
Transcriptomes of Wet Skin Biopsies Predict Outcomes after Ionizing Radiation Exposure with Potential Dosimetric Applications in a Mouse Model

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
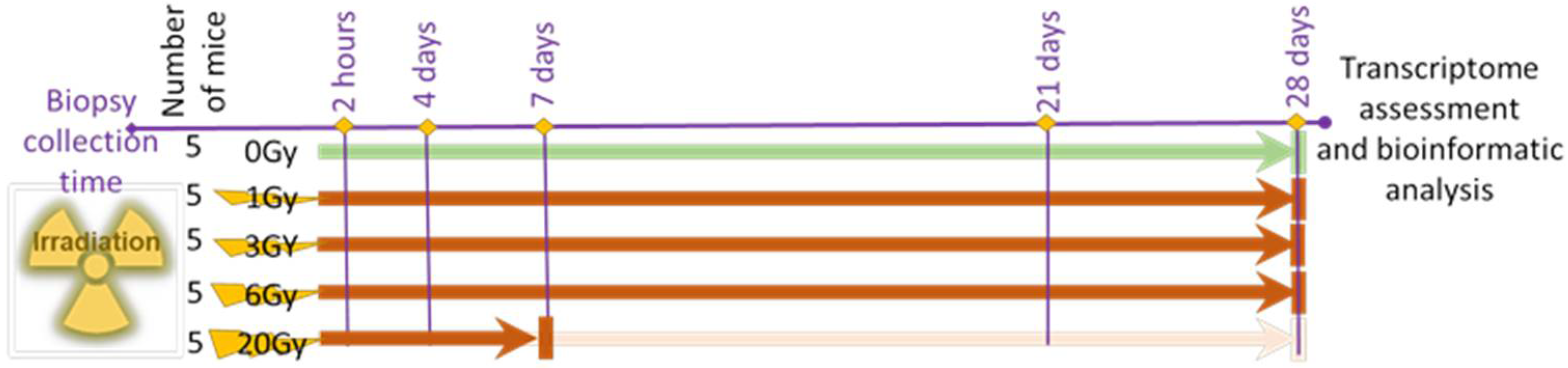
2.2. Animal Radiation Model and Sample Collection
2.3. Radiation Treatment
2.4. Samples Collection and Post-Irradiation Observation
2.5. Molecular Biology
2.6. Data Preparation and Analysis
3. Results
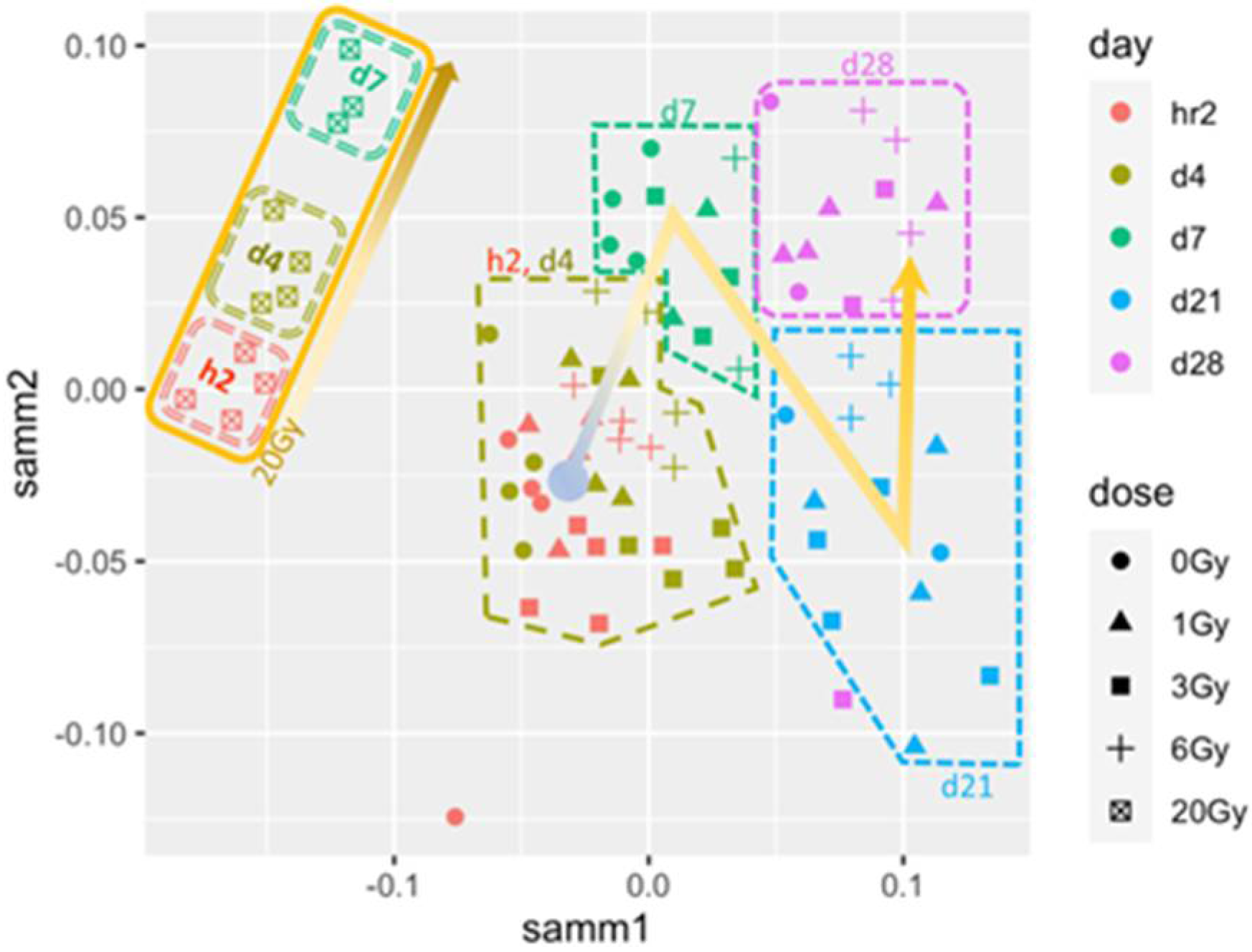
3.1. Skin Transcriptomes Predict the Short-Term Fate of Mice after IR Exposure
3.2. Idling Transcription Is a Principal Response Character in BLDs Contrary to That in BSDs
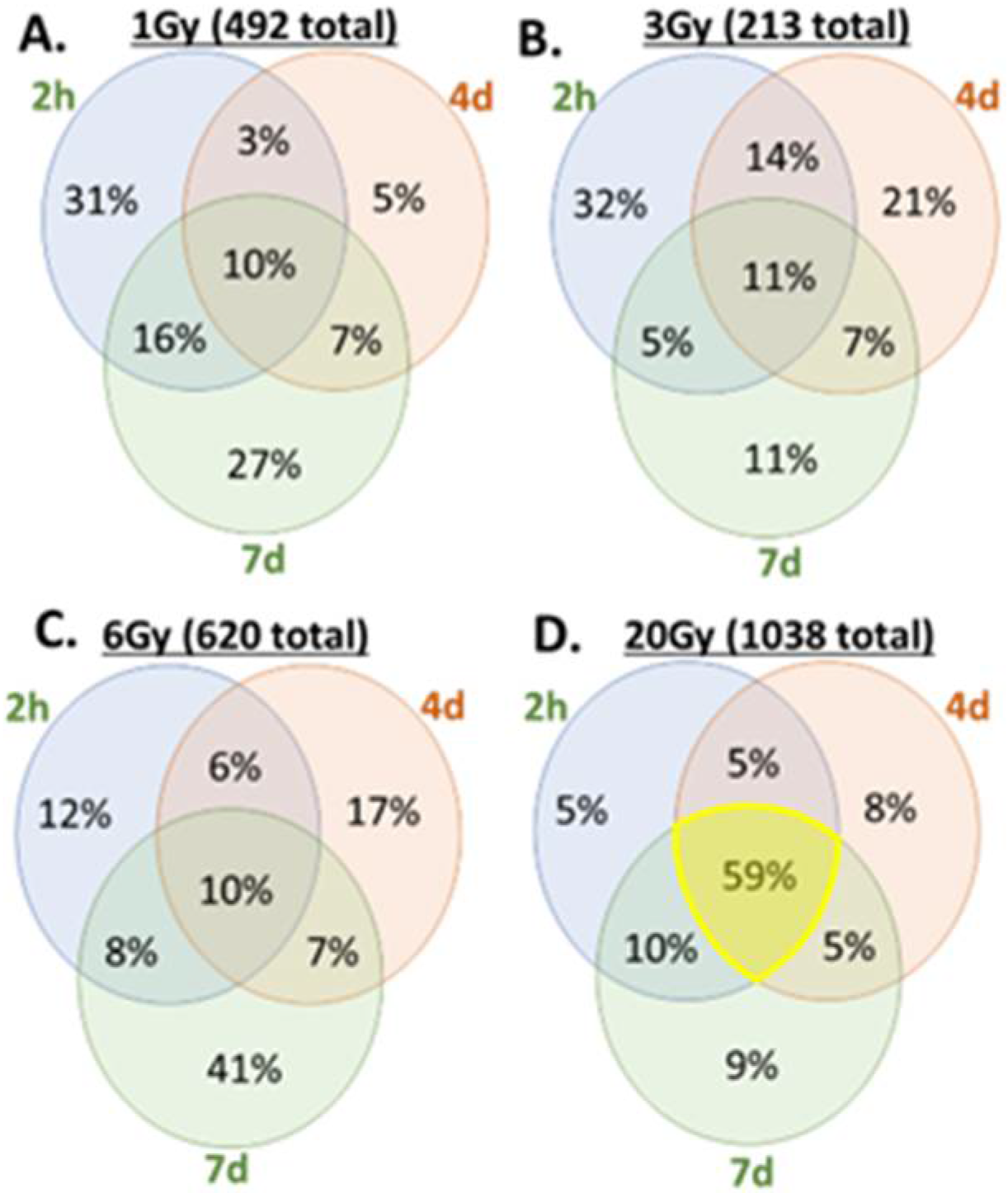
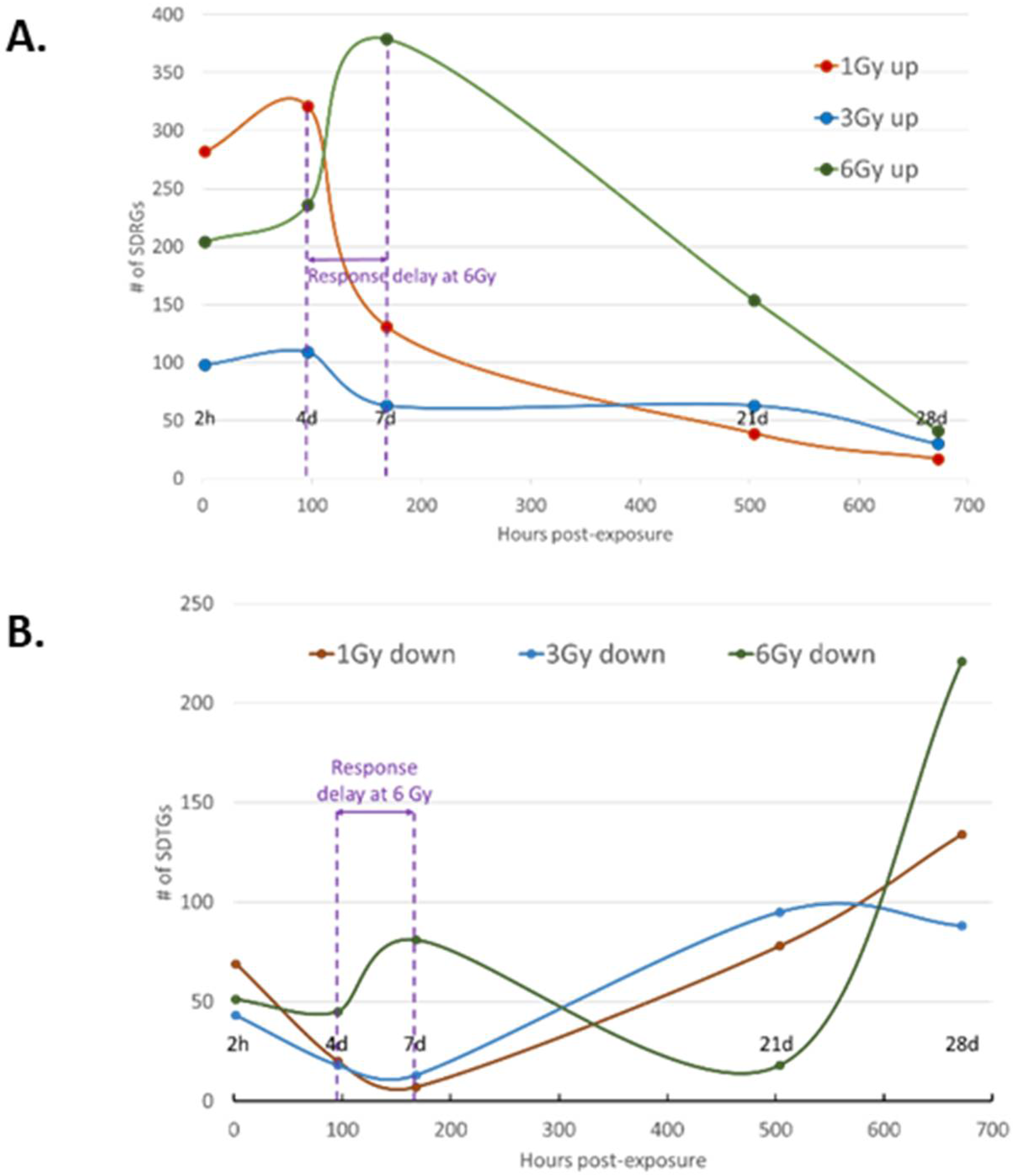
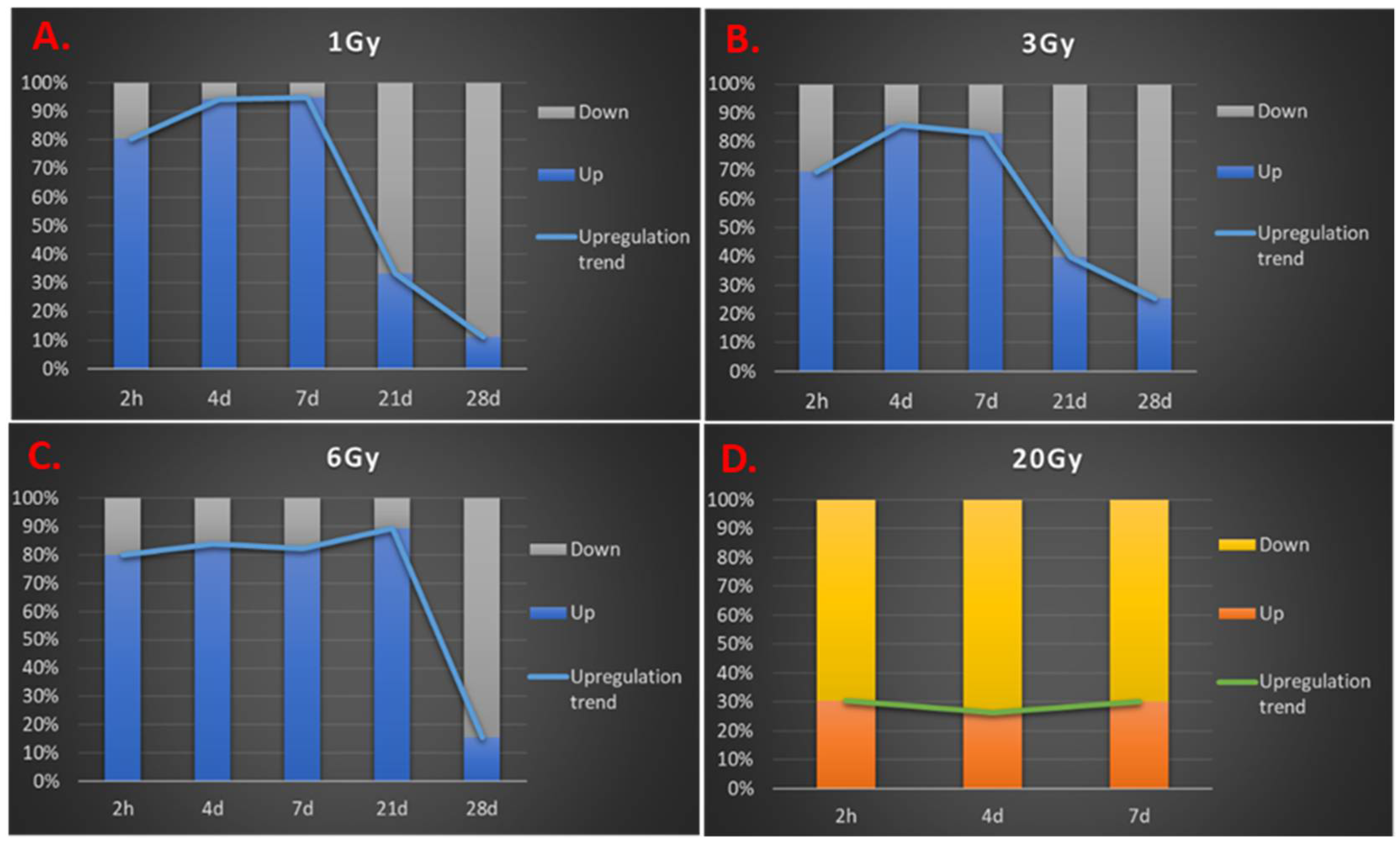
3.3. Transcription Regulation Dynamics and Mode of Gene Regulation Differentiate among Sublethal IR Doses
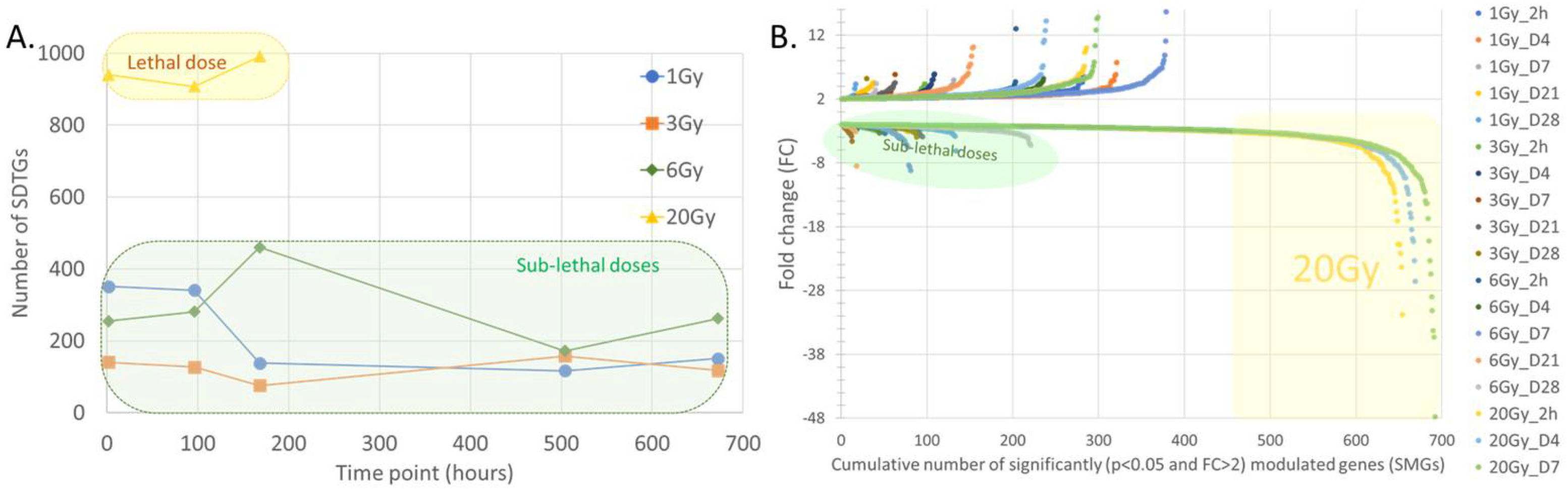
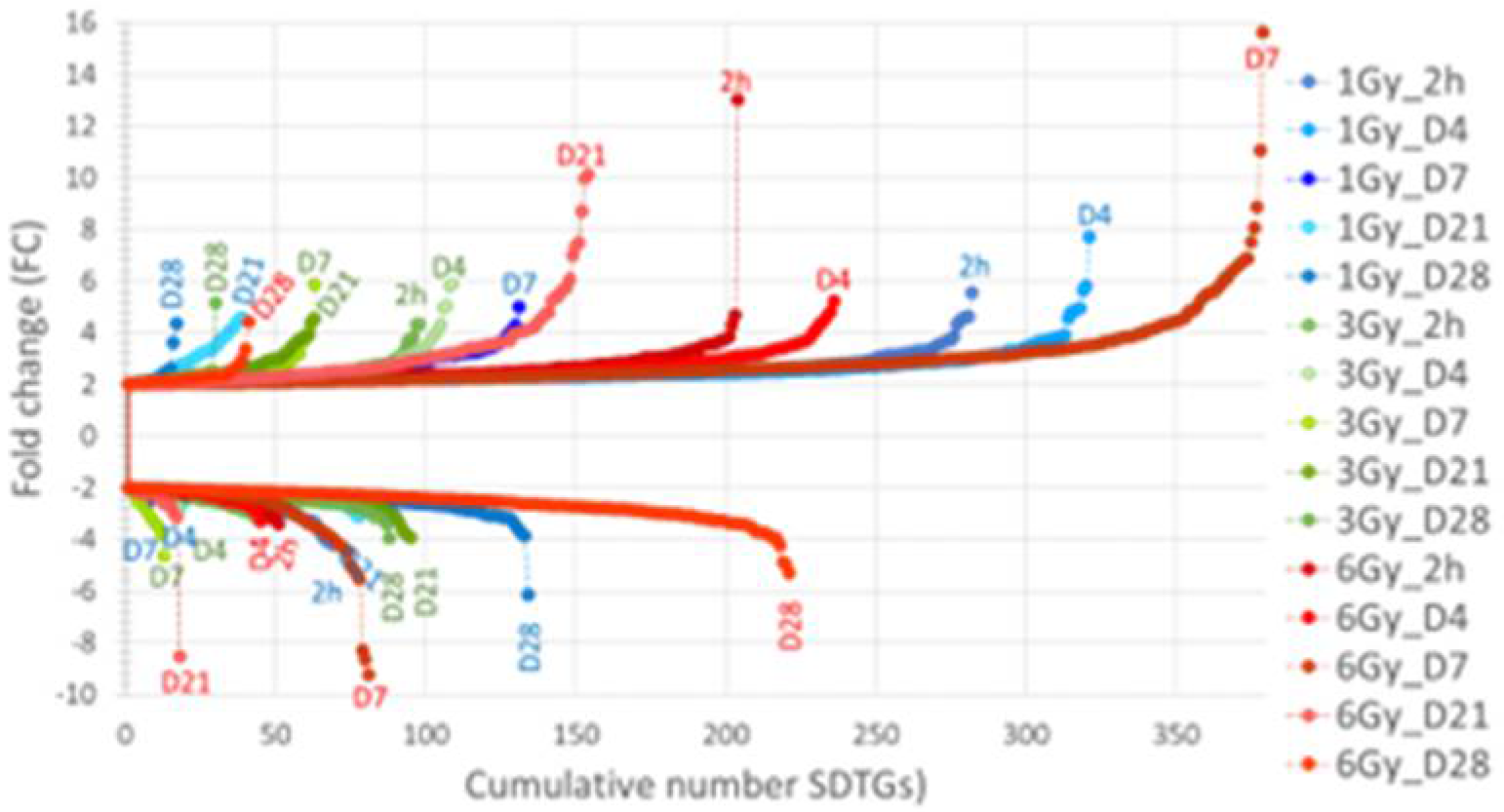
3.4. Distribution of SDTGs through the Time Points and in Different IR Doses Distinguishes Responses in Lethal and Sublethal IR Doses
3.5. Response Dynamics Are More Time-Based Than Dependent on IR Dose in the Sublethal Dose Range
3.6. Specific Significantly Differentially Transcribed Genes Distinguish between Exposure to Lethal and Sublethal IR Doses
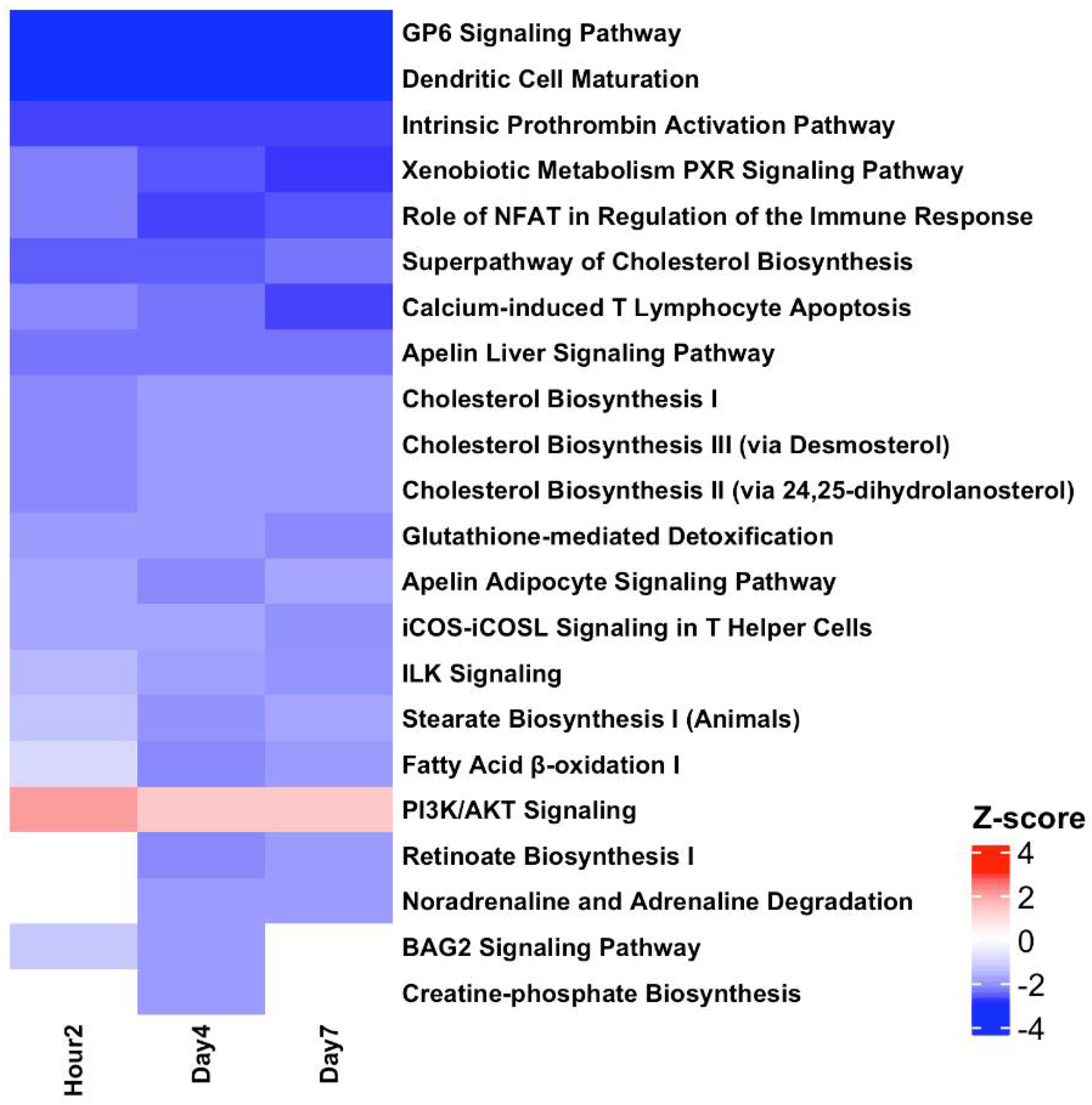
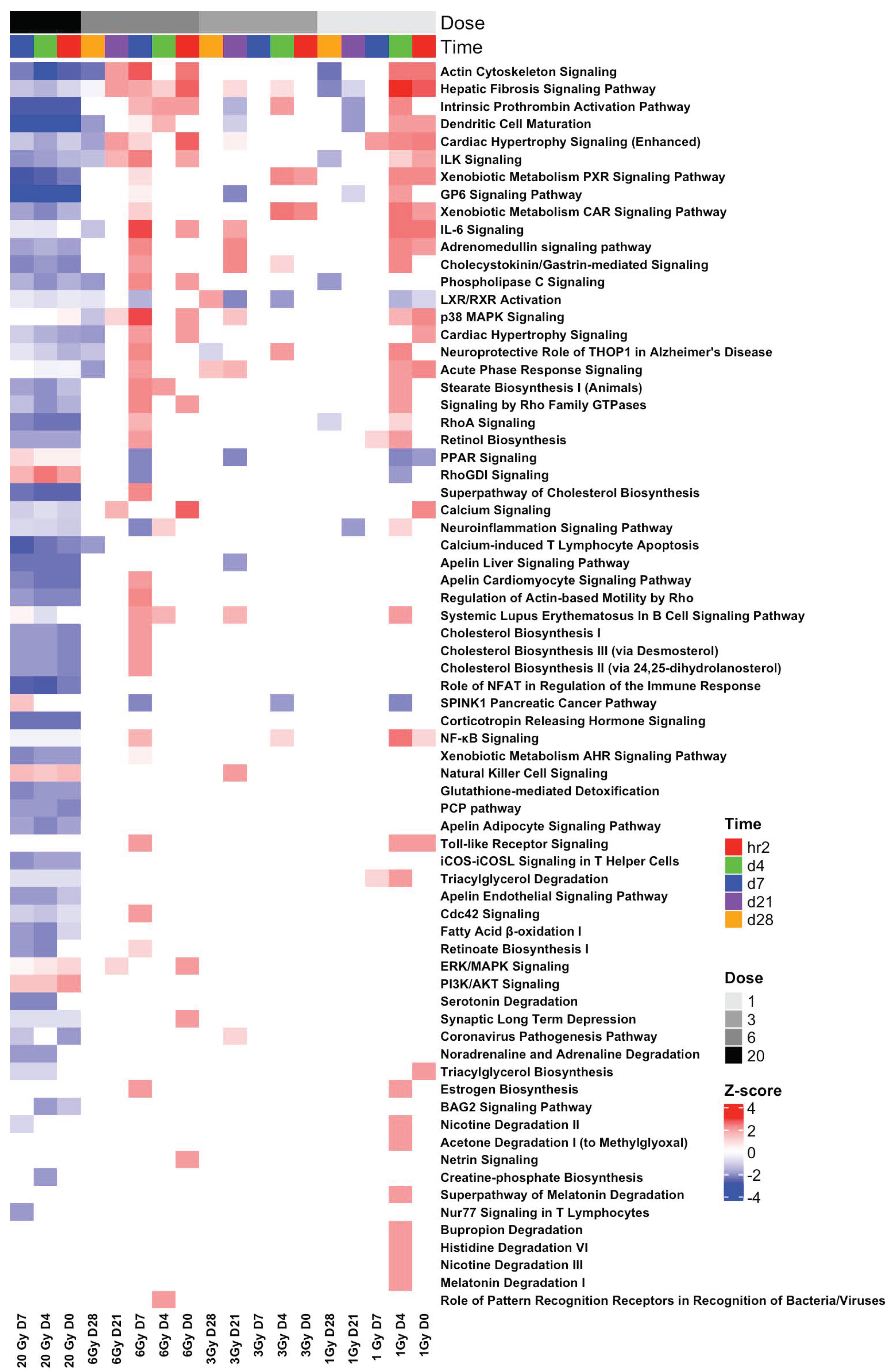
3.7. Pathway Analysis Revealed Inverted Biological Responses in the BLDs and BSDs at All TPs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thomas, G.A.; Symonds, P. Radiation Exposure and Health Effects—Is it Time to Reassess the Real Consequences? Clin. Oncol. 2016, 28, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Merrifield, M.; Kovalchuk, O. Epigenetics in radiation biology: A new research frontier. Front. Genet. 2013, 4, 40. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-F.; Chen, Y.-C.; Hsu, P.W.-C.; Liu, I.Y.; Wu, L.S.-H. MicroRNA expression profiling altered by variant dosage of radiation exposure. BioMed Res. Int. 2014, 2014, 456323. [Google Scholar] [CrossRef]
- Coy, S.L.; Cheema, A.K.; Tyburski, J.B.; Laiakis, E.C.; Collins, S.P.; Fornace, A.J., Jr. Radiation metabolomics and its potential in biodosimetry. Int. J. Radiat. Biol. 2011, 87, 802–823. [Google Scholar] [CrossRef]
- Donnelly, E.H.; Nemhauser, J.B.; Smith, J.M.; Kazzi, Z.N.; Farfán, E.B.; Chang, A.S.; Naeem, S.F. Acute Radiation Syndrome: Assessment and Management. South Med. J. 2010, 103, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Nepper-Christensen, S.; Heslet, L.; Bay, C. Acute radiation syndrome (ARS)—Treatment of the reduced host defense. Int. J. Gen. Med. 2012, 5, 105–115. [Google Scholar] [CrossRef]
- Grosovsky, A.J.; Parks, K.K.; Giver, C.R.; Nelson, S.L. Clonal analysis of delayed karyotypic abnormalities and gene mutations in radia-tion-induced genetic instability. Mol. Cell. Biol. 1996, 16, 6252–6262. [Google Scholar] [CrossRef] [PubMed]
- Roy, L.; Gruel, G.; Vaurijoux, A. Cell response to ionising radiation analysed by gene expression patterns. Ann. Ist. Super. Sanita 2009, 45, 272–277. [Google Scholar]
- Riecke, A.; Rufa, C.G.; Cordes, M.; Hartmann, J.; Meineke, V.; Abend, M. Gene Expression Comparisons Performed for Biodosimetry Purposes on In Vitro Peripheral Blood Cellular Subsets and Irradiated Individuals. Radiat. Res. 2012, 178, 234–243. [Google Scholar] [CrossRef]
- Rivina, L.; Davoren, M.J.; Schiestl, R.H. Mouse models for radiation-induced cancers. Mutagenesis 2016, 31, 491–509. [Google Scholar] [CrossRef]
- Rivina, L.; Schiestl, R. Mouse Models for Efficacy Testing of Agents against Radiation Carcinogenesis—A Literature Review. Int. J. Environ. Res. Public Health 2012, 10, 107–143. [Google Scholar] [CrossRef] [PubMed]
- Seed, T.M.; Inal, C.E.; Singh, V.K. Radioprotection of hematopoietic progenitors by low dose amifostine prophylaxis. Int. J. Radiat. Biol. 2014, 90, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Newman, V.L.; Romaine, P.L.; Hauer-Jensen, M.; Pollard, H.B. Use of biomarkers for assessing radiation injury and efficacy of countermeasures. Expert Rev. Mol. Diagn. 2015, 16, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Swartz, H.M.; Flood, A.B.; Gougelet, R.M.; Rea, M.E.; Nicolalde, R.J.; Williams, B.B. A critical assessment of biodosimetry methods for large-scale incidents. Health Phys. 2010, 98, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, J.; Fu, J.; Wang, J.; Ye, S.; Liu, W.; Shao, C. Role of ROS-mediated autophagy in radiation-induced bystander effect of hepatoma cells. Int. J. Radiat. Biol. 2015, 91, 452–458. [Google Scholar] [CrossRef]
- Singh, V.K.; Fatanmi, O.O.; Singh, P.K.; Whitnall, M.H. Role of radiation-induced granulocyte colony-stimulating factor in recovery from whole body gamma-irradiation. Cytokine 2012, 58, 406–414. [Google Scholar] [CrossRef]
- Singh, V.K.; Ducey, E.J.; Fatanmi, O.O.; Singh, P.K.; Brown, D.S.; Purmal, A.; Shakhova, V.V.; Gudkov, A.V.; Feinstein, E.; Shakhov, A. CBLB613: A TLR 2/6 Agonist, Natural Lipopeptide of Mycoplasma arginini, as a Novel Radiation Countermeasure. Radiat. Res. 2011, 177, 628–642. [Google Scholar] [CrossRef]
- Singh, V.K.; Christensen, J.; Fatanmi, O.O.; Gille, D.; Ducey, E.J.; Wise, S.Y.; Karsunky, H.; Sedello, A.K. Myeloid Progenitors: A Radiation Countermeasure that is Effective when Initiated Days after Irradiation. Radiat. Res. 2012, 177, 781–791. [Google Scholar] [CrossRef]
- Singh, V.K.; Brown, D.S.; Kao, T.C. Alpha-tocopherol succinate protects mice from gamma-radiation by induction of granulo-cyte-colony stimulating factor. Int. J. Radiat. Biol. 2010, 86, 12–21. [Google Scholar] [CrossRef]
- Krivokrysenko, V.I.; Shakhov, A.N.; Singh, V.; Bone, F.; Kononov, Y.; Shyshynova, I.; Cheney, A.; Maitra, R.K.; Purmal, A.; Whitnall, M.H.; et al. Identification of Granulocyte Colony-Stimulating Factor and Interleukin-6 as Candidate Biomarkers of CBLB502 Efficacy as a Medical Radiation Countermeasure. J. Pharmacol. Exp. Ther. 2012, 343, 497–508. [Google Scholar] [CrossRef]
- Ha, C.T.; Li, X.-H.; Fu, D.; Moroni, M.; Fisher, C.; Arnott, R.; Srinivasan, V.; Xiao, M. Circulating Interleukin-18 as a Biomarker of Total-Body Radiation Exposure in Mice, Minipigs, and Nonhuman Primates (NHP). PLoS ONE 2014, 9, e109249. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.K.; Trankle, C.R.; Carbone, S.; Billingsley, H.; Van Tassell, B.W.; Evans, R.K.; Garten, R.; Weiss, E.; Abbate, A. Increased C-reactive protein is associated with the severity of thoracic radiothera-py-induced cardiomyopathy. Cardio-Oncology 2020, 6, 2. [Google Scholar]
- Bertho, J.-M.; Demarquay, C.; Frick, J.; Joubert, C.; Arenales, S.; Jacquet, N.; Sorokine-Durm, I.; Chau, Q.; Lopez, M.; Aigueperse, J.; et al. Level of Flt3-ligand in plasma: A possible new bio-indicator for radiation-induced aplasia. Int. J. Radiat. Biol. 2001, 77, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Sproull, M.; Camphausen, K. State-of-the-Art Advances in Radiation Biodosimetry for Mass Casualty Events Involving Radiation Exposure. Radiat. Res. 2016, 186, 423–435. [Google Scholar] [CrossRef]
- Hu, S.; Blakely, W.F.; Cucinotta, F.A. HEMODOSE: A Biodosimetry Tool Based on Multi-type Blood Cell Counts. Health Phys. 2015, 109, 54–68. [Google Scholar] [CrossRef]
- Wong, K.F.; Siu, L.L.P.; Ainsbury, E.; Moquet, J. Cytogenetic biodosimetry: What it is and how we do it. Hong Kong Med. J. 2013, 19, 168–173. [Google Scholar]
- Haber, J.E.; Thorburn, P.C.; Rogers, D. Meiotic and mitotic behavior of dicentric chromosomes in saccharomyces cerevisiae. Genetics 1984, 106, 185. [Google Scholar] [CrossRef]
- Kravtsov, V.; Fedortseva, R.; Starkova, Y.; Yartseva, N.; Nikiforov, A. Tailed nuclei and dicentric chromosomes in irradiated subjects. Appl. Radiat. Isot. 2000, 52, 1121–1127. [Google Scholar] [CrossRef]
- Singh, V.K.; Santiago, P.T.; Simas, M.; Garcia, M.; Fatanmi, O.O.; Wise, S.Y.; Seed, T.M. Acute radiation syndrome: An update on biomarkers for radiation injury. J. Radiat. Cancer Res. 2018, 9, 132. [Google Scholar] [CrossRef]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. gammaH2AX: A sensitive molecular marker of DNA damage and repair. Leukemia 2010, 24, 679–686. [Google Scholar] [CrossRef]
- Redon, C.E.; Nakamura, A.J.; Martin, O.A.; Parekh, P.R.; Weyemi, U.S.; Bonner, W.M. Recent developments in the use of gamma-H2AX as a quantitative DNA dou-ble-strand break biomarker. Aging 2011, 3, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Ivashkevich, A.; Redon, C.E.; Nakamura, A.J.; Martin, R.F.; Martin, O.A. Use of the gamma-H2AX assay to monitor DNA damage and repair in trans-lational cancer research. Cancer Lett. 2012, 327, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Markova, E.; Torudd, J.; Belyaev, I. Long time persistence of residual 53BP1/gamma-H2AX foci in human lymphocytes in rela-tionship to apoptosis, chromatin condensation and biological dosimetry. Int. J. Radiat. Biol. 2011, 87, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Bourton, E.C.; Plowman, P.N.; Smith, D.; Arlett, C.F.; Parris, C.N. Prolonged expression of the gamma-H2AX DNA repair biomarker correlates with excess acute and chronic toxicity from radiotherapy treatment. Int. J. Cancer 2011, 129, 2928–2934. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.W.; Na, Y.J.; Lee, Y.J.; An, S.; Lee, J.E.; Jung, M.; Kim, H.; Nam, S.Y.; Kim, C.S.; Yang, K.H.; et al. Comprehensive analysis of time- and dose-dependent patterns of gene expression in a human mes-enchymal stem cell line exposed to low-dose ionizing radiation. Oncol. Rep. 2008, 19, 135–144. [Google Scholar] [PubMed]
- Aypar, U.; Morgan, W.F.; Baulch, J.E. Radiation-induced epigenetic alterations after low and high LET irradiations. Mutat. Res. Mol. Mech. Mutagen. 2011, 707, 24–33. [Google Scholar] [CrossRef]
- Aypar, U.; Morgan, W.F.; Baulch, J.E. Radiation-induced genomic instability: Are epigenetic mechanisms the missing link? Int. J. Radiat. Biol. 2011, 87, 179–191. [Google Scholar] [CrossRef]
- Kaup, S.; Grandjean, V.; Mukherjee, R.; Kapoor, A.; Keyes, E.; Seymour, C.B.; Mothersill, C.E.; Schofield, P.N. Radiation-induced genomic instability is associated with DNA methylation changes in cultured human keratinocytes. Mutat. Res. Mol. Mech. Mutagen. 2006, 597, 87–97. [Google Scholar] [CrossRef]
- Pernot, E.; Hall, J.; Baatout, S.; Benotmane, M.A.; Blanchardon, E.; Bouffler, S.; El Saghire, H.; Gomolka, M.; Guertler, A.; Harms-Ringdahl, M.; et al. Ionizing radiation biomarkers for potential use in epidemiological studies. Mutat. Res. Rev. Mutat. Res. 2012, 751, 258–286. [Google Scholar] [CrossRef]
- Pannkuk, E.L.; Fornace, A.J., Jr.; Laiakis, E.C. Metabolomic applications in radiation biodosimetry: Exploring radiation effects through small molecules. Int. J. Radiat. Biol. 2017, 93, 1151–1176. [Google Scholar] [CrossRef]
- Pannkuk, E.L.; Laiakis, E.C.; Authier, S.; Wong, K.; Fornace, A.J. Global Metabolomic Identification of Long-Term Dose-Dependent Urinary Biomarkers in Nonhuman Primates Exposed to Ionizing Radiation. Radiat. Res. 2015, 184, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Laiakis, E.C. Metabolomic Applications in Radiation Biodosimetry. Methods Mol. Biol. 2019, 1978, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Hyduke, D.R.; Laiakis, E.C.; Li, H.H.; Fornace, A.J., Jr. Identifying radiation exposure biomarkers from mouse blood transcriptome. Int. J. Bioinform. Res. Appl. 2013, 9, 365–385. [Google Scholar] [CrossRef] [PubMed]
- Almond, P.R.; Biggs, P.J.; Coursey, B.M.; Hanson, W.F.; Huq, M.S.; Nath, R.; Rogers, D.W. AAPM’s TG-51 protocol for clinical reference dosimetry of high-energy photon and electron beams. Med. Phys. 1999, 26, 1847–1870. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Belinda, P.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IR Dose | Union in All TPs | Common to All TPs | Number of TP-Unique Molecules | ||||
|---|---|---|---|---|---|---|---|
| 2 h | D4 | D7 | D21 | D28 | |||
| 1 Gy | 1665 | 0 | 37 | 16 | 19 | 8 | 5 |
| 2.22% | 0.96% | 1.14% | 0.48% | 0.3% | |||
| 3 Gy | 4 | 6 | 2 | 28 | 32 | ||
| 0.24% | 0.36% | 0.12% | 1.68% | 1.92% | |||
| 6 Gy | 4 | 43 | 55 | 35 | 18 | ||
| 0.24% | 2.58% | 3.3% | 2.1% | 1.08% | |||
| 20 Gy | 34 | 50 | 58 | NA | NA | ||
| 2.04% | 3% | 3.48% | |||||
| IR Dose | Union in All TPs | Common to All TPs | Number of TP-Unique Molecules | ||||
|---|---|---|---|---|---|---|---|
| 2 h | d4 | d7 | d21 | d28 | |||
| 1 Gy | 619 | 1 | 125 | 101 | 26 | 51 | 62 |
| 0.16% | 20.2% | 16.3% | 4.2% | 8.2% | 10% | ||
| 3 Gy | 411 | 0 | 61 | 35 | 20 | 99 | 82 |
| 14.8% | 8.5% | 4.9% | 24.1% | 20% | |||
| 6 Gy | 765 | 4 | 47 | 84 | 173 | 52 | 88 |
| 0.52% | 6.1% | 11.0% | 22.6% | 6.8% | 11.5% | ||
| 20 Gy | 1038 | 609 | 57 | 80 | 89 | NA | NA |
| 58.6% | 5.5% | 7.7% | 8.6% | ||||
| IR Dose | Time Point | Symbol | Upregulated | Downregulated | |||
|---|---|---|---|---|---|---|---|
| Gene Name | Expr FC | Symbol | Gene Name | Expr FC | |||
| 1 Gy | 2 h | TRIM63 | tripartite motif containing 63 | 4.607 | KRT31 | keratin 31 | −3.736 |
| MYLK4 | myosin light chain kinase family member 4 | 4.488 | |||||
| MSTN | Myostatin | 3.841 | |||||
| d4 | Stfa2/Stfa2l1 | stefin A2 | 5.821 | no genes < −3.5 | |||
| S100A9 | S100 calcium-binding protein A9 | 3.711 | |||||
| d7 | D030036P13Rik | RIKEN cDNA D030036P13 gene | 5.008 | no genes < −3.5 | |||
| 4930556J24Rik | RIKEN cDNA 4930556J24 gene | 4.293 | |||||
| MKNK1 | MAPK interacting serine/threonine kinase 1 | 4.078 | |||||
| NET1 | neuroepithelial cell transforming 1 | 4.051 | |||||
| LIN7A | lin-7 homolog A, crumbs cell polarity complex component | 3.728 | |||||
| d21 | PRR9 | proline rich 9 | 4.542 | ||||
| d28 | Kap | kidney androgen-regulated protein | 4.359 | PPP1R3C | protein phosphatase 1 regulatory subunit 3C | −3.856 | |
| KRT71 | keratin 71 | −6.13 | |||||
| 3 Gy | 2 h | Defb8 | defensin beta 8 | 4.342 | no genes | ||
| d7 | PON1 | paraoxonase 1 | 3.654 | no genes | |||
| d21 | NR4A1 | nuclear receptor subfamily 4 group A member 1 | 4.566 | MLANA | melan-A | −3.711 | |
| FOSB | FosB proto-oncogene, AP-1 transcription factor subunit | 3.909 | |||||
| FOS | Fos proto-oncogene, AP-1 transcription factor subunit | 3.734 | LGR5 | leucine-rich repeat containing G protein-coupled receptor 5 | −3.862 | ||
| CCL4 | C-C motif chemokine ligand 4 | 3.557 | |||||
| d28 | GJB2 | gap junction protein beta 2 | 5.18 | PPP1R3C | protein phosphatase 1 regulatory subunit 3C | −3.942 | |
| 6 Gy | d4 | PLAC8 | placenta-associated 8 | 5.227 | no genes < −3.5 | ||
| CCL2 | C-C motif chemokine ligand 2 | 4.079 | |||||
| FCGR1A | Fc fragment of IgG receptor Ia | 3.649 | |||||
| d7 | Stfa2/Stfa2l1 | stefin A2 | 11.053 | Krtap16-3 | keratin-associated protein 16-3 | −3.576 | |
| 1110025L11Rik | RIKEN cDNA 1110025L11 gene | −3.597 | |||||
| KLHL36 | kelch-like family member 36 | 5.555 | KRT34 | keratin 34 | −3.734 | ||
| FBP1 | fructose-bisphosphatase 1 | −3.755 | |||||
| CKMT2 | creatine kinase, mitochondrial 2 | 4.435 | Krtap8-1 | keratin-associated protein 8-1 | −3.864 | ||
| Krtap22-2 | keratin-associated protein 22-2 | −3.938 | |||||
| KLK6 | kallikrein-related peptidase 6 | 4.3 | Gm10229 | predicted gene 10229 | −4.033 | ||
| PRR9 | proline rich 9 | −4.416 | |||||
| KRT6B | keratin 6B | 3.847 | Krtap19-1 | keratin-associated protein 19-1 | −5.359 | ||
| KRT27 | keratin 27 | −5.532 | |||||
| KRT25 | keratin 25 | −8.664 | |||||
| d21 | CHRNA1 | cholinergic receptor nicotinic alpha 1 subunit | 5.57 | no genes < −3.5 | |||
| FOS | Fos proto-oncogene, AP-1 transcription factor subunit | 5.228 | |||||
| CHRNG | cholinergic receptor nicotinic gamma subunit | 4.627 | |||||
| HSPB7 | heat shock protein family B (small) member 7 | 4.321 | |||||
| ANKRD1 | ankyrin repeat domain 1 | 4.161 | |||||
| MYH3 | myosin heavy chain 3 | 3.54 | |||||
| d28 | no genes > 3.5 | Fam25c | family with sequence similarity 25, member C | −3.683 | |||
| Krt10 | keratin 10 | −3.758 | |||||
| PPP1R3C | protein phosphatase 1 regulatory subunit 3C | −3.964 | |||||
| 20 Gy | 2 h | no genes > 3.5 | KRT28 | keratin 28 | −3.725 | ||
| d4 | ENO3 | enolase 3 | −3.848 | ||||
| PPP1R3C | protein phosphatase 1 regulatory subunit 3C | −4.105 | |||||
| Time Point | Union | # of Common SDTGs | # of IR Dose-Unique SDTGs | |||
|---|---|---|---|---|---|---|
| 1 Gy | 3 Gy | 6 Gy | 20 Gy | |||
| 2 h | 1096 | 17 1.55% | 83 | 14 | 36 | 702 |
| 7.57% | 1.27% | 3.28% | 64.05% | |||
| d4 | 1079 | 24 2.22% | 78 | 15 | 107 | 620 |
| 7.22% | 1.39% | 9.91% | 57.46% | |||
| d7 | 1121 | 16 1.42% | 34 | 7 | 179 | 666 |
| 3.03% | 0.62% | 15.96% | 59.41% | |||
| d21 | 314 | 5 | 47 | 76 | 118 | NA |
| −1.47% | 14.96% | 24.20% | 37.57% | |||
| d28 | 322 | 19 | 21 | 66 | 112 | NA |
| 5.90% | 6.52% | 20.49% | 34.78% | |||
| Time Point | Symbol | Molecule Name | Expr Fold Change | Location | Type(s) | |||
|---|---|---|---|---|---|---|---|---|
| (1 Gy) | (3 Gy) | (6 Gy) | (20 Gy) | |||||
| 2 h | AADAC * | arylacetamide deacetylase | 2.41 | 2.86 | 2.89 | −7.09 | Cytoplasm | enzyme |
| ABRA | actin-binding Rho activating protein | 5.57 | 2.69 | 2.93 | 4.37 | Cytoplasm | other | |
| ACOXL | acyl-CoA oxidase-like | 2.57 | 2.75 | 3.08 | −3.92 | Other | enzyme | |
| ELOVL4 * | ELOVL fatty acid elongase 4 | 2.09 | 2.10 | 2.32 | −7.36 | Cytoplasm | enzyme | |
| ELOVL6 | ELOVL fatty acid elongase 6 | 2.03 | 2.20 | 2.34 | −6.34 | Cytoplasm | enzyme | |
| FA2H | fatty acid 2-hydroxylase | 2.34 | 2.32 | 2.38 | −20.85 | Cytoplasm | enzyme | |
| MYH2 | myosin heavy chain 2 | 4.55 | 2.74 | 4.69 | −3.49 | Cytoplasm | enzyme | |
| SEC14L4 * | SEC14-like lipid binding 4 | 2.58 | 2.38 | 2.72 | −6.28 | Other | transporter | |
| Wfdc3 | WAP four-disulfide core domain 3 | 2.30 | 2.23 | 2.78 | −9.16 | Other | other | |
| d4 | ACOXL | acyl-CoA oxidase-like | 3.11 | 2.12 | 3.11 | −4.04 | Other | enzyme |
| TMPRSS4 | transmembrane serine protease 4 | 4.94 | 3.07 | 2.05 | −2.69 | Cytoplasm | peptidase | |
| Serpina3b/Serpina3j | serine (or cysteine) peptidase inhibitor, clade A (alpha-1 antiproteinase, antitrypsin), member 3J | 3.53 | 2.22 | 2.41 | −4.66 | Extracellular Space | other | |
| HBB | hemoglobin subunit beta | 2.31 | −2.12 | −2.83 | −6.06 | Cytoplasm | transporter | |
| ELOVL6 | ELOVL fatty acid elongase 6 | 2.83 | 2.04 | 3.22 | −5.49 | Cytoplasm | enzyme | |
| ELOVL3 | ELOVL fatty acid elongase 3 | 2.72 | 2.06 | 2.90 | −6.43 | Cytoplasm | enzyme | |
| SEC14L4 * | SEC14-like lipid binding 4 | 3.46 | 2.18 | 2.88 | −6.46 | Other | transporter | |
| ELOVL4 * | ELOVL fatty acid elongase 4 | 3.48 | 2.57 | 3.05 | −5.93 | Cytoplasm | enzyme | |
| AADAC * | arylacetamide deacetylase | 3.89 | 2.49 | 2.43 | −7.44 | Cytoplasm | enzyme | |
| Wfdc3 | WAP four-disulfide core domain 3 | 2.91 | 2.25 | 2.73 | −8.40 | Other | other | |
| RNASE2 | ribonuclease A family member 2 | 2.93 | 4.32 | 4.32 | −10.35 | Cytoplasm | enzyme | |
| S100A9 | S100 calcium-binding protein A9 | 3.71 | 5.80 | 3.00 | 11.25 | Cytoplasm | other | |
| FA2H | fatty acid 2-hydroxylase | 3.78 | 2.29 | 3.45 | −16.57 | Cytoplasm | enzyme | |
| d7 | RNASE2 | ribonuclease A family member 2 | 3.23 | 3.20 | 6.71 | −12.54 | Cytoplasm | enzyme |
| AADAC * | arylacetamide deacetylase | 2.86 | 2.31 | 6.31 | −8.54 | Cytoplasm | enzyme | |
| SEC14L4 * | SEC14-like lipid binding 4 | 3.11 | 2.17 | 6.51 | −6.50 | Other | transporter | |
| Aldh3b2 | aldehyde dehydrogenase 3 family, member B2 | 2.06 | 2.19 | 4.40 | −9.45 | Cytoplasm | enzyme | |
| ELOVL4 * | ELOVL fatty acid elongase 4 | 2.73 | 2.32 | 5.50 | −6.79 | Cytoplasm | enzyme | |
| AQP3 | aquaporin 3 (Gill blood group) | 2.84 | 2.82 | 6.87 | −2.84 | Plasma Membrane | transporter | |
| Defb6 | defensin beta 6 | 2.47 | 2.00 | 5.36 | −5.33 | Extracellular Space | other | |
| Sdr16c6 | short chain dehydrogenase/reductase family 16C, member 6 | 2.59 | 2.58 | 4.89 | −3.77 | Other | other | |
| SLURP1 | secreted LY6/PLAUR domain containing 1 | 3.01 | 2.31 | 4.33 | −3.29 | Extracellular Space | cytokine | |
| Wfdc21 | WAP four-disulfide core domain 21 | 2.67 | 2.24 | 4.43 | −2.70 | Extracellular Space | other | |
| d21 | FLNC | filamin C | 2.81 | 2.95 | 5.74 | NA | Cytoplasm | other |
| d28 | PPP1R3C | protein phosphatase 1 regulatory subunit 3C | −3.86 | −3.94 | −3.96 | NA | Cytoplasm | phosphatase |
| Mup1 (includes others) | major urinary protein 1 | −3.11 | −3.44 | −3.81 | NA | Extracellular Space | other | |
| Kap | kidney androgen-regulated protein | 4.36 | 2.50 | 2.96 | NA | Extracellular Space | other | |
| Union (ALL 3 TPs) | Common SDTGs (ALL 3 TPs) | Time Point | Union | TP-Common SDTGs | IR Dose-Unique SDTGs | |||
|---|---|---|---|---|---|---|---|---|
| 1Gy | 3Gy | 6Gy | 20Gy | |||||
| 1499 | 5 | 2h | 1096 | 17 1.55% | 83 | 14 | 36 | 702 |
| 7.57% | 1.27% | 3.28% | 64.05% | |||||
| d4 | 1079 | 24 2.22% | 78 | 15 | 107 | 620 | ||
| 7.22% | 1.39% | 9.91% | 57.46% | |||||
| d7 | 1121 | 16 1.42% | 34 | 7 | 179 | 666 | ||
| 3.03% | 0.62% | 15.96% | 59.41% | |||||
| Symbol | Entrez Gene Name | 1 Gy-2 h | 1 Gy-d4 | 1 Gy-d7 | 3 Gy-2 h | 3 Gy-d4 | 3 Gy-d7 | 6 Gy-2 h | 6 Gy-d4 | 6 Gy-d7 | 20 Gy-2 h | 20 Gy-d4 | 20 Gy-d7 | Location | Function |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AADAC | arylacetamide deacetylase | 2.41 | 3.89 | 2.86 | 2.86 | 2.49 | 2.31 | 2.89 | 2.43 | 6.31 | −7.09 | −7.44 | −8.54 | Cytoplasm | enzyme |
| ELOVL4 | ELOVL fatty acid elongase 4 | 2.09 | 3.48 | 2.73 | 2.10 | 2.57 | 2.32 | 2.32 | 3.05 | 5.50 | −7.36 | −5.93 | −6.79 | Cytoplasm | enzyme |
| SEC14L4 | SEC14-like lipid binding 4 | 2.58 | 3.46 | 3.11 | 2.38 | 2.18 | 2.17 | 2.72 | 2.88 | 6.51 | −6.28 | −6.46 | −6.50 | Other | transporter |
| SLURP1 | secreted LY6/PLAUR domain containing 1 | 2.29 | 3.33 | 3.01 | 2.07 | 3.10 | 2.31 | 2.21 | 2.95 | 4.33 | −3.08 | −2.52 | −3.29 | Extracellular Space | cytokine |
| TMPRSS4 | transmembrane serine protease 4 | 3.01 | 4.94 | 2.39 | 3.30 | 3.07 | 3.11 | 2.08 | 2.05 | 3.35 | −2.67 | −2.69 | −2.77 | Cytoplasm | peptidase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkhalil, A.; Clifford, J.; Miller, S.M.; Gautam, A.; Jett, M.; Hammamieh, R.; Moffatt, L.T.; Shupp, J.W. Transcriptomes of Wet Skin Biopsies Predict Outcomes after Ionizing Radiation Exposure with Potential Dosimetric Applications in a Mouse Model. Curr. Issues Mol. Biol. 2022, 44, 3711-3734. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb44080254
Alkhalil A, Clifford J, Miller SM, Gautam A, Jett M, Hammamieh R, Moffatt LT, Shupp JW. Transcriptomes of Wet Skin Biopsies Predict Outcomes after Ionizing Radiation Exposure with Potential Dosimetric Applications in a Mouse Model. Current Issues in Molecular Biology. 2022; 44(8):3711-3734. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb44080254
Chicago/Turabian StyleAlkhalil, Abdulnaser, John Clifford, Stacyann M. Miller, Aarti Gautam, Marti Jett, Rasha Hammamieh, Lauren T. Moffatt, and Jeffrey W. Shupp. 2022. "Transcriptomes of Wet Skin Biopsies Predict Outcomes after Ionizing Radiation Exposure with Potential Dosimetric Applications in a Mouse Model" Current Issues in Molecular Biology 44, no. 8: 3711-3734. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb44080254