
Ecological Study of the Active Ciliate Community from Bromeliads during the Humid Season in Different Types of Forests of the Mexican Neotropics

Abstract
:1. Introduction
2. Materials and Methods
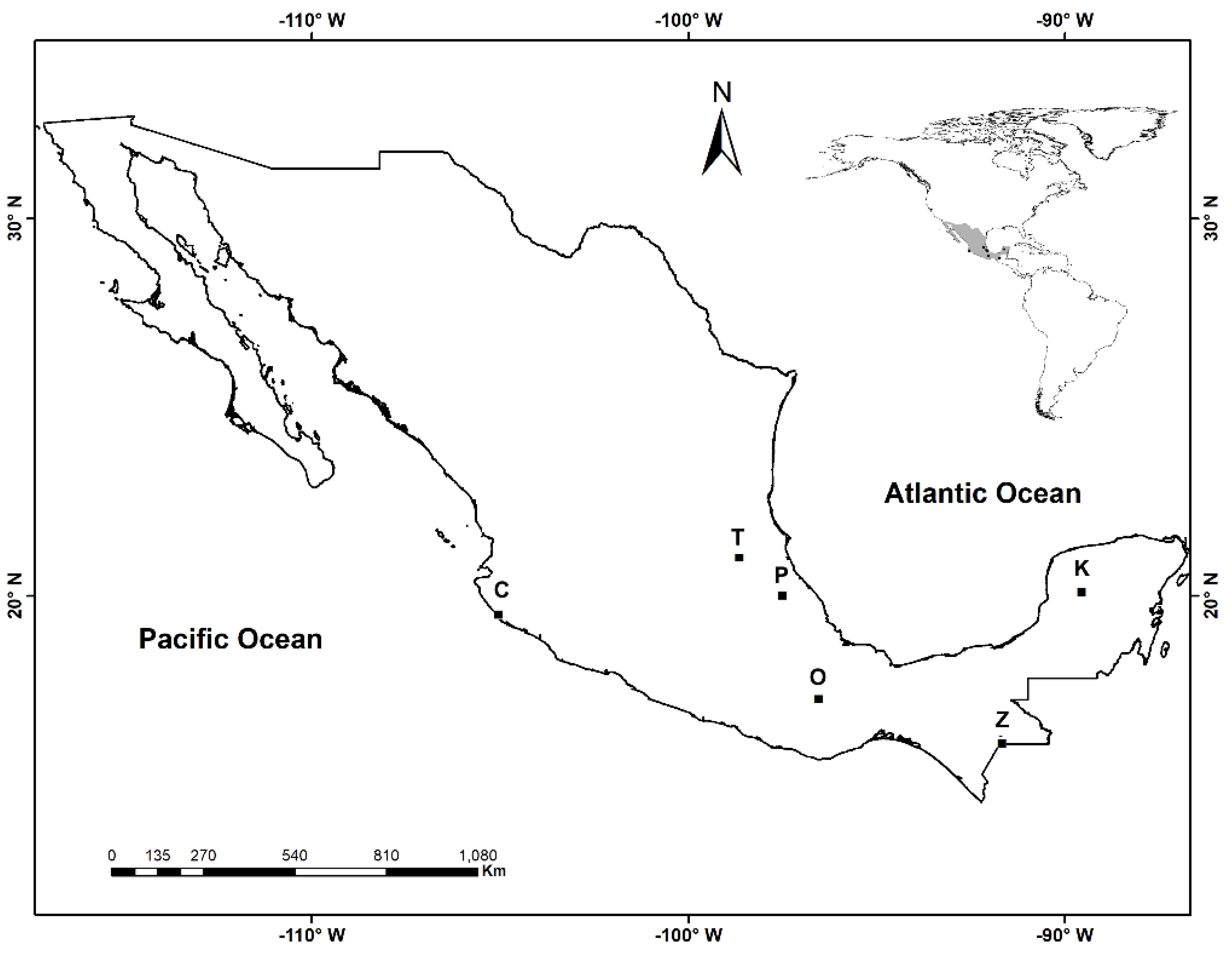
2.1. Area of Study
2.2. Sample Collection
2.3. Ciliate Identification
2.4. Laboratory Analysis
2.5. Data Analysis
3. Results
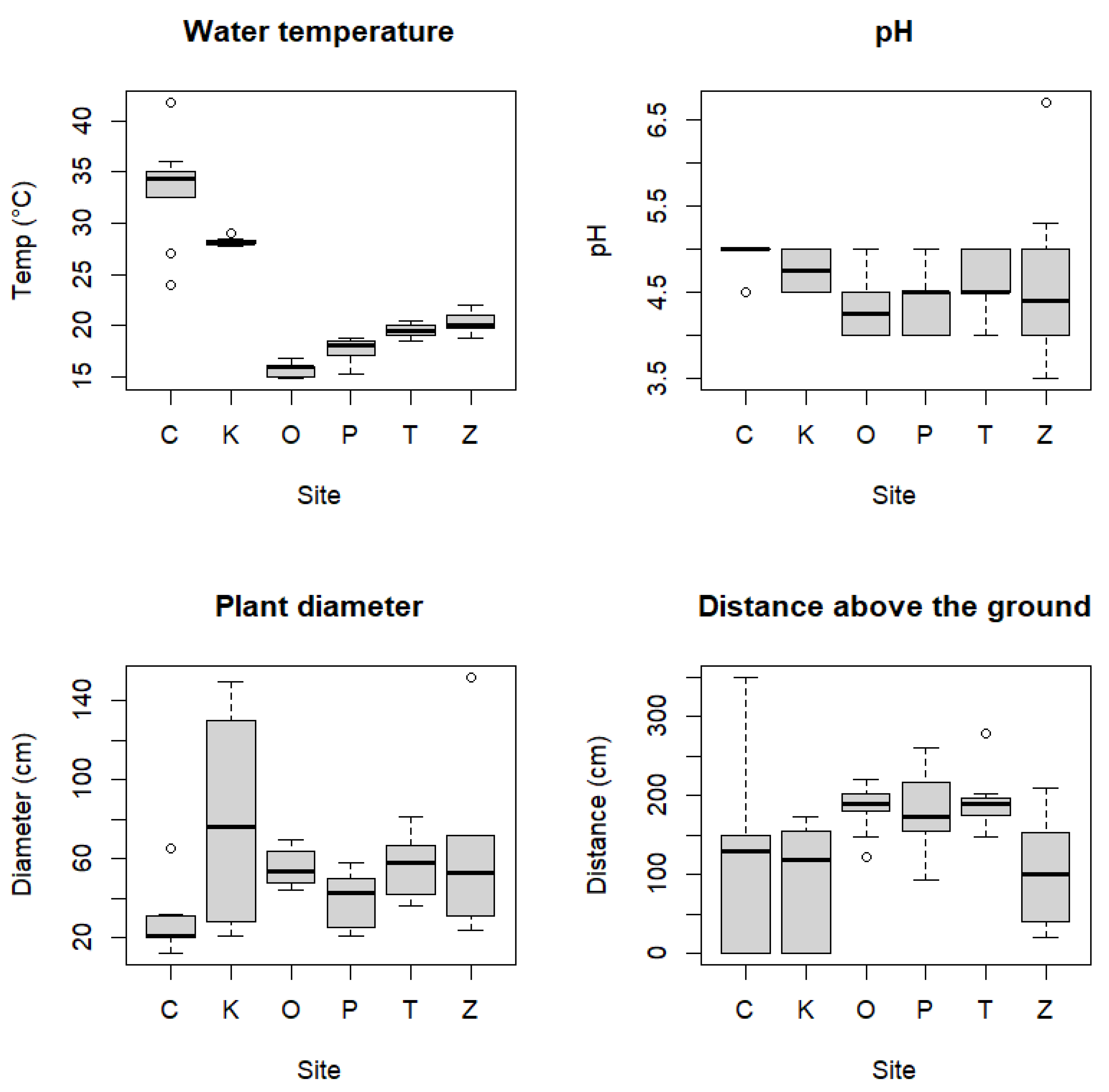
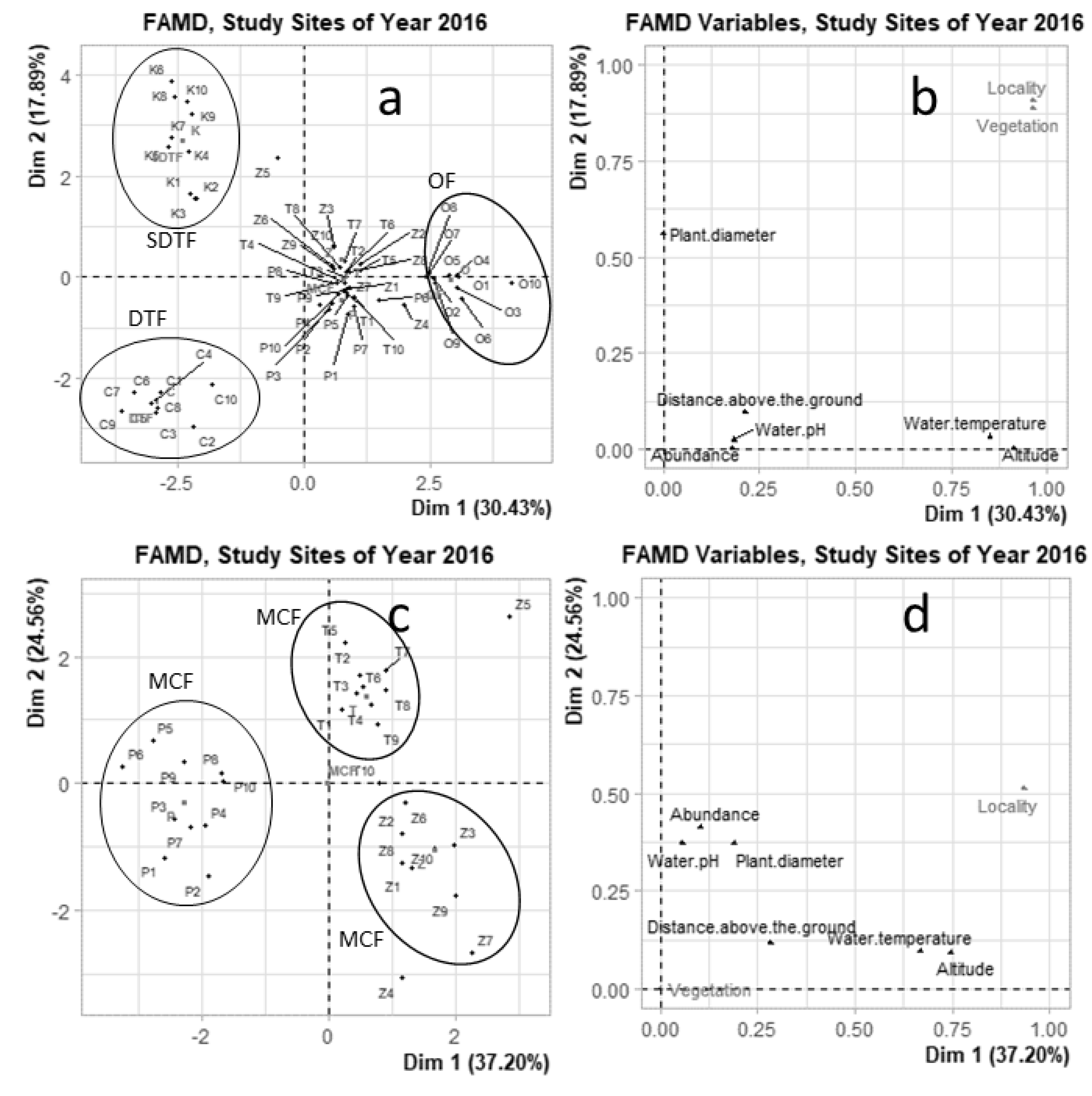
3.1. Microhabitat Characterization
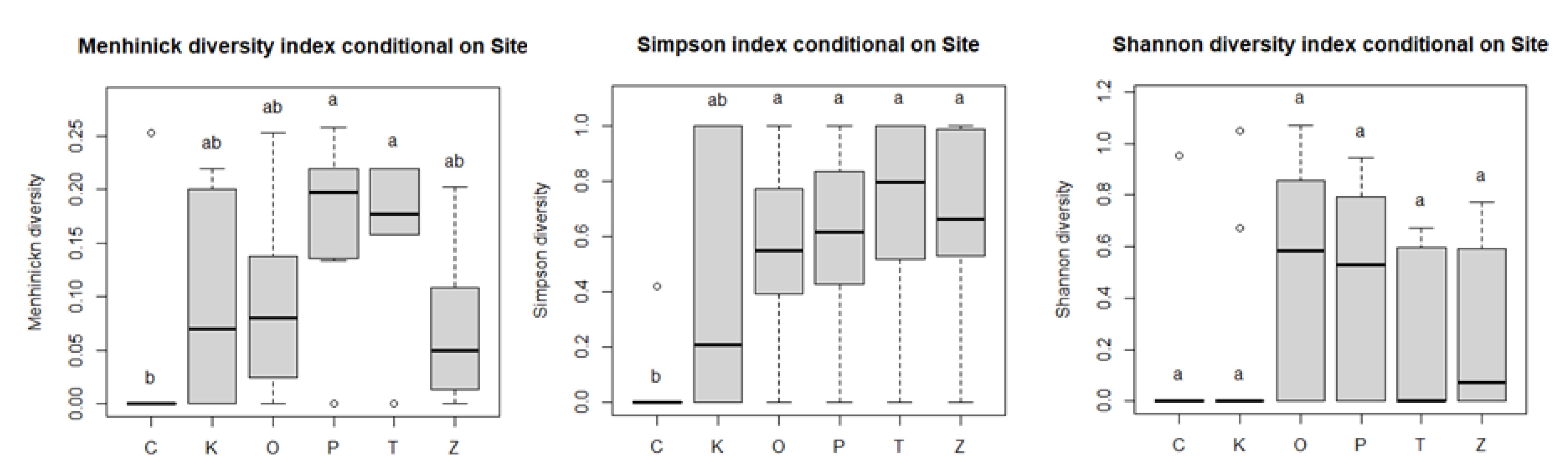
3.2. Species Richness
3.3. Abundance
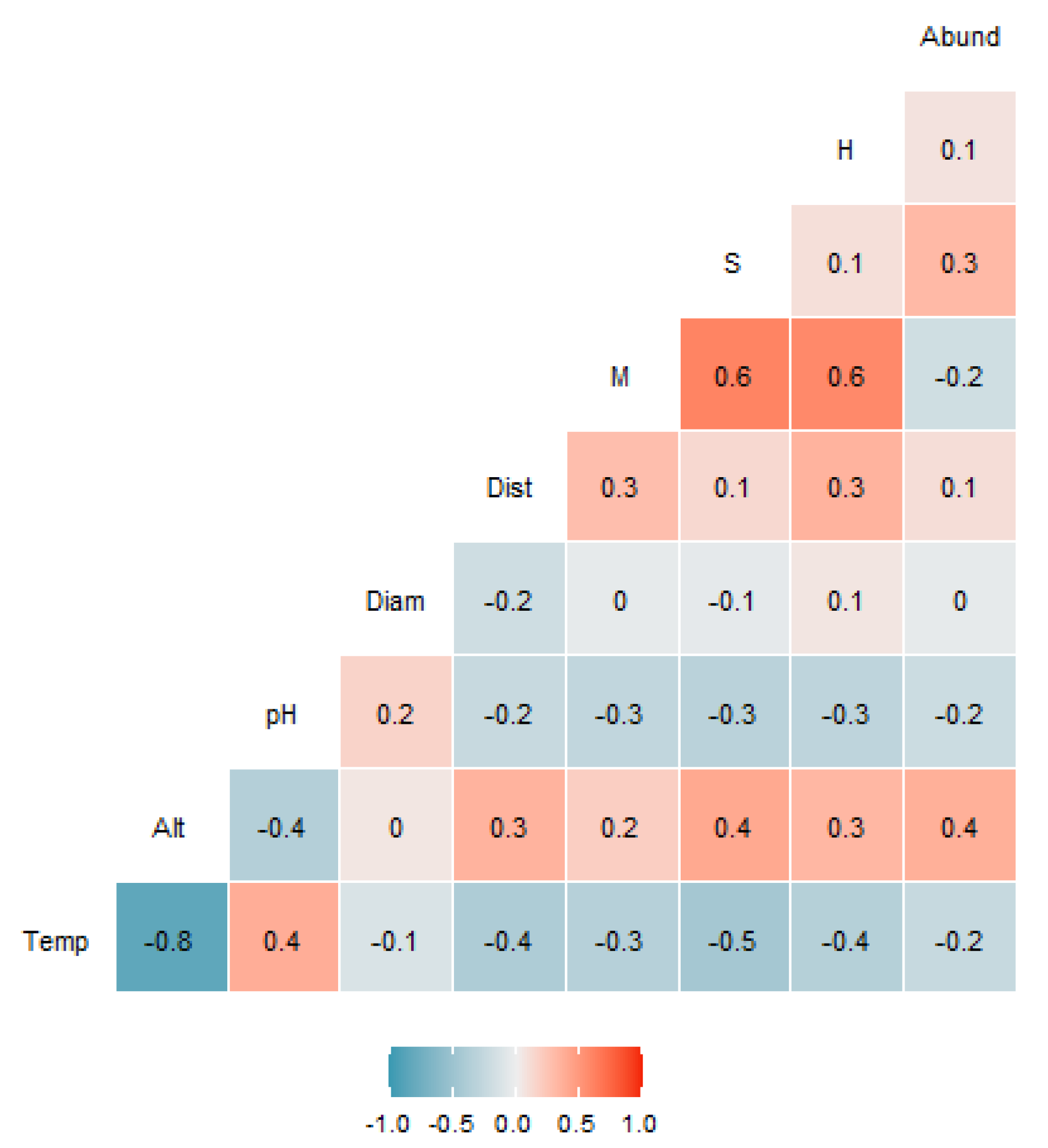
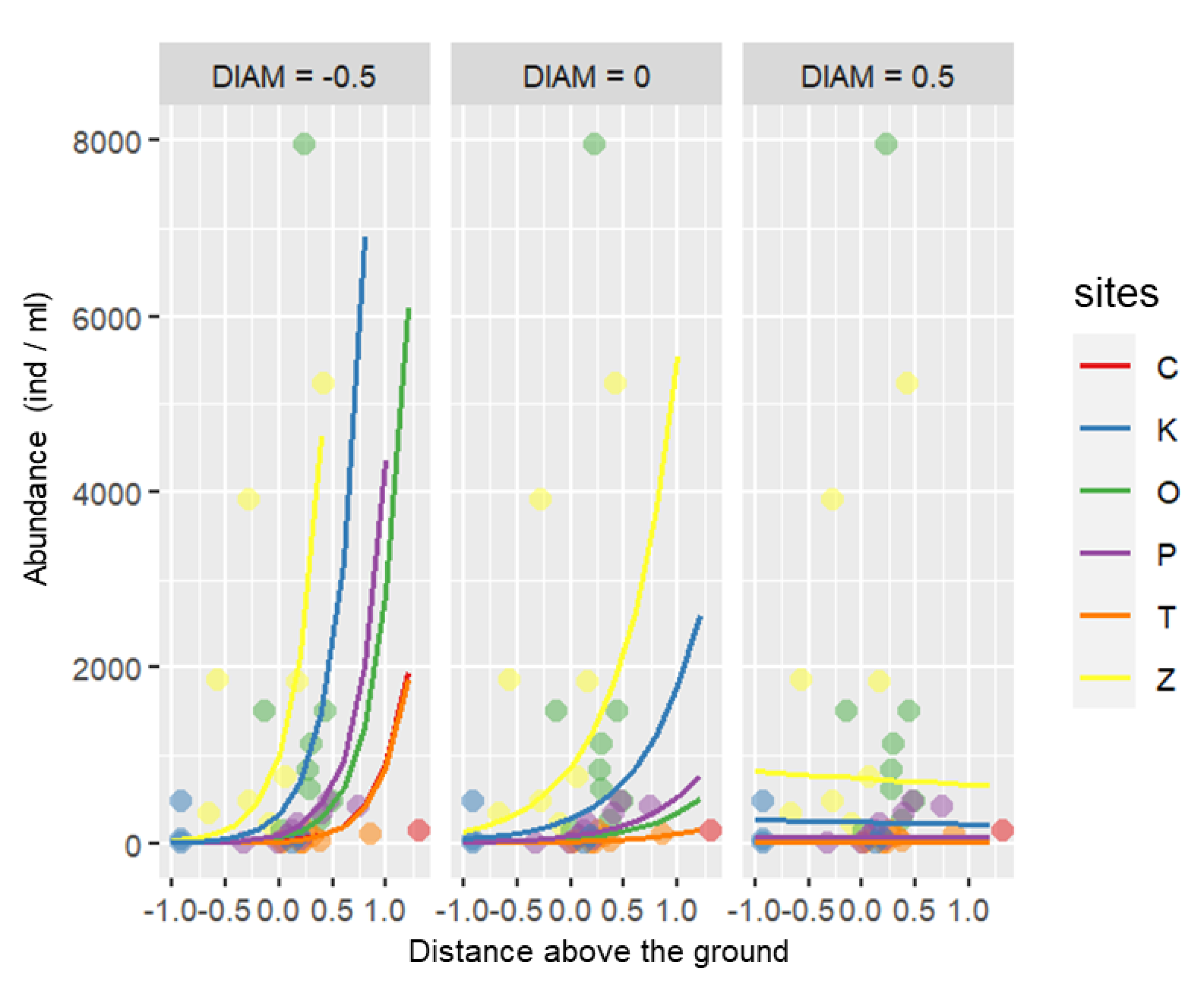
3.4. Environmental Variables and Their Effect on the Ciliate Community
4. Discussion
4.1. Species Richness
4.2. Abundance
4.3. Environmental Variables and Their Effect on the Ciliate Community
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Givnish, T.J.; Barfuss, M.H.; Van Ee, B.; Riina, R.; Schulte, K.; Horres, R.; Gonsiska, P.A.; Jabaily, R.S.; Crayn, D.M.; Smith, J.A.C.; et al. Adaptive radiation, correlated and contingent evolution, and net species diversification in Bromeliaceae. Mol. Phylogenetics Evol. 2014, 71, 55–78. [Google Scholar] [CrossRef] [PubMed]
- Nadkarni, N.M. Diversity of species and interactions in the upper tree canopy of forest ecosystems. Am. Zool. 1994, 34, 70–78. [Google Scholar] [CrossRef]
- Frank, J.H.; Lounibos, L.P. Phytotelmata: Swamps or islands? Fla. Entomol. 1987, 70, 14–20. [Google Scholar] [CrossRef]
- Kitching, R.L. Food Webs and Container Habitats: The Natural History and Ecology of Phytotelmata; Cambridge University Press: Cambridge, UK, 2000; 431p. [Google Scholar]
- Maguire, B., Jr. Phytotelmata: Biota and community structure determination in plant-held waters. Annu. Rev. Ecol. Syst. 1971, 2, 439–464. [Google Scholar] [CrossRef]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brownk, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [Green Version]
- Durán-Ramírez, C.A.; Mayén-Estrada, R.; Días, R.J.P. Checklist of ciliates (Alveolata: Ciliophora) that inhabit in bromeliads from the Neotropical Region. Zootaxa 2020, 4895, 1–36. [Google Scholar] [CrossRef]
- Dunthorn, M.; Stoeck, T.; Wolf, K.; Breiner, H.W.; Foissner, W. Diversity and endemism of ciliates inhabiting Neotropical phytotelmata. Syst Biodivers. 2012, 10, 195–205. [Google Scholar] [CrossRef]
- Foissner, W.; Strüder-Kypke, M.; van der Staay, G.; Moon-van der Staay, S.; Hackstein, J. Endemic ciliates (Protozoa, Ciliophora) from tank bromeliads (Bromeliaceae): A combined morphological, molecular, and ecological study. Eur. J. Protistol. 2003, 39, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Durán-Ramírez, C.A.; Mayén-Estrada, R. Ciliate species from tank-less bromeliads in a dry tropical forest and their geographical distribution in the Neotropics. Zootaxa 2018, 4497, 241–257. [Google Scholar] [CrossRef]
- Durán-Ramírez, C.A.; García-Franco, J.G.; Foissner, W.; Mayén-Estrada, R. Free-living ciliates from epiphytic tank bromeliads in Mexico. Eur. J. Protistol. 2015, 51, 15–33. [Google Scholar] [CrossRef]
- Lynn, D.H. The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature; Springer: Dordrecht, The Netherlands, 2008; 605p. [Google Scholar]
- Benzing, D.H. Bromeliaceae: Profile of an Adaptive Radiation; Cambridge University Press: New York, NY, USA, 2000; 655p. [Google Scholar]
- Sabagh, L.T.; Días, R.J.P.; Branco, C.W.C.; Rocha, C.F.D. News records of phoresy and hyperphoresy among treefrogs, ostracods, and ciliates in bromeliad of Atlantic forest. Biodivers. Conserv. 2011, 20, 1837–1841. [Google Scholar] [CrossRef]
- Bharti, D.; Kumar, S.; La Terza, A.; Chandra, K. Dispersal of ciliated protist cysts: Mutualism and phoresy on mites. Ecology 2020, 101, e03075. [Google Scholar] [CrossRef]
- Carrias, J.F.; Cussac, M.E.; Corbara, B. A preliminary study of freshwater protozoa in tank bromeliads. J. Trop. Ecol. 2001, 17, 611–617. [Google Scholar] [CrossRef]
- Kratina, P.; Petermann, J.S.; Marino, N.A.; MacDonald, A.A.; Srivastava, D.S. Environmental control of the microfaunal community structure in tropical bromeliads. Ecol. Evol. 2017, 7, 1627–1634. [Google Scholar] [CrossRef] [Green Version]
- Buosi, P.R.B.; Utz, L.R.P.; de Meira, B.R.; Segóvia da Silva, B.T.; Lansac-Tôha, F.M.; Lansac-Tôha, F.A.L.; Velho, L.F.M. Rainfall influence on species composition of the ciliate community inhabiting bromeliad phytotelmata. Zool. Stud. 2014, 53, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Buosi, P.R.B.; Cabral, A.F.; Utz, L.R.P.; Vieira, L.C.G.; Velho, L.F.M. Effects of seasonality and dispersal on the ciliate community inhabiting bromeliad phytotelmata in riparian vegetation of a large tropical river. J. Eukaryot. Microbiol. 2015, 62, 737–749. [Google Scholar] [CrossRef]
- Durán-Ramírez, C.A.; Mayén-Estrada, R.; Romero-Niembro, V.M. Ciliate community structure in bromeliads of different types of vegetation in eastern Mexico. Acta Protozool. 2019, 58, 155–165. [Google Scholar] [CrossRef]
- Malfatti, E.; Ferreira, P.; Utz, L.R.P. Eukaryotic communities in bromeliad phytotelmata: How do they respond to altitudinal differences? Diversity 2020, 12, 326. [Google Scholar] [CrossRef]
- Busse, A.; Antiqueira, P.A.; Neutzling, A.S.; Wolf, A.M.; Romero, G.Q.; Petermann, J.S. Different in the dark: The effect of habitat characteristics on community composition and beta diversity in bromeliad microfauna. PLoS ONE 2018, 13, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Wiackowski, K.; Kocerba-Soroka, W. Selective predation by harpacticoid copepod on ciliates in phytotelmata: A laboratory experiment. Hydrobiologia 2017, 790, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Espejo-Serna, A.; López-Ferrari, A.R. La familia Bromeliaceae en México. Bot Sci. 2018, 96, 533–554. [Google Scholar] [CrossRef]
- Morrone, J.J. Neotropical Biogeography: Regionalization and Evolution; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2017; 282p. [Google Scholar]
- Foissner, W. An update of basic light and scanning electron microscopic methods for taxonomic studies of ciliated protozoa. Int. J. Syst. Evol. Microbiol. 2014, 64, 271–292. [Google Scholar] [CrossRef] [Green Version]
- Foissner, W. Neue terrestrische und limnische Ciliaten (Protozoa, Ciliophora) aus Österreich und Deutschland. Sitzungsber Kaiserl. Akad. Wiss. Wien 1987, 195, 217–268. [Google Scholar]
- Foissner, W. Colpodea (Ciliophora); Gustav Fischer Verlag: New York, NY, USA, 1993; 798p. [Google Scholar]
- Foissner, W. Morphology and ontogenesis of Bromeliophrya brasiliensis gen. n., sp. n., a new ciliate (Protozoa: Ciliophora) from Brazilian tank bromeliads (Bromeliaceae). Acta Protozool. 2003, 42, 55–70. [Google Scholar]
- Foissner, W. Life cycle, morphology, ontogenesis, and phylogeny of Bromeliothrix metopoides nov. gen., nov. spec., a peculiar ciliate (Protista, Colpodea) from tank bromeliads (Bromeliaceae). Acta Protozool. 2010, 49, 159–193. [Google Scholar]
- Foissner, W. Description of Glaucomides bromelicola n. gen., n. sp. (Ciliophora, Tetrahymenida), a macrostome-forming inhabitant of bromeliads (Bromeliaceae), including redescriptions of Glaucoma scintillans and G. reniformis. J. Eukaryot. Microbiol. 2013, 60, 137–157. [Google Scholar] [CrossRef]
- Foissner, W. Terrestrial and semiterrestrial ciliates (Protozoa, Ciliophora) from Venezuela and Galapagos. Denisia 2016, 35, 1–912. [Google Scholar]
- Foissner, W.; Wolf, K.W.; Yashchenko, V.; Stoeck, T. Description of Leptopharynx bromelicola n. sp. and characterization of the genus Leptopharynx Mermod, 1914 (Protista, Ciliophora). J. Eukaryot. Microbiol. 2011, 58, 134–151. [Google Scholar] [CrossRef] [Green Version]
- Guggiari, M.; Peck, R. The bacterivorous ciliate Cyclidium glaucoma isolated from a sewage treatment plant: Molecular and cytological descriptions for barcoding. Eur. J. Protistol. 2008, 44, 168–180. [Google Scholar] [CrossRef]
- Lee, J.J.; Leedale, G.F.; Bradbury, P.C. An Illustrated Guide to the Protozoa, 2nd ed.; Society of Protozoologist: Lawrence, KS, USA, 2000; Volume I, 689p. [Google Scholar]
- Omar, A.; Foissner, W. Description of Leptopharynx bromeliophilus nov. spec. and Leptopharynx australiensis nov. spec. (Ciliophora, Nassulida). Acta Protozool. 2011, 50, 89–103. [Google Scholar]
- Omar, A.; Foissner, W. Neotypification and ontogenesis of Leptopharynx costatus costatus Mermod, 1914. J. Eukaryot. Microbiol. 2012, 59, 268–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penard, E. Études sur les Infusoires d’eau Douce; Georg and Cie: Geneve, Italy, 1922; 331p. [Google Scholar]
- Foissner, W. Soil Protozoa: Fundamental problems, ecological significance, adaptations in ciliates and testaceans, bioindicators, and guide to the literature. Prog. Protistol. 1987, 2, 69–212. [Google Scholar]
- Lüftenegger, G.; Petz, W.; Foissner, W.; Adam, H. The efficiency of a direct counting method in estimating the numbers of microscopic soil organisms. Pedobiologia 1988, 31, 95–101. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; 192p. [Google Scholar]
- Moreno, C.E. Métodos para Medir la Biodiversidad; M&T-Manuales y Tesis SEA: Zaragoza, Spain, 2001; 84p. [Google Scholar]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Kassambara, A. Practical Guide to Principal Component Methods in R: PCA, M (CA), FAMD, MFA, HCPC, Factoextra, Volume 2. STHDA. 2017. Available online: http://www.sthda.com/english/articles/31-principal-component-methods-in-r-practical-guide/ (accessed on 27 January 2021).
- Weisse, T. Ciliates and the rare biosphere-community ecology and population dynamics. J. Eukaryot. Microbiol. 2014, 61, 419–433. [Google Scholar] [CrossRef]
- Cavallero, L.; López, D.; Barberis, I.M. Morphological variation of Aechmea distichantha (Bromeliaceae) in a Chaco forest: Habitat and size-related effects. Plant Biol. 2009, 11, 379–391. [Google Scholar] [CrossRef]
- Caron, D.A. Past president’s address: Protistan biogeography: Why all the fuss? J. Eukaryot. Microbiol. 2009, 56, 105–112. [Google Scholar] [CrossRef]
- Caron, D.A.; Countway, P.D. Hypotheses on the role of protistan rare biosphere in a changing world. Aquat. Microb. Ecol. 2009, 57, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Caron, D.A.; Countway, P.D.; Jones, A.C.; Kim, D.Y.; Schnetzer, A. Marine protistan diversity. Ann. Rev. Mar. Sci. 2012, 4, 467–493. [Google Scholar] [CrossRef] [Green Version]
- Massana, R.; Logares, R. Eukaryotic versus prokaryotic marine picoplankton ecology. Environ. Microbiol. 2013, 15, 1254–1261. [Google Scholar] [CrossRef] [Green Version]
- Fernández, L.D.; Fournier, B.; Rivera, R.; Lara, E.; Mitchell, E.A.; Hernández, C.E. Water–energy balance, past ecological perturbations and evolutionary constraints shape the latitudinal diversity gradient of soil testate amoebae in south-western South America. Glob. Ecol. Biogeogr. 2016, 25, 1216–1227. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality Identifier | Locality | Type of Vegetation | Geographic Coordinates | Altitude (m asl) | Bromeliad Species |
|---|---|---|---|---|---|
| Locality C | Surroundings of the Estación de Biología Chamela-UNAM, inside the Reserve of the Biosphere Chamela-Cuixmala, municipality of La Huerta, Jalisco. | Dry tropical forest | 19°29′54.82″ N, 105°02′45.07″ W | 51 | Bromelia karatas L., Tillandsia makoyana Baker, T. rothii Rauh |
| Locality Z | Forest around Lake Tziscao and Lake Dos Lagunas, located at National Park Lagos de Montebello, municipality of La Independencia and Trinitaria, Chiapas. | Montane cloud forest | 16°05′47.43″ N, 91°41′04.77″ W | 1444 | T. guatemalensis L. B. Sm., T. multicaulis Steudel, Werauhia sp. |
| Locality O | Sierra Juárez, Km 175 of the road Oaxaca–Tuxtepec, near the village El Cerezal, municipality of Santa Catarina Ixtepeji, Oaxaca. | Oak forest | 17°15′33.87″ N, 96°32′28.70″ W | 2377 | T. prodigiosa Baker |
| Locality T | Forest area for conservation El Tegolome, near the town of Tlanchinol, municipality of Tlanchinol, Hidalgo. | Montane cloud forest | 21°01′19″ N, 98°38′45″ W | 1533 | T. imperialis E. Morren ex Mez, T. multicaulis Steudel, Tillandsia sp. |
| Locality P | Near the town of Cuetzalan, municipality of Cuetzalan del Progreso, Puebla. | Montane cloud forest | 20°00′21.438″ N, 97°30′30.56″ W | 983 | Catopsis sessiliflora (Ruiz and Pavón) Mez, T. heterophylla E. Morren |
| Locality K | Biocultural Reserve Kaxil Kiuic, located 27 KM southern of the town of Oxkutzcab, municipality of Oxkutzcab, Yucatán. | Semideciduous tropical forest | 20°06′10.8″ N, 89°33′43.2″ W | 92 | Aechmea bracteata (Sw.) Grisebach |
| Locality C | Locality Z | |||||||||||||||||||
| Ciliate species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| Bromeliophrya brasiliensis Foissner, 2003 | - | - | - | - | - | - | - | - | - | - | - | - | - | 5240 | - | 20 | 3900 | - | - | - |
| Bromeliothrix metopoides Foissner, 2010 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Colpoda cavicola Kahl, 1935 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Colpoda cucullus (Müller, 1773) | - | - | - | - | - | - | - | - | - | - | - | 580 | - | - | - | - | - | - | - | - |
| Colpoda lucida Greeff, 1888 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Colpoda maupasi Enriques, 1908 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Cyclidium glaucoma Müller, 1773 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Drepanomonas sphagni Kahl, 1931 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Glaucomides bromelicola Foissner, 2013 | - | - | - | - | - | - | - | - | - | - | 1800 | - | 1860 | - | - | 180 | - | - | 260 | - |
| Hymenostomatia 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 180 | - | - |
| Hymenostomatia 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Lambornella sp. | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 20 | - | - | - |
| Leptopharynx bromelicola Foissner et al., 2011 | - | - | - | - | - | - | - | - | - | - | 20 | - | - | - | - | 20 | - | - | - | - |
| Leptopharynx bromeliophilus Omar and Foissner, 2011 | - | - | - | - | - | - | - | - | - | - | - | 20 | - | - | - | - | - | - | 80 | - |
| Leptopharynx costatus Mermod, 1914 | - | 20 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Operculariidae | - | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Paracolpoda lajacola Foissner, 2016 | - | - | - | - | - | - | - | - | - | - | - | 80 | - | - | - | - | - | - | - | - |
| Pertrichia 1 | - | - | - | - | - | - | - | - | - | - | 20 | - | - | - | - | - | - | - | - | - |
| Peritrichia 2 | - | 80 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Sporadotrichida | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 300 | - | - |
| Stichotrichida | - | - | - | - | - | - | - | - | - | - | - | 80 | - | - | - | - | - | - | - | - |
| Tetrahymena sp. | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Number of species per bromeliad | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 4 | 1 | 1 | 0 | 3 | 2 | 2 | 2 | 0 |
| Total abundance of active species | 0 | 140 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1840 | 760 | 1860 | 5240 | 0 | 220 | 3920 | 480 | 340 | 0 |
| Bromeliad species | Bk | Tk | Tr | Tr | Tr | Bk | Bk | Tk | Tr | Tk | Tg | Tg | Tm | Tg | Ws | Tg | Tm | Ws | WS | Ws |
| Locality O | Locality T | |||||||||||||||||||
| Ciliate species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| Bromeliophrya brasiliensis Foissner, 2003 | 400 | - | 20 | - | 1100 | 20 | - | 60 | - | 1750 | - | - | - | - | - | - | - | - | - | - |
| Bromeliothrix metopoides Foissner, 2010 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Colpoda cavicola Kahl, 1935 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Colpoda cucullus (Müller, 1773) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Colpoda lucida Greeff, 1888 | - | - | - | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Colpoda maupasi Enriques, 1908 | - | - | 20 | - | - | - | 1520 | 20 | - | - | - | - | - | - | - | - | - | - | - | - |
| Cyclidium glaucoma Müller, 1773 | - | - | - | 540 | - | - | - | - | - | - | - | - | - | 60 | - | - | - | - | - | - |
| Drepanomonas sphagni Kahl, 1931 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Glaucomides bromelicola Foissner, 2013 | 740 | - | 140 | - | 260 | - | - | - | 620 | 6220 | - | - | 20 | 40 | 80 | 40 | 40 | 20 | 20 | 40 |
| Hymenostomatia 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Hymenostomatia 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Lambornella sp. | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Leptopharynx bromelicola Foissner et al., 2011 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | 100 | - | - | - | - |
| Leptopharynx bromeliophilus Omar and Foissner, 2011 | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Leptopharynx costatus Mermod, 1914 | - | - | - | 240 | 60 | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Operculariidae | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Paracolpoda lajacola Foissner, 2016 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Pertrichia 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Peritrichia 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Sporadotrichida | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Stichotrichida | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Tetrahymena sp. | - | - | 80 | 20 | - | 420 | - | 60 | - | - | - | - | - | - | - | - | - | - | - | - |
| Number of species per bromeliad | 2 | 0 | 4 | 4 | 4 | 3 | 1 | 3 | 1 | 2 | 0 | 0 | 1 | 2 | 2 | 2 | 1 | 1 | 1 | 1 |
| Total abundance of active species | 1140 | 0 | 260 | 840 | 1520 | 480 | 1520 | 140 | 620 | 7970 | 0 | 0 | 20 | 100 | 120 | 140 | 40 | 20 | 20 | 40 |
| Bromeliad species | To | To | To | To | To | To | To | To | To | To | Ti | Ti | Ti | T1 | Ti | Ti | Tm | Tm | T1 | T1 |
| Locality P | Locality K | |||||||||||||||||||
| Ciliate species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| Bromeliophrya brasiliensis Foissner, 2003 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Bromeliothrix metopoides Foissner, 2010 | - | 20 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Colpoda cavicola Kahl, 1935 | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - | - | - | 280 | - | - |
| Colpoda cucullus (Müller, 1773) | - | - | - | - | - | - | - | 20 | - | - | - | - | - | - | - | - | - | - | - | - |
| Colpoda lucida Greeff, 1888 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Colpoda maupasi Enriques, 1908 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 20 | - | - | - |
| Cyclidium glaucoma Müller, 1773 | - | - | - | 20 | - | 360 | - | - | - | - | - | - | 20 | - | - | - | - | 120 | - | - |
| Drepanomonas sphagni Kahl, 1931 | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Glaucomides bromelicola Foissner, 2013 | - | - | 20 | - | - | 20 | - | - | 40 | 240 | - | - | - | - | - | - | - | - | - | - |
| Hymenostomatia 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Hymenostomatia 2 | - | - | - | - | - | 20 | 20 | 80 | 120 | - | - | - | - | - | - | - | - | - | - | - |
| Lambornella sp. | - | - | - | - | - | - | - | - | 180 | 200 | - | 60 | - | - | - | - | - | - | - | - |
| Leptopharynx bromelicola Foissner et al., 2011 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Leptopharynx bromeliophilus Omar and Foissner, 2011 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | 60 | - | - |
| Leptopharynx costatus Mermod, 1914 | 20 | - | 200 | - | - | 20 | 40 | - | - | 40 | - | - | - | - | - | - | - | - | - | - |
| Operculariidae | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Paracolpoda lajacola Foissner, 2016 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Pertrichia 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Peritrichia 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Sporadotrichida | 20 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 20 | - | - |
| Stichotrichida | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Tetrahymena sp. | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Number of species per bromeliad | 3 | 1 | 2 | 1 | 0 | 4 | 2 | 2 | 3 | 3 | 0 | 2 | 1 | 0 | 0 | 1 | 1 | 4 | 0 | 0 |
| Total abundance of active species | 140 | 20 | 220 | 20 | 0 | 420 | 60 | 100 | 340 | 480 | 0 | 100 | 20 | 0 | 0 | 50 | 20 | 480 | 0 | 0 |
| Bromeliad species | Th | Th | Cs | Th | Th | Th | Th | Th | Th | Th | Ab | Ab | Ab | Ab | Ab | Ab | Ab | Ab | Ab | Ab |
| Menhinick Index (DMn) | Simpson’s Index (λ) | Shannon–Wiener Index (H’) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Estimates | CI | p | Estimates | CI | p | Estimates | CI | p | |
| Predictors | |||||||||
| Locality C | 0.025 | −0.028–0.079 | 0.349 | 0.042 | −0.178–0.262 | 0.703 | 0.095 | −0.128–0.319 | 0.396 |
| Locality K | 0.096 | 0.043–0.150 | 0.001 | 0.394 | 0.174–0.614 | 0.001 | 0.172 | −0.051–0.395 | 0.128 |
| Locality O | 0.102 | 0.049–0.156 | <0.001 | 0.581 | 0.361–0.800 | <0.001 | 0.538 | 0.315–0.761 | <0.001 |
| Locality P | 0.178 | 0.124–0.231 | <0.001 | 0.621 | 0.401–0.840 | <0.001 | 0.463 | 0.240–0.686 | <0.001 |
| Locality T | 0.153 | 0.099–0.207 | <0.001 | 0.666 | 0.446–0.886 | <0.001 | 0.191 | −0.033–0.414 | 0.093 |
| Locality Z | 0.068 | 0.015–0.122 | 0.014 | 0.639 | 0.419–0.859 | <0.001 | 0.271 | 0.048–0.494 | 0.018 |
| Abundance | |||
|---|---|---|---|
| Predictors | Incidence Rate Radios | CI | p |
| Count model | |||
| (Intercept) | 87.75 | 23.54–327.19 | <0.001 |
| Plant diameter | 0.74 | 0.15–3.68 | 0.711 |
| Distance above the ground | 6.42 | 2.02–20.47 | 0.002 |
| Altitude (asl) | 35.04 | 0.36–3454.15 | 0.129 |
| Water temperature | 0.41 | 0.00–46.41 | 0.713 |
| Plant diameter, distance above the ground | 0.02 | 0.00–0.29 | 0.004 |
| (Intercept) | 0.94 | ||
| Zero inflated model | |||
| (Intercept) | 0.28 | 0.12–0.62 | 0.002 |
| Random effects | |||
| σ2 | 5.69 | ||
| Τ00 ST | 2.17 | ||
| ICC | 0.28 | ||
| NST | 6 | ||
| Total observations | 60 | ||
| Marginal R2/Conditional R2 | 0.485/0.627 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durán-Ramírez, C.A.; Mayén-Estrada, R. Ecological Study of the Active Ciliate Community from Bromeliads during the Humid Season in Different Types of Forests of the Mexican Neotropics. Diversity 2022, 14, 122. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020122
Durán-Ramírez CA, Mayén-Estrada R. Ecological Study of the Active Ciliate Community from Bromeliads during the Humid Season in Different Types of Forests of the Mexican Neotropics. Diversity. 2022; 14(2):122. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020122
Chicago/Turabian StyleDurán-Ramírez, Carlos Alberto, and Rosaura Mayén-Estrada. 2022. "Ecological Study of the Active Ciliate Community from Bromeliads during the Humid Season in Different Types of Forests of the Mexican Neotropics" Diversity 14, no. 2: 122. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020122