
Physiological and Molecular Response of Prorocentrum minimum to Tannic Acid: An Experimental Study to Evaluate the Feasibility of Using Tannic Acid in Controling the Red Tide in a Eutrophic Coastal Water

Abstract
:1. Introduction
2. Materials and Methods
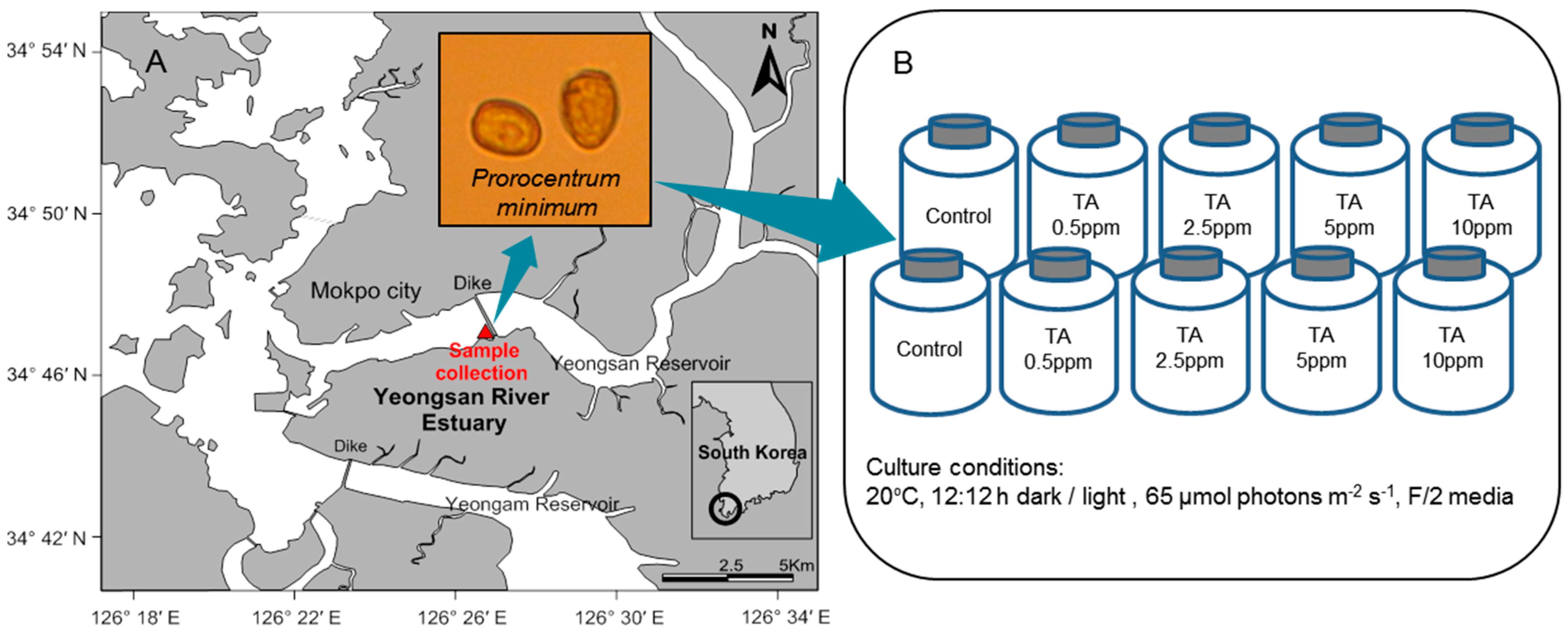
2.1. Sample Collection and Culture
2.2. Bioassay Experiment
2.3. Gene Expression Analysis
2.4. Statistical Analysis
3. Results
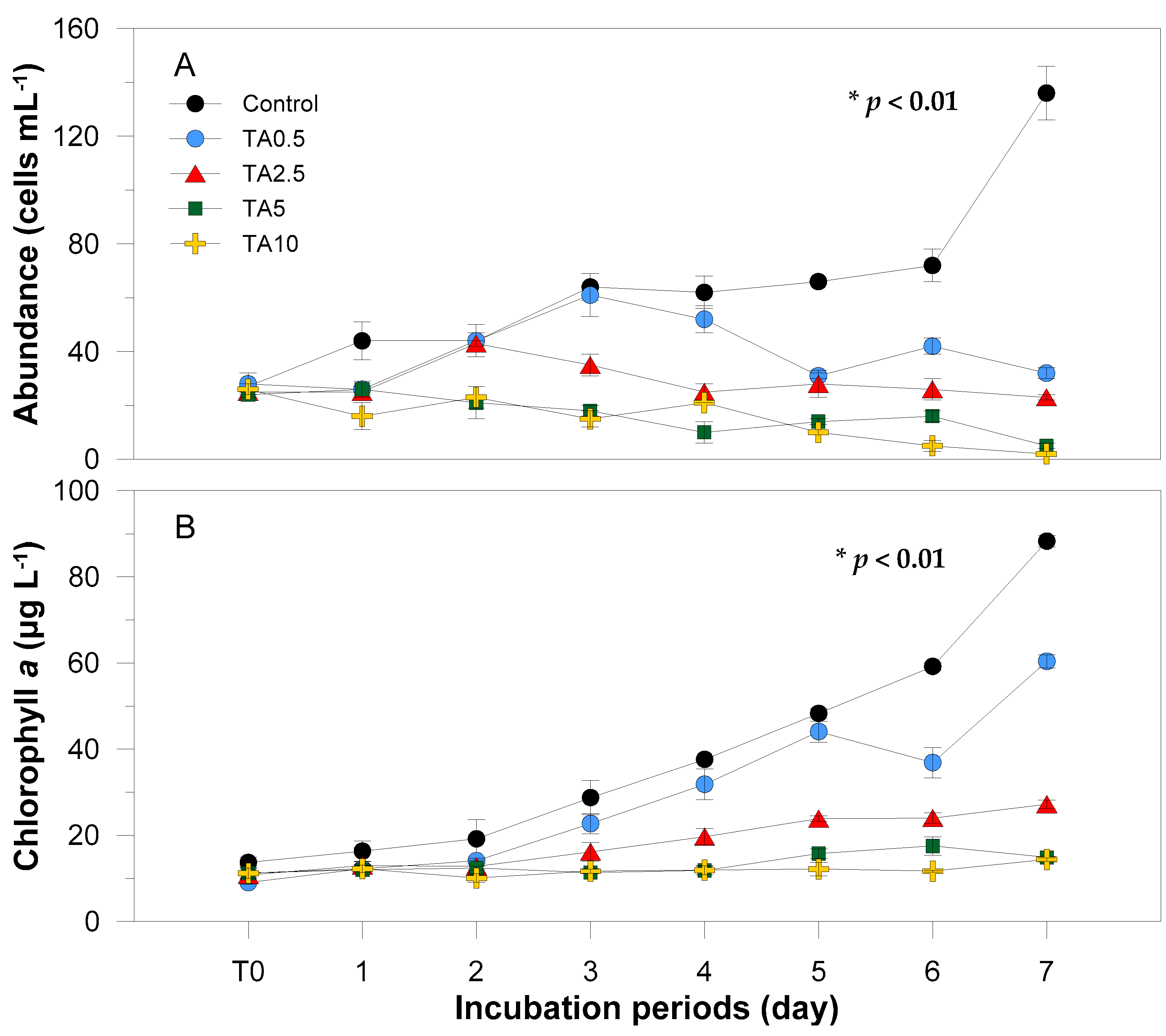
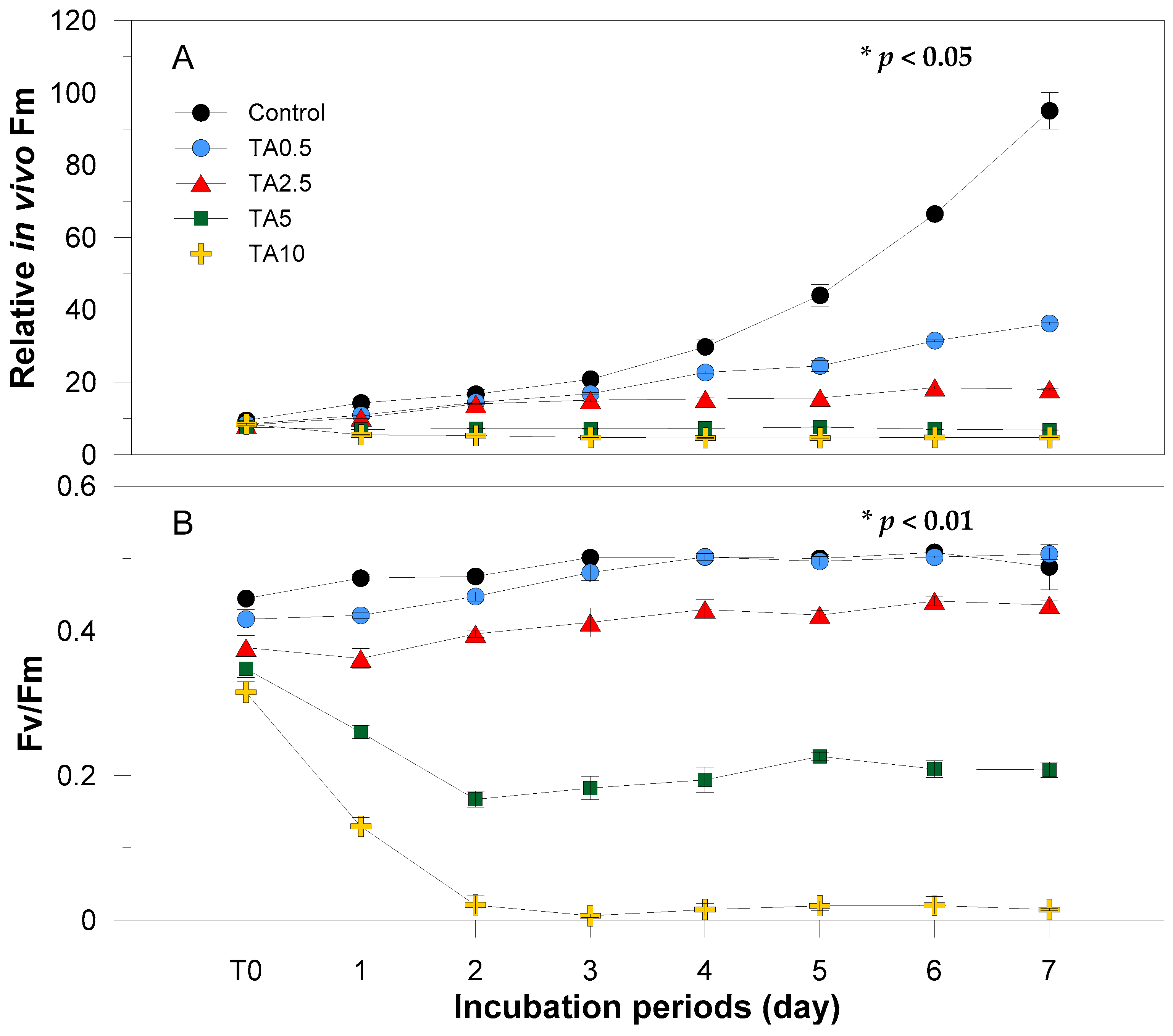
3.1. Physiological Response to Tannic Acid
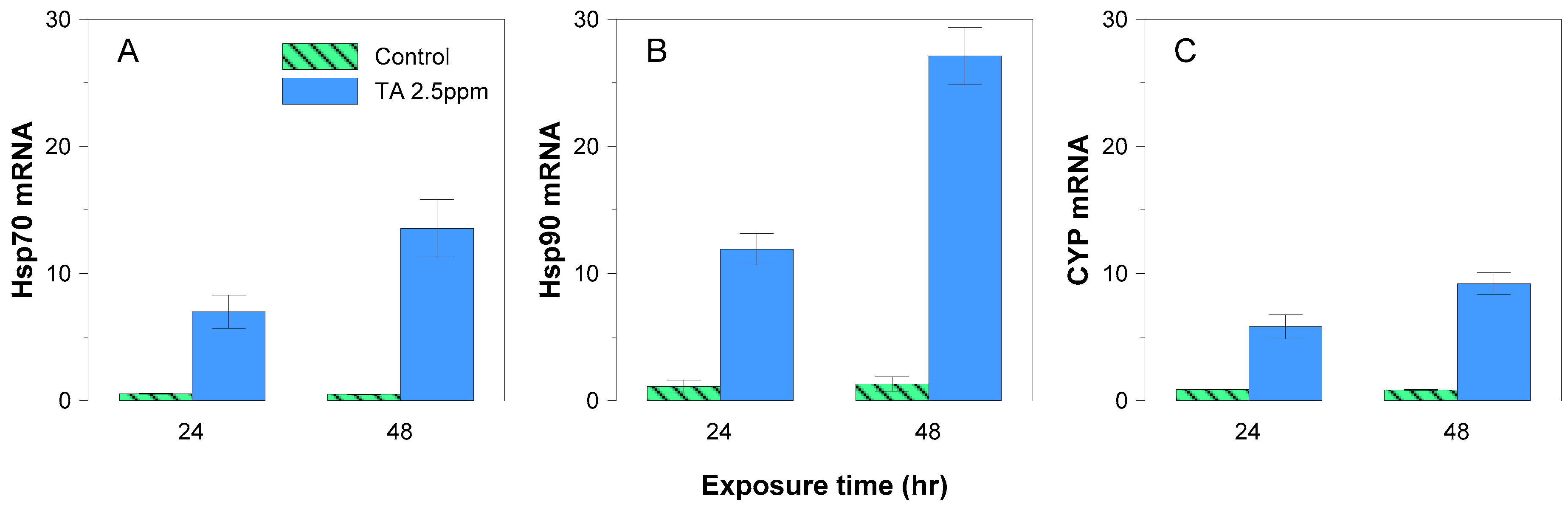
3.2. mRNA Expression
4. Discussion
4.1. Effects of Tannic Acid on Physiological Characteristics
4.2. Effect of Tannic Acid on Gene Expression
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef]
- Anderson, D.M. Turning back the harmful red tide. Nature 1997, 388, 513–514. [Google Scholar] [CrossRef]
- Smayda, T.J. What is a bloom? A commentary. Limnol. Oceanogr. 1997, 42, 1132–1136. [Google Scholar] [CrossRef]
- Lee, C.K.; Park, T.G.; Park, Y.T.; Lim, W.A. Monitoring and trends in harmful algal blooms and red tides in Korean coastal waters, with emphasis on Cochlodinium polykrikoides. Harmful Algae 2013, 30, 3–14. [Google Scholar] [CrossRef]
- Habas, E.J.; Gilbert, C.K. The economic effects of the 1971 Florida red tide and the damage it presages for future occurrences. Environ. Lett. 1974, 6, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Horner, R.A.; Garrison, D.L.; Plumley, F.G. Harmful algal blooms and red tide problems on the US west coast. Limnol. Oceanogr. 1997, 42, 1076–1088. [Google Scholar] [CrossRef]
- Asakawa, M.; Miyazawa, K.; Noguchi, T. Studies on Paralytic Shellfish Poison (PSP) Toxification of Bivalves, in Association with Appearance of Alexandrium tamarense, in Hiroshima Bay, Hiroshima Prefecture. J. Food Hyg. Soc. Jpn. 1992, 34, 50–54. [Google Scholar] [CrossRef]
- Asakawa, M.; Miyazawa, K.; Takayama, H.; Noguchi, T. Dinoflagellate Alexandrium tamarense as the source of Paralytic Shellfish Poison (PSP) contained in bivalves from Hiroshima Bay, Hiroshima Prefecture, Japan. Toxicon 1995, 33, 691–697. [Google Scholar] [CrossRef]
- Park, T.G.; Lim, W.A.; Park, Y.T.; Lee, C.K.; Jeong, H.J. Economic impact, management and mitigation of red tides in Korea. Harmful Algae 2013, 30, 131–143. [Google Scholar] [CrossRef]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Heil, C.A.; Glibert, P.M.; Fan, C. Prorocentrum minimum (Pavillard) Schiller A review of a harmful algal bloom species of growing worldwide importance. Harmful Algae 2005, 4, 449–470. [Google Scholar] [CrossRef]
- Kang, N.S.; Lee, K.H.; Jeong, H.J.; Yoo, Y.D.; Seong, K.A.; Potvin, É.; Hwanga, Y.J.; Yoond, E.Y. Red tides in Shiwha Bay, western Korea: A huge dike and tidal power plant established in a semi-enclosed embayment system. Harmful Algae 2013, 30, 114–130. [Google Scholar] [CrossRef]
- Kondo, K.; Seike, Y.; Date, Y. Red tides in the brackish Lake Nakanoumi. (II). Relationships between the occurrence of Prorocentrum minimum red tide and environmental conditions. Bull. Plankton Soc. Jpn. 1990, 37, 19–34. [Google Scholar]
- Smayda, T.J. Novel and nuisance phytoplankton blooms in the sea: Evidence for a global epidemic. In Proceedings of the Fourth International Conference on Toxic Marine Phytoplankton, Lund, Sweden, 26–30 June 1989; Graneli, E., Sundström, B., Edler, L., Anderson, D.M., Eds.; Elsevier: New York, NY, USA, 1990; pp. 29–40. [Google Scholar]
- Moncheva, S.; Petrova-Karadjova, V.; Palasov, A. Harmful algal blooms along the Bulgarian Black Sea coast and possible patterns of fish and zoobenthic mortalities. In Harmful Marine Algal Blooms, Proceedings of the Sixth International Conference on Toxic Marine Phytoplankton, Nantes, France, 6 October 1993; Lavoisier Publishing: Paris, France, 1995; pp. 193–198. [Google Scholar]
- Gallegos, C.L.; Jordan, T.E. Impact of the spring 2000 phytoplankton bloom in Chesapeake Bay on optical properties and light penetration in the Rhode River, Maryland. Estuaries 2002, 25, 508–518. [Google Scholar] [CrossRef]
- Gallegos, C.L.; Bergstrom, P.W. Effects of a Prorocentrum minimum bloom on light availability for and potential impacts on submersed aquatic vegetation in upper Chesapeake Bay. Harmful Algae 2005, 4, 553–574. [Google Scholar] [CrossRef]
- Tango, P.; Magnien, R.; Butler, W.; Luckett, C.; Luckenbach, M.; Lacouture, R.; Poukish, C. Impacts and potential effects due to Prorocentrum minimum blooms in Chesapeake Bay. Harmful Algae 2005, 4, 525–531. [Google Scholar] [CrossRef]
- Glibert, P.M.; Alexander, J.; Meritt, D.W.; North, E.W.; Stoecker, D.K. Harmful algae pose additional challenges for oyster restoration: impacts of the harmful algae Karlodinium veneficum and Prorocentrum minimum on early life stages of the oysters Crassostrea virginica and Crassostrea ariakensis. J. Shellfish Res. 2007, 2, 919–925. [Google Scholar] [CrossRef]
- Azanza, R.V.; Fukuyo, Y.; Yap, L.G.; Takayama, H. Prorocentrum minimum bloom and its possible link to a massive fish kill in Bolinao, Pangasinan, Northern Philippines. Harmful Algae 2005, 4, 519–524. [Google Scholar] [CrossRef]
- Saba, G.K.; Steinberg, D.K.; Bronk, D.A.; Place, A.R. The effects of harmful algal species and food concentration on zooplankton grazer production of dissolved organic matter and inorganic nutrients. Harmful Algae 2011, 10, 291–303. [Google Scholar] [CrossRef]
- Grzebyk, D.; Denardou, A.; Berland, B.; Pouchus, Y.F. Evidence of a new toxin in the red-tide dinoflagellate Prorocentrum minimum. J. Plankton Res 1997, 19, 1111–1124. [Google Scholar] [CrossRef]
- Maruyama, T.; Yamada, R.; Usui, K.; Suzuki, H.; Yoshida, T. The studies on removal of red tide plankton-II. Removal of marine red tide planktons with acid treated clay. Bull. Jpn. Soc. Sci. Fish. 1987, 53, 1811–1819. [Google Scholar] [CrossRef]
- Yu, Z.; Zou, J.; Ma, X. Application of clays to removal of red tide organisms I. Coagulation of red tide organisms with clays. Chin. J. Oceanol. Limnol. 1994, 12, 193–200. [Google Scholar]
- Yu, Z.; Zou, J.; Ma, X. Application of clays to removal of red tide organisms II. Coagulation of different species of red tide organisms with montmorillonite and effect of clay pretreatment. Chin. J. Oceanol. Limnol. 1994, 12, 316–324. [Google Scholar]
- Na, G.; Choi, W.; Chun, Y. A study on red tide control with loess suspension. J. Aquac. 1996, 9, 239–245. [Google Scholar]
- Sengco, M.; Anderson, D. Controlling harmful algal blooms through clay flocculation. J. Eukaryot. Microbiol. 2003, 51, 169–172. [Google Scholar] [CrossRef]
- Wulff, A.; Sundbäck, K.; Nilsson, C.; Carlson, L.; Jönsson, B. Effect of sediment load on the microbenthic community of a shallow-water sandy sediment. Estuaries 1997, 20, 547–558. [Google Scholar] [CrossRef]
- Raloff, J. Taming toxic tides: Can we slay poisonous algal blooms with clay? Sci. News 2002, 162, 344–345. [Google Scholar] [CrossRef]
- Shumway, S.E.; Frank, D.M.; Ewart, L.M. Effect of yellow loess on clearance rate in seven species of benthic, filter-feeding invertebrates. Aquac. Res. 2003, 34, 1391–1402. [Google Scholar] [CrossRef]
- Eigemman, F.; Vanormelingen, P.; Hilt, S. Sensitivity of green alga Pediastrum duplex Meyen to allelochemicals is strain-specific and not related to co-occurrence with allelopathic macrophytes. PLoS ONE 2013, 8, e78463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, N.; Blaschke, U.; Beutler, E.; Gross, E.M.; Jenett-Siems, K.; Siems, K.; Hilt, S. Seasonal and interannual dynamics of polyphenols in Myriophyllum verticillatum and their allelopathic activity on Anabaena variabilis. Aquat. Bot. 2009, 91, 110–116. [Google Scholar] [CrossRef]
- Leu, E.; Krieger-Liszkay, A.; Goussias, C.; Gross, E. Polyphenolic allelochemicals from the Aquatic angiosperm Myriophyllum spicatum inhibit photosystem II. Plant Physiol. 2002, 130, 2011–2018. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jin, L.; Ominski, K.H.; He, M.; Xu, Z.; Krause, D.O.; Acharya, S.N.; Wittenberg, K.M.; Liu, X.L.; Stanford, K.; et al. Screening of condensed tannins from Canadian prairie forages for anti-Escherichia coli O157:H7 with an emphasis on purple prairie clover (Dalea purpurea Vent). J. Food Prot. 2013, 76, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Chia, A.M.; Iortsuun, D.N.; Stephen, B.J.; Ayobamire, A.E.; Ladan, Z. Phytoplankton responses to changes in macrophyte density in a tropical artificial pond in Zaria, Nigeria. Afr. J. Aquat. Sci. 2011, 36, 35–46. [Google Scholar] [CrossRef]
- Bahrs, N.; Laue, P.; Chakrabarti, S.; Steinberg, C. Plant polyphenols: Do they control freshwater planktonic nuisance phototrophs? In Polyphenols in Plants: Isolation, Purification and Extract Purification; Watson, R.R., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 87–96. [Google Scholar]
- Henis, Y.; Tagari, H.; Volcani, R. Effect of water extracts of carob pods, tannic acid, and their derivatives on the morphology and growth of microorganisms. J. Appl. Microbiol. 1964, 12, 204–209. [Google Scholar]
- Field, J.A.; Lettinga, G. Toxicity of tannic compounds to microorganisms. Basic Life Sci. 1992, 59, 673–692. [Google Scholar]
- Laue, P.; Bährs, H.; Chakrabarti, S.; Steinberg, C.E.W. Natural xenobiotics to prevent cyanobacterial and algal growth in freshwater: Contrasting efficacy of tannic acid, gallic acid, and gramine. Chemosphere 2014, 104, 212–220. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, W.J.; Denoyelles, F., Jr. Response of three phytoplankton bioassay techniques in experimental ponds of known limiting nutrient. Hydrobiologia 1976, 49, 65–76. [Google Scholar] [CrossRef]
- Munawar, M.; Munawar, I. Phytoplankton bioassays for evaluating toxicity of in situ sediment contaminants, ecological effects of in situ sediment contaminants. Hydrobiologia 1987, 149, 87–105. [Google Scholar] [CrossRef]
- Hecky, R.E.; Kilham, P. Nutrient limitation of phytoplankton in freshwater and marine environments: A review of recent evidence on the effects of enrichment. Limnol. Oceanogr. 1988, 33, 796–822. [Google Scholar] [CrossRef]
- Zargar, S.; Krishnamurthi, K.; Devi, S.S.; Ghosh, T.; Chakrabarti, T. Temperature-induced stress on growth and expression of Hsp in freshwater alga Scenedesmus Quadricauda. Biomed. Environ. Sci. 2006, 19, 414. [Google Scholar] [PubMed]
- Krell, A.; Beszteri, B.; Dieckmann, G.; Glöckner, G.; Valentin, K.; Mock, T. A new class of ice-binding proteins discovered in a salt-stress-induced cDNA library of the psychrophilic diatom Fragilariopsis cylindrus (Bacillariophyceae). Eur. J. Phycol. 2008, 43, 423–433. [Google Scholar] [CrossRef]
- Guo, R.; Ebenezer, V.; Ki, J.S. Transcriptional responses of heat shock protein 70 (Hsp70) to thermal, bisphenol A, and copper stresses in the dinoflagellate Prorocentrum minimum. Chemosphere 2012, 89, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Ki, J.S. Differential transcription of heat shock protein 90 (HSP90) in the dinoflagellate Prorocentrum minimum by copper and endocrine-disrupting chemicals. Ecotoxicology 2012, 21, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Ki, J.S. Evaluation and validation of internal control genes for studying gene expression in the dinoflagellate Prorocentrum minimum using real-time PCR. Eur. J. Protistol. 2012, 48, 199–206. [Google Scholar] [PubMed]
- Sin, Y.; Hyun, B.; Jeong, B.; Soh, H.Y. Impacts of eutrophic freshwater inputs on water quality and phytoplankton size structure in a temperate estuary altered by a sea dike. Mar. Environ. Res. 2013, 85, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Sin, Y.; Jeong, B. Short term-variations of phytoplankton communities in response to anthropogenic stressors in a highly altered temperate estuary. Estuar. Coast. Shelf Sci. 2015, 156, 83–91. [Google Scholar] [CrossRef]
- Upadhyay, A.; Upadhyaya, I.; Johny, A.K.; Venkitanarayanan, K. Combating pathogenic microorganisms using plant-derived antimicrobials: A minireview of the mechanistic basis. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.; Puupponen-Pimia, R.; Dauer, A.; Aura, A.M.; Saura-Calixto, F. Tannins: Current knowledge of food sources, intake, bioavailability and biological effects. Mol. Nut. Food Res. 2009, 53, 310–329. [Google Scholar] [CrossRef] [PubMed]
- Gross, E.M.; Meyer, H.; Schilling, G. Release and ecological impact of algicidal hydrolysable polyphenols in Myriophyllum spicatum. Phytochemistry 1996, 41, 133–138. [Google Scholar] [CrossRef]
- Körner, S.; Nicklisch, A. Allelopathic growth inhibition of selected phytoplankton species by submerged macrophytes. J. Phycol. 2002, 38, 862–871. [Google Scholar] [CrossRef]
- Sanders, P. An Introduction to Algae Measurements Using in vivo Fluorescence. Available online: http://www.ott.com/en-us/products/download/fluorescence-white-paper/ (accessed on 5 September 2015).
- Ferreira, R.D.; Barbosa, C.C.F.; Novo, E.M. Assessment of in vivo fluorescence method for chlorophyll-a estimation in optically complex waters (Curuai floodplain, Para-Brazil). Acta Limnol. Bras. 2012, 24, 373–386. [Google Scholar] [CrossRef]
- Herrera-Silveira, J.A.; Ramirez-Ramirez, J. Effects of natural phenolic material (tannin) on phytoplankton growth. Limnol. Oceanogr. 1996, 41, 1018–1023. [Google Scholar] [CrossRef]
- Frommhold, I. Chlorophyll degradation: Effects of different concentrations of abscisic acid in barley leaf segments. Biochem. Physiol. Pflanze 1989, 184, 329–331. [Google Scholar] [CrossRef]
- Moharekar, S.T.; Lokhande, S.D.; Hara, T.; Tanaka, R.; Tanaka, A.; Chavan, P.D. Effect of salicylic acid on chlorophyll and carotenoid contents of wheat and moong seedlings. Photosynthetica 2003, 41, 315–317. [Google Scholar] [CrossRef]
- Xu, F.; Cheng, S.; Zhu, J.; Zhang, W. Effects of 5-aminolevulinic acid on chlorophyll, photosynthesis, soluble sugar and flavonoids of Gingko biloba. Not. Bot. Horti Agrobot. 2011, 39, 41. [Google Scholar]
- Schelbert, S.; Aubry, S.; Burla, B.; Agne, B.; Kessler, F.; Krupinska, K.; Hortensteiner, S. Pheophytin Pheophorbide Hydrolase (Pheophytinase) is involved in chlorophyll breakdown during leaf senescence in Arabidposis. Plant Cell 2009, 21, 767–785. [Google Scholar] [CrossRef] [PubMed]
- Henkin, B.M.; Sauer, K. Magnesium ion effects on chloroplast photosystem II fluorescence and photochemistry. J. Photochem. Photobiol. 2008, 26, 277–286. [Google Scholar] [CrossRef]
- Matorin, D.N.; Plekhanov, S.E.; Bratkovskaya, L.B.; Yakovleva, O.V.; Alekseev, A.A. The effect of phenols on the parameters of chlorophyll fluorescence and reactions of P700 in Green algae Scenedesmus quadricauda. Biophysics 2014, 59, 458–465. [Google Scholar] [CrossRef]
- Nakai, S.; Inoue, Y.; Hosomi, M. Algal growth inhibition effects and inducement modes by plant-producing phenols. Water Res. 2001, 35, 1855–1859. [Google Scholar] [CrossRef]
- Ozbay, G.; Chambliss, S.S.; Wikfors, G.H.; Adolf, J.E.; Chintapenta, L.K.; Place, A.R. The growth response of Prorocentrum minimum Pavill. (Dinophyta) to Karlotoxin Exposure. Int. J. Algae 2014, 16, 95–105. [Google Scholar] [CrossRef]
- Haslam, E. Plant polyphenols: Vegetable tannins revisited. In Chemistry and Pharmacology of Natural Products; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Bauer, N.; Zwirnmann, E.; Grossart, H.P.; Hilt, S. Tranformation and allelopathy of natural dissolved organic carbon and tannic acid are affected by solar radiation and bacteria. J. Phycol. 2012, 48, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Bauer, N.; Grossart, H.P.; Hilt, S. Effects of bacterial communities on the sensitivity of the phytoplankton Stephanodiscus minutulus and Desmodesmus armatus to tannic acid. Aquat. Microb. Ecol. 2010, 59, 295–306. [Google Scholar] [CrossRef]
- Cason, J. Synthesis of benzoquinones by oxidation. In Organic Reactions; Adams, R., Fieser, L.F., Blatt, A.H., Johnson, J.R., Snyder, H.R., Eds.; John Wiley: New York, NY, USA; p. 305.
- Roberts, R.J.; Aqius, C.; Saliba, C.; Bossier, P.; Sung, Y.Y. Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health: A review. J. Fish Dis. 2010, 33, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.G. Heat shock factors and the control of the stress response. Biochem. Pharmacol. 2000, 59, 55–63. [Google Scholar] [CrossRef]
- Basu, N.; Todgham, A.E.; Ackerman, P.A.; Bibeau, M.R.; Nakano, K.; Schulte, P.M.; Iwama, G.K. Heat shock protein genes and their functional significance in fish. Gene 2002, 295, 173–183. [Google Scholar] [CrossRef]
- Fu, A.; He, Z.; Cho, H.S.; Lima, A.; Buchann, B.B.; Luan, S. A chloroplast cyclophilin functions in the assembly and maintenance of photosystem II in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2007, 104, 15947–15952. [Google Scholar] [CrossRef] [PubMed]
- Kunmari, S.; Roy, S.; Singh, P.; Singla-Pareek, S.L.; Pareek, A. Cyclophilins: Proteins in search of function. Plant Signal Behav. 2012, 8, e22734. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Solis, J.R.; He, Z.; Lima, A.; Ting, J.; Buchanan, B.B.; Luan, S. A cyclophilin links redox and light signals to cysteine biosynthesis and stress responses in chloroplasts. Proc. Natl. Acad. Sci. USA 2009, 105, 16386–16391. [Google Scholar] [CrossRef] [PubMed]
- Doyle, V.; Virji, S.; Crompton, M. Evidence that cyclophilin-A protects cells against oxidative stress. J. Biochem. 1999, 341, 127–132. [Google Scholar] [CrossRef]
- US Food and Drug Administration. Available online: http://www.fda.gov (accessed on 2 September 2015).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Nucleotide Sequence (5′→3′) | Length (bp) | Cycle | GenBank Access No. and Reference |
|---|---|---|---|---|
| Actin | ||||
| Forward | CAG CGG AAT TCA CGA CAC CAC C | 117 | 25 | JF715156.1 |
| Reverse | CCG ATG CCT GGG AAC ATA GTC G | [47] | ||
| Hsp70 | ||||
| Forward | TGA TCG GTC GCA AAT TCG CCG | 120 | 30 | JN401970.1 |
| Reverse | TCT CCT CGC CCT GTG ATG TCA C | [45] | ||
| Hsp90 | ||||
| Forward | ACG AGG ACT CCA CCA AC | 120 | 30 | JN831315.2 |
| Reverse | TCT GGC CCT CCT TCA TAC GG | [46] | ||
| CYP | ||||
| Forward | AGT CCA TCT ACG GCA GCA AGT TTG | 143 | 30 | JF715159.1 |
| Reverse | TCG AGC CAG GAA GTC TTC ACG G | [47] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, B.; Jeong, E.-S.; Malazarte, J.M.; Sin, Y. Physiological and Molecular Response of Prorocentrum minimum to Tannic Acid: An Experimental Study to Evaluate the Feasibility of Using Tannic Acid in Controling the Red Tide in a Eutrophic Coastal Water. Int. J. Environ. Res. Public Health 2016, 13, 503. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph13050503
Jeong B, Jeong E-S, Malazarte JM, Sin Y. Physiological and Molecular Response of Prorocentrum minimum to Tannic Acid: An Experimental Study to Evaluate the Feasibility of Using Tannic Acid in Controling the Red Tide in a Eutrophic Coastal Water. International Journal of Environmental Research and Public Health. 2016; 13(5):503. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph13050503
Chicago/Turabian StyleJeong, Byungkwan, Eui-Suk Jeong, Jacqueline Martha Malazarte, and Yongsik Sin. 2016. "Physiological and Molecular Response of Prorocentrum minimum to Tannic Acid: An Experimental Study to Evaluate the Feasibility of Using Tannic Acid in Controling the Red Tide in a Eutrophic Coastal Water" International Journal of Environmental Research and Public Health 13, no. 5: 503. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph13050503