
The Spread of the Invasive Locust Digitate Leafminer Parectopa robiniella Clemens, 1863 (Lepidoptera: Gracillariidae) in Europe, with Special Reference to Ukraine


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
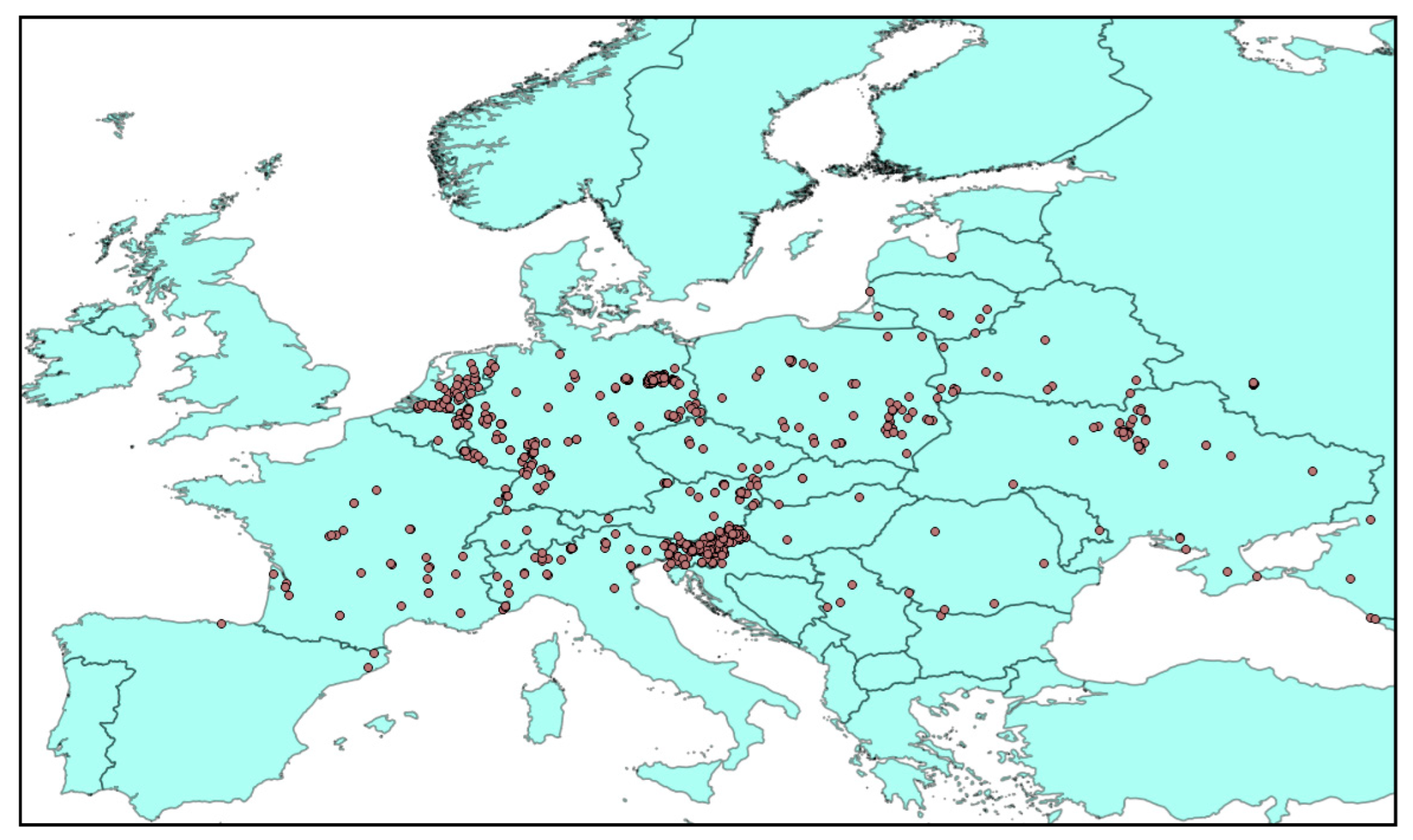
2.1. Occurrence Records and Environmental Variables
2.2. Modelling Procedure
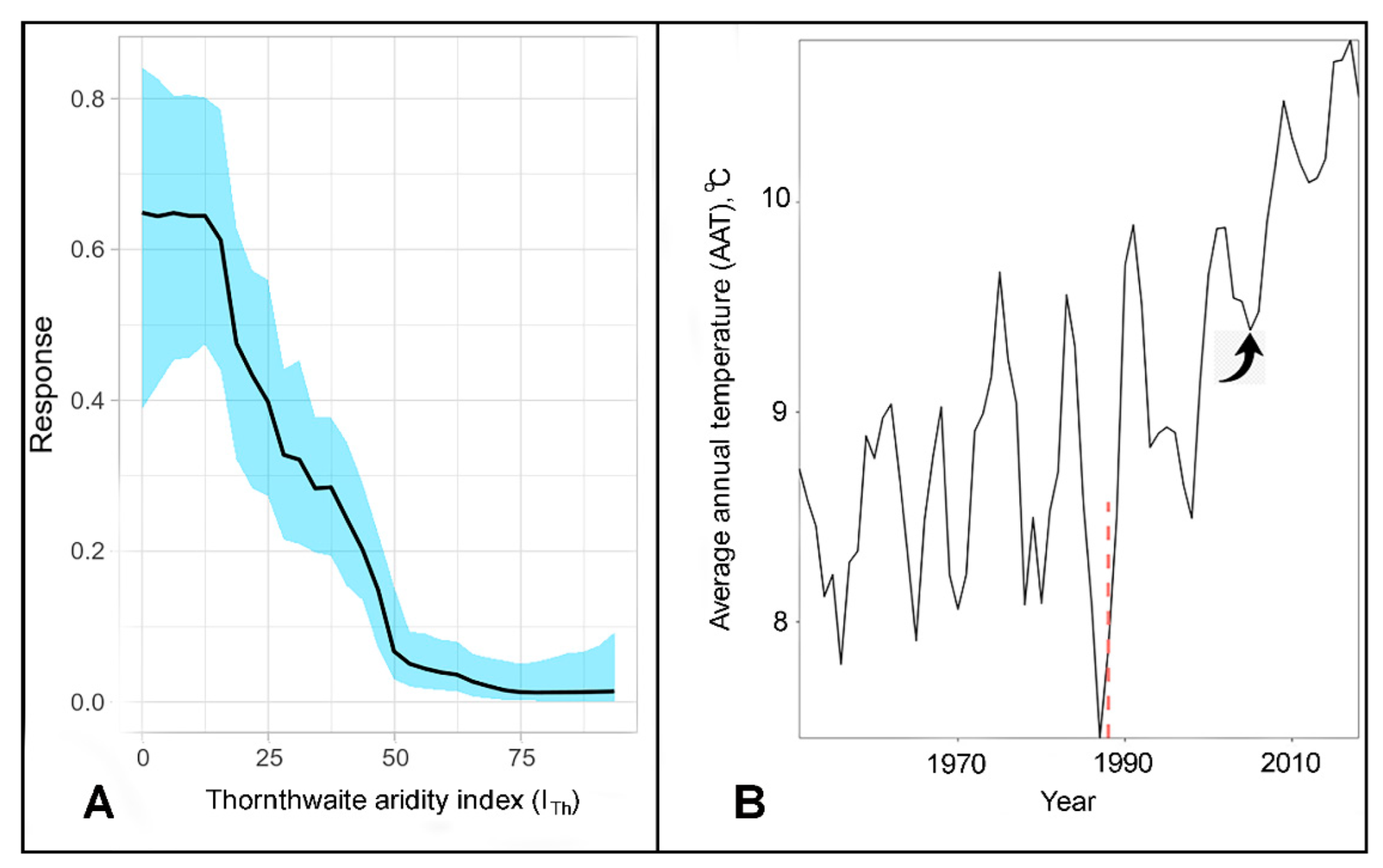
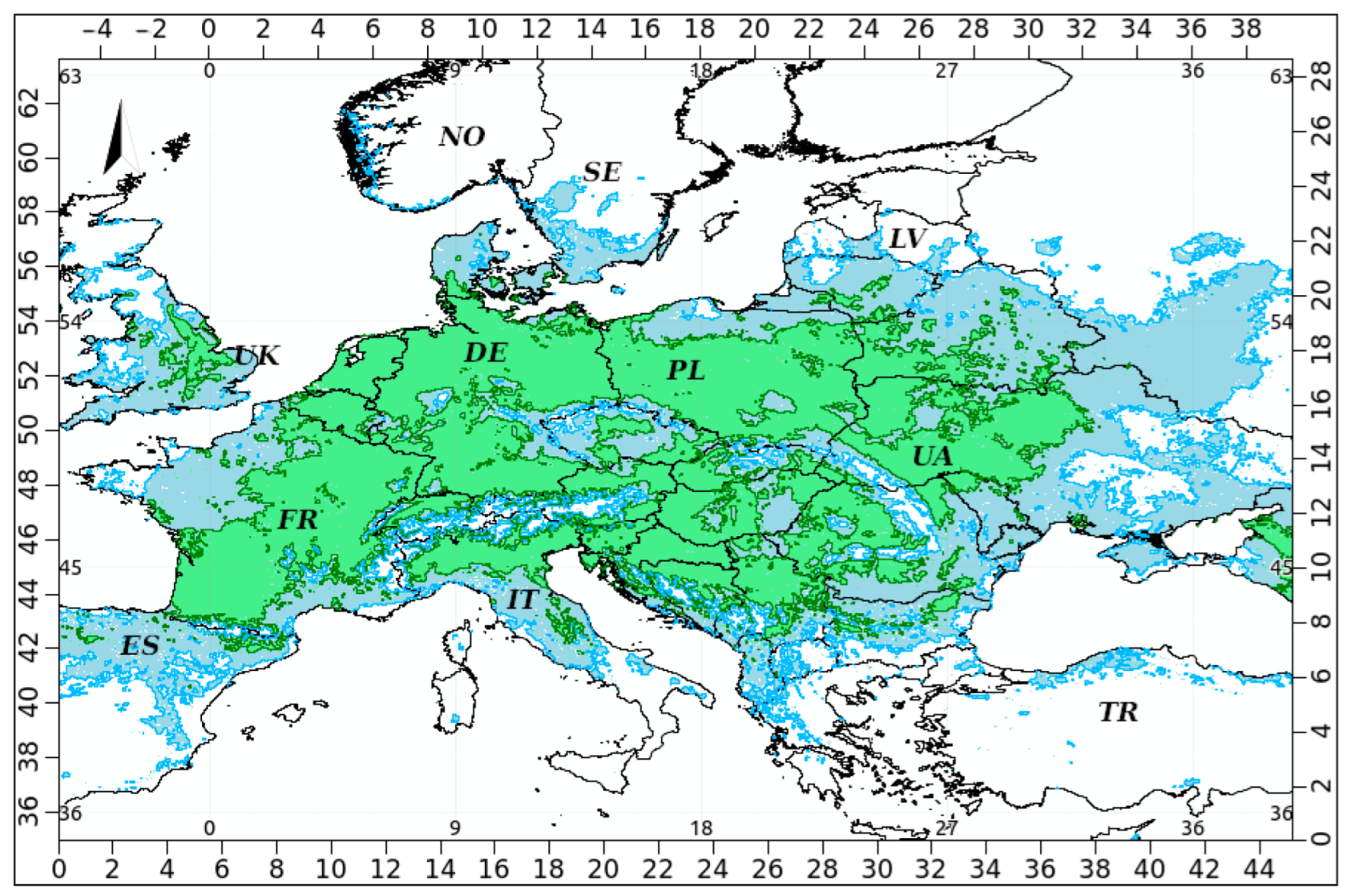
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- State of the Climate Report. 2021. Available online: https://www.ncdc.noaa.gov/sotc/ (accessed on 31 January 2022).
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- Root, T.L.; Price, J.T.; Hal, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Nekrasova, O.; Tytar, V.; Pupins, M.; Čeirāns, A.; Marushchak, O.; Skute, A. A GIS Modeling Study of the Distribution of Viviparous Invasive Alien Fish Species in Eastern Europe in Terms of Global Climate Change, as Exemplified by Poecilia reticulata Peters, 1859 and Gambusia holbrooki Girarg, 1859. Diversity 2021, 13, 385. [Google Scholar] [CrossRef]
- Nekrasova, O.; Marushchak, O.; Pupins, M.; Skute, A.; Tytar, V.; Čeirāns, A. Distribution and Potential Limiting Factors of the European Pond Turtle (Emys orbicularis) in Eastern Europe. Diversity 2021, 13, 280. [Google Scholar] [CrossRef]
- Jepsen, J.U.; Hagen, S.B.; Ims, R.A.; Yoccoz, N.G. Climate change and outbreaks of the geometrids Operophtera brumata and Epirrita autumnata in subarctic birch forest: Evidence of a recent outbreak range expansion. J. Anim. Ecol. 2008, 77, 257–264. [Google Scholar] [CrossRef]
- Vidano, C. Foglioline di Robinia pseudoacacia con mine di un microlepidopttero nuotore per l’Italia. L’Apicoltore Mod. 1970, 61, 1–11. [Google Scholar]
- Csóka, G.; Pénzes, Z.; Hirka, A.; Mikó, I.; Matošević, D. Parasitoid assemblages of two invading black locust leaf miners, Phyllonorycter robiniella and Parectopa robiniella in Hungary. Period. Biol. 2009, 111, 405–411. [Google Scholar]
- EPPO. EPPO Global Database. In EPPO Global Database; EPPO: Paris, France, 2021; Available online: https://gd.eppo.int/ (accessed on 31 January 2022).
- Gninenko, Y.I.; Kostukov, V.V.; Kosheleva, O.V. New invasive insects in the forests and greenery of the Krasnodar krai. Zashchita Karantin Rasteniĭ 2011, 4, 49–50. Available online: https://cyberleninka.ru/article/n/novye-invazivnye-nasekomye-v-lesah-i-ozelenitelnyh-posadkah-krasnodarskogo-kraya (accessed on 26 July 2022). (In Russian).
- Hrubík, P. Alien insect pests on introduced woody plants in Slovakia. Acta Entomol. Serbica 2007, 12, 81–85. [Google Scholar]
- Kollár, J. The harmful entomofauna of woody plants in Slovakia. Acta Entomol. Serbica 2007, 12, 67–79. [Google Scholar]
- Ivinskis, P.; Rimšaitė, J. Records of Phyllonorycter robiniella (Clemens, 1859) and Parectopa robiniella Clemens, 1863 (Lepidoptera, Gracillariidae) in Lithuania. Acta Zool. Litu. 2008, 8, 130–133. [Google Scholar] [CrossRef]
- Melika, G. Two invading black locust leaf miners, Parectopa robiniella and Phyllonorycter robiniella and their native parasitoid assemblages in Hungary. Biotic damage in forests. In Proceedings of the IUFRO (WP 7.03.10) Symposium, Mátrafüred, Hungary, 12–16 September 2004. [Google Scholar]
- Baugnée, J.-Y. Parectopa robiniella (Lepidoptera: Gracillariidae), a leafminer of black locust Robinia pseudoacacia new to the Belgian fauna. Phegea 2014, 42, 55–57. [Google Scholar]
- Holoborodko, K.K.; Rusynov, V.I.; Seliutina, O.V. Addition to analysis of morphological parameters of mines on two invasive leaf-mining Lepidoptera species (Parectopa robiniella Clemens, 1863 and Phyllonorycter robiniella Clemens, 1859) on black locust. Issues of bioindication and ecology 2018, 23, 134–141. [Google Scholar] [CrossRef]
- Revilla, T.; Gastón, F.J. Nuevas aportaciones a la fauna de Microlepidoptera de España y otras citas de interés (Insecta: Lepidoptera). SHILAP Rev. Lepidopterol. 2019, 47, 57–64. [Google Scholar]
- Matsiakh, I.; Kramarets, V. Invasive phyllophagous insects in Ukraine. Sci. Proc. For. Acad. Sci. Ukr. 2020, 20, 11–25. (In Ukrainian) [Google Scholar] [CrossRef]
- Shvydenko, I.M.; Stankevych, S.V.; Goroshko, V.V.; Bulat, A.G.; Cherkis, T.M.; Zabrodina, I.V.; Lezhenina, I.P.; Baidyk, H.V. Adventitious leafminer Parectopa robiniella Clemens, 1863 and Phyllonorycter robiniella Clemens, 1859 on a black locust tree in the Kharkiv region. Ukr. J. Ecol. 2021, 11, 22–32. [Google Scholar]
- Sautkin, F.V. Arthropod phytophages—Pests of Robinia (Robinia S. L.) in the conditions of Belarus. In Forestry, Nature Management and Processing. Renewable Resources.; Issue 1, BSTU: Minsk, Belarus, 2021; pp. 138–148. [Google Scholar] [CrossRef]
- Aarvik, L.; Bengt, A.; Hallvard, E.; Ivinskis, P.; Karsholt, O.; Mutanen, M.; Savenkov, N. Additions and corrections to the Nordic-Baltic Checklist of Lepidoptera. Nor. J. Entomol. 2021, 68, 1–14. [Google Scholar]
- Neţoiu, C.; Tomescu, R. Moliile miniere ale salcâmului (Parectopa robiniella Clemens, 1863 şi Phyllonorycter robiniella Clemens, 1859, Lepidoptera, Gracillariidae). An. ICAS 2006, 49, 119–131. [Google Scholar]
- Csiha, I.; Keserű, Z.; Rásó, J.; Rédei, K. Black locust (Robinia pseudoacacia L.) selection programmes in Hungary: A short review. Int. J. Hortic. Sci. 2016, 22, 31–34. [Google Scholar] [CrossRef] [Green Version]
- Rice, S.K.; Westerman, B.; Federici, R. Impacts of the exotic, nitrogen-fixing black locust (Robinia pseudoacacia) on nitrogen-cycling in a pine-oak ecosystem. Plant Ecol. 2004, 174, 97–107. [Google Scholar] [CrossRef]
- Rédei, K.; Keserű, Z.; Csiha, I.; Rásó, J.; Bakti, B.; Takács, M. Improvement of black locust (Robinia pseudoacacia L.) growing under marginal site conditions in Hungary: Case studies. Acta Agrar. Debr. 2018, 74, 129–133. [Google Scholar] [CrossRef]
- Stashenko, V.; Politshuk, V. Characteristics of black locust (Robinia pseudoacacia) as a honey plant in the middle Dnepr Region (Ukraine). Am. Bee J. 1998, 138, 665–668. [Google Scholar]
- Gninenko, Y.I.; Rakov, A.G. The Locust Digitate Leafminer Parectopa robiniella Cl.—New Invasive Phytophage. Pushkino VNIILM VPRS MOBB 2011, 1–14. Available online: http://www.vniilm.ru/docs/pdf/izdaniya/Edition-Beloakatcievaia-parektopa-robiniella-cl.pdf (accessed on 26 July 2022). (In Russian).
- Gubin, A.I.; Martynov, V.V. The first record of the locust digitate leafminer Parectopa robiniella (Clemens, 1863) (Lepidoptera: Gracillariidae) from Georgia. Euroasian Entomol. J. 2017, 16, 304–305. [Google Scholar]
- Retevoi, R. Ecological research on the Parectopa robiniella population. Curr. Trends Nat. Sci. 2018, 7, 269–273. [Google Scholar]
- Koch, F.H. Considerations regarding species distribution models for forest insects. Agric. For. Entomol. 2021, 23, 393–399. [Google Scholar] [CrossRef]
- Parectopa robiniella Clemens. 1863. Available online: https://0-doi-org.brum.beds.ac.uk/10.15468/dl.mqg95f (accessed on 10 June 2022).
- Vasyliuk, O.; Prylutskyi, O.; Marushchak, O.; Kuzemko, A.; Kutsokon, I.; Nekrasova, O.; Raes, N.; Rusin, M. An extended dataset of occurrences of species listed in Resolution 6 of the Bern Convention from Ukraine. Biodivers. Data J. 2022, 10, e84002. [Google Scholar] [CrossRef]
- Beale, C.M.; Lennon, J.J. Incorporating uncertainty in predictive species distribution modelling. Philos Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 247–258. [Google Scholar] [CrossRef]
- Huntley, J.C. Robinia pseudacacia L. (black locust). In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; Hardwoods Agricultural Handbook 654: Washington, DC, USA, 1990; Volume 2, pp. 755–761. [Google Scholar] [CrossRef] [Green Version]
- Franklin, J. Mapping Species Distributions, Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar] [CrossRef]
- Kriticos, D.J. Regional climate-matching to estimate current and future sources of biosecurity threats. Biol. Invasions 2012, 14, 1533–1544. [Google Scholar] [CrossRef]
- Venette, R.C. Climate analyses to assess risks from invasive forest insects: Simple matching to advanced models. Curr. For. Rep. 2017, 3, 255–268. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Escobar, L.E.; Lira-Noriega, A.; Medina-Vogel, G.; Peterson, A.T. Potential for spread of the white-nose fungus (Pseudogymnoascus destructans) in the Americas: Use of Maxent and NicheA to assure strict model transference. Geospat. Health 2014, 9, 221–229. [Google Scholar] [CrossRef]
- Datta, A.; Schweiger, O.; Kühn, I. Origin of climatic data can determine the transferability of species distribution models. NeoBiota 2020, 59, 61–76. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Jarošik, V.; Ota, N. Extending the suite of Bioclim variables: A proposed registry system and case study using principal components analysis. Methods Ecol. Evol. 2014, 5, 956–960. [Google Scholar] [CrossRef]
- Title, P.O.; Bemmels, J.B. ENVIREM: An expanded set of bioclimatic and topographic variables increases flexibility and improves performance of ecological niche modeling. Ecography 2018, 41, 291–307. [Google Scholar] [CrossRef] [Green Version]
- Tytar, V.M.; Baidashnikov, O. Associations between habitat quality and body size in the Carpathian land snail Vestia turgida: Species distribution model selection and assessment of performance. Zoodiversity 2021, 55, 25–40. [Google Scholar] [CrossRef]
- Robinia pseudoacacia L. Available online: https://0-doi-org.brum.beds.ac.uk/10.15468/dl.8f4mb4 (accessed on 5 February 2022).
- Nuñez, M.A.; Medley, K.A. Pine invasions: Climate predicts invasion success; something else predictsfailure. Divers. Distrib. 2011, 17, 703–713. [Google Scholar] [CrossRef]
- Rangel, T.F.; Diniz-Filho, J.A.F.; Bini, L.M. Towards an integrated computational tool for spatial analysis in macroecology and biogeography. Glob. Ecol. Biogeogr. 2006, 15, 321–327. [Google Scholar] [CrossRef]
- Lichstein, J.W.; Simons, T.R.; Shriner, S.A.; Franzreb, K.E. Spatial autocorrelation and autoregressive models in ecology. Ecol. Monogr. 2002, 72, 445–463. [Google Scholar] [CrossRef]
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.; Peterson, A.; Soberón, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Conrad, O.; Bechtel, B.; Bock, M.; Dietrich, H.; Fischer, E.; Gerlitz, L.; Wehberg, J.; Wichmann, V.; Böhner, J. System for automated geoscientific analyses (SAGA) v. 2.1.4. Geosci. Model Dev. Discuss. 2015, 8, 2271–2312. [Google Scholar] [CrossRef] [Green Version]
- Braunisch, V.; Coppes, J.; Arlettaz, R.; Suchant, R.; Schmid, H.; Bollmann, K. Selecting from correlated climate variables: A major source of uncertainty for predicting species distributions under climate change. Ecography 2013, 36, 971–983. [Google Scholar] [CrossRef]
- Leroy, B.; Meynard, C.N.; Bellard, C.; Courchamp, F. ‘virtualspecies’: An R package to generate virtual species distributions. Ecography 2016, 39, 599–607. [Google Scholar] [CrossRef]
- Carlson, C.J. ‘embarcadero’: Species distribution modelling with Bayesian additive regression trees in R. Methods Ecol. Evol. 2020, 11, 850–858. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Moreno, A.; Hasenauer, H. Spatial downscaling of European climate data. Int. J. Climatol. 2016, 36, 1444–1458. [Google Scholar] [CrossRef]
- Rumsey, D.J. Statistics for Dummies, 2nd ed.; John Wiley & Sons Inc.: New York, NY, USA, 2016. [Google Scholar]
- Mastitskiy, S.E. Analysis of Times Series with R.—Ebook; 2020; Available online: https://ranalytics.github.io/tsa-with-r (accessed on 26 July 2022). (In Russian)
- Beck, J.; Ballesteros-Mejia, L.; Nagel, P.; Kitching, I.J. Online solutions and the “Wallacean shortfall”: What does GBIF contribute to our knowledge of species’ ranges? Divers. Distrib. 2013, 19, 1043–1050. [Google Scholar] [CrossRef]
- Soberón, J.; Nakamura, M. Niches and distributional areas: Concepts, methods and assumptions. Proc. Natl. Acad. Sci. USA 2009, 106, 19644–19650. [Google Scholar] [CrossRef] [Green Version]
- Tytar, V.M. Analysis of home ranges in species: An approach based on modeling the ecological niche. Vestn. Zool. 2011, 25, 96. (In Ukrainian) [Google Scholar]
- Araújo, M.B.; Luoto, M. The importance of biotic interactions for modelling species distributions under climate change. Glob. Ecol. Biogeogr. 2007, 16, 743–753. [Google Scholar] [CrossRef]
- Lee-Yaw, A.; McCune, J.; Pironon, S.; Sheth, S.N. Species distribution models rarely predict the biology of real populations. Ecography 2022, 2022, e05877. [Google Scholar] [CrossRef]
- Li, G.; Xu, G.; Guo, K.; Du, S. Mapping the global potential geographical distribution of Black Locust (Robinia pseudoacacia L.) using herbarium data and a Maximum Entropy Model. Forests 2014, 5, 2773–2792. [Google Scholar] [CrossRef] [Green Version]
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geog. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Feddema, J.J. A revised Thornthwaite-type global climate classification. Phys. Geogr. 2005, 26, 442–466. [Google Scholar] [CrossRef]
- Dragot, C.-S.; Popovici, A.; Kucsicsa, G.; Grigorescu, I.; Dumitrascu, S. Land use and crop dynamics related to climate change signals during the post-communist period in the south Oltenia, Romania. Proc. Rom. Acad. Ser. B 2013, 15, 265–278. [Google Scholar]
- Mally, R.; Ward, S.F.; Trombik, J.; Buszko, J.; Medzihorsky, V.; Liebhold, A.M. Non-native plant drives the spatial dynamics of its herbivores: The case of black locust (Robinia pseudoacacia) in Europe. NeoBiota 2021, 69, 155–175. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M. Modeling of species distributions with MaxEnt: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Nakazato, T.; Warren, D.L.; Moyle, L.C. Ecological and geographic modes of species divergence in wild tomatoes. Am. J. Bot. 2010, 97, 680–693. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WorldClim v.2 | CliMond v1.2 | ENVIREM | |||||||
|---|---|---|---|---|---|---|---|---|---|
| SDMs * | P | R | P+R | P | R | P+R | P | R | P+R |
| AUC | 0.935 | 0.770 | 0.936 | 0.936 | 0.772 | 0.926 | 0.944 | 0.772 | 0.943 |
| ±SE | 0.009 | 0.010 | 0.009 | 0.012 | 0.011 | 0.012 | 0.009 | 0.010 | 0.008 |
| WorldClim v.2 | CliMond v.1.2 | ENVIREM |
|---|---|---|
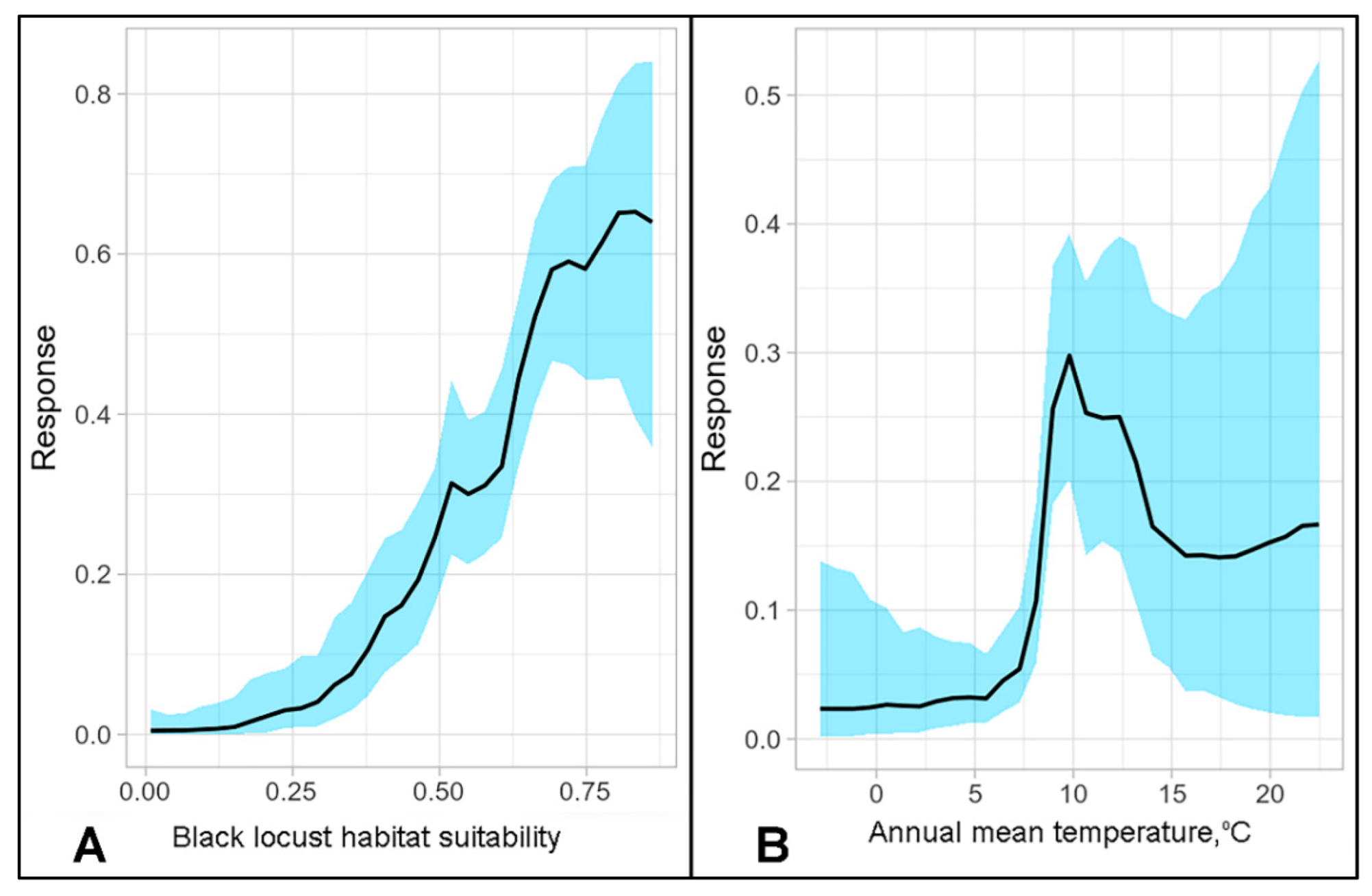
| 1. Annual mean temperature 2. Host habitat suitability 3. Temperature annual range 4. Annual mean precipitation | 1. Temperature seasonality 2. Annual mean temperature 3. Host habitat suitability | 1. Host habitat suitability 2. Thornthwaite aridity index (ITh) 3. gdd0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tytar, V.; Nekrasova, O.; Marushchak, O.; Pupins, M.; Skute, A.; Čeirāns, A.; Kozynenko, I. The Spread of the Invasive Locust Digitate Leafminer Parectopa robiniella Clemens, 1863 (Lepidoptera: Gracillariidae) in Europe, with Special Reference to Ukraine. Diversity 2022, 14, 605. https://0-doi-org.brum.beds.ac.uk/10.3390/d14080605
Tytar V, Nekrasova O, Marushchak O, Pupins M, Skute A, Čeirāns A, Kozynenko I. The Spread of the Invasive Locust Digitate Leafminer Parectopa robiniella Clemens, 1863 (Lepidoptera: Gracillariidae) in Europe, with Special Reference to Ukraine. Diversity. 2022; 14(8):605. https://0-doi-org.brum.beds.ac.uk/10.3390/d14080605
Chicago/Turabian StyleTytar, Volodymyr, Oksana Nekrasova, Oleksii Marushchak, Mihails Pupins, Arturs Skute, Andris Čeirāns, and Iryna Kozynenko. 2022. "The Spread of the Invasive Locust Digitate Leafminer Parectopa robiniella Clemens, 1863 (Lepidoptera: Gracillariidae) in Europe, with Special Reference to Ukraine" Diversity 14, no. 8: 605. https://0-doi-org.brum.beds.ac.uk/10.3390/d14080605