

Conservation of Host, Translocation of Parasites—Monitoring of Helminths during Population Reinforcement of the European Ground Squirrel (Spermophilus citellus)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reinforcement Process
2.2. Fecal Samples Collection
2.3. FLOTAC Calibration and Processing
2.4. Helminthological Examination
2.5. Statistical Analyses
3. Results
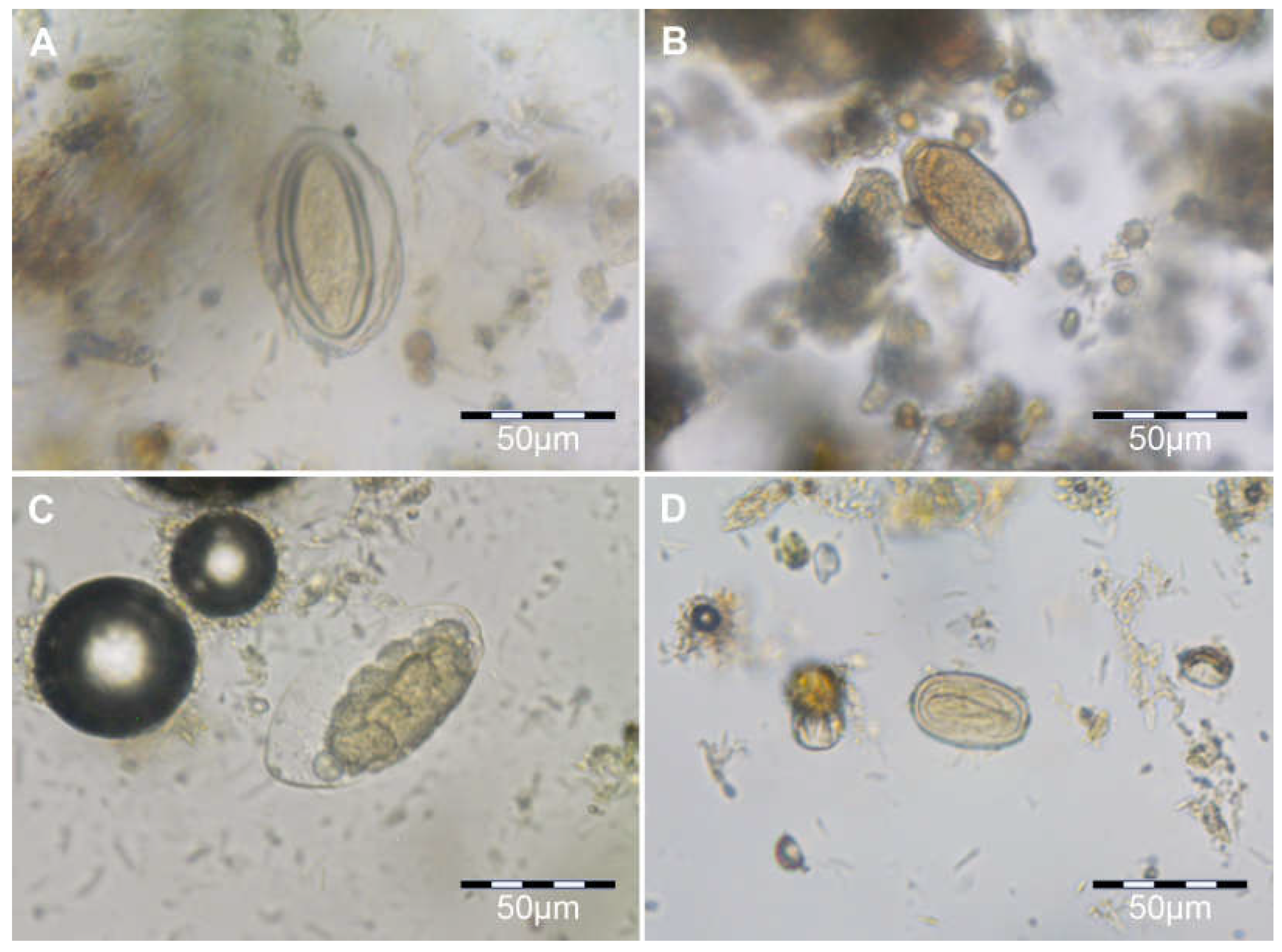
3.1. Helminth Diversity
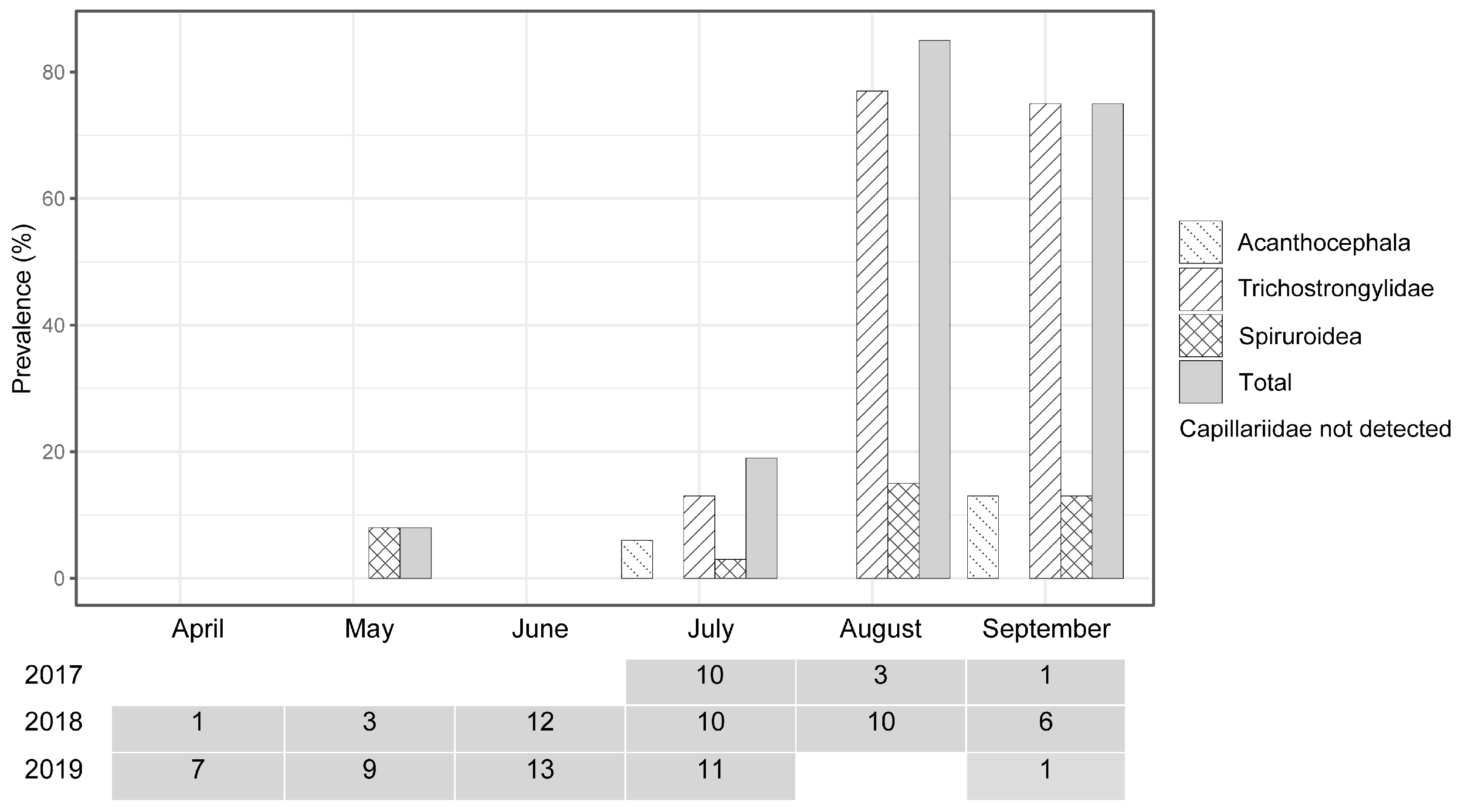
3.2. Dynamics of the Helminth Prevalence in Chronological Order
3.3. Seasonal, Age-, and Sex-Related Variations in Helminth Prevalence
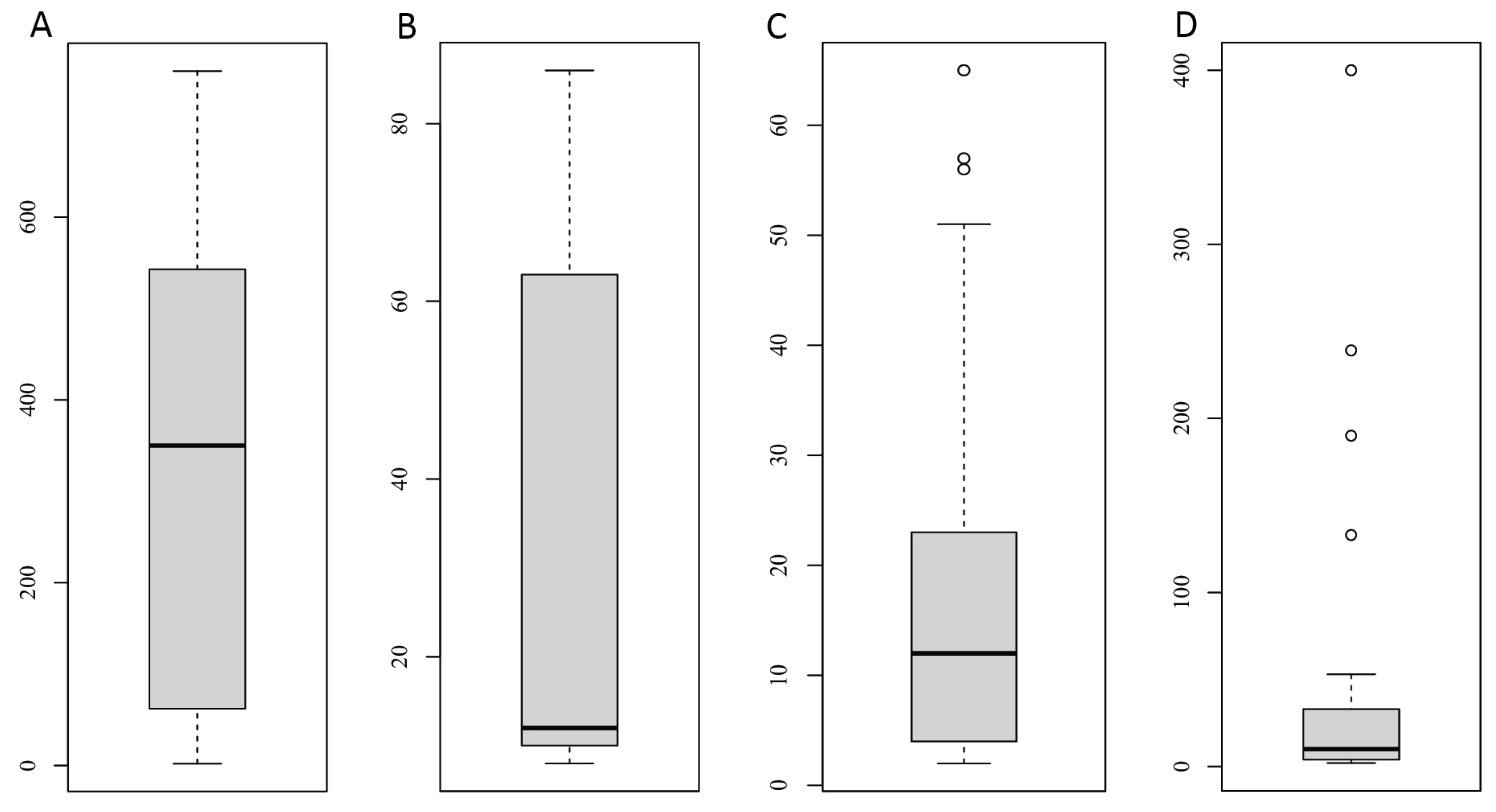
3.4. Variations in the EPG
4. Discussion
4.1. Helminth Diversity
4.2. Helminth Prevalence after the Translocation
4.3. Seasonal, Age-, and Sex- Related Variations in Helminth Prevalence
4.4. Variations in EPG
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aleuy, O.A.; Ruckstuhl, K.; Hoberg, E.P.; Veitch, A.; Simmons, N.; Kutz, S.J. Diversity of Gastrointestinal Helminths in Dall’s Sheep and the Negative Association of the Abomasal Nematode, Marshallagia marshalli, with Fitness Indicators. PLoS ONE 2018, 13, e0192825. [Google Scholar] [CrossRef] [PubMed]
- Gooderham, K.; Schulte-Hostedde, A. Macroparasitism Influences Reproductive Success in Red Squirrels (Tamiasciurus hudsonicus). Behav. Ecol. 2011, 22, 1195–1200. [Google Scholar] [CrossRef]
- Viggers, K.L.; Lindenmayer, D.B.; Spratt, D.M. The Importance of Disease in Reintroduction Programmes. Wildl. Res. 1993, 20, 687–698. [Google Scholar] [CrossRef]
- Cunningham, A.A. Disease Risks of Wildlife Translocations. Conserv. Biol. 1996, 10, 349–353. [Google Scholar] [CrossRef]
- Pedersen, A.B.; Jones, K.E.; Nunn, C.L.; Altizer, S. Infectious Diseases and Extinction Risk in Wild Mammals. Conserv. Biol. 2007, 21, 1269–1279. [Google Scholar] [CrossRef]
- Almberg, E.S.; Cross, P.C.; Dobson, A.P.; Smith, D.W.; Hudson, P.J. Parasite Invasion Following Host Reintroduction: A Case Study of Yellowstone’s Wolves. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 2840–2851. [Google Scholar] [CrossRef]
- Jørgensen, D. Conservation Implications of Parasite Co-Reintroduction. Conserv. Biol. 2015, 29, 602–604. [Google Scholar] [CrossRef]
- Ballweber, L.R.; Beugnet, F.; Marchiondo, A.A.; Payne, P.A. American Association of Veterinary Parasitologists’ Review of Veterinary Fecal Flotation Methods and Factors Influencing Their Accuracy and Use—Is There Really One Best Technique? Vet. Parasitol. 2014, 204, 73–80. [Google Scholar] [CrossRef]
- Dryden, M.W.; Payne, P.A.; Ridley, R.; Smith, V. Comparison of Common Fecal Flotation Techniques for the Recovery of Parasite Eggs and Oocysts. Vet. Ther. 2005, 6, 15–28. [Google Scholar]
- Rinaldi, L.; Coles, G.C.; Maurelli, M.P.; Musella, V.; Cringoli, G. Calibration and Diagnostic Accuracy of Simple Flotation, McMaster and FLOTAC for Parasite Egg Counts in Sheep. Vet. Parasitol. 2011, 177, 345–352. [Google Scholar] [CrossRef]
- D’Ovidio, D.; Rinaldi, L.; Ianniello, D.; Donnelly, T.M.; Pepe, P.; Capasso, M.; Cringoli, G. FLOTAC for Diagnosis of Endo-Parasites in Pet Squirrels in Southern Italy. Vet. Parasitol. 2014, 200, 221–224. [Google Scholar] [CrossRef]
- D’Ovidio, D.; Noviello, E.; Pepe, P.; Del Prete, L.; Cringoli, G.; Rinaldi, L. Survey of Hymenolepis spp. in Pet Rodents in Italy. Parasitol. Res. 2015, 114, 4381–4384. [Google Scholar] [CrossRef]
- Arok, M.; Nikolić, T.; Győri-Koósz, B.; Milinski, L.; Ćirović, D. Diet of the European Ground Squirrel (Spermophilus citellus) in the Southern Pannonian Plain. Arch. Biol. Sci. 2021, 73, 111–122. [Google Scholar] [CrossRef]
- Říčanová, Š.; Koshev, Y.; Říčan, O.; Ćosić, N.; Ćirović, D.; Sedláček, F.; Bryja, J. Multilocus Phylogeography of the European Ground Squirrel: Cryptic Interglacial Refugia of Continental Climate in Europe. Mol. Ecol. 2013, 22, 4256–4269. [Google Scholar] [CrossRef]
- Janák, M.; Marhoul, P.; Matějů, J. Action Plan for the Conservation of the European Ground Squirrel Spermophilus citellus in the European Union; European Commission: Brussels, Belgium, 2013.
- Stefanov, V. European Ground Squirrel (Spermophilus citellus Linnaeus, 1776). In Red Data Book of the Republic of Bulgaria. Part 2: Animals; Golemansky, V., Peev, D., Eds.; BAS & MOEW: Sofia, Bulgaria, 2015; p. 232. [Google Scholar]
- Koshev, Y. Action Plan for the Conservation of the European Ground Squirrel (Spermophilus citellus) in Bulgaria 2022–2031; MoEW, BSPB: Sofia, Bulgaria, 2022. [Google Scholar]
- Golemansky, V.G.; Koshev, Y.S. Coccidian Parasites (Eucoccidia: Eimeriidae) in European Ground Squirrel (Spermophilus citellus L., 1766) (Rodentia: Sciuridae) from Bulgaria. Acta Zool. Bulg. 2007, 59, 81–85. [Google Scholar]
- Diakou, A.; Kapantaidakis, E.; Youlatos, D. Endoparasites of the European Ground Squirrel (Spermophilus citellus) (Rodentia: Sciuridae) in Central Macedonia, Greece. J. Nat. Hist. 2014, 49, 359–370. [Google Scholar] [CrossRef]
- Tenora, F.; Barus, V. Hymenolepis Megaloon (Linstow, 1901) Parasitizing Citellus citellus L. in Czechoslovakia. Folia Parasitol. 1972, 19, 272. [Google Scholar]
- Genov, T. Helminths of Insectivores and Rodents in Bulgaria; Bulgarian Academy of Sciences: Sofia, Bulgaria, 1984. [Google Scholar]
- Stefanov, V.; Georgiev, B.; Genov, T.; Chipev, N. New Data on the Species Composition and Distribution of the Helminth Parasites of Spermophilus citellus L. (Rodentia, Sciuridae) in Bulgaria. In Proceedings of the Sixth National Conference of Parasitology, Sofia, Bulgaria, 5–7 October 2001. [Google Scholar]
- Simitch, T.; Petrovitch, Z. Ce Qu’il Advient Avec Les Helminthes Du Citellus citellus Au Cours Du Sommeil Hibernal de Ce Rongeur. Riv. Parassitol. 1954, 15, 655–662. [Google Scholar]
- Golemansky, V.G.; Koshev, Y.S. Systematic and Ecological Survey on Coccidians (Apicomplexa: Eucoccidida) in European Ground Squirrel (Spermophilus citellus L.) (Rodentia: Sciuridae) from Bulgaria. Acta Zool. Bulg. 2009, 61, 143–150. [Google Scholar]
- Rhodes, C.J.; Anderson, R.M. Contact Rate Calculation for a Basic Epidemic Model. Math. Biosci. 2008, 216, 56–62. [Google Scholar] [CrossRef]
- Kachamakova, M.; Koshev, Y.; Millesi, E. Resident European Ground Squirrels Exhibit Higher Stress Levels than Translocated Individuals after Conservation Reinforcement. Mamm. Biol. 2021, 101, 127–136. [Google Scholar] [CrossRef]
- Koshev, Y.; Kachamakova, M.; Arangelov, S.; Ragyov, D. Translocations of European Ground Squirrel (Spermophilus citellus) along Altitudinal Gradient in Bulgaria—An Overview. Nat. Conserv. 2019, 35, 63–95. [Google Scholar] [CrossRef]
- Kachamakova, M.; Koshev, Y.; Rammou, D.-L.; Spasov, S. Rise and Fall: Results of a Multidisciplinary Study and 5-Year Long Monitoring of Conservation Translocation of the European Ground Squirrel. Biodivers. Data J. 2022, 10, e83321. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.L.; Liu, G.; Wang, W.X.; Zhou, R.; Liu, S.Q.; Li, L.H.; Hu, D.F. Methods of Preservation and Flotation for the Detection of Nematode Eggs and Coccidian Oocysts in Faeces of the Forest Musk Deer. J. Helminthol. 2016, 90, 680–684. [Google Scholar] [CrossRef]
- Cringoli, G.; Rinaldi, L.; Maurelli, M.P.; Utzinger, J. FLOTAC: New Multivalent Techniques for Qualitative and Quantitative Copromicroscopic Diagnosis of Parasites in Animals and Humans. Nat. Protoc. 2010, 5, 503–515. [Google Scholar] [CrossRef]
- Cringoli, G. FLOTAC Manual Appendix N.1. Flotation Solutions and Parasitic Elements; Veterinary Parasitology and Parasitic Diseases Department of Pathology and Animal Health Faculty of Veterinary Medicine, University of Naples Federico II: Naples, Italy, 2009. [Google Scholar]
- Czapski, Z. Fauna Robaków Pasożytniczych Przewodu Pokarmowego u Susła Moręgowanego Citellus citellus L. Wiad. Parazytol. 1958, 4, 697–699. [Google Scholar] [PubMed]
- Petrotschenko, V.I. Acanthocephala of Domestic and Wild Animals; Academii Nauk SSSR: Moscow, Russia, 1958; Volume 2, p. 459. [Google Scholar]
- Kozlov, D. Keys to the Helminths of Carnivores of the USSR; Nauka: Moscow, Russia, 1977; p. 275. [Google Scholar]
- Bettiol, S.; Goldsmid, J.M. A Case of Probable Imported Moniliformis moniliformis Infection in Tasmania. J. Travel Med. 2000, 7, 336–337. [Google Scholar] [CrossRef]
- Anderson, R.C. Nematode Parasites of Vertebrates: Their Development and Transmission; CABI: Wallingford, UK, 2000; ISBN 978-0-85199-786-5. [Google Scholar] [CrossRef]
- Wertheim, G. Cuticular Markings in Species Differentiation of Streptopharagus (Nematoda-Spiruroidea) Parasitic in Rodents. Ann. Parasitol. Hum. Comp. 1993, 68, 49–60. [Google Scholar] [CrossRef]
- Chabaud, A.G. Sur Le Cycle Évolutif Des Spirurides et de Nématodes Ayant Une Biologie Comparable. Valeur Systématique Des Caractères Biologiques. Ann. Parasitol. Hum. Comp. 1954, 29, 42–88. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; May, R.M. Regulation and Stability of Host-Parasite Population Interactions: I. Regulatory Processes. J. Anim. Ecol. 1978, 47, 219–247. [Google Scholar] [CrossRef]
- Arneberg, P. An Ecological Law and Its Macroecological Consequences as Revealed by Studies of Relationships between Host Densities and Parasite Prevalence. Ecography 2001, 24, 352–358. [Google Scholar] [CrossRef]
- Marcogliese, D.J. Parasites of the Superorganism: Are They Indicators of Ecosystem Health? Int. J. Parasitol. 2005, 35, 705–716. [Google Scholar] [CrossRef]
- Cahill, J.E.; Lewert, R.M.; Jaroslow, B.N. Effect of Hibernation on Course of Infection and Immune Response in Citellus tridecemlineatus Infected with Nippostrongylus brasiliensis. J. Parasitol. 1967, 53, 110–115. [Google Scholar] [CrossRef]
- Ford, B.R. Hymenolepis citelli: Development and Chemical Composition in Hypothermic Ground Squirrels. Exp. Parasitol. 1972, 32, 62–70. [Google Scholar] [CrossRef]
- Callait, M.-P.; Gauthier, D. Parasite Adaptations to Hibernation in Alpine Marmots (Marmota marmota). In Life in the Cold; Heldmaier, G., Klingenspor, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 139–146. [Google Scholar] [CrossRef]
- Hillegass, M.A.; Waterman, J.M.; Roth, J.D. The Influence of Sex and Sociality on Parasite Loads in an African Ground Squirrel. Behav. Ecol. 2008, 19, 1006–1011. [Google Scholar] [CrossRef]
- Anderson, R.M.; Schad, G.A. Hookworm Burdens and Faecal Egg Counts: An Analysis of the Biological Basis of Variation. Trans. R. Soc. Trop. Med. Hyg. 1985, 79, 812–825. [Google Scholar] [CrossRef]
- Sinniah, B. Daily Egg Production of Ascaris lumbricoides: The Distribution of Eggs in the Faeces and the Variability of Egg Counts. Parasitology 1982, 84, 167–175. [Google Scholar] [CrossRef]
- Berger-Tal, O.; Blumstein, D.T.; Swaisgood, R.R. Conservation Translocations: A Review of Common Difficulties and Promising Directions. Anim. Conserv. 2020, 23, 121–131. [Google Scholar] [CrossRef]
- Nichols, E.; Gómez, A. Conservation Education Needs More Parasites. Biol. Conserv. 2011, 144, 937–941. [Google Scholar] [CrossRef]
- Carlson, C.J.; Hopkins, S.; Bell, K.C.; Doña, J.; Godfrey, S.S.; Kwak, M.L.; Lafferty, K.D.; Moir, M.L.; Speer, K.A.; Strona, G.; et al. A Global Parasite Conservation Plan. Biol. Conserv. 2020, 250, 108596. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Captured Individuals | ||
|---|---|---|
| Recapture Sessions | Resident | Translocated |
| 2017: Jul | 13 | 12 |
| 2017: Aug | 3 | 9 |
| 2017: Sep | 2 | 6 |
| 2018: Apr | 2 | 5 |
| 2018: May | 4 | 6 |
| 2018: Jun | 18 | 1 |
| 2018: Jul | 22 | 2 |
| 2018: Aug | 17 | 12 |
| 2018: Sep | 7 | 2 |
| 2019: Apr | 7 | 5 |
| 2019: May | 9 | 3 |
| 2019: Jun | 13 | 0 |
| 2019: Jul | 11 | 0 |
| 2019: Sep * | 1 | 7 |
| Flotation Solution | Specific Gravity | Floating Parasitic Elements (Mean ± SE) |
|---|---|---|
| Sucrose and formaldehyde | 1.2 | 6.8 (±5) |
| Sodium chloride | 1.2 | 0.3 (±0.2) |
| Zinc sulphate | 1.2 | 0 |
| Sodium nitrate | 1.2 | 0 |
| Sucrose and potassium iodomercurate (Rinaldi’s solution) | 1.25 | 0 |
| Magnesium sulphate | 1.28 | 11 (±2) |
| Zinc sulphate | 1.35 | 30 (±4) |
| Potassium iodomercurate | 1.44 | 1.6 (±0.4) |
| Zinc sulphate and potassium iodomercurate | 1.45 | 0.2 (±0.2) |
| Groups | Periods | N | Response Variable | Explanatory Variables | Model | Significant and Nearly Significant Variables | |
|---|---|---|---|---|---|---|---|
| 1 | T + R | 2017: Jul | 30 | Prevalence—all helminths | Sex, age, origin | Binomial glm | |
| 2 | R | 2017: Jul–Aug 2018: Jul–Aug | 33 | Prevalence—all helminths | Year, sex, age | Binomial glm | Year: z = 3.027, p = 0.002 (2018 > 2017) |
| 3 | T | 2017: Jul 2018: Jun–Jul | 32 | Prevalence—all helminths | Sex, age, year | Binomial glm | – |
| 4 | T | 2017: Jul 2018: Jun–Jul | 32 | Prevalence—Spiruroidea | Sex, age, year | Binomial glm | Year: z = 2.171, p = 0.030 (2017 > 2018) Sex: z = −2.137, p = 0.033 (m < f) Age: z = −1.747, p = 0.080 (juv < ad) |
| 5 | T | 2017: Jul 2018: Jun–Jul | 32 | Prevalence—Trichostrongylidae | Sex, age, year | Binomial glm | Year: z = −1.945, p = 0.05 (2017 > 2018) |
| 6 | T + R | 2017: Aug–Sep 2018: Aug–Sep 2019: Aug–Sep | 47 | Prevalence—all helminths | Sex, age, year, month, origin | Binomial glm | – |
| 7 | T + R | 2017: Aug–Sep 2018: Aug–Sep 2019: Aug–Sep | 47 | Prevalence—Spiruroidea | Sex, age, year, month, origin | Binomial glm | Origin: z = 2.221, p = 0.026 (T > R) |
| 8 | T + R | 2017: Aug–Sep 2018: Aug–Sep 2019: Aug–Sep | 47 | Prevalence—Trichostrongylidae | Sex, age, year, month, origin | Binomial glm | – |
| 9 | T + R | 2018: Apr–May 2019: Apr–May | 35 | Prevalence—all helminths | Sex, year, origin | Binomial glm | Origin: z = 2.201, p = 0.028 (T > R) |
| Period | Acanthocephala | Capillariidae | Trichostrongylidae | Spiruroidea | Sample Size | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| R | T | R | T | R | T | R | T | R | T | |
| 2017: Jul | 10 | 0 | 0 | 12 | 0 | 47 | 0 | 12 | n = 10 | n = 17 |
| 2017: Aug and Sep | 0 | 10 | 0 | 10 | 25 | 80 | 0 | 70 | n = 4 | n = 12 |
| 2018: Apr, May and Jun | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 33 | n = 16 | n = 12 |
| 2018: Jul | 10 | 20 | 0 | 13 | 40 | 13 | 10 | 53 | n = 10 | n = 15 |
| 2018: Aug and Sep | 6 | 14 | 0 | 0 | 88 | 57 | 19 | 43 | n = 16 | n = 7 |
| 2019: Apr, May and Jun | 0 | 0 | 0 | 0 | 0 | 38 | 29 | 13 | n = 7 | n = 8 |
| 2019: Jul * | 0 | - | 0 | - | 0 | - | 0 | - | n = 12 | n = 0 |
| 2019: Sep | 0 | - | 0 | - | 100 | - | 14 | - | n = 7 | n = 0 |
| Month | Acanthocephala | Capillariidae | Trichostrongylidae | Spiruroidea | Sample Size | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Juv | Ad | Juv | Ad | Juv | Ad | Juv | Ad | Juv | Ad | |
| Jun | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 22 | n = 21 | n = 9 |
| Jul | 7 | 9 | 2 | 14 | 27 | 23 | 10 | 27 | n = 41 | n = 22 |
| Aug | 0 | 10 | 0 | 10 | 67 | 80 | 20 | 50 | n = 15 | n = 10 |
| Sep | 12 | 0 | 0 | 0 | 76 | 100 | 29 | 20 | n = 17 | n = 5 |
| Groups | Periods | N | Response Variable | Explanatory Variables | Model | Significant and Nearly Significant Variables | |
|---|---|---|---|---|---|---|---|
| 1 | T | 2017: Jul 2018: Jun–Jul | 19 | EPG—all helminths | Sex, age, year | quasi–Poisson glm | Year: z = 2.215; p = 0.041 (2018 > 2017) |
| 2 | T + R | 40 | EPG—all helminths | Sex, age, year, month, origin | quasi–Poisson glm | – | |
| 2017: Aug–Sep 2018: Aug–Sep 2019: Aug–Sep | |||||||
| 3 | T + R | 2017: Aug–Sep 2018: Aug–Sep 2019: Aug–Sep | 15 | EPG—Spiruroidea | Sex, age, year, month, origin | quasi–Poisson glm | Age: z = −2.393, p = 0.021 (juv < ad) |
| 4 | T + R | 2017: Aug–Sep 2018: Aug–Sep 2019: Aug–Sep | 36 | EPG— Trichostron–gylidae | Sex, age, year, month, origin | quasi–Poisson glm | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kachamakova, M.; Mutafchiev, Y.; Nikolov, P.N.; Koshev, Y. Conservation of Host, Translocation of Parasites—Monitoring of Helminths during Population Reinforcement of the European Ground Squirrel (Spermophilus citellus). Diversity 2023, 15, 266. https://0-doi-org.brum.beds.ac.uk/10.3390/d15020266
Kachamakova M, Mutafchiev Y, Nikolov PN, Koshev Y. Conservation of Host, Translocation of Parasites—Monitoring of Helminths during Population Reinforcement of the European Ground Squirrel (Spermophilus citellus). Diversity. 2023; 15(2):266. https://0-doi-org.brum.beds.ac.uk/10.3390/d15020266
Chicago/Turabian StyleKachamakova, Maria, Yasen Mutafchiev, Pavel N. Nikolov, and Yordan Koshev. 2023. "Conservation of Host, Translocation of Parasites—Monitoring of Helminths during Population Reinforcement of the European Ground Squirrel (Spermophilus citellus)" Diversity 15, no. 2: 266. https://0-doi-org.brum.beds.ac.uk/10.3390/d15020266