
Pipecolisporin, a Novel Cyclic Peptide with Antimalarial and Antitrypanosome Activities from a Wheat Endophytic Nigrospora oryzae

 ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results and Discussion
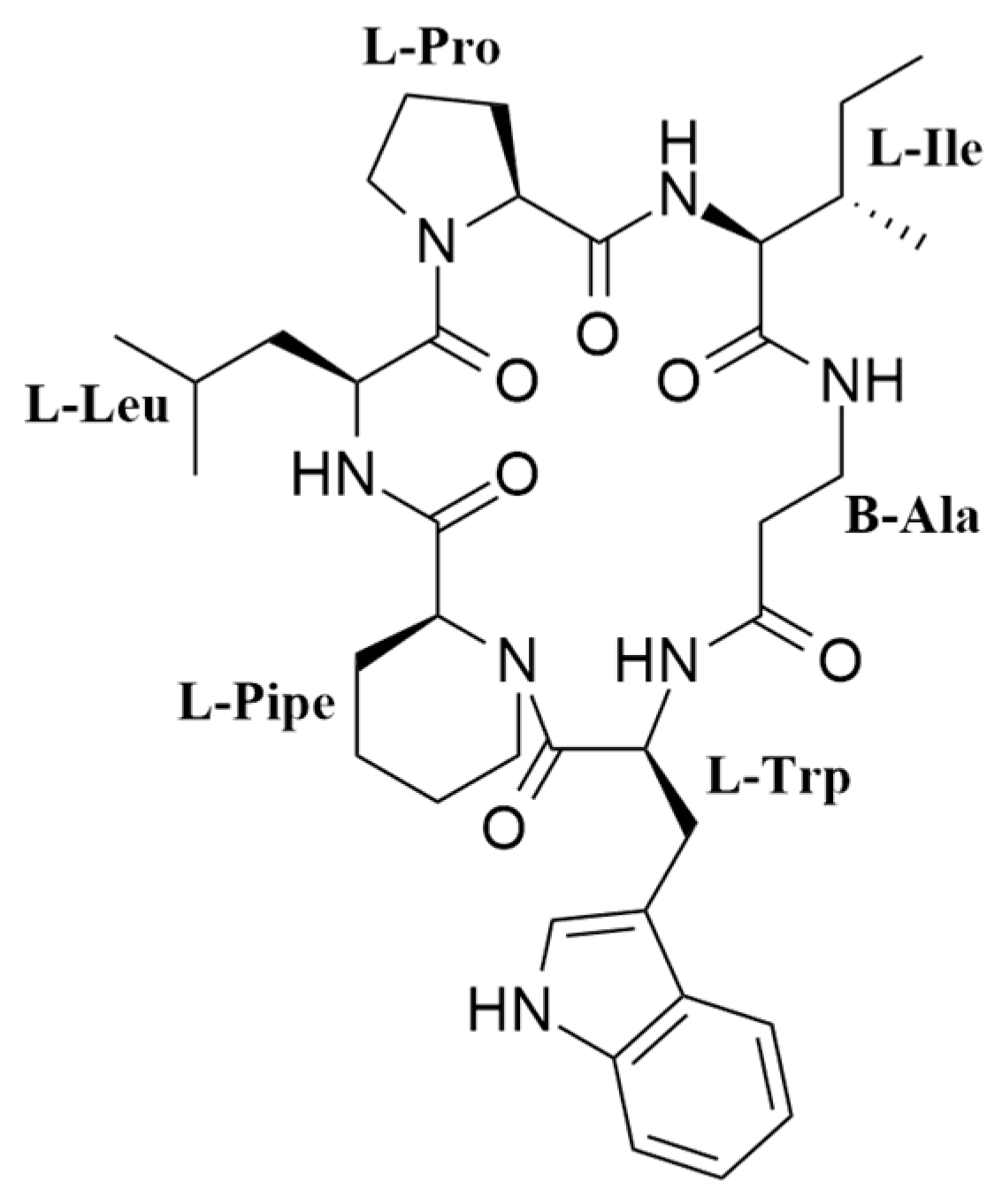
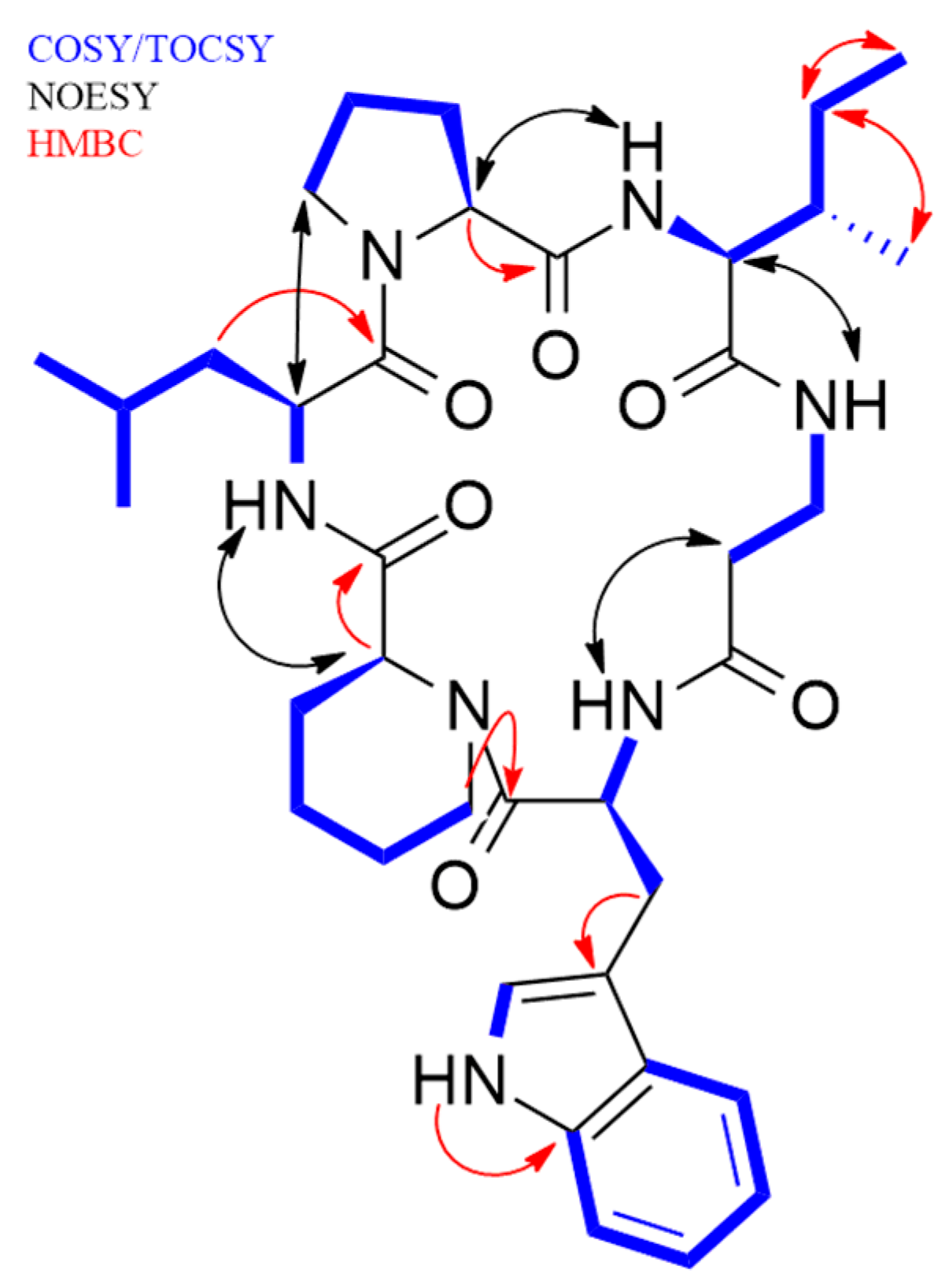
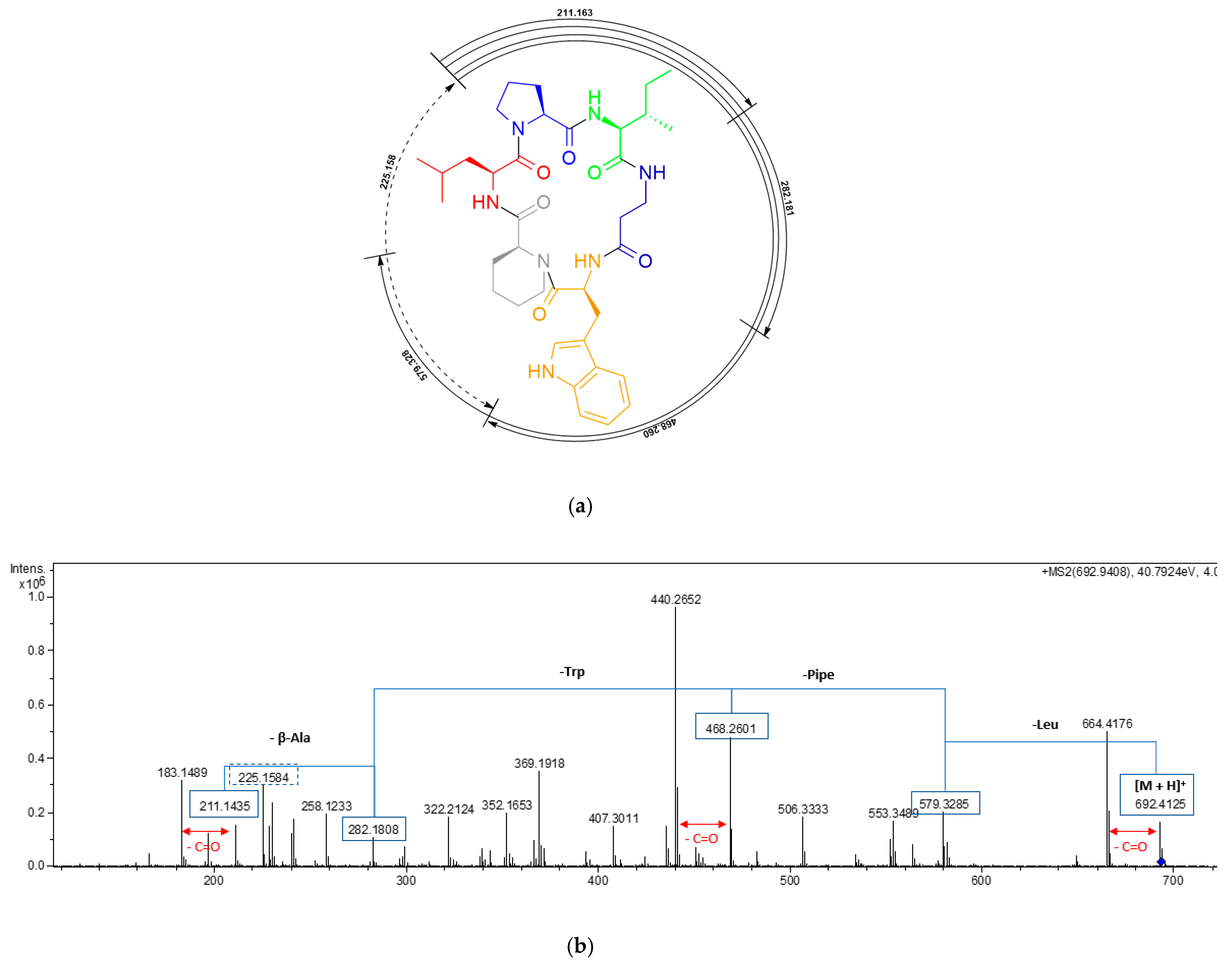
2.1. Isolation and Structural Elucidation of Pipecolisporin
2.2. Biological Activity
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Microbial Isolation and Identification
3.3. Fungal Solid-State Fermentation (SSF)
3.4. Isolation and Identification of Pipecolisporin
Marfey’s Analysis of Compound 1
3.5. Biological Assays
3.5.1. Plasmodium Falciparum 3D7 Lactase Dehydrogenase in Vitro Assay
3.5.2. Transgenic Trypanosoma cruzi β-D-galactosidase in vitro Assay
3.5.3. MTT/Cell Viability Assay
3.5.4. Antimicrobial Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nisa, H.; Kamili, A.N.; Nawchoo, I.A.; Shafi, S.; Shameem, N.; Bandh, S.A. Fungal endophytes as prolific source of phytochemicals and other bioactive natural products: A review. Microb. Pathog. 2015, 82, 50–59. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Vasundhara, M.; Sudhakara Reddy, M.; Kumar, A. Secondary metabolites from endophytic fungi and their biological activities. In New and Future Developments in Microbial Biotechnology and Bioengineering: Microbial Secondary Metabolites Biochemistry and Applications; Gupta, V.K., Pandey, A., Eds.; Elsevier: Oxford, UK, 2019; pp. 237–258. [Google Scholar]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebada, S.S.; Eze, P.; Okoye, F.B.C.; Esimone, C.O.; Proksch, P. The Fungal Endophyte Nigrospora oryzae Produces Quercetin Monoglycosides Previously Known Only from Plants. ChemistrySelect 2016, 1, 2767–2771. [Google Scholar] [CrossRef]
- Ganley, R.J.; Brunsfeld, S.J.; Newcombe, G. A community of unknown, endophytic fungi in western white pine. Proc. Natl. Acad. Sci. USA 2004, 101, 10107–10112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vector-Borne Diseases. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 17 November 2020).
- Tajuddeen, N.; Van Heerden, F.R. Antiplasmodial natural products: An update. Malar. J. 2019, 18, 404. [Google Scholar] [CrossRef] [Green Version]
- Cota, B.B.; Rosa, L.H.; Caligiorne, R.B.; Rabello, A.L.T.; Almeida Alves, T.M.; Rosa, C.A.; Zani, C.L. Altenusin, a biphenyl isolated from the endophytic fungus Alternaria sp., inhibits trypanothione reductase from Trypanosoma cruzi. FEMS Microbiol. Lett. 2008, 285, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Tansuwan, S.; Pornpakakul, S.; Roengsumran, S.; Petsom, A.; Muangsin, N.; Sihanonta, P.; Chaichit, N. Antimalarial Benzoquinones from an Endophytic Fungus, Xylaria sp. J. Nat. Prod. 2007, 70, 1620–1623. [Google Scholar] [CrossRef]
- Kontnik, R.; Clardy, J. Codinaeopsin, an Antimalarial Fungal Polyketide. Org. Lett. 2008, 10, 4149–4151. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Abdallah, H.M.; Elkhayat, E.S.; Al Musayeib, N.M.; Asfour, H.Z.; Zayed, M.F.; Mohamed, G.A. Fusaripeptide A: New antifungal and anti-malarial cyclodepsipeptide from the endophytic fungus Fusarium sp. J. Asian Nat. Prod. Res. 2018, 20, 75–85. [Google Scholar] [CrossRef]
- Annang, F.; Pérez-Moreno, G.; González-Menéndez, V.; Lacret, R.; Pérez-Victoria, I.; Martín, J.; Cantizani, J.; de Pedro, N.; Choquesillo-Lazarte, D.; Ruiz-Pérez, L.M.; et al. Strasseriolides A-D, A Family of Antiplasmodial Macrolides Isolated from the Fungus Strasseria geniculata CF-247251. Org. Lett. 2020, 22, 6709–6713. [Google Scholar] [CrossRef]
- Wang, G.; Liu, Z.; Lin, R.; Li, E.; Mao, Z.; Ling, J.; Yang, Y.; Yin, W.B.; Xie, B. Biosynthesis of Antibiotic Leucinostatins in Bio-control Fungus Purpureocillium lilacinum and Their Inhibition on Phytophthora Revealed by Genome Mining. PLoS Pathog. 2016, 12, 1005685. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.M.; Jerkovic, A.; Truong, T.T.; Foote, S.J.; McCarthy, J.S.; McMorran, B.J. Griseofulvin impairs intraerythrocytic growth of Plasmodium falciparum through ferrochelatase inhibition but lacks activity in an experimental human infection study. Sci. Rep. 2017, 7, 41975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barúa, J.E.; de la Cruz, M.; de Pedro, N.; Cautain, B.; Hermosa, R.; Cardoza, R.E.; Gutiérrez, S.; Monte, E.; Vicente, F.; Collado, I.G. Synthesis of Trichodermin Derivatives and Their Antimicrobial and Cytotoxic Activities. Molecules 2019, 24, 3811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, B.; Wang, Y.; Sun, X.; Tang, K. Bioactive natural products from endophytes: A review. Appl. Biochem. Microbiol. 2008, 44, 136–142. [Google Scholar] [CrossRef]
- Linington, R.G.; González, J.; Ureña, L.D.; Romero, L.I.; Ortega-Barría, E.; Gerwick, W.H. Venturamides A and B: Antimalarial constituents of the Panamanian marine cyanobacterium Oscillatoria sp. J. Nat. Prod. 2007, 70, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Búa, J.; Fichera, L.E.; Fuchs, A.G.; Potenza, M.; Dubin, M.; Wenger, R.O.; Moretti, G.; Scabone, C.M.; Ruiz, A.M. Anti-Trypanosoma cruzi effects of cyclosporin A derivatives: Possible role of a P-glycoprotein and parasite cyclophilins. Parasitology 2008, 135, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Sweeney-Jones, A.M.; Gagaring, K.; Antonova-Koch, J.; Zhou, H.; Mojib, N.; Soapi, K.; Skolnick, J.; McNamara, C.W.; Kubanek, J. Antimalarial Peptide and Polyketide Natural Products from the Fijian Marine Cyanobacterium Moorea producens. Mar. Drugs 2020, 18, 167. [Google Scholar] [CrossRef]
- Lee, Y.; Phat, C.; Hong, S.C. Structural diversity of marine cyclic peptides and their molecular mechanisms for anticancer, antibacterial, antifungal, and other clinical applications. Peptides 2017, 95, 94–105. [Google Scholar] [CrossRef]
- Stawikowski, M.; Cudic, P. A novel strategy for the solid-phase synthesis of cyclic lipodepsipeptides. Tetrahedron Lett. 2006, 47, 8587–8590. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Fukushima, T.; Tsujino, Y.; Fujimori, T. Nigrosporins A and B, New Phytotoxic and Antibacterial Metabolites Produced by a Fungus Nigrospora oryzae. Biosci. Biotechnol. Biochem. 1997, 61, 1848–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.J.; Yuan, W.; Liao, X.J.; Han, B.N.; Wang, S.P.; Li, Z.Y.; Xu, S.H.; Zhang, W.; Lin, H.W. Oryzamides A-E, Cyclodepsipeptides from the Sponge-Derived Fungus Nigrospora oryzae PF18. J. Nat. Prod. 2016, 79, 2045–2052. [Google Scholar] [CrossRef]
- Ortiz-López, F.J.; Carretero-Molina, D.; Sánchez-Hidalgo, M.; Martín, J.; González, I.; Román-Hurtado, F.; Cruz, M.; García-Fernández, S.; Reyes, F.; Deisinger, J.P.; et al. Cacaoidin, First Member of the New Lanthidin RiPP Family. Angew. Chem. Int. Ed. 2020, 59, 12654–12658. [Google Scholar] [CrossRef]
- Gonzalez-Menendez, V.; Martin, J.; Siles, J.A.; Gonzalez-Tejero, M.R.; Reyes, F.; Platas, G.; Tormo, J.R.; Genilloud, O. Biodiversity and chemotaxonomy of Preussia isolates from the Iberian Peninsula. Mycol. Prog. 2017, 16, 713–728. [Google Scholar] [CrossRef]
- Blaxter, M.; Mann, J.; Chapman, T.; Thomas, F.; Whitton, C.; Floyd, R.; Abebe, E. Defining operational taxonomic units using DNA barcode data. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1935–1943. [Google Scholar] [CrossRef]
- Hofstetter, V.; Buyck, B.; Eyssartier, G.; Schnee, S.; Gindro, K. The unbearable lightness of sequenced-based identification. Fungal Divers. 2019, 96, 243–284. [Google Scholar] [CrossRef] [Green Version]
- Widmer, T.; Kirk, A.; Kirk, G.; Guermache, F. Foliar and Cane Rot of Arundo donax Caused by Nigrospora oryzae in Europe. Plant Dis. 2006, 90, 1107. [Google Scholar] [CrossRef]
- Pelaez, F.; Cabello, A.; Platas, G.; Díez, M.T.; Del Val, A.G.; Basilio, A.; Martán, I.; Vicente, F.; Bills, G.F.; Giacobbe, R.A.; et al. The Discovery of Enfumafungin, a Novel Antifungal Compound Produced by an Endophytic Hormonema Species Biological Activity and Taxonomy of the Producing Organisms. Syst. Appl. Microbiol. 2000, 23, 333–343. [Google Scholar] [CrossRef]
- Pérez-Moreno, G.; Cantizani, J.; Sánchez-Carrasco, P.; Ruiz-Pérez, L.M.; Martín, J.; El Aouad, N.; Pérez-Victoria, I.; Tormo, J.R.; González-Menendez, V.; González, I.; et al. Discovery of New Compounds Active against Plasmodium falciparum by High Throughput Screening of Microbial Natural Products. PLoS ONE 2016, 11, e0145812. [Google Scholar] [CrossRef]
- Annang, F.; Pérez-Moreno, G.; García-Hernández, R.; Cordon-Obras, C.; Martín, J.; Tormo, J.R.; Rodríguez, L.; De Pedro, N.; Gómez-Pérez, V.; Valente, M.; et al. High-Throughput Screening Platform for Natural Product-Based Drug Discovery Against 3 Neglected Tropical Diseases: Human African Trypanosomiasis, Leishmaniasis, and Chagas Disease. J. Biomol. Screen. 2015, 20, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Präbst, K.; Engelhardt, H.; Ringgeler, S.; Hübner, H. Basic colorimetric proliferation assays: MTT, WST, and resazurin. In Methods in Molecular Biology; Gilbert, D.F., Friedrich, O., Eds.; Springer Nature: New York, NY, USA, 2017; Volume 1601, pp. 1–17. [Google Scholar]
- Koagne, R.R.; Annang, F.; Cautain, B.; Martín, J.; Pérez-Moreno, G.; Thierry, M.; Bitchagno, G.; González-Pacanowska, D.; Vicente, F.; Simo, I.K.; et al. Cytotoxicity and antiplasmodial activity of phenolic derivatives from Albizia zygia (DC.) J.F. Macbr. (Mimosae). BMC Complement. Med. Ther. 2020, 20, 8. [Google Scholar] [CrossRef] [PubMed]
- Audoin, C.; Bonhomme, D.; Ivanisevic, J.; Cruz, M.D.l.; Cautain, B.; Monteiro, M.C.; Reyes, F.; Rios, L.; Perez, T.; Thomas, O.P. Balibalosides, an Original Family of Glucosylated Sesterterpenes Produced by the Mediterranean Sponge Oscarella balibaloi. Mar. Drugs 2013, 11, 1477–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Position | δH, m, J (Hz) | δC, Mult | HMBC (H to C) | NOESY |
|---|---|---|---|---|---|
| Pro | 1 | 3.99, m | 62.9, CH | CO Pro | NH Ile |
| 2 | 2.19, m, 1.77, m | 29.4, CH2 | CO Pro | ||
| 3 | 1.99, m, 1.84, m | 25.7, CH2 | |||
| 4 | 3.72, m 3.56, m | 47.3, CH2 | Leu 1 | ||
| CO | 171.8, C | ||||
| Ile | NH amide | 6.47, d, (8.0) | Pro 1 | ||
| 1 | 4.01, m | 57.5, CH | NH β-Ala | ||
| 2 | 1.91, m | 37.0, CH | |||
| 3 | 1.30, m, 1.08, m | 24.7, CH2 | |||
| 4 | 0.82, m | 12.1, CH3 | Ile 3 | ||
| 2-Me | 0.81, m | 16.2, CH3 | Ile 3 | ||
| CO | 171.2, C | ||||
| β-Ala | NH amide | 7.05, m | CO Ile | Ile 1 | |
| 1 | 2.43 m, 2.11, m | 35.2 CH2 | |||
| 2 | 3.71, m, 3.12, m | 35.0, CH2 | NH Trp | ||
| CO | 171.0, C | ||||
| Trp | NH amide | 8.56, d, (8.9) | |||
| 1 | 4.49, m | 51.8, CH | Trp 2 | ||
| 2 | 3.06, d, (14.8) 3.17, dd, (14.8, 4.1) | 27.6, CH2 | Trp 1, Trp 3 | ||
| 3 | 108.9, C | ||||
| 4 | 7.29, d, (2.0) | 125.0, CH | |||
| NH | 11.00, bs | Trp 5 | |||
| 5 | 136.7, C | ||||
| 6 | 7.34, d, (8.1) | 118.7, CH | |||
| 7 | 7.06, m | 121.7, CH | |||
| 8 | 6.97, m | 108.9, CH | |||
| 9 | 7.44, m, (8.0) | 118.7, CH | |||
| 10 | 127.7, C | ||||
| CO | 170.9, C | ||||
| 1 | 3.66, m | 56.7, CH | Pipe 5 | NH Leu | |
| 2 | 1.45, m, −0.70 m | 23.6, CH2 | NH Leu | ||
| Pipe | 3 | 0.99, m, 0.78, m | 20.7, CH2 | ||
| 4 | 1.25 m, 0.50 m | 23.9, CH2 | |||
| 5 | 4.32 m, 2.10 m | 39.4, CH2 | CO Trp | ||
| CO | 169.6, C | ||||
| NH amide | 8.95, d, (8.5) | CO Pipe | |||
| 1 | 4.53, m | 50.3, CH | Pro 4 | ||
| 2 | 1.85, m, 1.53, m | 38.3, CH2 | |||
| Leu | 3 | 1.62, m | 25.2, CH | CO Leu | |
| 4 | 0.81, m | 20.6, CH3 | |||
| 4’ | 0.90, m | 23.9, CH3 | |||
| CO | 173.5, C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Pastor, I.; González-Menéndez, V.; Annang, F.; Toro, C.; Mackenzie, T.A.; Bosch-Navarrete, C.; Genilloud, O.; Reyes, F. Pipecolisporin, a Novel Cyclic Peptide with Antimalarial and Antitrypanosome Activities from a Wheat Endophytic Nigrospora oryzae. Pharmaceuticals 2021, 14, 268. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030268
Fernández-Pastor I, González-Menéndez V, Annang F, Toro C, Mackenzie TA, Bosch-Navarrete C, Genilloud O, Reyes F. Pipecolisporin, a Novel Cyclic Peptide with Antimalarial and Antitrypanosome Activities from a Wheat Endophytic Nigrospora oryzae. Pharmaceuticals. 2021; 14(3):268. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030268
Chicago/Turabian StyleFernández-Pastor, Ignacio, Victor González-Menéndez, Frederick Annang, Clara Toro, Thomas A. Mackenzie, Cristina Bosch-Navarrete, Olga Genilloud, and Fernando Reyes. 2021. "Pipecolisporin, a Novel Cyclic Peptide with Antimalarial and Antitrypanosome Activities from a Wheat Endophytic Nigrospora oryzae" Pharmaceuticals 14, no. 3: 268. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030268