachus). Mol. Ecol. Notes 2007, 7, 990–992. [Google Scholar] [CrossRef]
- Klauke, N.; Masello, J.F.; Quillfeldt, P.; Segelbacher, G. Isolation of tetranucleotide microsatellite loci in the burrowing parrot (Cyanoliseus patagonus). J. Ornithol. 2009, 150, 921–924. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.; Amato, G. Thick-Billed Parrot Primers. 2009; unpublished. [Google Scholar]
- Acosta, D. Development and Characterization of Microsatellite Markers for the Endangered Thick-Billed Parrot (Rhynchopsitta pachyrhyncha); New Mexico State University: Las Cruces, NM, USA, 2009. [Google Scholar]
- Van Oosterhout, C.; Hutchinson, W.; Wills, D.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Sinnwell, J.P.; Therneau, T.M.; Schaid, D.J. The kinship2 R package for pedigree data. Hum. Hered. 2014, 78, 91–93. [Google Scholar] [CrossRef] [Green Version]
- Kalinowski, S.; Wagner, A.; Taper, M. ML-RELATE: A computer program for maximum likelihood estimation of relatedness and relationship. Mol. Ecol. Notes 2006, 6, 576–579. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (version-1.2)—Population-genetics software for exact tests and ecumenicsim. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Earl, D.; Vonholdt, B. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, J.; Mj, S.; Donnelly, P.J. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, M.; Rosenberg, N. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, N. DISTRUCT: A program for the graphical display of population structure. Mol. Ecol. Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P. GenAlEx 6.5: Genetic analysis in excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paetkau, D.; Slade, R.; Burden, M.; Estoup, A. Genetic assignment methods for the direct, real-time estimation of migration rate: A simulation-based exploration of accuracy and power. Mol. Ecol. 2004, 13, 55–65. [Google Scholar] [CrossRef]
- Piry, S.; Alapetite, A.; Cornuet, J.; Paetkau, D.; Baudouin, L.; Estoup, A. GENECLASS2: A software for genetic assignment and first-generation migrant detection. J. Hered. 2004, 95, 536–539. [Google Scholar] [CrossRef]
- Goudet, J. hierfstat, a package for R to compute and test hierarchical F-statistics. Mol. Ecol. Notes 2005, 5, 184–186. [Google Scholar] [CrossRef] [Green Version]
- Cornuet, J.M.; Luikart, G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 1996, 144, 2001–2014. [Google Scholar] [CrossRef]
- Piry, S.; Luikart, G.; Cornuet, J.M. BOTTLENECK: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Beck, N.; Double, M.; Cockburn, A. Microsatellite evolution at two hypervariable loci revealed by extensive avian pedigrees. Mol. Biol. Evol. 2003, 20, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.; Haig, S.; Mullins, T.; Popper, K.; Green, M. Evidence for population bottlenecks and subtle genetic structure in the yellow rail. Condor 2012, 114, 100–112. [Google Scholar] [CrossRef]
- Vergara-Tabares, D.L.; Cordier, J.M.; Landi, M.A.; Olah, G.; Nori, J. Global trends of habitat destruction and consequences for parrot conservation. Glob. Chang. Biol. 2020, 26, 4251–4262. [Google Scholar] [CrossRef]
- Russello, M.A.; Smith-Vidaurre, G.; Wright, T.F. Genetics of invasive parrot populations. In Naturalized Parrots of the World: Distribution, Ecology, and Impacts of the World’s Most Colorful Colonizers; Pruitt-Jones, S., Ed.; Princeton University Press: Princeton, NJ, USA, 2021. [Google Scholar]
- Klauke, N.; Schaefer, H.M.; Bauer, M.; Segelbacher, G. Limited dispersal and significant fine - scale genetic structure in a tropical montane parrot species. PLoS ONE 2016, 11, e0169165. [Google Scholar] [CrossRef] [Green Version]
- Blanco, G.; Morinha, F.; Roques, S.; Hiraldo, F.; Rojas, A.; Tella, J.L. Fine-scale genetic structure in the critically endangered red-fronted macaw in the absence of geographic and ecological barriers. Sci. Rep. 2021, 11, 556. [Google Scholar] [CrossRef]
- Raisin, C.; Frantz, A.C.; Kundu, S.; Greenwood, A.G.; Jones, C.G.; Zuel, N.; Groombridge, J.J. Genetic consequences of intensive conservation management for the Mauritius parakeet. Conserv. Genet. 2012, 13, 707–715. [Google Scholar] [CrossRef]
- Tollington, S.; Jones, C.G.; Greenwood, A.; Tatayah, V.; Raisin, C.; Burke, T.; Dawson, D.A.; Groombridge, J.J. Long-term, fine-scale temporal patterns of genetic diversity in the restored Mauritius parakeet reveal genetic impacts of management and associated demographic effects on reintroduction programmes. Biol. Conserv. 2013, 161, 28–38. [Google Scholar] [CrossRef]
- Lacy, R.; Petric, A.; Warneke, M. Inbreeding and outbreeding depression in captive populations of wild animals. In The Natural History of Inbreeding and Outbreeding: Theoretical and Empirical Perspectives; Thornhill, N.W., Ed.; University of Chicago Press: Chicago, IL, USA, 1993. [Google Scholar]
- Lande, R. Anthropogenic, ecological and genetic factors in extinction and conservation. Popul Ecol. 1998, 40, 259–269. [Google Scholar] [CrossRef]
- Stojanovic, D.; Olah, G.; Webb, M.; Peakall, R.; Heinsohn, R. Genetic evidence confirms severe extinction risk for critically endangered swift parrots: Implications for conservation management. Anim. Conserv. 2018, 21, 313–323. [Google Scholar] [CrossRef]
- BirdLife International. Lathamus Discolor. Available online: https://0-dx-doi-org.brum.beds.ac.uk/10.2305/IUCN.UK.2018-2.RLTS.T22685219A130886700.en (accessed on 1 June 2021).
- BirdLife International. Amazona Leucocephala. Available online: https://0-dx-doi-org.brum.beds.ac.uk/10.2305/IUCN.UK.2020-3.RLTS.T22686201A179212864.en (accessed on 1 June 2021).
- Bergner, L.M.; Jamieson, I.G.; Robertson, B.C. Combining genetic data to identify relatedness among founders in a genetically depauperate parrot, the Kakapo (Strigops habroptilus). Conserv. Genet. 2014, 15, 1013–1020. [Google Scholar] [CrossRef]
- BirdLife International. Strigops Habroptila. Available online: https://0-dx-doi-org.brum.beds.ac.uk/10.2305/IUCN.UK.2018-2.RLTS.T22685245A129751169.en (accessed on 1 June 2021).
- Yamashita, C.; Barros, Y.M. The Blue-throated Macaw (Ara glaucogularis), characterization of its distinctive habitat in the savannas of Beni, Bolivia. Ararajuba 1997, 5, 141–150. [Google Scholar]
- Clout, M.N.; Merton, D.V. Saving the kakapo: The conservation of the world’s most peculiar parrot. Bird Conserv. Int. 1998, 8, 281–296. [Google Scholar] [CrossRef] [Green Version]
- Collar, N.; Sharpe, C.J.; Boesman, P.F.D. Kakapo (Strigops habroptila). In Birds of the World, 1st ed.; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- White, K.L.; Eason, D.K.; Jamieson, I.G.; Robertson, B.C. Evidence of inbreeding depression in the critically endangered parrot, the kakapo. Anim. Conserv. 2015, 18, 341–347. [Google Scholar] [CrossRef]
- Beissinger, S.R.; Wunderle, J.M.; Meyers, J.M.; Sæther, B.E.; Engen, S. Anatomy of a bottleneck: Diagnosing factors limiting population growth in the Puerto Rican parrot. Ecol. Monogr. 2008, 78, 185–203. [Google Scholar] [CrossRef] [Green Version]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Shields, W.M. The natural and unnatural history of inbreeding and outbreeding. In The Natural History of Inbreeding and Outbreeding; Thornhill, N.W., Ed.; University of Chicago Press: Chicago, IL, USA, 1993; pp. 143–169. [Google Scholar]
- Edmands, S. Between a rock and a hard place: Evaluating the relative risks of inbreeding and outbreeding for conservation and management. Mol. Ecol. 2007, 16, 463–475. [Google Scholar] [CrossRef]
- Marr, A.B.; Keller, L.F.; Arcese, P. Heterosis and outbreeding depression in descendants of natural immigrants to an inbred population of song sparrows (Melospiza melodia). Evolution 2002, 56, 131–142. [Google Scholar] [CrossRef]
- Herzog, S.; Bürger, J.; Troncoso, A.; Vargas-Rodriguez, R.; Boorsma, T.; Soria-Auza, R.W. Deterministic population growth models and conservation translocation as a management strategy for the critically endangered blue-throated macaw (Ara glaucogularis): A critique of Maestri et al. Ecol. Model. 2018, 388, 145–148. [Google Scholar] [CrossRef]
- IUCN/SSP. Guidelines for Reintroductions and Other Conservation Translocations; Version 1.0; IUCN Species Survival Commission: Gland, Switzerland, 2013; p. 57. [Google Scholar]



{kind=link}
{kind=link}
| Blue-Throated Macaw | Thick-Billed Parrot | ||||||
|---|---|---|---|---|---|---|---|
| Population | Captive | North Wild | South Wild | Population | Captive | Madera | Tutuaca |
| Captive | - | Captive | - | ||||
| North Wild | 0.026 | - | Madera | 0.017 | - | ||
| South Wild | 0.047 ** | 0.048 ** | - | Tutuaca | 0.045 | 0.053 | - |
| Whole Wild | 0.025 * | NA | NA | Whole Wild | 0.014 | NA | NA |
| Blue-Throated Macaw | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Locus | Captive | North Wild | South Wild | Whole Wild | ||||||||||||||||
| N | Na | Ho | He | F | N | Na | Ho | He | F | N | Na | Ho | He | F | N | Na | Ho | He | F | |
| SCMA09 [25] | 29 | 2 | 0.034 | 0.034 | −0.018 | 9 | 1 | 0 | 0 | NA | 10 | 2 | 0.200 | 0.180 | −0.111 | 19 | 2 | 0.105 | 0.100 | −0.056 |
| SCMA46 [25] | 27 | 9 | 0.852 | 0.846 | −0.007 | 9 | 7 | 0.889 | 0.821 | −0.083 | 10 | 5 | 0.700 | 0.725 | −0.034 | 19 | 7 | 0.789 | 0.812 | 0.027 |
| SCMA19 [25] | 23 | 9 | 0.609 | 0.635 | 0.026 | 9 | 5 | 0.667 | 0.728 | 0.085 | 10 | 2 | 0.500 | 0.455 | −0.099 | 19 | 5 | 0.579 | 0.614 | 0.056 |
| SCMA31 [25] | 25 | 8 | 0.800 | 0.814 | 0.017 | 9 | 7 | 0.778 | 0.815 | 0.045 | 10 | 6 | 0.800 | 0.740 | −0.081 | 19 | 8 | 0.789 | 0.812 | 0.027 |
| SCMA41 [25] | 28 | 10 | 0.750 | 0.839 | 0.106 | 9 | 7 | 0.778 | 0.765 | −0.016 | 10 | 8 | 1.000 | 0.820 | −0.220 | 19 | 10 | 0.895 | 0.820 | −0.091 |
| SCMA27 [25] | 29 | 6 | 0.759 | 0.707 | −0.072 | 9 | 4 | 0.444 | 0.648 | 0.314 | 10 | 2 | 0.700 | 0.455 | −0.538 | 19 | 4 | 0.579 | 0.611 | 0.052 |
| SCMA11 [25] | 28 | 2 | 0.179 | 0.219 | 0.184 | 9 | 2 | 0.111 | 0.105 | −0.059 | 10 | 2 | 0.400 | 0.320 | −0.250 | 19 | 2 | 0.263 | 0.229 | −0.152 |
| SCMA22 [25] | 28 | 5 | 0.679 | 0.668 | −0.015 | 9 | 2 | 0.444 | 0.444 | 0.000 | 10 | 2 | 0.500 | 0.455 | −0.099 | 19 | 2 | 0.474 | 0.450 | −0.052 |
| SCMA26 [25] | 29 | 10 | 0.724 | 0.825 | 0.122 | 9 | 5 | 0.889 | 0.735 | −0.210 | 10 | 8 | 0.900 | 0.805 | −0.118 | 19 | 9 | 0.895 | 0.817 | −0.095 |
| SCMA02 [25] | 25 | 7 | 0.800 | 0.731 | −0.094 | 9 | 5 | 0.778 | 0.716 | 0.086 | 10 | 4 | 0.900 | 0.655 | −0.374 | 19 | 6 | 0.842 | 0.720 | −0.169 |
| Mean (SE) | 27 | 7 | 0.618 (0.089) | 0.631 (0.088) | 0.025 (0.028) | 9 | 4.5 | 0.578 (0.100) | 0.578 (0.094) | −0.001 (0.046) | 10 | 4.1 | 0.660 (0.081) | 0.561 (0.069) | −0.186 (0.053) | 19 | 5.5 | 0.621 (0.087) | 0.598 (0.082) | −0.045 (0.026) |
| Thick-billed parrot | ||||||||||||||||||||
| Locus | Captive | Madera | Tutuaca | Whole Wild | ||||||||||||||||
| N | Na | Ho | He | F | N | Na | Ho | He | F | N | Na | Ho | He | F | N | Na | Ho | He | F | |
| UnaCT74 [26] | 25 | 3 | 0.120 | 0.114 | −0.049 | 10 | 2 | 0.100 | 0.095 | −0.053 | 5 | 3 | 0.200 | 0.460 | 0.565 | 15 | 3 | 0.133 | 0.240 | 0.444 |
| UnaCT21 [26] | 26 | 3 | 0.500 | 0.536 | 0.066 | 10 | 3 | 0.700 | 0.505 | −0.386 | 5 | 2 | 0.400 | 0.320 | −0.250 | 15 | 3 | 0.600 | 0.464 | −0.292 |
| UnaCT55 [26] | 27 | 8 | 0.815 | 0.767 | −0.063 | 10 | 7 | 0.700 | 0.700 | 0.000 | 5 | 5 | 0.800 | 0.680 | −0.176 | 15 | 7 | 0.733 | 0.702 | −0.044 |
| UnaCT43 [26] | 25 | 2 | 0.120 | 0.114 | −0.064 | 10 | 2 | 0.100 | 0.095 | −0.053 | 5 | 1 | 0.000 | 0.000 | NA | 15 | 2 | 0.067 | 0.064 | −0.034 |
| TBP2−39 [29] | 27 | 10 | 0.778 | 0.684 | −0.136 | 10 | 7 | 0.900 | 0.785 | −0.146 | 5 | 6 | 1.000 | 0.800 | −0.250 | 15 | 10 | 0.933 | 0.856 | −0.091 |
| TBP2−61 [29] | 25 | 4 | 0.640 | 0.578 | −0.107 | 10 | 4 | 0.900 | 0.625 | −0.440 | 5 | 3 | 0.600 | 0.460 | −0.304 | 15 | 5 | 0.800 | 0.616 | −0.300 |
| Rhpac149 [30] | 26 | 5 | 0.654 | 0.604 | −0.082 | 10 | 5 | 0.600 | 0.700 | 0.143 | 5 | 4 | 0.800 | 0.700 | −0.143 | 15 | 5 | 0.667 | 0.722 | 0.077 |
| Rhpac074 [30] | 25 | 8 | 0.800 | 0.766 | −0.044 | 10 | 5 | 0.900 | 0.690 | −0.304 | 5 | 5 | 0.800 | 0.680 | −0.176 | 15 | 7 | 0.867 | 0.744 | −0.164 |
| MmGT057 [27] | 26 | 3 | 0.500 | 0.452 | −0.106 | 10 | 2 | 0.500 | 0.495 | −0.010 | 5 | 2 | 0.400 | 0.480 | 0.167 | 15 | 2 | 0.467 | 0.491 | 0.050 |
| CyanP05 [28] | 26 | 7 | 0.808 | 0.794 | −0.017 | 10 | 5 | 0.700 | 0.765 | 0.085 | 5 | 5 | 0.600 | 0.680 | 0.118 | 15 | 5 | 0.667 | 0.771 | 0.135 |
| Mean (SE) | 26 | 5.3 | 0.573 (0.084) | 0.541 (0.079) | −0.060 (0.018) | 10 | 4.2 | 0.610 (0.095) | 0.546 (0.081) | −0.116 (0.063) | 5 | 3.6 | 0.560 (0.098) | 0.526 (0.075) | −0.050 (0.089) | 15 | 4.9 | 0.593 (0.092) | 0.567 (0.080) | 0.022 (0.069) |
| Blue-Throated Macaw: (Left) Southern and (Right) Northern Population | ||||||
|---|---|---|---|---|---|---|
| Stepwise Mutation Model | Two-Phase Model | Stepwise Mutation Model | Two-Phase Model | |||
| 60% Stepwise | 80% Stepwise | 60% Stepwise | 80% Stepwise | |||
| Sign test | 0.158 | 0.153 | 0.176 | 0.535 | 0.074 | 0.092 |
| Wilcoxon signed rank test (1-tail) | 0.285 | 0.082 | 0.125 | 0.161 | 0.009* | 0.016 * |
| Wilcoxon signed rank test (2-tail) | 0.570 | 0.164 | 0.250 | 0.322 | 0.018* | 0.032 * |
| Thick-billed parrot: whole wild population | ||||||
| Stepwise Mutation Model | Two-Phase Model | |||||
| 60% Stepwise | 80% Stepwise | |||||
| Sign test | 0.193 | 0.449 | 0.428 | |||
| Wilcoxon signed rank test (1-tail) | 0.838 | 0.422 | 0.577 | |||
| Wilcoxon signed rank test (2-tail) | 0.375 | 0.845 | 0.921 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campos, C.I.; Martinez, M.A.; Acosta, D.; Diaz-Luque, J.A.; Berkunsky, I.; Lamberski, N.L.; Cruz-Nieto, J.; Russello, M.A.; Wright, T.F. Genetic Diversity and Population Structure of Two Endangered Neotropical Parrots Inform In Situ and Ex Situ Conservation Strategies. Diversity 2021, 13, 386. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080386
Campos CI, Martinez MA, Acosta D, Diaz-Luque JA, Berkunsky I, Lamberski NL, Cruz-Nieto J, Russello MA, Wright TF. Genetic Diversity and Population Structure of Two Endangered Neotropical Parrots Inform In Situ and Ex Situ Conservation Strategies. Diversity. 2021; 13(8):386. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080386
Chicago/Turabian StyleCampos, Carlos I., Melinda A. Martinez, Daniel Acosta, Jose A. Diaz-Luque, Igor Berkunsky, Nadine L. Lamberski, Javier Cruz-Nieto, Michael A. Russello, and Timothy F. Wright. 2021. "Genetic Diversity and Population Structure of Two Endangered Neotropical Parrots Inform In Situ and Ex Situ Conservation Strategies" Diversity 13, no. 8: 386. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080386