Responses of the Pheromone-Binding Protein of the Silk Moth Bombyx mori on a Graphene Biosensor Match Binding Constants in Solution

 , ,
, ,
Abstract
:1. Introduction
- Being the natural carriers for odorant and pheromones, with dissociation constants in the micromolar range, they represent the best candidates to trap and detect environmental odors.
- OBPs can be expressed in high yields in bacterial and eucaryotic systems and can be easily purified using standard protocols [11].
- Currently, the affinity of a ligand for an OBP is evaluated using a displacement assay, where a fluorescent probe is adopted as reporter [6]. This method is fast and simple, allowing for a wide screening of ligands. However, it suffers for being an indirect approach, and sometimes different fluorescent reporters can yield different affinity values for the same ligand [17]. Therefore, a label-free approach, such as the use of an electronic biosensor, would be highly desirable.
2. Materials and Methods
2.1. Protein Expression and Purification
2.2. Fluorescence Binding Assay
2.3. Fabrication of BmorPBP1 Based Biosensor Devices
2.4. Electrical Measurements
2.5. Analysis of Odorants Binding to BmorPBP1, Immobilized on rGO-FET
3. Results
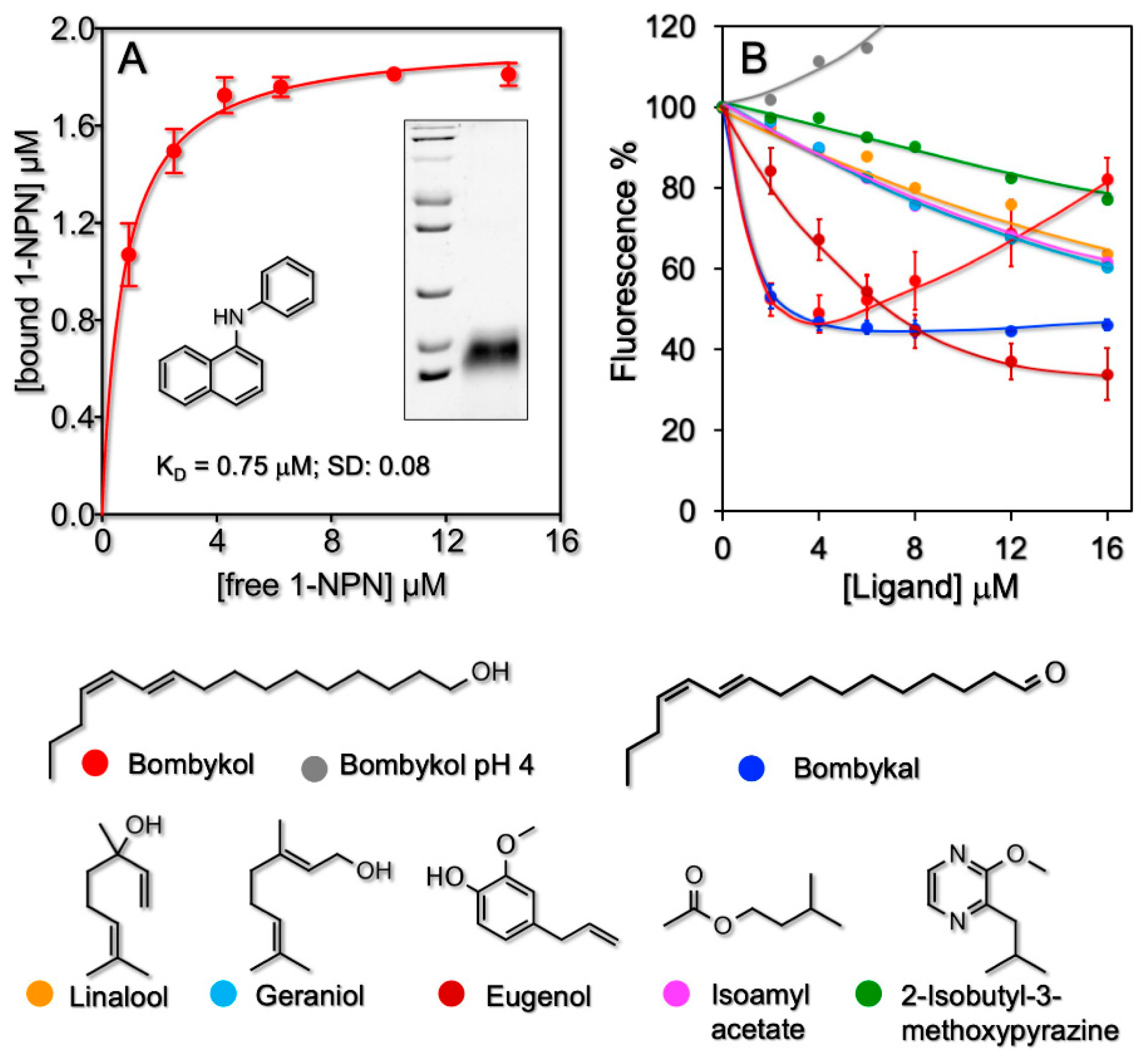
3.1. Affinities of BmorPBP1 to Odorants and Pheromones in Solution
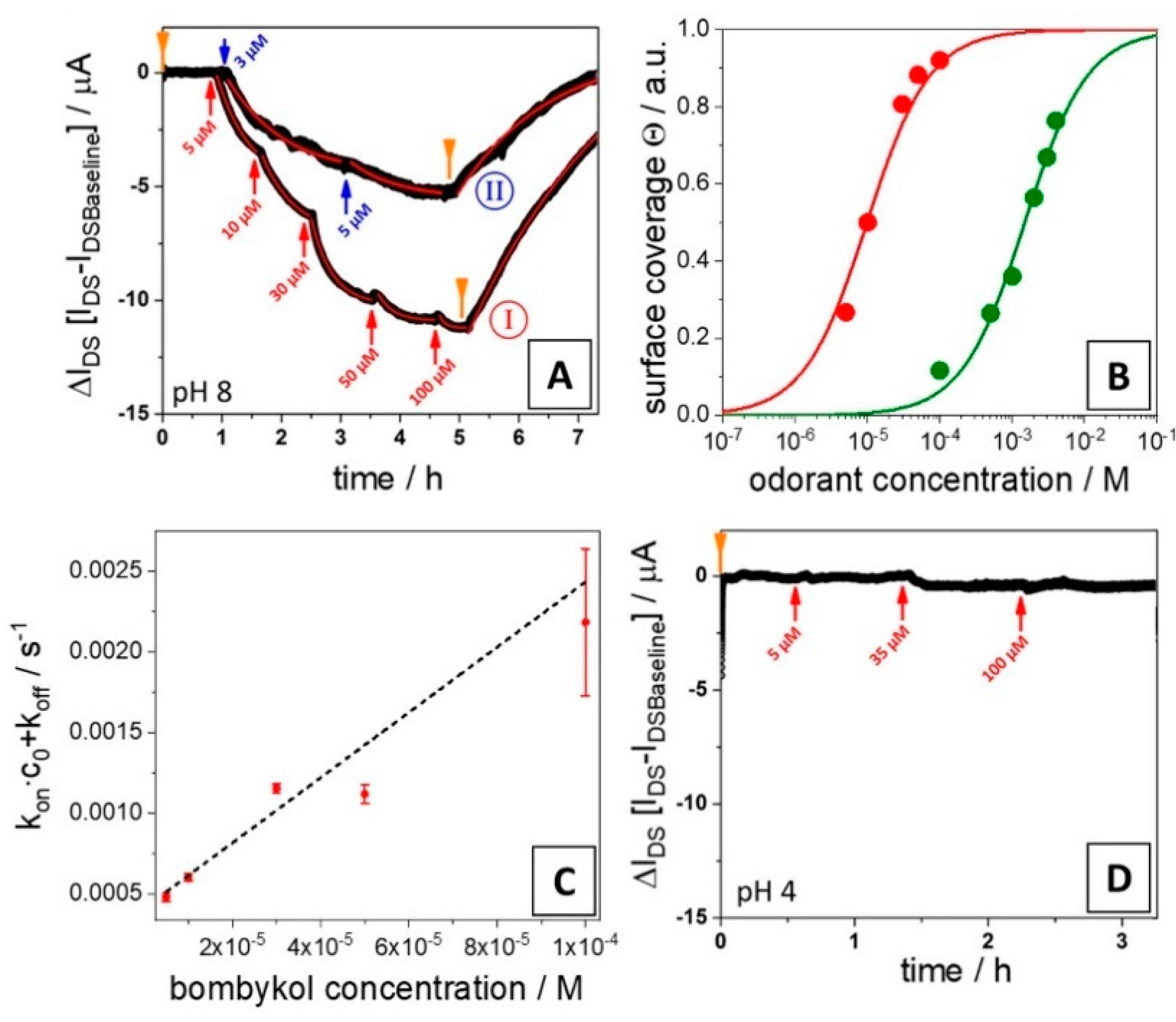
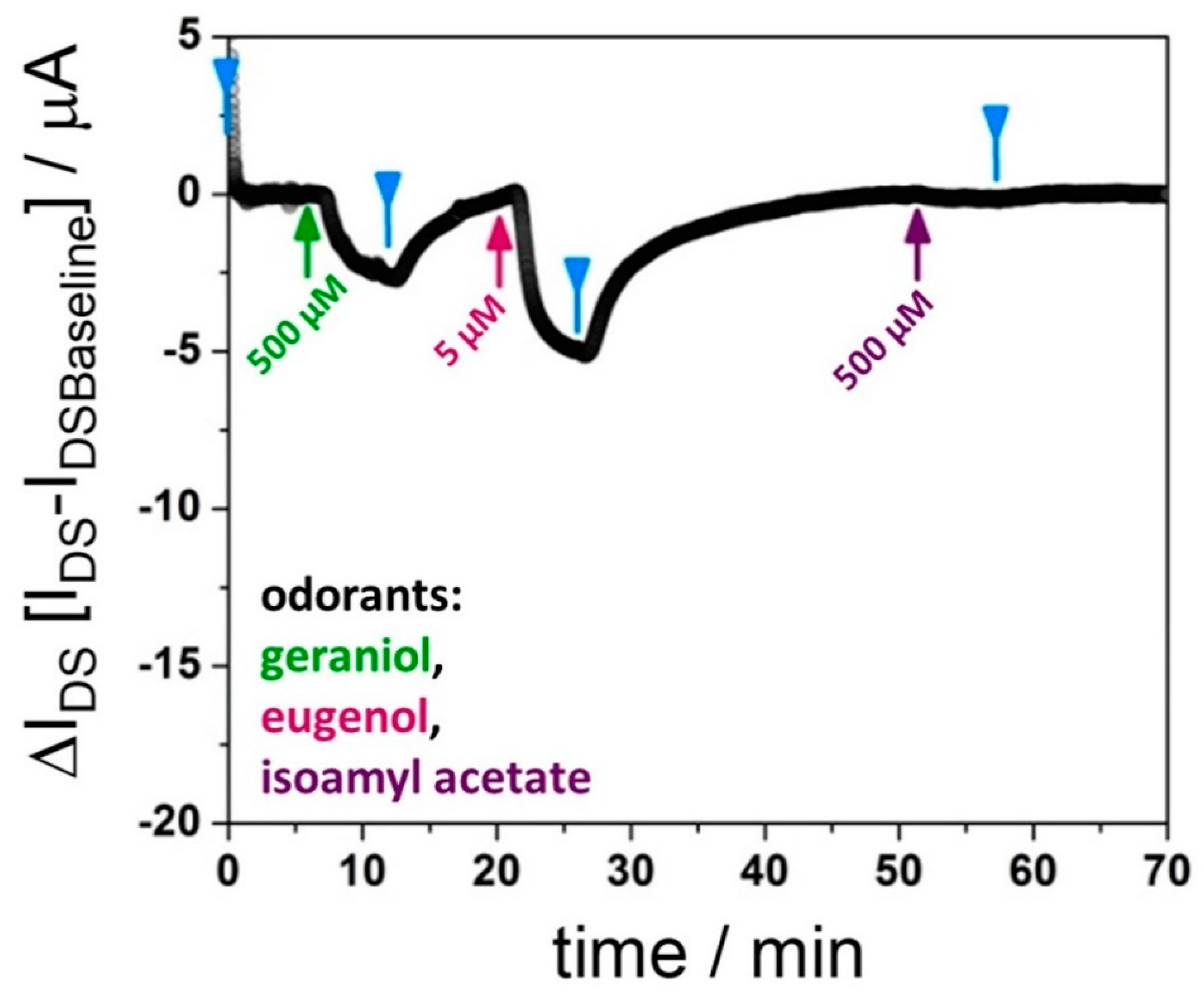
3.2. Affinities of BmorPBP1 to Odorants and Pheromones Measured by an Electronic Sensor
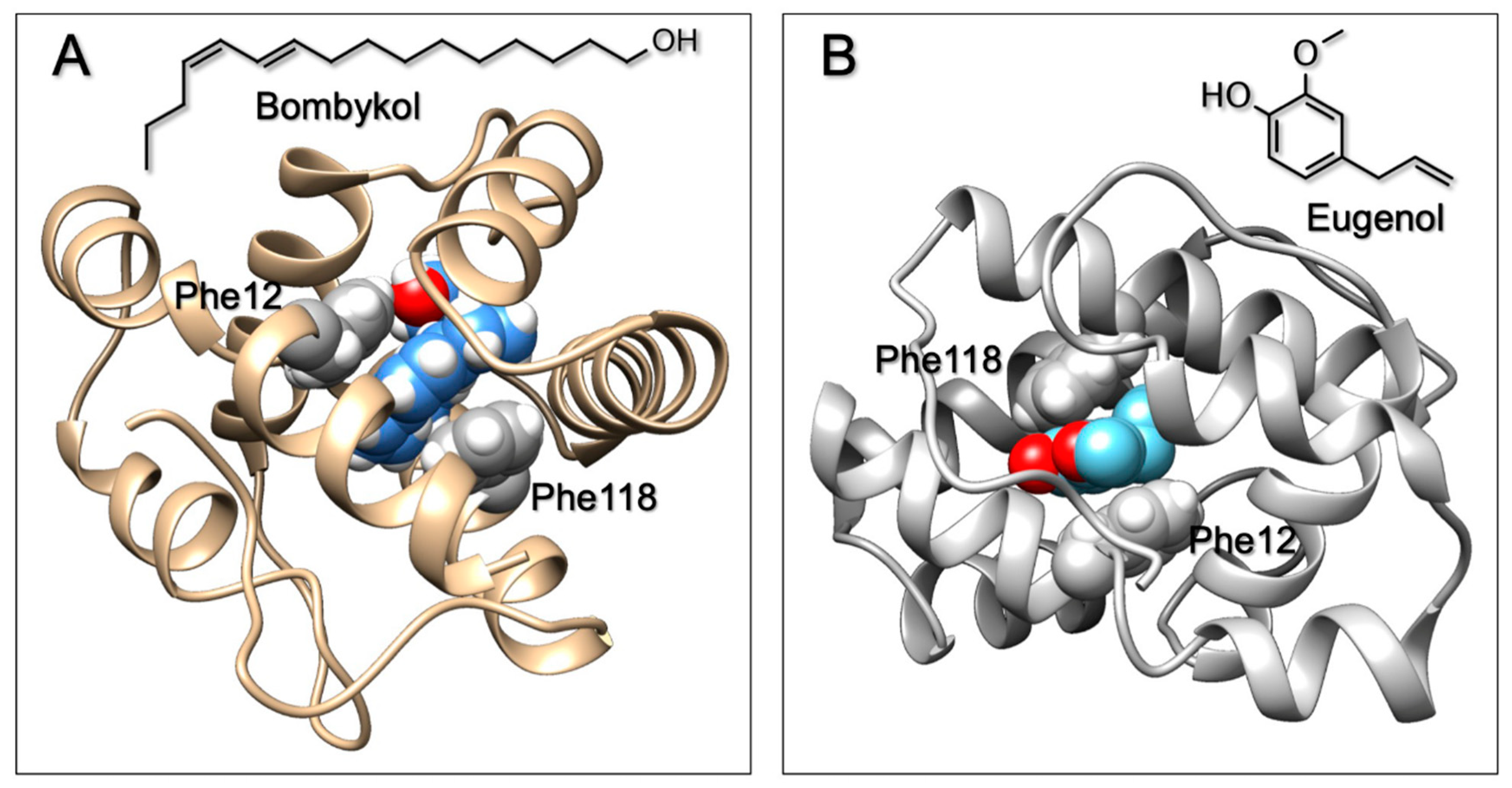
4. Discussion
Author Contributions
Funding

Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vogt, R.G.; Riddiford, L.M. Pheromone Binding and Inactivation by Moth Antennae. Nature 1981, 293, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond Chemoreception: Diverse Tasks of Soluble Olfactory Proteins in Insects: Soluble Olfactory Proteins in Insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelosi, P.; Zhou, J.-J.; Ban, L.P.; Calvello, M. Soluble Proteins in Insect Chemical Communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Baldaccini, N.E.; Pisanelli, A.M. Identification of a Specific Olfactory Receptor for 2-Isobutyl-3-Methoxypyrazine. Biochem. J. 1982, 201, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Pelosi, P. Odorant-Binding Proteins. Crit. Rev. Biochem. Mol. Biol. 1994, 29, 199–228. [Google Scholar] [CrossRef]
- Pelosi, P.; Zhu, J.; Knoll, W. From Radioactive Ligands to Biosensors: Binding Methods with Olfactory Proteins. Appl. Microbiol. Biotechnol. 2018, 102, 8213–8227. [Google Scholar] [CrossRef]
- Pelosi, P.; Zhu, J.; Knoll, W. Odorant-Binding Proteins as Sensing Elements for Odour Monitoring. Sensors 2018, 18, 3248. [Google Scholar] [CrossRef] [Green Version]
- Pelosi, P.; Zhu, J.; Knoll, W. From Gas Sensors to Biomimetic Artificial Noses. Chemosensors 2018, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Paolini, S.; Tanfani, F.; Fini, C.; Bertoli, E.; Pelosi, P. Porcine Odorant-Binding Protein: Structural Stability and Ligand A¤nities Measured by Fourier-Transform Infrared Spectroscopy and Fluorescence Spectroscopy. Biochim. Biophys. Acta 1999, 1431, 179–188. [Google Scholar] [CrossRef]
- Schwaighofer, A.; Kotlowski, C.; Araman, C.; Chu, N.; Mastrogiacomo, R.; Becker, C.; Pelosi, P.; Knoll, W.; Larisika, M.; Nowak, C. Honey Bee Odorant-Binding Protein 14: Effects on Thermal Stability upon Odorant Binding Revealed by FT-IR Spectroscopy and CD Measurements. Eur. Biophys. J. 2014, 43, 105–112. [Google Scholar] [CrossRef]
- Brulé, M.; Glaz, M.; Belloir, C.; Poirier, N.; Moitrier, L.; Neiers, F.; Briand, L. Chapter Six—Bacterial expression and purification of vertebrate odorant-binding proteins. In Methods in Enzymology; Odorant Binding and Chemosensory Proteins; Pelosi, P., Knoll, W., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 642, pp. 125–150. [Google Scholar]
- Tegoni, M.; Pelosi, P.; Vincent, F.; Spinelli, S.; Campanacci, V.; Grolli, S.; Ramoni, R.; Cambillau, C. Mammalian Odorant Binding Proteins. Biochim. Biophys. Acta Struct. Mol. Enzymol. 2000, 1482, 229–240. [Google Scholar] [CrossRef]
- Tegoni, M.; Campanacci, V.; Cambillau, C. Structural Aspects of Sexual Attraction and Chemical Communication in Insects. Trends Biochem. Sci. 2004, 29, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Arena, S.; Spinelli, S.; Liu, D.; Zhang, G.; Wei, R.; Cambillau, C.; Scaloni, A.; Wang, G.; Pelosi, P. Reverse Chemical Ecology: Olfactory Proteins from the Giant Panda and Their Interactions with Putative Pheromones and Bamboo Volatiles. Proc. Natl. Acad. Sci. USA 2017, 114, E9802–E9810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Ban, L.; Song, L.-M.; Liu, Y.; Pelosi, P.; Wang, G. General Odorant-Binding Proteins and Sex Pheromone Guide Larvae of Plutella Xylostella to Better Food. Insect Biochem. Mol. Biol. 2016, 72, 10–19. [Google Scholar] [CrossRef]
- Zaremska, V.; Tan, J.; Lim, S.; Knoll, W.; Pelosi, P. Isoleucine Residues Determine Chiral Discrimination of Odorant-Binding Protein. Chem. A Eur. J. 2020, 26, 8720–8724. [Google Scholar] [CrossRef]
- Tan, J.; Zaremska, V.; Lim, S.; Knoll, W.; Pelosi, P. Probe-Dependence of Competitive Fluorescent Ligand Binding Assays to Odorant-Binding Proteins. Anal. Bioanal. Chem. 2019, 412, 547–554. [Google Scholar] [CrossRef]
- Xu, Y.; Bai, P.; Zhou, X.; Akimov, Y.; Png, C.E.; Ang, L.-K.; Knoll, W.; Wu, L. Optical Refractive Index Sensors with Plasmonic and Photonic Structures: Promising and Inconvenient Truth. Adv. Opt. Mater. 2019, 7, 1801433. [Google Scholar] [CrossRef]
- Wang, Y.; Dostalek, J.; Knoll, W. Magnetic Nanoparticle-Enhanced Biosensor Based on Grating-Coupled Surface Plasmon Resonance. Anal. Chem. 2011, 83, 6202–6207. [Google Scholar] [CrossRef]
- Kotlarek, D.; Curti, F.; Vorobii, M.; Corradini, R.; Careri, M.; Knoll, W.; Rodriguez-Emmenegger, C.; Dostálek, J. Surface Plasmon Resonance-Based Aptasensor for Direct Monitoring of Thrombin in a Minimally Processed Human Blood. Sens. Actuators B Chem. 2020, 320, 128380. [Google Scholar] [CrossRef]
- Piscevic, D.; Lawall, R.; Veith, M.; Liley, M.; Okahata, Y.; Knoll, W. Oligonucleotide Hybridization Observed by Surface Plasmon Optical Techniques. Appl. Surf. Sci. 1995, 90, 425–436. [Google Scholar] [CrossRef]
- Benilova, I.; Chegel, V.I.; Ushenin, Y.V.; Vidic, J.; Soldatkin, A.P.; Martelet, C.; Pajot, E.; Jaffrezic-Renault, N. Stimulation of Human Olfactory Receptor 17-40 with Odorants Probed by Surface Plasmon Resonance. Eur. Biophys. J. 2008, 37, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Sanmartí-Espinal, M.; Iavicoli, P.; Calò, A.; Taulés, M.; Galve, R.; Marco, M.P.; Samitier, J. Quantification of Interacting Cognate Odorants with Olfactory Receptors in Nanovesicles. Sci. Rep. 2017, 7, 17483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulla, M.Y.; Tuccori, E.; Magliulo, M.; Lattanzi, G.; Palazzo, G.; Persaud, K.; Torsi, L. Capacitance-Modulated Transistor Detects Odorant Binding Protein Chiral Interactions. Nat. Commun. 2015, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Li, H.; Zhuang, S.; Zhang, D.; Zhang, Q.; Zhou, J.; Dong, S.; Liu, Q.; Wang, P. Olfactory Biosensor Using Odorant-Binding Proteins from Honeybee: Ligands of Floral Odors and Pheromones Detection by Electrochemical Impedance. Sens. Actuators B Chem. 2014, 193, 420–427. [Google Scholar] [CrossRef]
- Lu, Y.; Yao, Y.; Zhang, Q.; Zhang, D.; Zhuang, S.; Li, H.; Liu, Q. Olfactory Biosensor for Insect Semiochemicals Analysis by Impedance Sensing of Odorant-Binding Proteins on Interdigitated Electrodes. Biosens. Bioelectron. 2015, 67, 662–669. [Google Scholar] [CrossRef]
- Larisika, M.; Kotlowski, C.; Steininger, C.; Mastrogiacomo, R.; Pelosi, P.; Schütz, S.; Peteu, S.F.; Kleber, C.; Reiner-Rozman, C.; Nowak, C.; et al. Electronic Olfactory Sensor Based on A. Mellifera Odorant-Binding Protein 14 on a Reduced Graphene Oxide Field-Effect Transistor. Angew. Chem. Int. Ed. 2015, 54, 13245–13248. [Google Scholar] [CrossRef]
- Kotlowski, C.; Larisika, M.; Guerin, P.M.; Kleber, C.; Kröber, T.; Mastrogiacomo, R.; Nowak, C.; Pelosi, P.; Schütz, S.; Schwaighofer, A.; et al. Fine Discrimination of Volatile Compounds by Graphene-Immobilized Odorant-Binding Proteins. Sens. Actuators B Chem. 2018, 256, 564–572. [Google Scholar] [CrossRef]
- Bai, Y.; Xu, T.; Zhang, X. Graphene-Based Biosensors for Detection of Biomarkers. Micromachines 2020, 11, 60. [Google Scholar] [CrossRef] [Green Version]
- Sandler, B.H.; Nikonova, L.; Leal, W.S.; Clardy, J. Sexual Attraction in the Silkworm Moth: Structure of the Pheromone-Binding-Protein–Bombykol Complex. Chem. Biol. 2000, 7, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.-J.; Robertson, G.; He, X.; Dufour, S.; Hooper, A.M.; Pickett, J.A.; Keep, N.H.; Field, L.M. Characterisation of Bombyx Mori Odorant-Binding Proteins Reveals That a General Odorant-Binding Protein Discriminates Between Sex Pheromone Components. J. Mol. Biol. 2009, 389, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Damberger, F.; Nikonova, L.; Horst, R.; Peng, G.; Leal, W.S.; Wüthrich, K. NMR Characterization of a PH-Dependent Equilibrium between Two Folded Solution Conformations of the Pheromone-Binding Protein from Bombyx Mori. Protein Sci. 2000, 9, 1038–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larisika, M.; Huang, J.; Tok, A.; Knoll, W.; Nowak, C. An Improved Synthesis Route to Graphene for Molecular Sensor Applications. Mater. Chem. Phys. 2012, 136, 304–308. [Google Scholar] [CrossRef]
- Huang, J.; Larisika, M.; Fam, W.H.D.; He, Q.; Nimmo, M.A.; Nowak, C.; Tok, I.Y.A. The Extended Growth of Graphene Oxide Flakes Using Ethanol CVD. Nanoscale 2013, 5, 2945–2951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamczyk, M.; Moore, J.A.; Yu, Z. Application of Surface Plasmon Resonance toward Studies of Low-Molecular-Weight Antigen–Antibody Binding Interactions. Methods 2000, 20, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, R.; Fält, A. Experimental Design for Kinetic Analysis of Protein-Protein Interactions with Surface Plasmon Resonance Biosensors. J. Immunol. Methods 1997, 200, 121–133. [Google Scholar] [CrossRef]
- Khan, H.U.; Jang, J.; Kim, J.-J.; Knoll, W. In Situ Antibody Detection and Charge Discrimination Using Aqueous Stable Pentacene Transistor Biosensors. J. Am. Chem. Soc. 2011, 133, 2170–2176. [Google Scholar] [CrossRef]
- Mirsky, V.M. Quantitative Characterization of Affinity Properties of Immobilized Receptors. In Artificial Receptors for Chemical Sensors; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 1–15. ISBN 978-3-527-63248-0. [Google Scholar]
- Reiner-Rozman, C.; Larisika, M.; Nowak, C.; Knoll, W. Graphene-Based Liquid-Gated Field Effect Transistor for Biosensing: Theory and Experiments. Biosens. Bioelectron. 2015, 70, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Grosdidier, A.; Zoete, V.; Michielin, O. SwissDock, a Protein-Small Molecule Docking Web Service Based on EADock DSS. Nucleic Acids Res. 2011, 39, W270–W277. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sensor Data | Solution Data | ||
|---|---|---|---|
| Ligand | Kd/μM | Kd/μM | |
| Titration | Kinetics | ||
| Bombykol (pH 8.0) | 9.8 | 8.2 | 0.94 |
| Bombykol (pH 4.0) | - | >1000 | >100 |
| Bombykal | - | 7.4 | 1.1 |
| Eugenol | 7.6 | 8.1 | 2.8 |
| 2-Isobutyl-3-methoxypyrazine | 2800 | 1300 | >100 |
| Geraniol | - | 470 | ~20 |
| Linalool | - | >1000 | ~40 |
| Isoamyl acetate | - | >1000 | ~20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonazza, C.; Zhu, J.; Hasler, R.; Mastrogiacomo, R.; Pelosi, P.; Knoll, W. Responses of the Pheromone-Binding Protein of the Silk Moth Bombyx mori on a Graphene Biosensor Match Binding Constants in Solution. Sensors 2021, 21, 499. https://0-doi-org.brum.beds.ac.uk/10.3390/s21020499
Bonazza C, Zhu J, Hasler R, Mastrogiacomo R, Pelosi P, Knoll W. Responses of the Pheromone-Binding Protein of the Silk Moth Bombyx mori on a Graphene Biosensor Match Binding Constants in Solution. Sensors. 2021; 21(2):499. https://0-doi-org.brum.beds.ac.uk/10.3390/s21020499
Chicago/Turabian StyleBonazza, Caroline, Jiao Zhu, Roger Hasler, Rosa Mastrogiacomo, Paolo Pelosi, and Wolfgang Knoll. 2021. "Responses of the Pheromone-Binding Protein of the Silk Moth Bombyx mori on a Graphene Biosensor Match Binding Constants in Solution" Sensors 21, no. 2: 499. https://0-doi-org.brum.beds.ac.uk/10.3390/s21020499