
Physiological and Biochemical Characterization of Isolated Bacteria from a Coccolithophore Chrysotila dentata (Prymnesiophyceae) Culture


1
Research Centre for Indian Ocean Ecosystem, Tianjin University of Science and Technology, Tianjin 300457, China
2
Institute of Marine Science and Technology, Shandong University, Qingdao 266237, China
3
College of Marine Science and Technology, China University of Geosciences (Wuhan), Wuhan 430079, China
*
Author to whom correspondence should be addressed.
†
Xueru Wang and Yang Liu contributed equally to this work.
Diversity 2022, 14(1), 2; https://0-doi-org.brum.beds.ac.uk/10.3390/d14010002
Submission received: 16 November 2021
/
Revised: 10 December 2021
/
Accepted: 13 December 2021
/
Published: 22 December 2021
(This article belongs to the Special Issue Linking Plankton Diversity with Ecosystem Functioning and Services)
Abstract
:Coccolithophores are involved in oceanic carbon and nitrogen cycles, and they also have an impact on global climate change. Chrysotila dentata have a complex and close relationship with phycosphere bacteria. In this study, culturable phycosphere bacteria (free-living bacteria and attached bacteria) are isolated from C. dentata by a gradient dilution method and identified based on the 16S rRNA gene sequencing analysis. The phylogenetic tree (neighbor-joining tree, N-J tree) was constructed using the bacterial sequences and closest related sequences from GenBank. Colony characteristics, Gram nature, and physiological and biochemical characteristics were obtained based on a series of tests, such as the sugar utilization (glucose, arabinose, xylose, maltose, and mannitol) test, Voges–Proskauer reaction, urease tests, gelatin liquefaction, Gram test, starch hydrolysis, among others. In this study, seven strains (CF1, CF2, CF3, CF5, CF6, and CF7) of free-living bacteria (CF) and five strains (CA1, CA2, CA3, CA4, and CA5) of attached bacteria (CA) are isolated and identified. We found that the culturable phycosphere bacteria of C. dentata were mainly α-proteobacteria and γ-proteobacteria, with a small part of the CFB (Cytophaga-Flexibacter-Bacteroides) group bacteria and firmicutes. In this study, most α-proteobacteria can utilize malonate and positive in the urease test, meanwhile they can grow in a 7% NaCl medium. Differently to α-proteobacteria, γ-proteobacteria are more reactive, and can utilize maltose, glucose, arabinose, malonate, aesculin, and starch hydrolysis. Meanwhile, γ-proteobacteria can growth in a 7% NaCl and pH 5.7 medium, and some bacteria of this strain were positive in nitrate reduction. Firmicutes are similar to γ-proteobacteria: they are similar in reactivity, as they can utilize maltose, glucose, arabinose, malonate, aesculin, and starch hydrolysis, and can growth in a 7% NaCl and pH 5.7 medium. The difference is that some of firmicutes were positive in gelatin liquefaction and can utilize mannitol. The CFB group of bacteria were only positive in malonate, aesculin, and starch hydrolysis. The above results provide basic experimental data for further studies on the relationship between the coccolithophores and culturable phycosphere bacteria.
1. Introduction
Chrysotila dentata belong to the Prymnesiophyceae of haptophyta [1], which accounts for about 20% of the total phytoplankton carbon pool [2]. C. dentata are marine microalgae with a biomineralizing effect in the ocean [3]. The surface of C. dentata consists of calcium carbonate produced by the core of coccolithophores. However, the ability of C. dentata to calcify is reduced under high temperatures and acidifying conditions [4]. C. dentata make impact of the light permeability of the ocean and global heat fluxes because its surface is covered with coccolith [5]. Coccolithophores are involved in the biological and carbonate pumps when they photosynthesize and calcify. They have a profound impact on the carbon cycle and carbonate system of marine ecosystems [1]. In addition, coccolithophores produce dimethyl sulfur gas, which is a precursor of dimethyl phosphorothioic acid. Dimethyl sulfur participates in the regulation of global climate by altering cloud condensation nodules through the sea–air exchange, as well as in the oceanic sulfur cycle. In brief, dimethyl sulfur has a significant impact on the marine environment.
C. dentata are influenced by bacteria from the phycosphere [6]. The phycosphere bacteria in the natural environment can be divided into two communities (free-living bacteria and attached bacteria), and there are differences in the composition of the two communities [6]. The phycosphere environment is similar to the inter-rooted environment in terrestrial ecosystems. In the ocean, microalgal cells are the starting point and extend outward for a distance. The microalgae can produce a variety of organic compounds, such as sugars, vitamins, toxins, enzymes, and other extracellular products that can stimulate and induce the action of epiphytic bacteria [7]. Additionally, epiphytic bacteria can provide organics, such as vitamins, phytohormones, chelator, or organic compounds, to create a favorable environment for microalgae in the phycosphere [8]. The phycosphere bacteria can break down organic carbon and aquatic plant debris [9]. Some bacteria have a competitive relationship with microalgae that may inhibit microalgal growth. Moreover, certain bacteria may promote the growth of microalgae in the early stage or inhibit the growth of microalgae in the later stage [8]. The existence of complex relationships between microalgae and heterotrophic bacteria is also influenced by a variety of factors (e.g., nitrogen, phosphorus, and dissolved organic carbon) in the ocean. It has been shown that the mutually beneficial symbiotic relationship between phytoplankton and heterotrophic bacteria is enhanced when nitrogen and phosphorus are added to the environment in a stoichiometric balance. However, when excess organic carbon is introduced into the environment, it can cause a nutritional competition between phytoplankton and heterotrophic bacteria [10].
Fandino et al. [11] showed that free-living bacteria mainly belong to α-proteobacteria and γ-proteobacteria during the algal bloom of flagellated algae, and attached bacteria are mainly dominated by CFB [12]. There is a clear phylogenetic difference between the bacterial community of phycosphere and the free-living bacterial community in the ocean, suggesting that the structural composition of bacterial populations from the two different environments are influenced by different selective pressures [13,14].
Coccolithophores can influence the environment and climate by mediating certain mechanisms, and they also have an important impact on marine and coastal ecology. However, the survival and growth of coccolithophores are importantly related to the phycosphere bacteria [15]. However, bacterial abundance and community structure depend heavily on the algal species, growth, and physiological status [13]. In this study, we study the bacterial community composition [free-living bacteria (CF) and attached bacteria (CA)] of C. dentata, which facilitates the exploration of the effects of C. dentata on photosynthesis and calcification.
2. Materials and Methods
2.1. Microalgae Collection and Culture Conditions
The size of the coccolithophore, Chrysotila dentata, was between 13–15 μm originally isolated from the Bohai Sea, China [16], in the collection of microalgal cultures of our laboratory. C. dentata was grown in a nutrient enriched artificial sea water sterilized at 121 °C for 20 min. Then, the algal cells were cultured for a week under the growth-saturating irradiance of 100 μmol m−2 s−1 photon flux density and a 14:10 h light: dark cycle at a constant temperature of 25 °C.
2.2. Isolation and Purification of Culturable Bacteria
Due to the size of C. dentata between 13–15 μm [16], the bacteria attached to C. dentata (the volume of algae and bacteria >3 μm) cannot pass the filtration. Moreover, free-living bacteria cannot attach to C. dentata if the size of the bacteria is <3 μm, so free-living bacteria can pass through a 3 μm pore-size filter paper. Therefore, we used the 3 μm pore-size filter paper to separate both types of bacteria from C. dentata at a plateau stage in an ultraclean bench [16]. The bacteria were diluted into six gradients of 10−1–10−6, and finally 20 μL of the bacterial solution were applied to the Zobells 2216E marine agar [17]. The bacteria were incubated in an incubator at 28 °C for 5–7 days according to the physical characteristics of the bacteria, and then the picked monoclonal colonies were purified 2–4 times and placed in an incubator at 28 °C for 4–6 days to obtain the purified monoclonal colonies. The monoclone was expanded with Zobells 2216E marine broth and cultured for 5 days on a shaker at 28 °C with 180 rpm. The cultured bacteria were stored at −80 °C.
2.3. Physiological and Biochemical Characterization
A total of 12 strains of bacteria isolated from C. dentata culture were selected for further studies. A part of isolated pure bacteria was made into bacterial suspensions by sanitary saline. Then, the bacterial suspensions were cultured 24–48 h in a broth of test ampoules (Qingdao Hope Bio-Technology Co., Ltd., Qingdao, China), and the colors of bacterial broth were observed. For this part, some indicators were used for physiological and biochemical tests. Different biochemical tests, such as the sugar utilization test (arabinose, maltose, glucose, xylose, and mannitol), nitrate reduction, aesculin, urease, Gram test, Voges–Proskauer reaction, gelatin liquefaction, malonate, starch hydrolysis, pH 5.7 and 7% NaCl [18].
2.4. DNA Extraction
Total DNA was extracted using a TIANamp Bacteria Genomic DNA Kit (Tiangen Biotech, Co., Ltd., Beijing, China). Then, the purified DNA was stored at −20 °C until further use.
2.5. PCR Amplification
Bar-coded fragments of the 16S rRNA gene were amplified using the primer 27F (5′-AGAGTTTGATCCTGGCAG-3′) and 1492R (5′-TACGGTTACCTTGTTACGACTT-3′) [19]. The quantitative PCR was carried out in a volume of 25 μL, including 12.5 μL 2 × Taq PCR Master Mix (Tiangen Biotech Co., Ltd., Beijing, China), 0.75 μM of each primer (27F and 1492R), 10 μL RNase-free water, and 1μL of template DNA. Thermal cycling involved incubation at 95 °C for 3 min to denature the DNA; the amplification was performed for 30 cycles at 98 °C for 10 s, 55 °C for 15 s, 72 °C for 10 s, and a final extension for 5 min at 72 °C [20]. All measurements were performed in triplicate. Then, the PCR products were checked with a 1% agarose gel and run in a 0.5 × TBE buffer. Finally, the PCR products were sent to the Huada Company (HuaDa, Beijing, China) for sequencing and artificial splicing.
2.6. Construction of Phylogenetic Trees
The sequences were submitted to the NCBI’s nucleotide program of the database project to detect the possible closest phylogenetic neighbors. A phylogenetic tree was constructed following the model by the neighbor-joining (N-J) approach using 1000 bootstrap replicates in MEGA 6.1 software [21].
3. Results
3.1. Identification of Culturable Phycosphere Bacteria
3.1.1. Isolation and Identification of Culturable Phycosphere Bacteria
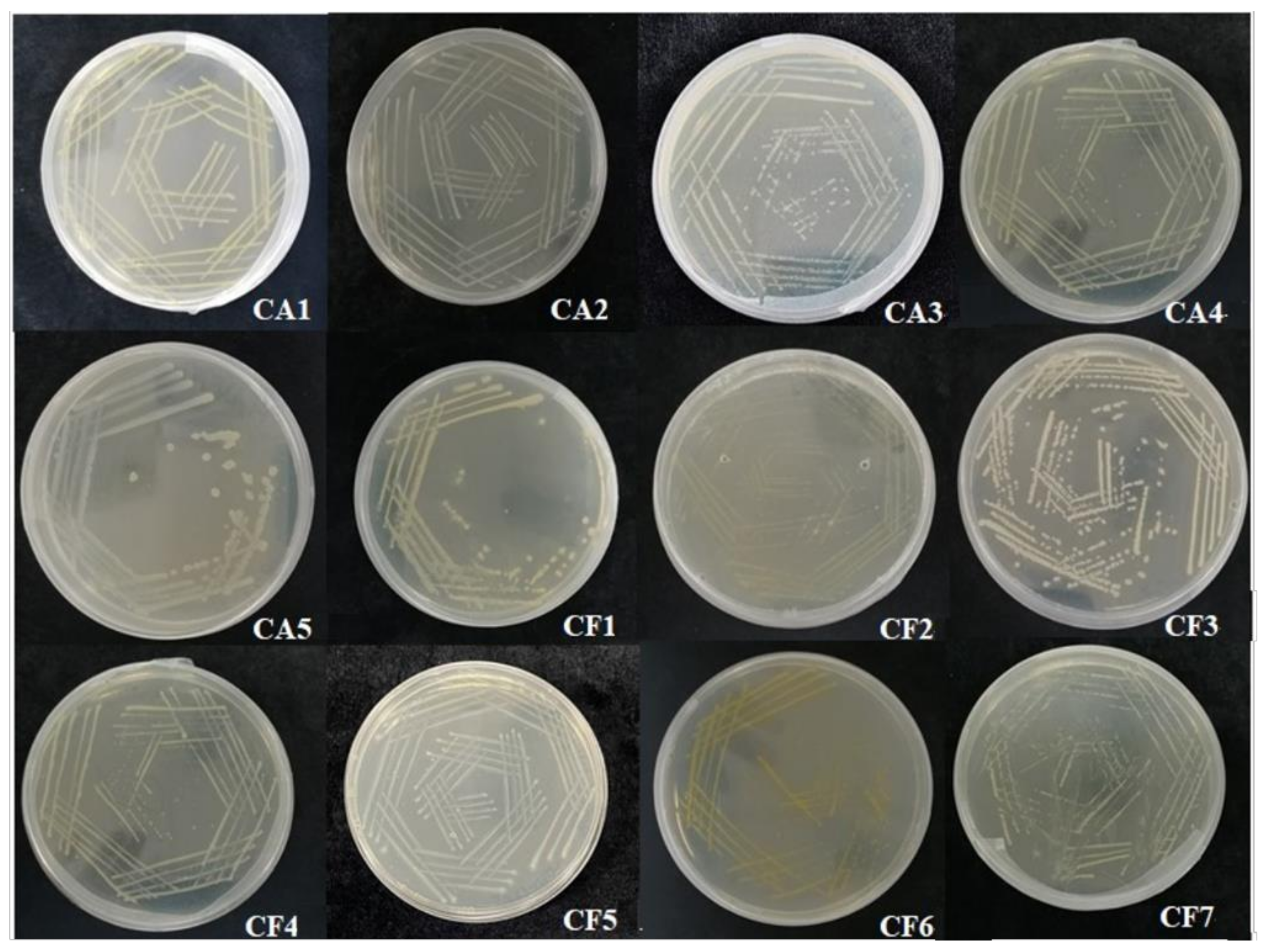
Several culturable phycosphere bacteria were isolated from the samples of C. dentata, and then we analyzed the 16S rRNA gene sequence of each culturable bacterial sample. Finally, the results obtained by sequencing were compared using BLAST on NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 26 November 2021). We obtained a total of 12 different bacterial strains, numbered CA1, CA2, CA3, CA4, CA5, CF1, CF2, CF3, CF4, CF5, CF6, and CF7 (Figure 1).
3.1.2. Identification of Physiological and Biochemical Characteristics of Phycosphere Bacteria in C. dentata
The associated bacteria to the C. dentata culture were isolated and three groups of bacteria were identified as follows (Table 1): CFB group (CA1, CF2, and CF6), α-proteobacteria (CA2, CF1, CF4, and CF7), γ-proteobacteria (CA4, CA3, and CF5), and firmicutes (CA5 and CF3). The dominant specie of bacteria from coccolithophores was proteobacteria. And the CFB group and Bacillus which account for about 40% of the total isolated bacteria. However, the CFB group, proteobacteria (α-proteobacteria and γ-proteobacteria), and firmicutes were present in both of the attached bacteria and in free-living bacteria around C. dentata.
The CA1, CF2, and CF3 in the CFB group bacteria belong to the genus Flavobacterium; CA5 and CF3 belong to the genus Bacillus; CA2 and CF7 belong to the genus Stappia sp.; and the strain CF1 belongs to Alpha proteobacterium. CF4 is a bacterium belonging to Thalassococcus sp.; the strains CF5 and CA3 belong to the Marinobacter alkaliphilus and Marinobacter hydrocarbonoclasticus, respectively. CA4 belongs to the genus Alteromonas. The Stappia sp. (CA2 and CF7) and Marinobacter (CF5 and CA3) genera were found in attached and free-living bacteria.
3.2. Phylogenetic Tree of Culturable Phycosphere Bacteria from C. dentata
The 16S rRNA gene sequences of the phycosphere bacteria isolated from C. dentata and closer sequences strains were obtained from the BLAST search. Then, they were imported into the MEGA X64 Software to construct the phylogenetic tree using the neighbor-joining method (Figure 2).
3.3. Phenotype Charateristics of Culturable Phycosphere Bacteria
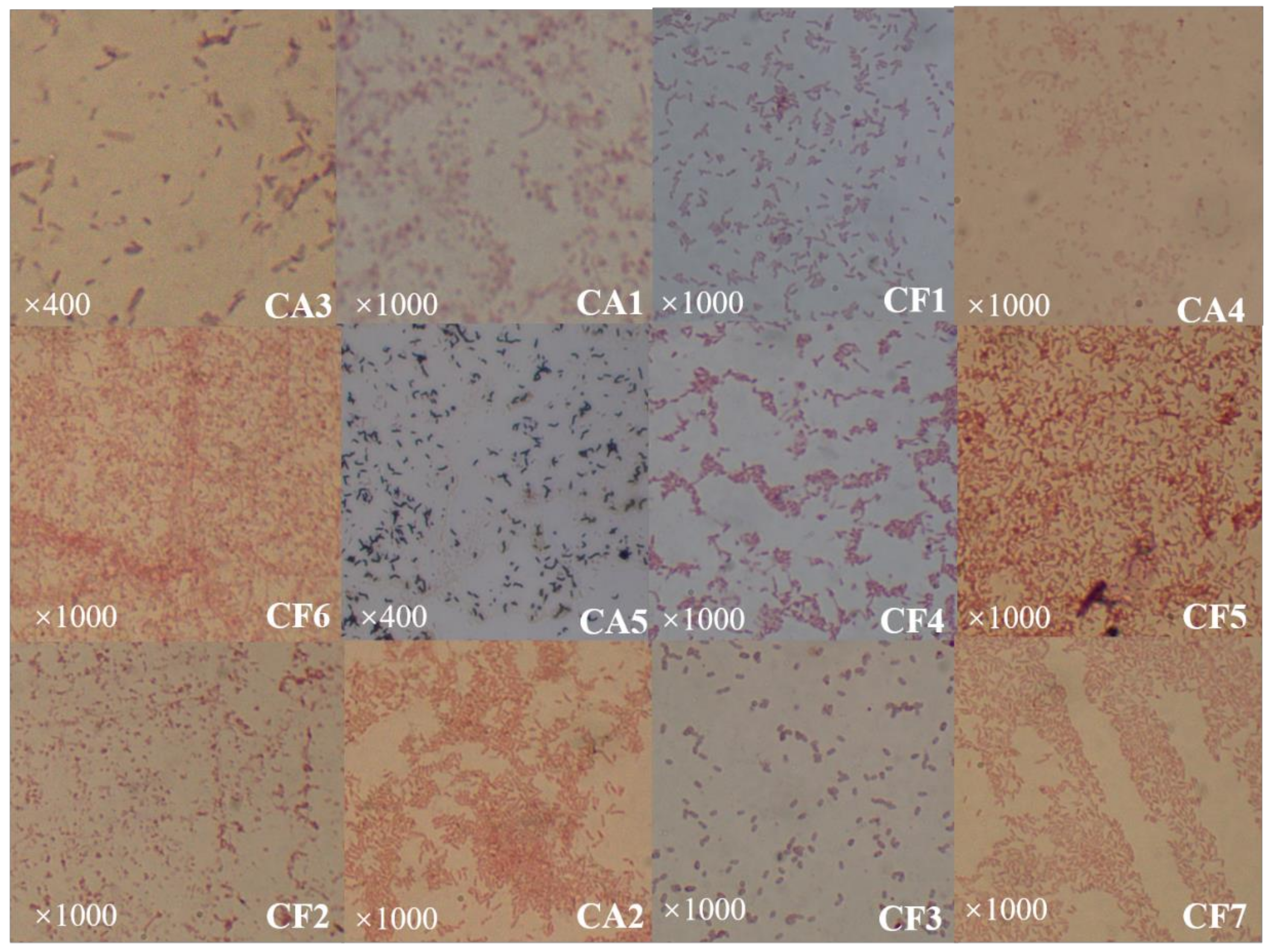
In Figure 1, it can be seen that the phycosphere bacteria presented in the surroundings of the coccolithophores were predominantly yellow, white, and a few were transparent (CF5). These colonies are about 0.5 mm in size and most bacteria are circular with only a few being oval in shape. Most of the colonies have apophysis in their surfaces with no radiolucent edges (Table 2). Based on the staining, both Gram positive (CA5 and CF3) and Gram negative bacteria were obtained (Figure 3).
3.4. Physiological and Biochemical Characteristics
The growth of CA3, CA5, CF5, and CF7 was observed in a medium of pH 5.7 and 7% NaCl. The strains CA2, CA4, and CF1 can grow in 7% NaCl. The strain CF3 was observed in a pH 5.7 medium. Unlike the other related phycosphere bacteria, strain CF3 was positive in the Voges–Proskauer reaction.
In this study, the strain CA5 was positive for gelatin liquefaction, and CA2, CF1, and CA7 were positive in the urease test. CA3 and CF5 were positive for the maltose test. Only the strain CA5 can utilize glucose. CA3, CA5, and CF6 were negative for malonate. The stains CA3, CA5, CF3, CF5 CF6, and CF7 can hydrolyze aesculin. In addition, CF7 only make utilization of xylose. CA3, CA5, CF3, CF5, CF6, and CF7 have the function of to hydrolyze starch. The strains CA3 and CF1 were positive in a nitrate reduction. CA2, CA5, and CF3 were positive in the mannitol test, and CA3, CA5, CF3, CF5, and CF7 were positive in the arabinose test. The physiological and biochemical characteristics of the 12 strains are shown in Table 3.
4. Discussion
In natural ecosystems, there are complex and varied relationships (e.g., symbiosis and inhibition) between microalgae and epiphytic bacteria. The community composition of epiphytic bacteria plays an important role for microalgae. In this study, the culturable bacteria of the C. dentata were isolated and identified. The experimental results show that firmicutes, α-proteobacteria, γ-proteobacteria, and the CFB group were found in the cultured attached bacteria of C. dentata. The culture of free-living bacteria include mainly proteobacteria and the CFB group, of which α-proteobacteria was the dominant bacterial taxa in the cultures. The bacterial community composition was also similar between cultures of free-living and attached bacteria, but γ-proteobacteria account for a part of the culture of attached bacteria. It can be observed that most of the culturable bacteria from C. dentata are similar to marine bacteria. In addition, the firmicutes strain was found in the attached bacteria of C. dentata, which can exist in soil, water, and air. Certain strains of firmicutes play an important role in harmful algal blooms, for example bacillemia secreted by the bacterium Bacillus sp. SY-1 had a strong algicidal effect on chlodinium polykrikoides [22]. Previous studies have shown that α-proteobacteria, γ-proteobacteria, and the CFB group are common taxa in the marine environment [23,24]. Kimbrel et al. [25] explained that there are differences in the community composition of attached and free-living bacteria. Some of the most important factors (CO2, nutrients, pH and seasonal change, among others) that affect algal growth are well known [26]. However, free-living bacteria partially overlap with attached bacteria in the composition of bacteria. It has been speculated that the composition of free-living and attached bacterial communities seems to be related to the species of microalgae and the living environment [27].
Cho et al. [28] identified 14 strains culturable bacteria isolated from Chlorella sp., and the results showed that actinobacteria, α-proteobacteria, β-proteobacteria, and the CFB group are the main bacterial taxa, with α-proteobacteria and CFB accounting for the majority of bacterial taxa. Several studies have shown that the culturable bacterial taxa of Dunaliella salina include α-proteobacteria, γ-proteobacteria, CFB group, and Gram-positive bacteria [29]. For marine diatoms, α-proteobacteria were present in free-living bacteria, the CFB group was present in attached bacteria, and γ-proteobacteria were present in both of the above bacteria [13]. Steven et al. [30] found that α-, β-, γ-proteobacteria, and Bacteroides are widely found in surface layers of ocean. In summary, proteobacteria are the dominant bacterial taxon in marine microalgae, consistent with the results of this study.
Biomineralization links organism with elements of the solid earth, for example the silicification of diatoms and the calcification of coccolithophores [31], and certain microalgae cells surface are covered with coccolith [5]. Schäfer et al. [12] analyzed the "satellite" bacteria of six marine diatoms species during repeated passage and culture without significant genetic fingerprint variability, suggesting that the relationship between microalgae and phycosphere bacteria is stable [13]. Microalgae release soluble organic compounds to provide a carbon source for epiphytic bacteria, and the extracellular enzymes produced by the epiphytic bacteria degrade the organic polymers [32]. Therefore, the interaction of epiphytic bacteria with diatoms and coccolithophores is involved in the decomposition and elemental cycling of organic matter in the ocean [13]. Based on the above result, we found that proteobacteria is the main bacterial taxon in the phycosphere. β-proteobacteria were not found in C. dentata, but the presence of β-proteobacteria in other marine microalgae suggests that the community distribution of bacteria is related to a variety of factors [33].
In this study, γ-proteobacteria dominated in cultured attached bacteria and they can utilize various sugars, including maltose, glucose, arabinose, malonate, aesculin, and starch, with certain strains of γ-proteobacteria being positive in a nitrate reduction. Most of α-proteobacteria can only utilize malonate and grow in 7% NaCl medium and are positive in a urease test, which is the difference with γ-proteobacteria. Firmicutes that exist in cultural attached and free-living bacteria are similar to γ-proteobacteria, such as in their sugar utilization. Some of the firmicutes were positive in gelatin liquefaction and can utilize mannitol, which is different from the case of γ-proteobacteria. The CFB group bacteria were only positive in malonate, aesculin, and starch hydrolysis in this study. In cultured free-living bacteria, there are more types of α-proteobacteria than there are of the others, but only a small part of α-proteobacteria can perform certain functions. In terms of sugar utilization, most of phycosphere bacteria can utilized arabinose and malonate. When malonate is used as a carbon source, it is decomposed to sodium carbonate, so malonate can supply carbonate ion to C. dentata. The present study provided some basic information for further studies on the relationship between the coccolithophores and culturable phycosphere bacteria.
5. Conclusions
The results of this research demonstrate that proteobacteria is the dominant cultured bacterial taxon of C. dentata in the collection of microalgal cultures of our laboratory conditions. The free-living and attached bacteria were mainly α-proteobacteria and γ-proteobacteria, respectively. Firmicutes and the CFB group bacteria are also found in two different living status of the bacterial communities of C. dentata. In addition, the cultured free-living and attached bacteria of coccolithophores contain similar bacterial communities. Then, physiological and biochemical characteristics indicate that free-living bacteria performed some functions, such as the utilized malonate, aesculin, mannitol, arabinose, and starch hydrolysis, and can growth in a pH 5.7 and 7% NaCl medium. The attached bacteria utilized maltose, aesculin, and arabinose; grew in a pH 5.7 and up to 7% NaCl medium, and can hydrolyze starch.
Author Contributions
Conceptualization, J.S. and Y.L.; methodology, X.W. and Y.L.; formal analysis, X.W. and Y.L.; resources, J.S.; writing—original draft preparation, X.W.; writing—review and editing, X.W., Y.L.; visualization, X.W.; supervision, J.S.; project administration, J.S.; funding acquisition, J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the National Key Research and Development Project of China (2019YFC1407805), National Natural Science Foundation of China (41876134) and the Changjiang Scholar Program of Chinese Ministry of Education (T2014253) to Jun Sun.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Sun, J.; Jin, S.F. Species diversity of living coccolithophores in Chinese sea waters. Biodivers. Sci. 2011, 19, 787–797. (In Chinese) [Google Scholar]
- Krumhardt, K.M.; Lovenduski, N.S.; Iglesias-Rodriguez, M.D.; Kleypas, J.A. Coccolithophores growth and calcification in a changing ocean. Prog. Oceanogr. 2017, 159, 276–295. [Google Scholar] [CrossRef]
- Weiner, S.; Dove, P.M. An Overview of Biomineralization Processes and the Problem of the Vital Effect. Rev. Mineral. Geochem. 2003, 54, 1–29. [Google Scholar] [CrossRef]
- Matoo, O.B.; Ivanina, A.V.; Ullstad, C.; Beniash, E.; Sokolova, I.M. Interactive effects of elevated temperature and CO2 levels on metabolism and oxidative stress in two common marine bivalves (Crassostrea virginica and Mercenaria mer-cenaria). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 164, 545–553. [Google Scholar] [CrossRef]
- Sun, J. Organic carbon pump and carbonate counter pump of living coccolithophorid. Adv. Earth Sci. 2007, 22, 23–31. [Google Scholar]
- Miao, Z.; Du, Z.J.; Li, H.R.; Lou, Y.Y.; Luo, W. Analysis of bacterial diversity in the phycosphere of five Arcticmi-croalgae. Acta Ecol. Sin. 2015, 35, 1587–1600. [Google Scholar]
- Bell, W.; Mitchell, R. Chemotactic and growth responses of marine bacteria to algal extracellular products. Biol. Bull. 1972, 143, 265–277. [Google Scholar] [CrossRef]
- Li, Y.; Liu, F.L.; Wang, Q.H.; Liu, Y.; Li, J.Y.; Ging, Q.L. Effects of cultivable phycosphere bacteria on growth of Haematococcus pluvialis and its astaxanthin accumulation. Period. Ocean Univ. China 2020, 50, 44–55. (In Chinese) [Google Scholar]
- Anesio, A.; Abreu, P.; Biddanda, B. The role of free and attached microorganisms in the decomposition of estuarine macrophyte detritus. Estuar. Coast. Shelf Sci. 2003, 56, 197–201. [Google Scholar] [CrossRef]
- Cao, X.; Li, H.; Zhou, Y.; Song, C. The shift of mutualistic relationships among algae, free-living and attached bacteria through different nutrient addition mode: A mesocosm study. J. Freshw. Ecol. 2020, 35, 535–548. [Google Scholar] [CrossRef]
- Fandino, L.; Riemann, L.; Steward, G.; Long, R.; Azam, F. Variations in bacterial community structure during a dinoflagellate bloom analyzed by DGGE and 16S rDNA sequencing. Aquat. Microb. Ecol. 2001, 23, 119–130. [Google Scholar] [CrossRef]
- 1Schäfer, H.; Abbas, B.; Witte, H.; Muyzer, G. Genetic diversity of ‘satellite’ bacteria present in cultures of marine diatoms. FEMS Microbiol. Ecol. 2002, 42, 25–35. [Google Scholar]
- Grossart, H.-P.; Levold, F.; Allgaier, M.; Simon, M.; Brinkhoff, T. Marine diatom species harbour distinct bacterial communities. Environ. Microbiol. 2005, 7, 860–873. [Google Scholar] [CrossRef] [PubMed]
- Rooney-Varga, J.; Giewat, M.; Savin, M.; Sood, S.; LeGresley, M.; Martin, J. Links between Phytoplankton and Bacterial Community Dynamics in a Coastal Marine Environment. Microb. Ecol. 2005, 49, 163–175. [Google Scholar] [CrossRef]
- Amin, S.A.; Green, D.H.; Hart, M.C.; Kupper, F.C.; Sunda, W.G.; Carrano, C.J. Photolysis of iron-siderophore chelates promotes bacterial-algal mutualism. Proc. Natl. Acad. Sci. USA 2009, 106, 17071–17076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhang, X.; Sun, J.; Satheeswaran, T.; Zhang, G.; Li, H.; An, X. Morphology, phylogenetic position, and ecophysiological features of the coccolithophore Chrysotila dentata (Prymnesiophyceae) isolated from the Bohai Sea, China. Phycologia 2019, 58, 628–639. [Google Scholar] [CrossRef]
- Green, D.H.; Llewellyn, L.E.; Negri, A.; Blackburn, S.I.; Bolch, C.J. Phylogenetic and functional diversity of the cultivable bacterial community associated with the paralytic shellfish poisoning dinoflagellate Gymnodinium catenatum. FEMS Microbiol. Ecol. 2004, 47, 345–357. [Google Scholar] [CrossRef]
- Mehta, A.; Bhardwaj, K.K.; Shaiza, M.; Gupta, R. Isolation, characterization and identification of pesticide de-grading bacteria from contaminated soil for bioremediation. Biol. Futur. 2021, 72, 317–323. [Google Scholar] [CrossRef]
- Bosshard, P.P.; Santini, Y.; Grüter, D.; Stettler, R.; Bachofen, R. Bacterial diversity and community composition in the chemocline of the meromictic alpine Lake Cadagno as revealed by 16S rDNA analysis. FEMS Microbiol. Ecol. 2000, 31, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishna, K.P.; Macadangdang, B.R.; Rogers, M.B.; Tometich, J.T.; Firek, B.A.; Baker, R.; Ji, J.; Burr, A.H.P.; Ma, C.; Good, M.; et al. Maternal IgA protects against the development of necrotizing enterocolitis in preterm infants. Nat. Med. 2019, 25, 1110–1115. [Google Scholar] [CrossRef]
- Hagström, Å.; Pinhassi, J.; Zweifel, U.L. Biogeographical diversity among marine bacterioplankton. Aquat. Microb. Ecol. 2000, 21, 231–244. [Google Scholar] [CrossRef]
- Wang, Y.; Coyne, K.J. Immobilization of algicidal bacterium Shewanella sp. IRI-160 and its application to control harmful dinoflagellates. Harmful Algae 2020, 94, 101798. [Google Scholar] [CrossRef] [PubMed]
- Eilers, H.; Pernthaler, J.; Peplies, J.; Glöckner, F.O.; Gerdts, G.; Amann, R. Isolation of Novel Pelagic Bacteria from the German Bight and Their Seasonal Contributions to Surface Picoplankton. Appl. Environ. Microbiol. 2001, 67, 5134–5142. [Google Scholar] [CrossRef] [Green Version]
- Agogué, H.; Casamayor, E.O.; Bourrain, M.; Obernosterer, I.; Joux, F.; Herndl, G.J.; LeBaron, P.; Agogué, H. A survey on bacteria inhabiting the sea surface microlayer of coastal ecosystems. FEMS Microbiol. Ecol. 2005, 54, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Kimbrel, J.A.; Samo, T.; Ward, C.; Nilson, D.; Thelen, M.P.; Siccardi, A.; Zimba, P.; Lane, T.W.; Mayali, X. Host selection and stochastic effects influence bacterial community assembly on the microalgal phycosphere. Algal Res. 2019, 40, 101489. [Google Scholar] [CrossRef]
- Huang, Y.-J.; Jiang, Z.-B.; Zeng, J.-N.; Chen, Q.-Z.; Zhao, Y.-Q.; Liao, Y.-B.; Shou, L.; Xu, X.-Q. The chronic effects of oil pollution on marine phytoplankton in a subtropical bay, China. Environ. Monit. Assess. 2011, 176, 517–530. [Google Scholar] [CrossRef]
- Leflaive, J.; Danger, M.; Lacroix, G.; Lyautey, E.; Oumarou, C.; Ten-Hage, L. Nutrient effects on the genetic and functional diversity of aquatic bacterial communities. FEMS Microbiol. Ecol. 2008, 66, 379–390. [Google Scholar] [CrossRef]
- Cho, D.H.; Ramanan, R.; Heo, J.; Lee, J.; Kim, B.H.; Oh, H.M.; Kim, H.S. Enhancing microalgal biomass produc-tivity by engineering a microalgal–bacterial community. Bioresour. Technol. 2015, 175, 578–585. [Google Scholar] [CrossRef]
- Le Chevanton, M.; Garnier, M.; Bougaran, G.; Schreiber, N.; Lukomska, E.; Bérard, J.-B.; Fouilland, E.; Bernard, O.; Cadoret, J.-P. Screening and selection of growth-promoting bacteria for Dunaliella cultures. Algal Res. 2013, 2, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Stevens, H.; Stübner, M.; Simon, M.; Brinkhoff, T. Phylogeny of Proteobacteria and Bacteroidetes from oxic habitats of a tidal flat ecosystem. FEMS Microbiol. Ecol. 2005, 54, 351–365. [Google Scholar] [CrossRef]
- Raven, J.A.; Giordano, M. Biomineralization by photosynthetic organisms: Evidence of coevolution of the organisms and their environment? Geobiology 2009, 7, 140–154. [Google Scholar] [CrossRef] [PubMed]
- Middelboe, M.; Letarte, Y.; Borch, N.H. Attached and free-living bacteria: Production and polymer hydrolysis during a diatom bloom. Microb. Ecol. 1995, 29, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.J.; Sun, J.; Feng, Y.Y. Study on modern coccolithophores in coastal region along the east Hainan Island. Haiyang Xuebao 2015, 37, 27–40. [Google Scholar]
Figure 1.
Colonies of the 12 phycosphere bacteria isolated from C. dentata.

Figure 2.
Phylogenetic evolutionary tree of the phycosphere bacteria isolated from C. dentata. (a) free-living bacteria; (b) attached bacteria.
Figure 2.
Phylogenetic evolutionary tree of the phycosphere bacteria isolated from C. dentata. (a) free-living bacteria; (b) attached bacteria.

Figure 3.
Gram staining of the 12 strains of phycosphere bacteria isolated from C. dentata. The “×400” and “×1000” repreScheme.
Figure 3.
Gram staining of the 12 strains of phycosphere bacteria isolated from C. dentata. The “×400” and “×1000” repreScheme.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Identification of 12 strains of phycosphere bacteria.
| Samples | Accession | Closest Relative in Database | Accession Number | Similarity (%) |
|---|---|---|---|---|
| CFB group | ||||
| CF2 | SRR17041857 | Flavobacteriaceae bacterium LA8 | AF513435.1 | 99.57 |
| CF6 | SRR17041863 | Flavobacterium sp. Strain 7002-040 | KY770468.1 | 99.71 |
| CA1 | SRR17041865 | Flavobacterium sp. Strain 7002-040 | KY770468.1 | 99.64 |
| α-proteobacteria | ||||
| CA2 | SRR17041864 | Stappia sp. strain FG-4 | JF451107.1 | 100.00 |
| CF1 | SRR17041858 | Alpha proteobacterium 45A11 | JF263527.1 | 99.92 |
| CF4 | SRR17041855 | Thalassococcus sp. strain S24 | KX989355.1 | 100.00 |
| CF7 | SRR17041862 | Stappia sp. Strain VB | MH454495.1 | 100.00 |
| γ-proteobacteria | ||||
| CA4 | SRR17041860 | Alteromonas sp. strain KSTI56 | KX989422.1 | 100.00 |
| CA3 | SRR17041861 | Marinobacterhydrocarbonoclasticus strain RMR 21 | KP769865.1 | 99.46 |
| CF5 | SRR17041854 | Marinobacter alkaliphilus strain Ad 1 | MF029656.2 | 99.85 |
| firmicutes | ||||
| CF3 | SRR170418856 | Bacillus firmus strain sctcc471 | HQ622343.1 | 99.86 |
| CA5 | SRR17041859 | Bacillus xiamenensis strain SN3-8 | MT071697.1 | 99.93 |
Table 2.
Colony characteristics and Gram nature of phycosphere bacteria isolated from C. dentata.
| Strain Number | Color | Diameter (mm) | Shape | Elevation | Margin | Gram Nature |
|---|---|---|---|---|---|---|
| CA1 | Yellow | 0.7 | Circular | Flat | Entire | Negative |
| CA2 | White | 0.4 | Circular | Flat | Entire | Negative |
| CA3 | White | 0.4 | Circular | Flat | Entire | Negative |
| CA4 | Pale yellow | 0.7 | Circular | Flat | Entire | Negative |
| CA5 | White | 1.2 | Circular | Flat | Entire | Positive |
| CF1 | Pale yellow | 0.6 | Circular | Flat | Entire | Negative |
| CF2 | Bright yellow | 0.4 | Circular | Flat | Entire | Negative |
| CF3 | White | 0.7 | Circular | Flat | Entire | Positive |
| CF4 | Pale yellow | 0.25 | Circular | Flat | Entire | Negative |
| CF5 | Transparent | 0.35 | Circular | Bulge | Extension | Negative |
| CF6 | Bright yellow | 0.45 | Oval | Flat | Entire | Negative |
| CF7 | Pale yellow | 0.15 | Circular | Bulge | Entire | Negative |
Table 3.
Physiological and biochemical characteristics of phycosphere bacteria isolated from C. dentata.
Table 3.
Physiological and biochemical characteristics of phycosphere bacteria isolated from C. dentata.
| Target | Physiological and Biochemical Characteristics | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CA1 | CA2 | CA3 | CA4 | CA5 | CF1 | CF2 | CF3 | CF4 | CF5 | CF6 | CF7 | |
| Voges–Proskauer | − | − | − | − | − | − | − | − | − | − | − | − |
| Gelatin liquefaction | − | − | − | − | + | − | − | − | − | − | − | − |
| Urease test | − | + | - | − | − | + | − | − | − | − | − | + |
| Maltose | − | − | + | − | − | − | − | − | − | + | − | − |
| Glucose | − | − | − | − | + | − | − | − | − | − | − | − |
| Xylose | − | − | − | − | − | − | − | − | − | − | − | + |
| Arabinose | − | − | + | − | + | − | − | + | − | + | − | + |
| Mannitol | − | − | − | − | + | − | − | − | − | − | − | − |
| Starch hydrolysis | − | − | + | − | + | − | − | + | − | + | + | + |
| Malonate | + | + | − | + | − | + | + | + | + | + | + | + |
| Aesculin | − | − | + | − | + | − | − | + | − | + | + | + |
| Nitrate reduction | − | − | + | − | − | + | − | − | − | − | − | + |
| pH 5.7 | − | − | + | − | + | − | − | + | − | + | − | + |
| 7% NaCl | − | + | + | + | + | + | − | − | − | + | − | + |
Plus sign (+) indicates a positive reaction; minus sign (−) indicates a negative reaction. Flavobacterium sp. (KY770468.1) is the closest reference strain with CA1; Stappia sp. (JF451107.1) is closest with CA2; Marinobacter hydrocarbonoclasticus (KP769865.1) is the closest microalgae bacteria to CA3; Alteromonas sp. (KX989422.1) is the closest reference stain with CA4 from the BLAST search. Among the free-living bacteria, Alpha proteobacterium (JF263527.1), Flavobacteriaceae bacterium (AF513435.1), Bacillus firmus (HQ622343.1), Thalassococcus sp. (KX989355.1), Marinobacter alkaliphilus icroa (MF029656.2), Flavobacterium sp. (KY770468.1), and Stappia sp. (MH454495.1) are the closest reference strains with CF1, CF2, CF3, CF4, CF5, CF6, and CF7, respectively.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, X.; Liu, Y.; Sun, J. Physiological and Biochemical Characterization of Isolated Bacteria from a Coccolithophore Chrysotila dentata (Prymnesiophyceae) Culture. Diversity 2022, 14, 2. https://0-doi-org.brum.beds.ac.uk/10.3390/d14010002
AMA Style
Wang X, Liu Y, Sun J. Physiological and Biochemical Characterization of Isolated Bacteria from a Coccolithophore Chrysotila dentata (Prymnesiophyceae) Culture. Diversity. 2022; 14(1):2. https://0-doi-org.brum.beds.ac.uk/10.3390/d14010002
Chicago/Turabian StyleWang, Xueru, Yang Liu, and Jun Sun. 2022. "Physiological and Biochemical Characterization of Isolated Bacteria from a Coccolithophore Chrysotila dentata (Prymnesiophyceae) Culture" Diversity 14, no. 1: 2. https://0-doi-org.brum.beds.ac.uk/10.3390/d14010002
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.