
Brain Perivascular Macrophages Do Not Mediate Interleukin-1-Induced Sickness Behavior in Rats


1
UMR CNRS 5287 Aquitaine Institute for Integrative and Cognitive Neuroscience, University of Bordeaux, 33076 Bordeaux, France
2
Department of Symptom Research, MD Anderson Cancer Center, The University of Texas, Houston, TX 770030, USA
*
Author to whom correspondence should be addressed.
Pharmaceuticals 2021, 14(10), 1030; https://0-doi-org.brum.beds.ac.uk/10.3390/ph14101030
Submission received: 2 September 2021
/
Revised: 1 October 2021
/
Accepted: 7 October 2021
/
Published: 11 October 2021
(This article belongs to the Special Issue Cerebral Production and Action of Pro-inflammatory Cytokines)
{kind=link}
{kind=link}
Abstract
:Sickness behavior, characterized by on overall reduction in behavioral activity, is commonly observed after bacterial infection. Sickness behavior can also be induced by the peripheral administration of Gram-negative bacterial lipopolysaccharide (LPS) or interleukin-1beta (IL-1β), a pro-inflammatory cytokine released by LPS-activated macrophages. In addition to the microglia, the brain contains perivascular macrophages, which express the IL-1 type 1 receptor (IL-1R1). In the present study, we assessed the role of brain perivascular macrophages in mediating IL-1β-induced sickness behavior in rats. To do so, we used intracerebroventricular (icv) administration of an IL-1β-saporin conjugate, known to eliminate IL-R1-expressing brain cells, prior to systemic or central IL-1β injection. Icv IL-1β-saporin administration resulted in a reduction in brain perivascular macrophages, without altering subsequent icv or ip IL-1β-induced reductions in food intake, locomotor activity, and social interactions. In conclusion, the present work shows that icv IL-1β-saporin administration is an efficient way to target brain perivascular macrophages, and to determine whether these cells are involved in IL-1β-induced sickness behavior.
1. Introduction
Sickness behavior, consisting of reductions in food intake (hypophagia), locomotor activity, and social interactions, is commonly associated with bacterial infection [1,2,3]. Pro-inflammatory cytokines, such as interleukin-1 (IL-1), produced by activated immune cells in response to bacterial lipopolysaccharides (LPS) [4,5,6], also reduce food intake, activity, and social interactions, and mediate sickness behavior [7,8,9,10,11,12]. Over the past decades, prostaglandin synthesis at the blood–brain barrier (BBB), activation of the vagus nerve, and the central transport or synthesis of IL-1 have been proposed to take part in the immune-to-brain signaling underlying sickness-associated changes in behavior [13]. In accordance with the first hypothesis, the endothelial-targeted knockdown of the IL-1 type 1 receptor (IL-1R1) in mice has been found to abrogate sickness behavior after intravenous (iv), but not after intraperitoneal (ip), administration of IL-1β [14]. However, we recently showed that IL-1R-expressing neurons in the arcuate hypothalamus of rats play a role in hypophagia after ip IL-1β administration [15], thus corroborating the hypothesis that IL-1β action in the brain is important for sickness behavior.
However, given that the arcuate nucleus seems much more accessible to high-molecular-weight components circulating in the blood, rather than to those present in the cerebrospinal fluid [16,17], IL-1-receptors in this structure may be a preferential target for the action of systemic IL-1β, instead of IL-1β produced in the brain. In addition to neurons, rodent brain endothelial cells and macrophages, and, to a lesser extent, astrocytes, express IL-1R1 [14,18,19]. Interestingly, peripherally administered LPS induces IL-1β immunoreactivity in the macrophages of brain circumventricular organs, such as the organum vasculosum lamina terminalis (OVLT) and choroid plexus [20,21], as well as bioactive IL-1β in the rat ventricular cerebrospinal fluid [22]. Moreover, intracerebroventricular (icv) administration of IL-1β rapidly spreads through the perivascular spaces and induces nuclear translocation of the transcription factor NF-κb in perivascular cells [23,24], in addition to sickness behavior [25,26,27]. Furthermore, circulating IL-1β has been proposed to act on rat brain perivascular macrophages to induce the prostaglandin-synthesizing enzyme cyclooxygenase-2 [28]. Thus, these findings raise the question as to whether central and/or peripheral IL-1β acts on brain perivascular macrophages to induce sickness behavior.
The cell types mediating the behavioral effects of IL-1 have been mostly studied using different promotor-specific Cre-Lox recombinant mice. During mouse development, the angiopoietin-1 receptor promotor Tie-2 is highly expressed in brain endothelial cells [29], but also in myeloid precursors that give rise to brain F4/80-positive cells [30], while expression of the fractalkine receptor promoter Cx3cr1 occurs in the microglia and, to a lesser extent, in brain macrophages, which both express Iba-1 [31,32]. In contrast, the thyroid hormone transporter and solute carrier organic anion transporter family member 1C1 (Slco1c1) promoter is expressed in rodent brain parenchymal endothelial cells, tanycytes, choroid plexus cuboid cells, and some cortical and hippocampal neurons [33,34,35]. Thus, it has been shown that Tie-2-dependent, but not Cx3cr1- or Slco1c1-dependent, genetic deficiency for IL-R1, or the IL-R1-associated signaling protein Myd88, abrogates sickness behavior induced by icv, but not ip, IL-1β [14,19,24]. Given the still-debated expression of Cx3cr1 in brain macrophages [31,32,36,37], these findings can be interpreted to suggest that IL-1R1 signaling in the fenestrated capillaries of brain circumventricular organs and choroid plexus [24], or in brain macrophages, plays a role in IL-1β-induced sickness behavior.
The present experiments were designed to test the hypothesis that IL-1R1-expressing brain perivascular macrophages mediate IL-1β-induced sickness behavior. Brain perivascular macrophages are a distinct population of myeloid cells that differ from microglia in their developmental origin, location, and activity profile [32,37,38]. To specifically target IL-1R1-expressing brain perivascular macrophages, we used icv administration of an IL-1ß-saporin conjugate that we recently validated to lesion brain IL-1 receptor-expressing cells [15]. Saporin is a ribosomal-inactivating protein [39], which is non-toxic outside the cells, but can enter the cell if coupled to a ligand that is internalized once bound to its receptor [40,41]. Given that icv IL-1 preferentially spreads along the perivascular spaces [23], the administration of IL-1ß-saporin via this route is expected to selectively eliminate IL-R1-expressing brain perivascular macrophages and, therefore, allow the role of these cells in IL-1β-induced sickness behavior to be established.
2. Results
2.1. Ip IL-1 β-Alexa Conjugate Decreased Food Intake
A one-way ANOVA on food intake after ip injection showed that 60 µg/kg of mouse (m)IL-1β-Alexa-488 significantly reduced cumulated food intake 2, 3.5 and 5 h later, as compared to the administration of a vehicle (F(1,14) = 5.94, p < 0.05; F(1,12) = 8.22, p < 0.05; F(1,10) = 5.92, p < 0.05, respectively (data not shown)), thus indicating that the conjugation reaction did not affect IL-1β’s bioactivity.
2.2. Icv IL-1β-Saporin Conjugate Eliminated Brain Perivascular Macrophages, but Not Endothelial Cells
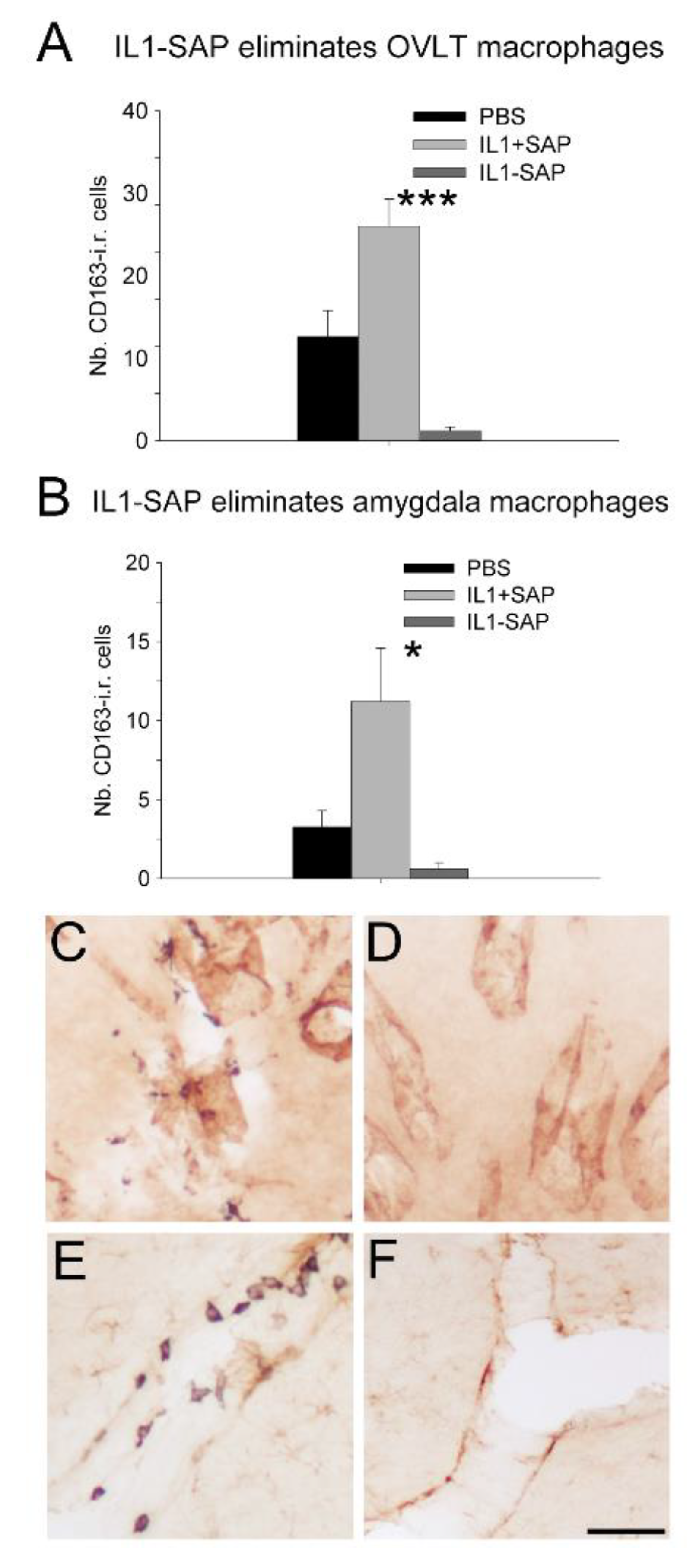
After semi-quantitative immunohistochemistry, one-way ANOVA on the number of CD163-positive cells in the OVLT, where many perivascular macrophages are concentrated, revealed a significant effect of icv pretreatment (F(2,26) = 19.0, p < 0.001). Post hoc tests showed that mIL-1β-saporin conjugate administration reduced the number of CD163-positive cells in the OVLT, as compared to PBS or mIL-1β + saporin (not conjugated) administration (p < 0.05 and p < 0.001, respectively; Figure 1A,C). Similar analyses on blood vessels within the amygdala indicated a significant effect of icv pretreatment on the number of CD163-positive cells (F(2,27) = 6.54, p < 0.01). Post hoc tests then revealed that the administration of mIL-1β + saporin induced an increase in CD163-positive cells in the amygdala, as compared to that of PBS (p < 0.05), but also indicated that the icv administration of the mIL-1β-saporin conjugate reduced the number of perivascular macrophages, compared to mIL-1β + saporin administration (p < 0.05. Figure 1B). Finally, and according to one-way ANOVA, icv pretreatment had a significant effect on the surface of CD163-positive cells, relative to the surface of vessels located between the cortex and the striatum (F(2,27) = 8.75, p < 0.001). Subsequent post hoc tests showed that the mIL-1β-saporin conjugate decreased the relative surface of CD163-positive cells, compared to PBS or mIL-1β + saporin (p < 0.05 and p < 0.001, respectively; data not shown). At those locations where icv administration of the mIL-1β-saporin conjugate reduced the number of perivascular macrophages, no major differences in GFAP and vWF immunoreactivity were observed between the icv pretreatments (Figure 1C,D).
2.3. Icv IL-1β-Saporin Conjugate Did Not Alter Behavioral Responses to Subsequent IL-1β Injection
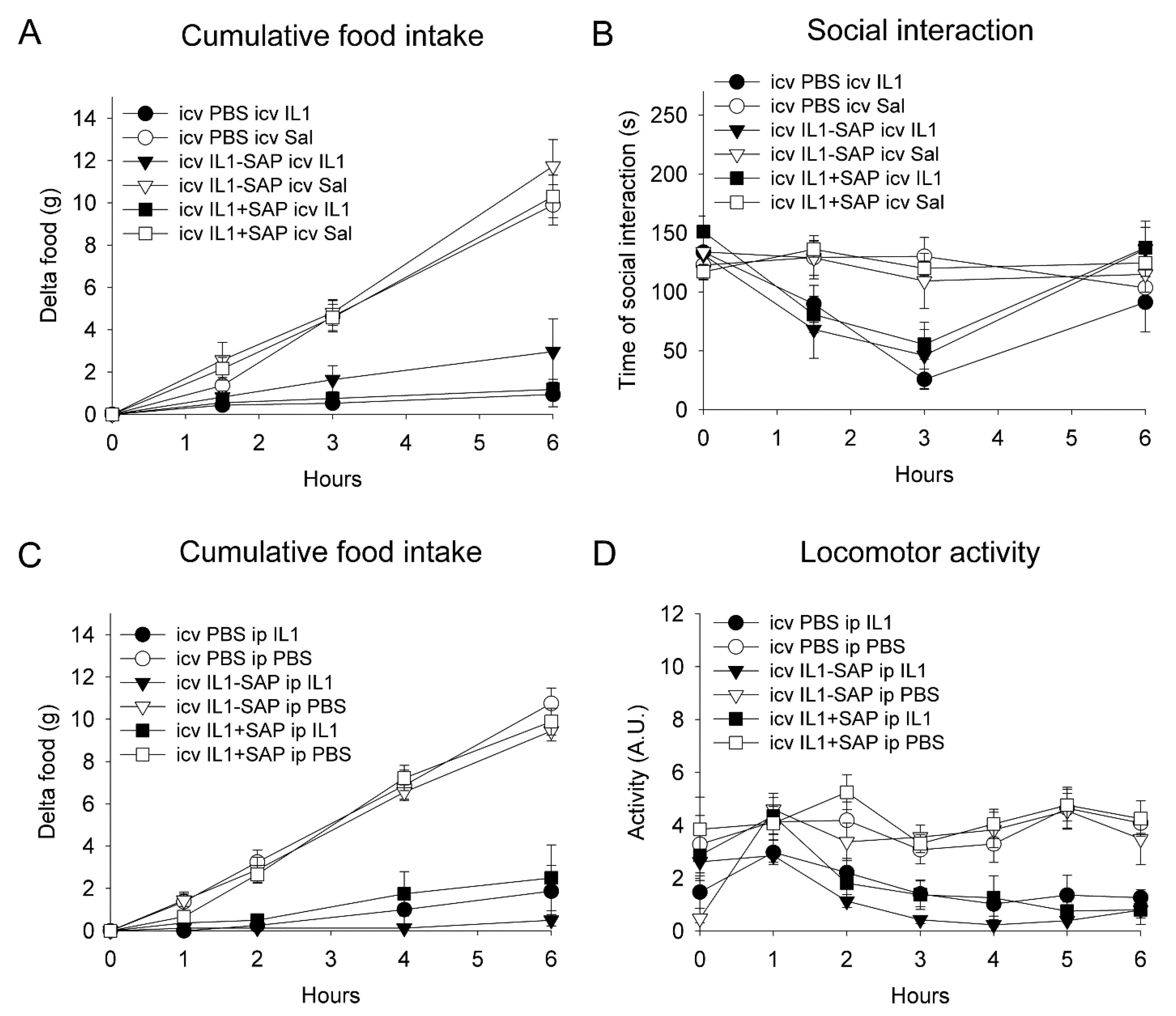
A two-way ANOVA on cumulative food intake, over six hours after icv rat recombinant (rr)IL-1β or saline administration, five days after pre-treatment, indicated a significant reduction in icv rrIL-1β-injected rats as compared to vehicle-treated animals (F(1,29) = 112, p < 0.001), with no significant effect of the pre-treatment or interaction between the pre-treatment and treatment (Figure 2A). Similarly, a three-way repeated measures ANOVA on social exploration, 1.5, 3 and 6 h after icv rrIL-1β or saline administration (treatment), showed that rrIL-1β induced a significant decrease (F(1,29) = 16.5, p < 0.001), without any significant effect of the pre-treatment (Figure 2B).
A two-way ANOVA on cumulative food intake, over six hours after ip rrIL-1β or saline administration, five days after pre-treatment, indicated a significant reduction in icv rrIL-1β-injected rats as compared to vehicle-treated animals (F(1,41) = 31.1, p < 0.001), with no significant effect of the pre-treatment or interaction between the pre-treatment and treatment (Figure 2C). Similarly, a three-way repeated measures ANOVA on locomotor activity, during the first six hours after ip rrIL-1β or saline administration (treatment), showed that rrIL-1β induced a significant overall decrease (F(1,34) = 77.4, p < 0.001), without a significant effect of the pre-treatment or interaction between the treatment and pre-treatment (Figure 2D).
3. Discussion
The aim of the present experiments was to determine the role of IL-1R1-expressing brain perivascular macrophages in mediating IL-1β-induced sickness behavior, using an IL-1R1-saponin conjugate to specifically eliminate cells containing IL-1R1 [15]. The administration of this conjugate reduced the number of CD163-positive brain perivascular macrophages, known to express IL-1R1 [18], without altering the immunoreactivity for endothelial cells or astrocytes, but did not affect sickness behavior in response to either icv or ip administration of IL-1β. These findings are important because they allow, for the first time, IL-1R1-expressing brain perivascular macrophages to be targeted in a more direct way, as compared to the use of promotor-specific Cre-Lox recombinant mice. In addition, they indicate that perivascular macrophages do not play an important role in the behavioral effects of exogenously administered IL-1β.
In experimental life sciences, it is of the utmost importance to intervene on a target causal candidate in different ways. Thus, it has been argued that repeating the same experiments is not enough to promote robust research, but that, in addition, different lines of evidence are required to mitigate the biases associated with findings obtained using a particular approach to address a research question [42]. When it comes to the question of which cell types mediate the behavioral effects of IL-1, the vast majority of recent studies have employed different promotor-specific Cre-Lox recombinant mice. One of the major limitations of such approaches is the specificity of the promotors used. Indeed, specific promotors may not yet be widely available for all cell types, and the specificity of promotors that have already been used may be less than initially presumed. In particular, there is an ongoing discussion about the expression of the Cx3cr1 promotor in brain cells. While some authors have presented Cx3cr1-Cre-Lox recombinant mice as lacking expression of a gene of interest only in the microglia, there is increasing evidence to suggest that this promotor is expressed by brain macrophages and, in some conditions, by neurons [31,32,36,37,38,43]. Similarly, the Ttie-2 promotor Tie-2 is highly expressed in brain endothelial cells [29], but has also been found in myeloid precursors that give rise to brain cells expressing F4/80 [30], which is one of the markers expressed by perivascular macrophages [32,37,38]. Thus, the currently widely employed promotor-specific Cre-Lox recombinant mice do not seem to be adequate to address the role of brain perivascular macrophages in the behavioral effects of IL-1β. Another important limitation of the studies published thus far, addressing the brain cell types mediating IL-1β’s effects on behavior, is that they have all been conducted in mice.
The role of brain perivascular macrophages in the effects of IL-1β on behavior was, therefore, studied in rats using non-genetically modified animals. One of the traditional ways of targeting these cells is the icv administration of liposomes containing clodronate. This treatment results in phagocytosis of these particles, by perivascular macrophages, and subsequent uptake of the apoptosis-inducing drug clodronate [44]. Although the initial article based on this technique reported no effect of such treatment on the microglia [44], recent work indicates that the icv administration of liposomes containing clodronate can also reduce the number of microglia in adult rodent brains [45,46]. Since microglia do not express IL-1R1 under basal conditions [18,19], icv administration of the IL-1ß-saporin conjugate that was previously shown to specifically lesion brain IL-1 receptor-expressing neurons in the hippocampus [15] appears to be an efficient way to eliminate brain perivascular macrophages without affecting the microglia. However, since icv-administered IL-1 preferentially spreads along the brain perivascular spaces [23], perivascular cell types, other than macrophages, known to express IL-R1, such as endothelial cells and astrocytes [18,19], may also be a target of icv IL-1ß-saporin. This is the reason why, in our previous and present work, the effect of IL-1ß-saporin administration on brain endothelial cells and astrocytes was studied. Importantly, we have shown that immunoreactivity for the endothelial cell marker von Willebrand factor, and that for the astrocyte marker glial fibrillary acidic protein, was not affected by intracerebral or icv administration of the IL-1ß-saporin conjugate [15]. The absence of an effect of IL-1ß-saporin on these cell types may be explained by postulating that IL-R1s are neither expressed on the abluminal side of brain endothelial cells nor on astrocytic end feet. Indeed, the only rat brain cell types that were affected by the IL-1ß-saporin conjugate were the hippocampal and arcuate hypothalamic neurons after local microinjection [15], and perivascular macrophages after icv administration (present work). Icv administration of IL-1ß-saporin is, therefore, an important experimental tool to study the contribution of IL-R1-expressing brain perivascular macrophages to mediating IL-1β-induced sickness behavior.
Because of their location in the perivascular spaces, and their close contact with the abluminal site of brain endothelial cells, perivascular macrophages have long been thought to connect the immune and nervous systems [47,48,49]. Beyond their roles as scavenger phagocytes in neuropathologies, these cells have been shown to play a role in hypertension-associated neurovascular and cognitive dysfunction, activation of the paraventricular nucleus of the hypothalamus, and corticosterone and noradrenaline release after systemic IL-1β administration or myocardial infarction in rodents [36]. However, their role in mediating IL-1β-induced sickness behavior has not been studied directly. Since this pro-inflammatory cytokine can be detected peripherally as well as in the brain in animal models of systemic inflammation [21,50], IL-1β was injected either ip or icv after icv administration of IL-1ß-saporin conjugate to study the role of brain macrophages in sickness behavior. However, even though icv IL-1β-saporin administration significantly reduced the number of brain perivascular macrophages, it did not alter subsequent icv or ip IL-1ß-induced reduced food intake, locomotor activity, and social interactions. These findings indicate that IL-1R1-expressing brain perivascular macrophages do not mediate sickness behavior after intracerebroventricular IL-1β injection. This lack of effect could theoretically be due to the fact that an in vivo behavioral assessment was performed at five days after administration, several days before the post-mortem histological examination. However, this is rather unlikely, as other populations of macrophages have been shown to be reduced within one week after an injection of saporin conjugates [51,52,53]. Moreover, increasing the interval between IL-1β-saporin conjugate administration and subsequent IL-1ß injection to thirteen days did not affect the sickness behavior [15]. Finally, IL-R1s in the rat arcuate hypothalamus mediate in part hypohagia after systemic IL-1ß injection, since the local administration of IL-1β-saporin into this brain structure mitigated, without preventing, ip IL-1ß-induced reduced food intake [15]. Therefore, the most likely explanation for our finding is that brain perivascular macrophages do not play a role in IL-1ß-induced sickness behavior.
4. Materials and Methods
4.1. Animals
Thirty-three and fifty male sexually naïve Wistar Han IGS: [Crl:WI(Han)] rats that weighed 175–225 g upon arrival (Charles River, L’Arbresle, France) were housed five per polypropylene cage (30 × 45 × 19 cm) containing corncob bedding (UAR, Epinay-sur-Orge, France) and left undisturbed for fourteen days in a room with controlled temperature (23 ± 1 °C) and light (12/12 h light–dark cycle) conditions and ad libitum access to food (Extralabo, Provins, France) and water. Twenty-one- to thirty-day-old juvenile animals of the same strain were used as social stimuli and housed in groups of ten in a similarly controlled room. Studies were carried out in accordance with the INRA quality reference system (www.international.inra.fr/content/download/947/11111/file/requirements.pdf accessed on 1 September 2014), approved by the Aquitaine Poitou-Charente regional ethical committee on animal experiments (AP 1 March 2004) and followed European recommendations on animal research (European Council Directive of 24 November 1986 (86/609/EEC) and European Parliament and Council Directive of 22 September 2010 (2010/63/UE)).
4.2. Biologically Active Molecules
Purified rat recombinant interleukin-1beta (rrIL-1ß; biological activity: 317 IU/mg, NIBSC, Potters Bar, UK) was dissolved in sterile phosphate-buffered saline (PBS), as previously described [54], and used for intracerebroventricular (icv) injections. Non-targeted saporin, a ribosome-inactivating protein from Saponaria officinalis (Advanced Targeting Systems, San Diego, CA, USA), dissolved in sterile PBS, was used for the conjugation of saporin to IL-1ß and as a control for treatment with this conjugate (see below). Since rat IL-1ß contains four cysteine residues, coupling to saporin would be likely to result in a heterogeneous molecule with variable toxicity. However, mouse, similar to human, IL-1ß only contains one cysteine residue and is bioactive in rats [55,56,57]. Furthermore, this cysteine residue does not affect biological activity [58], which allowed this residue to be changed to serine by site-directed mutagenesis to result in mouse [S71]IL-1ß. Next, a new cysteine was introduced by changing the C-terminal serine to cysteine resulting in [S71,C152]mIL-1ß. This molecule was used as starting material for conjugation to Alexa-488, a chemically modified fluorescein molecule, or to saporin, and as a supplementary control for the effects of the conjugate. The molecules were first derivatized by N-succinimidyl-3-(2-pyridyl) dithiopropionate (SPDP; Pierce, Perbio Science, Brebières France) at room temperature for 24 h and subsequently coupled to the cysteine residue of mouse [S71,C152]IL-1ß for another 24 h at room temperature. The molar ratio of Alexa-488 or saporin to mouse [S71,C152]IL-1ß was 1:1. The coupling reaction between [C71,C152]mIL-1ß and Alexa-488 resulted in a single peak of emission centered around 494 nm (not shown). The coupling reaction between [C71,C152]mIL-1ß and saporin yielded a conjugate molecule migrating at the expected size on an electrophoresis gel [15].
4.3. Sickness Behavior Testing after Intraperitoneal Injection of Interleukin-1β-Alexa-488
To assure that mouse [S71,C152]IL-1β conjugates were biologically active and induced sickness behavior, rats were given an ip injection of 60 µg/kg [S71,C152]mIL-1β-Alexa-488 or saline two hours before the start of the dark phase. Food intake was monitored by weighing food pellets in ruffs just before and 2, 3.5 and 5 h after injection on a high-precision balance. Even though rats show their food intake peak during the first hours of the dark phase if they have ad libitum access to food, each animal has its own pattern that is repeated everyday [59]. Cumulative food intake over 5 h was therefore chosen for data presentation and analysis. One week after the first injection, treatments were reversed according to a Latin square design.
4.4. Intracerebroventricular Injection of Interleukin-1β ± Saporin
Rats were first anesthetized through an ip injection (1 mL/kg) of ketamine (61 mg/kg) and xylazine (9 mg/kg) and secured in a stereotaxic apparatus. A midline incision was made and skin flaps were pulled aside.
In thirty-three animals, a stainless-steel 7 mm long guide cannula (23 Gauge) was placed above the lateral brain ventricle for subsequent icv administration of [S71,C152] mouse interleukin-1β ± saporin or vehicle and ultimately ip rat interleukin-1β or vehicle (see below). This guide cannula was lowered unilaterally 3.2 mm below the skull surface (2.2 mm below the dura mater) through a hole drilled in the skull 0.6 mm posterior to bregma and 1.5 mm lateral to the midline. Coordinates were chosen to place the tip of this guide cannula 1 mm above the roof of the lateral ventricle. Guide cannulas were permanently connected to the skull with dental cement and screws. The head wound was closed by suturing skin flaps and rats were allowed a two-week recovery period before icv injections and behavioral testing. For icv injections into the lateral ventricle, a 30-gauge 8 mm long injection cannula was lowered through the guide cannula and connected to a 10 µL Hamilton syringe with plastic tubing. Ten microliters of the following solutions were infused to deliver (1) 1.75 μg conjugated [S71,C152]mIL-1β-saporin dissolved in phosphate-buffered saline (PBS; [S71,C152]mIL-1ß-saporin; n = 12), (2) equivalent amounts of unconjugated saporin and [S71,C152]mIL-1ß dissolved in PBS ([S71,C152]mIL-1ß + saporin; n = 9), or (3) PBS (PBS; n = 12) to hand-held animals. The injection device was left in place for another thirty seconds to allow diffusion of chemicals or vehicle and then retracted. These rats were subsequently used to study the effects of these pretreatments on icv IL-1ß-induced sickness behavior (see below).
The doses of [S71,C152]mIL-1β-saporin conjugate used were based on the finding that 6–600 ng of mouse IL-1β administered icv reduces food intake in the rat [55], bearing in mind that only 40% (17 kDa/42 kDa), about 700 ng, of the weight of the IL-1β-saporin construct corresponds to IL-1β. A 10-fold excess of the IL-1β-saporin construct was infused icv relative to the amount of rrIL-1β administered five days later via the same guide cannula (see below).
In fifty animals, a glass micropipette with a tip diameter of 100–120 µm was placed 8.5 mm below the midline dural surface, 1.8 mm posterior to bregma, for third brain ventricle administration of [S71,C152] mouse interleukin-1β ± saporin or vehicle. This micropipette was connected to a 10 µL Hamilton syringe with plastic tubing with the aid of which five µL of solution was infused to administer the following: (1) 875 ng conjugated [S71,C152]mIL-1β-saporin dissolved in PBS ([S71,C152]mIL-1ß-saporin; n = 17), (2) equivalent amounts of unconjugated saporin (330 ng) and [S71,C152]mIL-1ß (544 ng) dissolved in PBS ([S71,C152]mIL-1ß + saporin; n = 17), or (3) PBS (PBS; n = 16). These rats were subsequently used to study the effects of these pretreatments on ip IL-1ß-induced sickness behavior (see below).
After the head wound was closed by suturing skin flaps, a telemetric transmitter (TA10TA-F40, Data Sciences, St. Paul, MN, USA) was inserted into the peritoneal cavity of each rat through a 2 cm incision in the linea alba. The abdominal wall and skin were then sutured. A ten day period was given to the animals to recover from surgery and to allow enough time for saporin to provoke lesions. Food intake and body weight were measured daily from the day before surgery until the end of the experiments.
4.5. Sickness Behavior Assessment after Intracerebroventricular Interleukin-1β Injection
Since conjugated saporin constructs can kill 66–75% of cells, including macrophages, within 5 days [53,60,61], a challenge with icv rr IL-1β or saline was given five days after icv pretreatment of IL-1ß ± saporin or PBS. Thus, during the early dark phase, rats were administered icv with either 70 ng of rrIL-1ß dissolved in 2 µL of saline or vehicle at a rate of 0.5 µL every thirty seconds. Social interaction with a juvenile rat of the same strain and food intake were measured (see below) 1.5, 3 and 6 h later as measures of sickness behavior. Social behavioral testing took place in the home cage of the test animal as previously described [15,50]. A trained observer, blind to the treatment given, scored the duration of social exploration of the test rat directed towards the juvenile rat from a video screen fed by a camera placed over the home cage using pre-set keys on the keyboard of a microcomputer. Social exploration consisted mainly of ano-genital sniffing, but also included nosing, sniffing of body parts, and grooming behaviors. Food intake and body weight changes were obtained by measuring weight of food pellets in ruffs and rats just before and 1.5, 3 and 6 h after injection on a high-precision balance. Cumulative food intake and changes in body weight over 6 h were chosen for data presentation and analysis.
4.6. Sickness Behavior Assessment after Intraperitoneal Interleukin-1β Injection
Ten days after third brain ventricle administration, half of the animals of each experimental group received an intraperitoneal injection (1 mL/kg) of sterile PBS and the other half an intraperitoneal injection of IL-1β (30 µg/kg) one hour before the lights went off. Horizontal locomotor activity and food intake were used as measures of sickness behavior. Food intake was measured at the time of ip injection and 1, 2, 4, 6 h after injection. Locomotor activity was monitored every 10 min for 10 s with the implanted telemetry devices based on the changes in power of the transmitted signal due to the transmitter position changes and fed into a computer-based data acquisition system (Dataquest IV, Data Sciences, St. Paul, MN, USA).
4.7. Brain Tissue Preparation
Twenty-four hours after the last injections, animals were anesthetized with pentobarbital (150 mg/kg) and perfused transcardially with 0.9% NaCl followed by 4% paraformaldehyde (pH 7.4). Fixed brains were removed from the skull and placed for another 4 h in the same fixative before being transferred to 30% sucrose and stored at 4 °C for 48 h. Brains were subsequently frozen and stored at −80 °C. Forty-micrometer brain sections were cut on a cryostat and stored in a cryoprotective solution (20% glycerol, 30% ethylene glycol in PBS, 0.1 M, pH 7.4) at −20 °C.
4.8. Immunohistochemistry
To determine to what extent icv administration of IL-1β-saporin conjugate affected brain macrophages, astrocytes and endothelial cells, immunohistochemistry was employed to detect the macrophage-specific marker CD163, the astrocyte-specific marker glial fibrillary acidic protein (GFAP) and the endothelial-specific marker von Willebrand factor (vWF), respectively. Immunohistochemistry was performed as previously described [15,50]. Briefly, free-floating sections were washed four times in 0.1 M PBS (pH 7.4). Non-specific binding sites were blocked by a 45-minute incubation in PBS containing 0.3% Triton X-100 and 1.0% bovine serum albumin (BSA). The first antibody was subsequently diluted in the same buffer and added to sections overnight at room temperature. Commercially available antisera raised against CD163 (Serotec, mouse anti-rat CD163, MCA342R), GFAP (rabbit anti-GFAP, Sigma, 63893, Sigma-Aldrich, St-Fallavier, France), and vWF (rabbit anti-vWF, ab6994, Abcam, Paris, France), all diluted 1:1000 were used. These antisera have been well characterized according to the Journal of Comparative Neurology antibody database available at antibodyregistry.org (IDs for CD163, GFAP, and vWF antibodies were AB_321966, AB_477010, and AB_305689, respectively). After four rinses in PBS, sections were treated for 30 min with 0.3% (v/v) hydrogen peroxide to block endogenous peroxidases followed by additional rinses in PBS. Sections were then incubated for 2 h with biotinylated antisera raised against mouse or rabbit IgGs (Vector Laboratories, Burlingame, CA, USA) diluted 1:1000 in PBS containing 0.3% Triton X-100 and 1% BSA. After four washes in PBS, sections were incubated for 2 h with a complex of avidin and biotinylated peroxidase (Vector Laboratories, Burlingame, CA, USA) diluted 1:500 in PBS. Finally, sections were transferred to a sodium acetate buffer and stained using diaminobenzidine as a chromogen in the presence of Ni ions, thus yielding a dark grey to black precipitate. In the case of double-labeling immunohistochemistry, CD163 immunoreactivity was revealed following this staining procedure, after which sections were incubated with GFAP or vWF antibodies. The binding of these antibodies was then revealed following the same protocol, except that staining occurred in the absence of Ni ions and thus resulted in brown labeling.
4.9. Microscopy
Stained sections were examined with a light microscope (Nikon Eclipse, Nikon France or Leica DM5500B, Leica Microsystems, Nanterre, France) and images were captured using a high-resolution CCD video camera and fed into a computer. To quantify CD163-immunoreactive cells, image processing was performed using Image J (http://imagej.nih.gov/ij/ accessed on 1 September 2021) on grey images by defining brightness and surface area above which labeling was to be taken into account. Once established, these parameters remained unchanged. CD163-positive cells were thus quantified along vessels of the organum vasculosum laminae terminalis (OVLT), the amygdala and between cortex and striatum in at least three sections. Given (1) that cerebral arteries and veins almost never run in parallel in rats [62], (2) that arteries and arterioles stain for alkaline phosphatase, an arterial and arteriole cell marker [18], and (3) that more peri-arteriole perivascular macrophages than peri-venule perivascular macrophages are found in rodent brain [38,63,64], these latter vessels were considered to correspond to the anterior choroid artery and posterior striate artery, respectively.
4.10. Data Presentation and Statistical Analysis
All data are reported as means ± standard error of the mean (SEM). Food intake after ip [S71,C152]mIL-1β-Alexa-488 conjugate or saline administration was analyzed with one-way ANOVA. Social interaction after icv [S71,C152]mIL-1β ± saporin or PBS (pre-treatment) was expressed as time that the test rat spent in exploration of the juvenile conspecific and compared using one-way repeated measures ANOVA. Two-way ANOVA was used to determine the effects of icv [S71,C152]mIL-1ß±saporin or PBS pretreatment on subsequent icv rrIL-1ß-induced changes in food intake. The effects of pre-treatment and treatment on time spent in social interactions and locomotor activity were compared using a three-way repeated measures ANOVA. Finally, the effect of icv pre-treatment with [S71,C152]mIL-1β ± saporin or PBS on the number ED2-positive cells was analyzed using one-way ANOVA. Significant effects were further analyzed by Newman–Keuls post hoc tests. When normality and equal variance criteria were not met, non-parametric Mann–Whitney tests or ANOVA on ranks were performed. In the latter case, significant effects were further analyzed by Dunn’s post hoc test. In all cases, a level of p < 0.05 was considered as statistically significant.
5. Conclusions
In conclusion, the present study confirms that IL-1β-saporin is a useful experimental tool to study the brain cell types mediating sickness behavior, and shows that rat brain perivascular macrophages are not involved in bringing about IL-1ß-induced sickness behavior. Further research will need to determine which cell types, beyond arcuate hypothalamic neurons, play a role in mediating features of sickness behavior after IL-1ß injection in rats.
Author Contributions
L.C. investigation, R.D. conceptualization and manuscript editing and J.P.K. conceptualization, investigation, supervision and writing—original draft preparation editing. All authors have read and agreed to the published version of the manuscript.
Funding
RD’s research is supported by the National Institutes of Health, United States [R01 CA193522, R01 NS073939, and an MD Anderson Cancer Center Support Grant (P30 CA016672)).
Institutional Review Board Statement
Studies were carried out in accordance with the INRA Quality Reference system (www.international.inra.fr/content/download/947/11111/file/requirements.pdf accessed on 1 September 2014), approved by the Aquitaine Poitou-Charente regional ethical committee on animal experiments (AP 1 March 2004) and followed European recommendations on animal research (European Council Directive of 24 November 1986 (86/609/EEC) and European Parliament and Council Directive of 22 September 2010 (2010/63/UE)).
Data Availability Statement
Data is contained within the article.
Acknowledgments
The authors would like to thank Adrian Bristow (National Institute for Biological Standards and Control, Potters Bar, UK) for the synthesis of the IL-1β-saporin conjugate.
Conflicts of Interest
R.D. has received an honorarium from Compass Pathways UK not related to the objective of this study.
References
- Hart, B.L. Biological basis of the behavior of sick animals. Neurosci. Biobehav. Rev. 1988, 12, 123–137. [Google Scholar] [CrossRef]
- Sylvia, K.E.; Demas, G.E. A return to wisdom: Using sickness behaviors to integrate ecological and translational research. Integr. Comp. Biol. 2017, 57, 1204–1213. [Google Scholar] [CrossRef]
- Lasselin, J.; Schedlowski, M.; Karshikoff, B.; Engler, H.; Lekander, M.; Konsman, J.P. Comparison of bacterial lipopolysaccharide-induced sickness behavior in rodents and humans: Relevance for symptoms of anxiety and depression. Neurosci. Biobehav. Rev. 2020, 115, 15–24. [Google Scholar] [CrossRef]
- Rietschel, E.T.; Schletter, J.; Weidemann, B.; El-Samalouti, V.; Mattern, T.; Zähringer, U.; Seydel, U.; Brade, H.; Flad, H.D.; Kusumoto, S.; et al. Lipopolysaccharide and peptidoglycan: Cd14-dependent bacterial inducers of inflammation. Microb. Drug Resist. 1998, 4, 37–44. [Google Scholar] [CrossRef]
- Pedra, J.H.; Cassel, S.L.; Sutterwala, F.S. Sensing pathogens and danger signals by the inflammasome. Curr. Opin. Immunol. 2009, 21, 10–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinarello, C.A. Il-1: Discoveries, controversies and future directions. Eur. J. Immunol. 2010, 40, 599–606. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, D.O.; Kluger, M.J.; Vander, A.J. Suppression of food intake during infection: Is interleukin-1 involved? Am. J. Clin. Nutr. 1985, 42, 1179–1182. [Google Scholar] [CrossRef] [PubMed]
- Moldawer, L.L.; Andersson, C.; Gelin, J.; Lundholm, K.G. Regulation of food intake and hepatic protein synthesis by recombinant-derived cytokines. Am. J. Physiol. 1988, 254, G450–G456. [Google Scholar] [CrossRef] [PubMed]
- Hellerstein, M.K.; Meydani, S.N.; Meydani, M.; Wu, K.; Dinarello, C.A. Interleukin-1-induced anorexia in the rat. Influence of prostaglandins. J. Clin. Investig. 1989, 84, 228–235. [Google Scholar] [CrossRef]
- Crestani, F.; Seguy, F.; Dantzer, R. Behavioural effects of peripherally injected interleukin-1: Role of prostaglandins. Brain Res. 1991, 542, 330–335. [Google Scholar] [CrossRef]
- Bluthé, R.M.; Dantzer, R.; Kelley, K.W. Effects of interleukin-1 receptor antagonist on the behavioral effects of lipopolysaccharide in rat. Brain Res. 1992, 573, 318–320. [Google Scholar] [CrossRef]
- Swiergiel, A.H.; Smagin, G.N.; Johnson, L.J.; Dunn, A.J. The role of cytokines in the behavioral responses to endotoxin and influenza virus infection in mice: Effects of acute and chronic administration of the interleukin-1-receptor antagonist (il-1ra). Brain Res. 1997, 776, 96–104. [Google Scholar] [CrossRef]
- Konsman, J.P.; Parnet, P.; Dantzer, R. Cytokine-induced sickness behaviour: Mechanisms and implications. Trends Neurosci. 2002, 25, 154–159. [Google Scholar] [CrossRef]
- Ching, S.; Zhang, H.; Belevych, N.; He, L.; Lai, W.; Pu, X.A.; Jaeger, L.B.; Chen, Q.; Quan, N. Endothelial-specific knockdown of interleukin-1 (IL-1) type 1 receptor differentially alters CNS responses to IL-1 depending on its route of administration. J. Neurosci. 2007, 27, 10476–10486. [Google Scholar] [CrossRef]
- Chaskiel, L.; Bristow, A.D.; Bluthe, R.M.; Dantzer, R.; Blomqvist, A.; Konsman, J.P. Interleukin-1 reduces food intake and body weight in rat by acting in the arcuate hypothalamus. Brain Behav. Immun. 2019, 81, 560–573. [Google Scholar] [CrossRef]
- Cheunsuang, O.; Stewart, A.L.; Morris, R. Differential uptake of molecules from the circulation and csf reveals regional and cellular specialisation in cns detection of homeostatic signals. Cell Tissue Res. 2006, 325, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Morita, S.; Miyata, S. Different vascular permeability between the sensory and secretory circumventricular organs of adult mouse brain. Cell Tissue Res. 2012, 349, 589–603. [Google Scholar] [CrossRef]
- Konsman, J.P.; Vigues, S.; Mackerlova, L.; Bristow, A.; Blomqvist, A. Rat brain vascular distribution of interleukin-1 type-1 receptor immunoreactivity: Relationship to patterns of inducible cyclooxygenase expression by peripheral inflammatory stimuli. J. Comp. Neurol. 2004, 472, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Nemeth, D.P.; McKim, D.B.; Zhu, L.; DiSabato, D.J.; Berdysz, O.; Gorantla, G.; Oliver, B.; Witcher, K.G.; Wang, Y.; et al. Cell-type-specific interleukin 1 receptor 1 signaling in the brain regulates distinct neuroimmune activities. Immunity 2019, 50, 317–333 e316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dam, A.M.; Brouns, M.; Louisse, S.; Berkenbosch, F. Appearance of interleukin-1 in macrophages and in ramified microglia in the brain of endotoxin-treated rats: A pathway for the induction of non-specific symptoms of sickness? Brain Res. 1992, 588, 291–296. [Google Scholar] [CrossRef]
- Konsman, J.P.; Kelley, K.; Dantzer, R. Temporal and spatial relationships between lipopolysaccharide-induced expression of fos, interleukin-1beta and inducible nitric oxide synthase in rat brain. Neuroscience 1999, 89, 535–548. [Google Scholar] [CrossRef]
- Quan, N.; Sundar, S.K.; Weiss, J.M. Induction of interleukin-1 in various brain regions after peripheral and central injections of lipopolysaccharide. J. Neuroimmunol. 1994, 49, 125–134. [Google Scholar] [CrossRef]
- Konsman, J.P.; Tridon, V.; Dantzer, R. Diffusion and action of intracerebroventricularly injected interleukin-1 in the CNS. Neuroscience 2000, 101, 957–967. [Google Scholar] [CrossRef]
- Knoll, J.G.; Krasnow, S.M.; Marks, D.L. Interleukin-1beta signaling in fenestrated capillaries is sufficient to trigger sickness responses in mice. J. Neuroinflammation 2017, 14, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spadaro, F.; Dunn, A.J. Intracerebroventricular administration of interleukin-1 to mice alters investigation of stimuli in a novel environment. Brain Behav. Immun. 1990, 4, 308–322. [Google Scholar] [CrossRef]
- Kent, S.; Bluthe, R.M.; Dantzer, R.; Hardwick, A.J.; Kelley, K.W.; Rothwell, N.J.; Vannice, J.L. Different receptor mechanisms mediate the pyrogenic and behavioral effects of interleukin 1. Proc. Natl. Acad. Sci. USA 1992, 89, 9117–9120. [Google Scholar] [CrossRef] [Green Version]
- Plata-Salamán, C.R. Meal patterns in response to the intracerebroventricular administration of interleukin-1 beta in rats. Physiol. Behav. 1994, 55, 727–733. [Google Scholar] [CrossRef]
- Schiltz, J.C.; Sawchenko, P.E. Distinct brain vascular cell types manifest inducible cyclooxygenase expression as a function of the strength and nature of immune insults. J. Neurosci. 2002, 22, 5606–5618. [Google Scholar] [CrossRef] [Green Version]
- Schnurch, H.; Risau, W. Expression of tie-2, a member of a novel family of receptor tyrosine kinases, in the endothelial cell lineage. Development 1993, 119, 957–968. [Google Scholar] [CrossRef]
- Gomez Perdiguero, E.; Klapproth, K.; Schulz, C.; Busch, K.; Azzoni, E.; Crozet, L.; Garner, H.; Trouillet, C.; de Bruijn, M.F.; Geissmann, F.; et al. Tissue-resident macrophages originate from yolk-sac-derived erythro-myeloid progenitors. Nature 2015, 518, 547–551. [Google Scholar] [CrossRef]
- Nishiyori, A.; Minami, M.; Ohtani, Y.; Takami, S.; Yamamoto, J.; Kawaguchi, N.; Kume, T.; Akaike, A.; Satoh, M. Localization of fractalkine and Cx3cr1 mrnas mRNAs in rat brain: Does fractalkine play a role in signaling from neuron to microglia? FEBS Lett. 1998, 429, 167–172. [Google Scholar] [CrossRef] [Green Version]
- Goldmann, T.; Wieghofer, P.; Jordao, M.J.; Prutek, F.; Hagemeyer, N.; Frenzel, K.; Amann, L.; Staszewski, O.; Kierdorf, K.; Krueger, M.; et al. Origin, fate and dynamics of macrophages at central nervous system interfaces. Nat. Immunol. 2016, 17, 797–805. [Google Scholar] [CrossRef]
- Sugiyama, D.; Kusuhara, H.; Taniguchi, H.; Ishikawa, S.; Nozaki, Y.; Aburatani, H.; Sugiyama, Y. Functional characterization of rat brain-specific organic anion transporter (oatp14) at the blood-brain barrier: High affinity transporter for thyroxine. J. Biol. Chem. 2003, 278, 43489–43495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, L.M.; Woodford, K.; Zhou, M.; Black, D.S.; Haggerty, J.E.; Tate, E.H.; Grindstaff, K.K.; Mengesha, W.; Raman, C.; Zerangue, N. Expression of the thyroid hormone transporters monocarboxylate transporter-8 (slc16a2) and organic ion transporter-14 (slco1c1) at the blood-brain barrier. Endocrinology 2008, 149, 6251–6261. [Google Scholar] [CrossRef] [PubMed]
- Lang, M.F.; Salinin, S.; Ridder, D.A.; Kleesiek, J.; Hroudova, J.; Berger, S.; Schutz, G.; Schwaninger, M. A transgenic approach to identify thyroxine transporter-expressing structures in brain development. J. Neuroendocrinol. 2011, 23, 1194–1203. [Google Scholar] [CrossRef]
- Faraco, G.; Park, L.; Anrather, J.; Iadecola, C. Brain perivascular macrophages: Characterization and functional roles in health and disease. J. Mol. Med. 2017, 95, 1143–1152. [Google Scholar] [CrossRef] [Green Version]
- Prinz, M.; Masuda, T.; Wheeler, M.A.; Quintana, F.J. Microglia and central nervous system-associated macrophages-from origin to disease modulation. Annu. Rev. Immunol. 2021, 39, 251–277. [Google Scholar] [CrossRef]
- Yang, T.; Guo, R.; Zhang, F. Brain perivascular macrophages: Recent advances and implications in health and diseases. CNS Neurosci. Ther. 2019, 25, 1318–1328. [Google Scholar] [CrossRef]
- Ferreras, J.M.; Barbieri, L.; Girbes, T.; Battelli, M.G.; Rojo, M.A.; Arias, F.J.; Rocher, M.A.; Soriano, F.; Mendez, E.; Stirpe, F. Distribution and properties of major ribosome-inactivating proteins (28 s rRNA n-glycosidases) of the plant saponaria officinalis l. (caryophyllaceae). Biochim. Biophys. Acta 1993, 1216, 31–42. [Google Scholar] [CrossRef]
- Santanche, S.; Bellelli, A.; Brunori, M. The unusual stability of saporin, a candidate for the synthesis of immunotoxins. Biochem. Biophys. Res. Commun. 1997, 234, 129–132. [Google Scholar] [CrossRef]
- Wiley, R.G. Targeting toxins to neural antigens and receptors. Methods Mol. Biol. 2001, 166, 267–276. [Google Scholar]
- Munafo, M.R.; Davey Smith, G. Robust research needs many lines of evidence. Nature 2018, 553, 399–401. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.F.; Alam, M.M.; Liao, Y.; Huang, T.; Mathur, R.; Zhu, X.; Huang, Y. Targeting microglia using Cx3cr1-Cre lines: Revisiting the specificity. eNeuro 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Polfliet, M.M.; Goede, P.H.; van Kesteren-Hendrikx, E.M.; van Rooijen, N.; Dijkstra, C.D.; van den Berg, T.K. A method for the selective depletion of perivascular and meningeal macrophages in the central nervous system. J. Neuroimmunol. 2001, 116, 188–195. [Google Scholar] [CrossRef]
- Michels, M.; Avila, P.; Pescador, B.; Vieira, A.; Abatti, M.; Cucker, L.; Borges, H.; Goulart, A.I.; Junior, C.C.; Barichello, T.; et al. Microglial cells depletion increases inflammation and modifies microglial phenotypes in an animal model of severe sepsis. Mol. Neurobiol. 2019, 56, 7296–7304. [Google Scholar] [CrossRef]
- Takagi, S.; Murayama, S.; Torii, K.; Takemura-Morita, S.; Kurganov, E.; Nagaoka, S.; Wanaka, A.; Miyata, S. Depletion of microglia and macrophages with clodronate liposomes attenuates zymosan-induced fos expression and hypothermia in the adult mouse. J. Neuroimmunol. 2020, 344, 577244. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.C.; Corey, S.; Westmoreland, S.V.; Pauley, D.; Knight, H.; DeBakker, C.; Alvarez, X.; Lackner, A.A. Perivascular macrophages are the primary cell type productively infected by simian immunodeficiency virus in the brains of macaques: Implications for the neuropathogenesis of aids. J. Exp. Med. 2001, 193, 905–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.K.; Alvarez, X.; Fisher, J.; Bronfin, B.; Westmoreland, S.; McLaurin, J.; Williams, K. CD163 identifies perivascular macrophages in normal and viral encephalitic brains and potential precursors to perivascular macrophages in blood. Am. J. Pathol. 2006, 168, 822–834. [Google Scholar] [CrossRef] [Green Version]
- Ransohoff, R.M.; Engelhardt, B. The anatomical and cellular basis of immune surveillance in the central nervous system. Nat. Rev. Immunol. 2012, 12, 623–635. [Google Scholar] [CrossRef]
- Konsman, J.P.; Veeneman, J.; Combe, C.; Poole, S.; Luheshi, G.N.; Dantzer, R. Central nervous action of interleukin-1 mediates activation of limbic structures and behavioural depression in response to peripheral administration of bacterial lipopolysaccharide. Eur. J. Neurosci. 2008, 28, 2499–2510. [Google Scholar] [CrossRef] [PubMed]
- Kanai, T.; Watanabe, M.; Okazawa, A.; Sato, T.; Yamazaki, M.; Okamoto, S.; Ishii, H.; Totsuka, T.; Iiyama, R.; Okamoto, R.; et al. Macrophage-derived IL-18-mediated intestinal inflammation in the murine model of crohn’s disease. Gastroenterology 2001, 121, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Kanai, T.; Uraushihara, K.; Totsuka, T.; Nemoto, Y.; Fujii, R.; Kawamura, T.; Makita, S.; Sawada, D.; Yagita, H.; Okumura, K.; et al. Ameliorating effect of saporin-conjugated anti-CD11b monoclonal antibody in a murine T-cell-mediated chronic colitis. J. Gastroenterol. Hepatol. 2006, 21, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Hozain, S.; Cottrell, J. CDllb+ targeted depletion of macrophages negatively affects bone fracture healing. Bone 2020, 138, 115479. [Google Scholar] [CrossRef]
- Anforth, H.R.; Bluthe, R.M.; Bristow, A.; Hopkins, S.; Lenczowski, M.J.; Luheshi, G.; Lundkvist, J.; Michaud, B.; Mistry, Y.; Van Dam, A.M.; et al. Biological activity and brain actions of recombinant rat interleukin-1alpha and interleukin-1beta. Eur. Cytokine Netw. 1998, 9, 279–288. [Google Scholar]
- Terao, A.; Matsumura, H.; Saito, M. Interleukin-1 induces slow-wave sleep at the prostaglandin D2-sensitive sleep-promoting zone in the rat brain. J. Neurosci. 1998, 18, 6599–6607. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Ikoma, K.; Morioka, N.; Kumagai, K.; Hashimoto, T.; Hide, I.; Nakata, Y. Interleukin-1beta induces substance p release from primary afferent neurons through the cyclooxygenase-2 system. J. Neurochem. 1999, 73, 2206–2213. [Google Scholar] [PubMed]
- Serou, M.J.; DeCoster, M.A.; Bazan, N.G. Interleukin-1 beta activates expression of cyclooxygenase-2 and inducible nitric oxide synthase in primary hippocampal neuronal culture: Platelet-activating factor as a preferential mediator of cyclooxygenase-2 expression. J. Neurosci. Res. 1999, 58, 593–598. [Google Scholar] [CrossRef]
- Boraschi, D.; Bossu, P.; Macchia, G.; Ruggiero, P.; Tagliabue, A. Structure-function relationship in the IL-1 family. Front. Biosci. 1996, 1, d270–d308. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, G.; Strubbe, J.H. Time-dependent effects of neuropeptide Y infusion in the paraventricular hypothalamus on ingestive and associated behaviors in rats. Physiol. Behav. 2003, 79, 575–580. [Google Scholar] [CrossRef]
- Bolognesi, A.; Tazzari, P.L.; Olivieri, F.; Polito, L.; Lemoli, R.; Terenzi, A.; Pasqualucci, L.; Falini, B.; Stirpe, F. Evaluation of immunotoxins containing single-chain ribosome-inactivating proteins and an anti-CD22 monoclonal antibody (om124): In vitro and in vivo studies. Br. J. Haematol. 1998, 101, 179–188. [Google Scholar] [CrossRef]
- Pistillo, M.P.; Tazzari, P.L.; Palmisano, G.L.; Pierri, I.; Bolognesi, A.; Ferlito, F.; Capanni, P.; Polito, L.; Ratta, M.; Pileri, S.; et al. CTLA-4 is not restricted to the lymphoid cell lineage and can function as a target molecule for apoptosis induction of leukemic cells. Blood 2003, 101, 202–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scremin, O.U. Cerebral vascular system. In The Rat Nervous System; Paxinos, G., Ed.; Academic Press: San Diego, CA, USA, 1995; pp. 3–35. [Google Scholar]
- Mato, M.; Ookawara, S.; Sakamoto, A.; Aikawa, E.; Ogawa, T.; Mitsuhashi, U.; Masuzawa, T.; Suzuki, H.; Honda, M.; Yazaki, Y.; et al. Involvement of specific macrophage-lineage cells surrounding arterioles in barrier and scavenger function in brain cortex. Proc. Natl. Acad. Sci. USA 1996, 93, 3269–3274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ookawara, S.; Mitsuhashi, U.; Suminaga, Y.; Mato, M. Study on distribution of pericyte and fluorescent granular perithelial (fgp) cell in the transitional region between arteriole and capillary in rat cerebral cortex. Anat. Rec. 1996, 244, 257–264. [Google Scholar] [CrossRef]
Figure 1.
Effects of intracerebroventricular IL-1ß-saporin conjugate administration on brain perivascular cells in the organum vasculosum laminae terminalis (OVLT; A,C,D) and amygdala (B,E,F). In these structures, the IL-1ß-saporin conjugate reduced the number of CD163 immunoreactive (CD163 i.r.) cells (A–F) without affecting labelling for the endothelial cell marker von Willebrand factor (C,D) and the astrocyte marker glial fibrillary acidic protein (E,F). Brown labeling indicates von Willebrand factor (C,D) and glial fibrillary acidic protein (E,F) and black staining CD163. * p < 0.05; *** p < 0.001. IL1-SAP compared to IL1 + SAP group. IL1: interleukin-1ß; IL1-SAP: interleukin-1ß conjugated to saporin; IL1 + SAP: interleukin-1ß not conjugated to saporin; PBS: phosphate-buffered saline; SAP: saporin. Group sizes: n = 5–9. Scale bar represents 100 µm.
Figure 1.
Effects of intracerebroventricular IL-1ß-saporin conjugate administration on brain perivascular cells in the organum vasculosum laminae terminalis (OVLT; A,C,D) and amygdala (B,E,F). In these structures, the IL-1ß-saporin conjugate reduced the number of CD163 immunoreactive (CD163 i.r.) cells (A–F) without affecting labelling for the endothelial cell marker von Willebrand factor (C,D) and the astrocyte marker glial fibrillary acidic protein (E,F). Brown labeling indicates von Willebrand factor (C,D) and glial fibrillary acidic protein (E,F) and black staining CD163. * p < 0.05; *** p < 0.001. IL1-SAP compared to IL1 + SAP group. IL1: interleukin-1ß; IL1-SAP: interleukin-1ß conjugated to saporin; IL1 + SAP: interleukin-1ß not conjugated to saporin; PBS: phosphate-buffered saline; SAP: saporin. Group sizes: n = 5–9. Scale bar represents 100 µm.

Figure 2.
Effects of intracerebroventricular IL-1ß-saporin conjugate administration on subsequent IL-1β-induced sickness behavior. Intracerebroventricular pre-treatment with IL-1β-saporin conjugate did not affect the reduction in food intake and social interaction after intracerebroventricular IL-1ß administration (A,B). Intracerebroventricular administration of IL-1β-saporin conjugate did not affect the reduction in food intake and locomotor activity to subsequent intraperitoneal IL-1ß injection (C,D). IL1: interleukin-1ß; IL1-SAP: interleukin-1ß conjugated to saporin; IL1 + SAP: interleukin-1ß not conjugated to saporin; PBS: phosphate-buffered saline; Sal: saline; SAP: saporin. Group sizes: n = 5–9.
Figure 2.
Effects of intracerebroventricular IL-1ß-saporin conjugate administration on subsequent IL-1β-induced sickness behavior. Intracerebroventricular pre-treatment with IL-1β-saporin conjugate did not affect the reduction in food intake and social interaction after intracerebroventricular IL-1ß administration (A,B). Intracerebroventricular administration of IL-1β-saporin conjugate did not affect the reduction in food intake and locomotor activity to subsequent intraperitoneal IL-1ß injection (C,D). IL1: interleukin-1ß; IL1-SAP: interleukin-1ß conjugated to saporin; IL1 + SAP: interleukin-1ß not conjugated to saporin; PBS: phosphate-buffered saline; Sal: saline; SAP: saporin. Group sizes: n = 5–9.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chaskiel, L.; Dantzer, R.; Konsman, J.P. Brain Perivascular Macrophages Do Not Mediate Interleukin-1-Induced Sickness Behavior in Rats. Pharmaceuticals 2021, 14, 1030. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14101030
AMA Style
Chaskiel L, Dantzer R, Konsman JP. Brain Perivascular Macrophages Do Not Mediate Interleukin-1-Induced Sickness Behavior in Rats. Pharmaceuticals. 2021; 14(10):1030. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14101030
Chicago/Turabian StyleChaskiel, Léa, Robert Dantzer, and Jan Pieter Konsman. 2021. "Brain Perivascular Macrophages Do Not Mediate Interleukin-1-Induced Sickness Behavior in Rats" Pharmaceuticals 14, no. 10: 1030. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14101030
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.