Effects of Phthalate Esters on Ipomoea aquatica Forsk. Seedlings and the Soil Microbial Community Structure under Different Soil Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Standards, Assay Kits, and Reagents
2.2. Test Soil and Plants
2.3. Toxicity Tests
2.4. Quantitative Analysis of PAE Compounds
2.5. Determination of Physiological and Biochemical Indices
2.6. DNA Extraction, Pyrosequencing, and Pyrosequencing Analysis
2.7. Statistical Analysis
3. Results
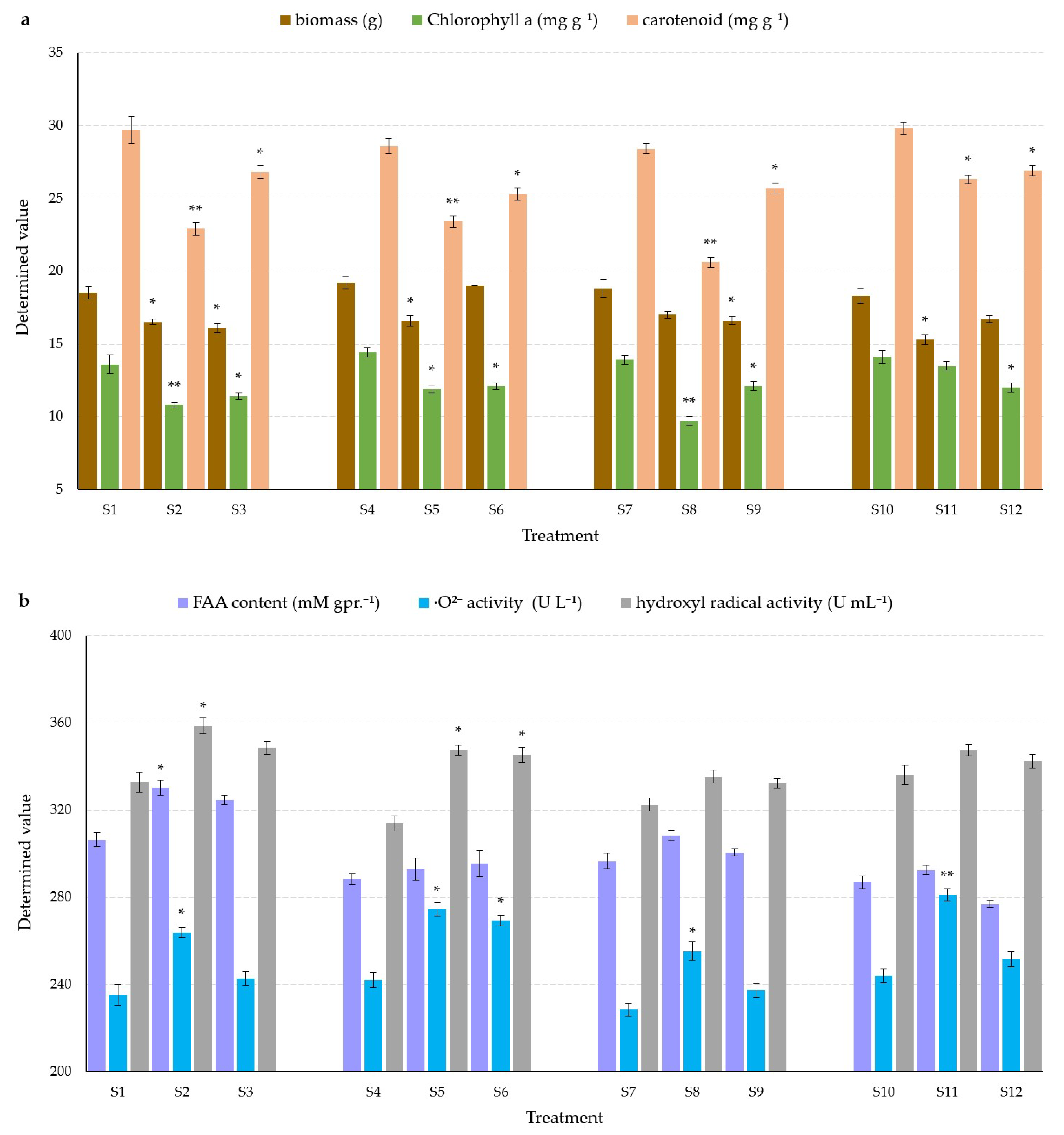
3.1. Effects on Plant Biomass
3.2. Effects on Phytochromes
3.3. Effects on Plant FAA Contents
3.4. Effects on Plant Superoxide Anion Free Radical and Hydroxyl Radical Activity
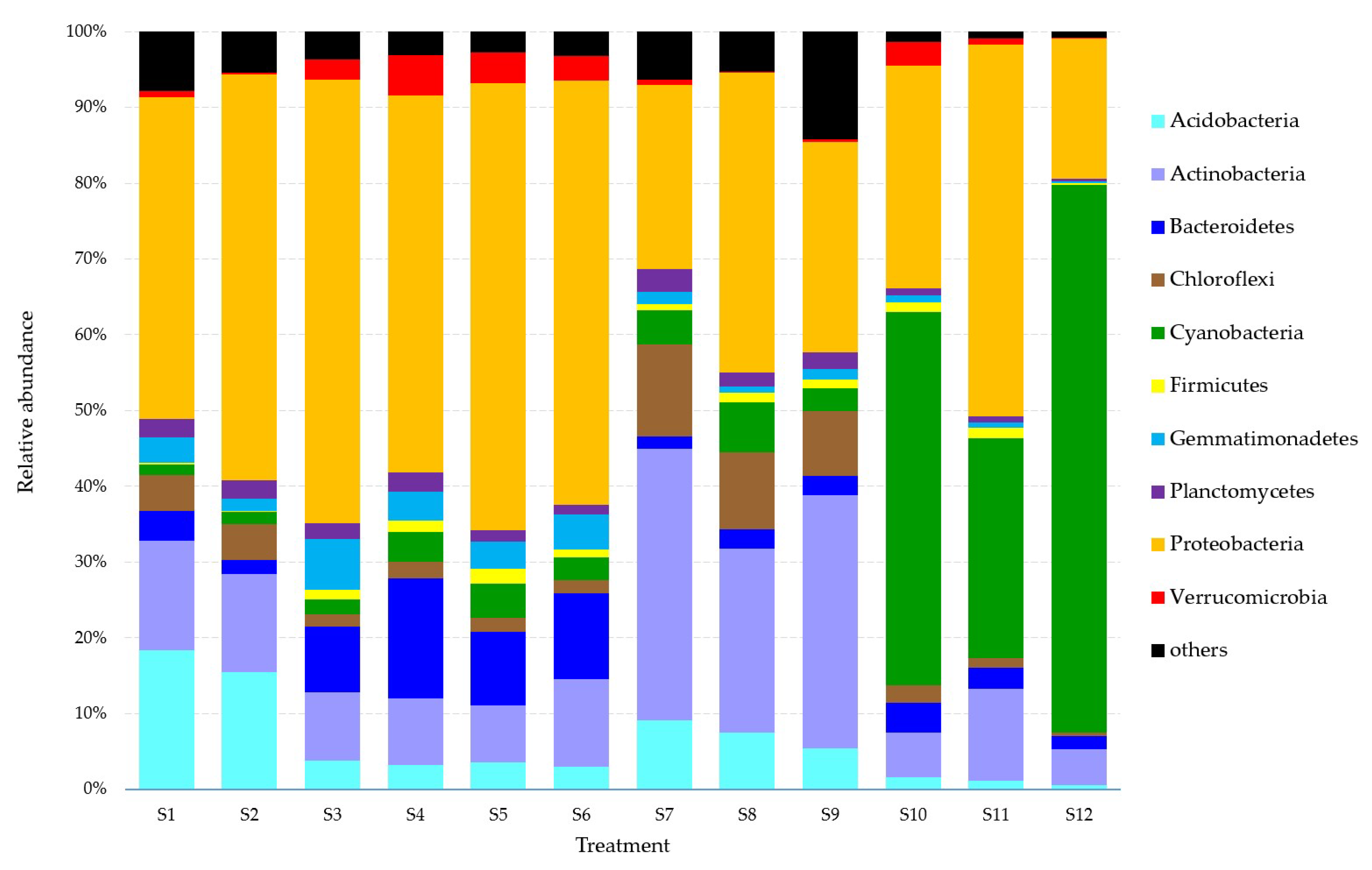
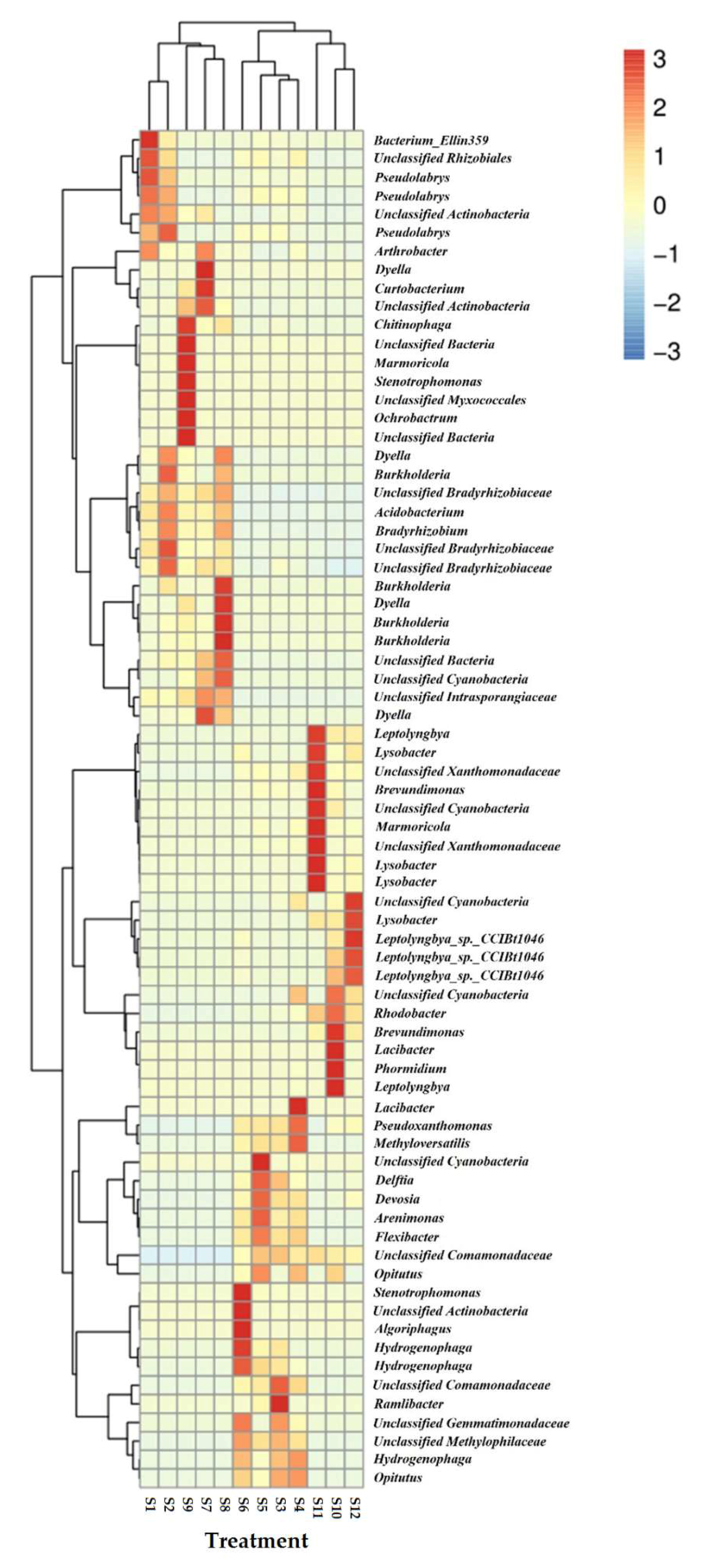
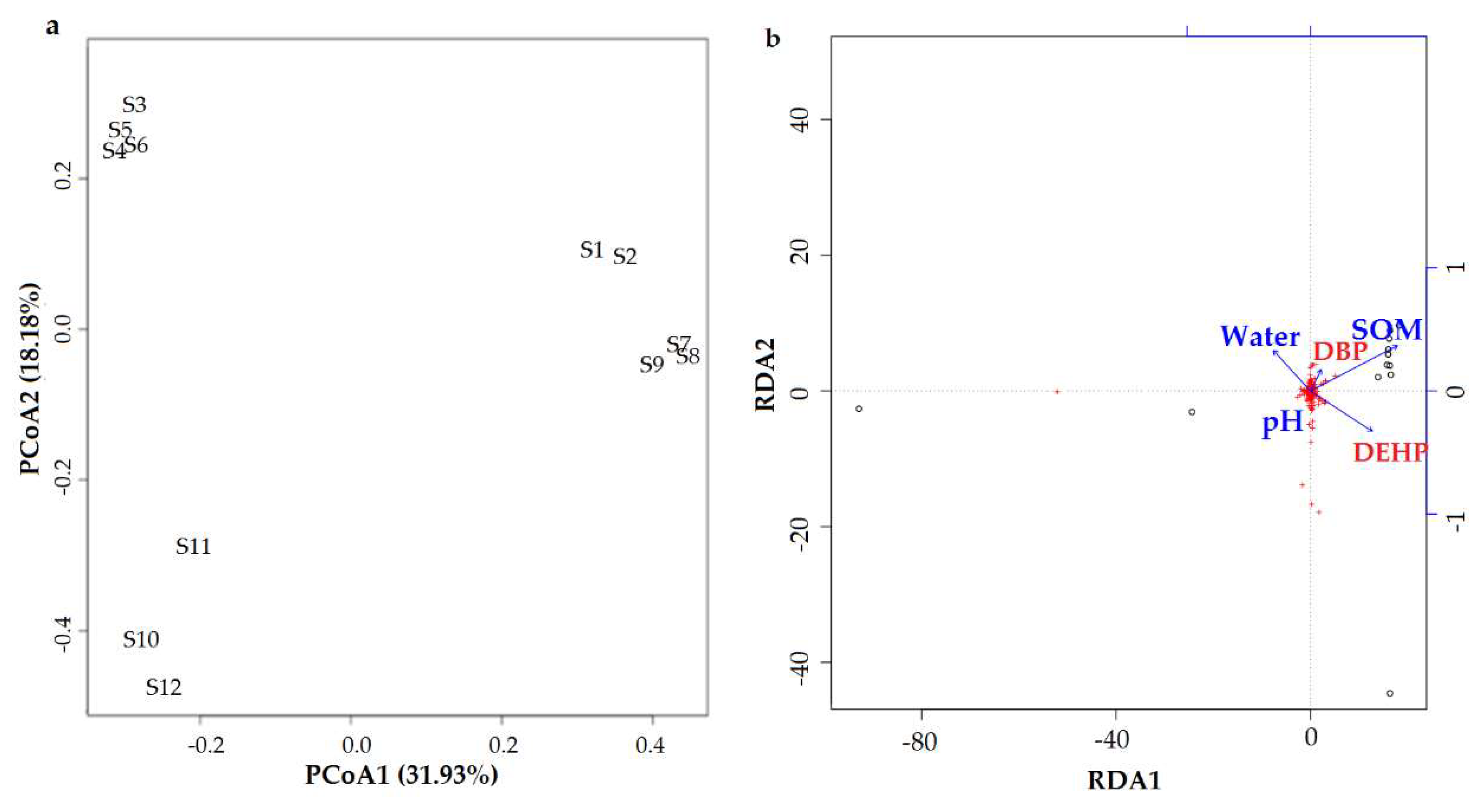
3.5. Effects on the Soil Microbial Community
4. Discussion
4.1. Plant Biomass
4.2. Phytochromes
4.3. FAA Content, Superoxide Anion Free Radical Activity, and OH− Activity
4.4. Effects on the Soil Microbial Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Organization for Economic Cooperation and Development (OECD). Draft Detailed Review Paper: Appraisal of Test Methods for Sex Hormone Disrupting Chemicals; Organization for Economic Cooperation and Development (OECD): Paris, France, 1997.
- United States Environmental Protection Agency (USEPA). Draft Detailed Review Paper on Fish Screening Assays for Endocrine Disruption; United States Environmental Protection Agency (USEPA): Columbus, OH, USA, 2002.
- Liu, X.; Shi, J.; Bo, T.; Zhang, H.; Wu, W.; Chen, Q.; Zhan, X. Occurrence of phthalic acid esters in source waters: A nationwide survey in China during the period of 2009–2012. Environ. Pollut. 2014, 184, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Net, S.; Sempere, R.; Delmont, A.; Paluselli, A.; Ouddane, B. Occurrence, fate, behavior and ecotoxicological state of phthalates in different environmental matrices. Environ. Sci. Technol. 2015, 49, 4019–4035. [Google Scholar] [CrossRef] [PubMed]
- Vinas, P.; Campillo, N.; Pastor-Belda, M.; Oller, A.; Hernandez-Cordoba, M. Determination of phthalate esters in cleaning and personal care products by dispersive liquid-liquid microextraction and liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2015, 1376, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Ikonomou, M.G.; Kelly, B.C.; Blair, J.D.; Gobas, F.A.P.C. An interlaboratory comparison study for the determination of dialkyl phthalate esters in environmental and biological samples. Environ. Toxicol. Chem. 2012, 31, 1948–1956. [Google Scholar] [CrossRef] [PubMed]
- Qian, B. Development of Plasticizers in China; Shanghai Chemical Industry: Shanghai, China, 2016; pp. 36–41. [Google Scholar]
- Benson, R. Hazard to the developing male reproductive system from cumulative exposure to phthalate esters-dibutyl phthalate, diisobutyl phthalate, butylbenzyl phthalate, diethylhexyl phthalate, dipentyl phthalate, and diisononyl phthalate. Regul. Toxicol. Pharmacol. 2009, 53, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Cheng, B. A strategy for nonmigrating highly plasticized PVC. Sci. Rep. 2017, 7, 9277–9282. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.L.; Wu, Q.Y.; Wang, C.; He, T.; Hu, H.Y. Health risk assessment of phthalate esters (PAEs) in drinking water sources of China. Environ. Sci. Pollut. Res. 2015, 22, 3620–3630. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.; Conti, G.O.; Caltabiano, R.; Buffone, A.; Zuccarello, P.; Cormaci, L.; Cannizzaro, M.A.; Ferrante, M. Role of Emerging Environmental Risk Factors in Thyroid Cancer: A Brief Review. Int. J. Environ. Res. Public Health 2019, 16, 1185. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liang, H.; Gao, D.W. Occurrence and risk assessment of phthalate esters (PAEs) in agricultural soils of the Sanjiang plain, Northeast China. Environ. Sci. Pollut. Res. 2017, 24, 19723–19732. [Google Scholar] [CrossRef]
- Cheng, Z.; Li, H.H.; Yu, L.; Yang, Z.B.; Xu, X.X.; Wang, H.S.; Wong, M.H. Phthalate esters distribution in coastal mariculture of Hong Kong, China. Environ. Sci. Pollut. Res. 2018, 25, 17321–17329. [Google Scholar] [CrossRef]
- Ma, T.T.; Wu, L.H.; Chen, L.; Zhang, H.B.; Teng, Y.; Luo, Y.M. Phthalate esters contamination in soils and vegetables of plastic film greenhouses of suburban Nanjing, China and the potential human health risk. Environ. Sci. Pollut. Res. 2015, 22, 12018–12028. [Google Scholar] [CrossRef]
- He, X.; Zang, J.; Liao, P.; Zheng, Y.; Lu, Y.; Zhu, Z.; Shi, Y.; Wang, W. Distribution and Dietary Predictors of Urinary Phthalate Metabolites among Pregnant Women in Shanghai, China. Int. J. Environ. Res. Public Health 2019, 16, 1366. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.T.; Teng, Y.; Christie, P.; Luo, Y.M. Phytotoxicity in seven higher plant species exposed to di-n-butyl phthalate or bis (2-ethylhexyl) phthalate. Front. Environ. Sci. Eng. 2015, 9, 259–268. [Google Scholar] [CrossRef]
- Cai, Q.Y.; Mo, C.H.; Li, Y.H.; Zeng, Q.Y.; Wang, B.G.; Xiao, K.E.; Li, H.Q.; Xu, G.S. The study of PAEs in soils from typical vegetable fields in areas of Guangzhou and Shenzhen, South China. Acta Ecol. Sin. 2005, 25, 283–288. [Google Scholar]
- Chai, C.; Cheng, H.; Ge, W.; Ma, D.; Shi, Y. Phthalic acid esters in soils from vegetable greenhouses in Shandong Peninsula, East China. PLoS ONE 2014, 9, e95701. [Google Scholar] [CrossRef]
- Zheng, S.A.; Xue, Y.H.; Li, X.H.; Duan, Q.H.; Gao, S.B. Phthalate acid esters (PAEs) pollution in soils and agricultural products of vegetable greenhouses in Shouguang City, Shandong Province. J. Agro Environ. Sci. 2016, 35, 492–499. [Google Scholar]
- Christina, J.; Oscar, L.A.; Anne-Charlotte, H.; Rebecka, L.; Emma, H.; Sandra, R. Microplastics shedding from textiles—Developing analytical method for measurement of shed material representing release during domestic washing. Sustainability 2018, 10, 2457–2473. [Google Scholar]
- Xu, Y.; Dai, S.; Meng, K.; Wang, Y.; Ren, W.; Zhao, L.; Christie, P.; Teng, Y. Occurrence and risk assessment of potentially toxic elements and typical organic pollutants in contaminated rural soils. Sci. Total Environ. 2018, 630, 618–629. [Google Scholar] [CrossRef]
- Gao, M.; Dong, Y.; Zhang, Z.; Song, W.; Qi, Y. Growth and antioxidant defense responses of wheat seedlings to di-n-butyl phthalate and di (2-ethylhexyl) phthalate stress. Chemosphere 2017, 172, 418–428. [Google Scholar] [CrossRef]
- Wang, X.J.; Jin, L.; Chen, J.K. Effects of dibutyl phthalate on morphogenesis of Arabidopsis thaliana in vitro. Acta Bot. Boreali Occident. Sin. 2003, 23, 1889–1893. [Google Scholar]
- Wang, X.J.; Jin, L.; Chen, J.K. Effects of DBP on the ultra-structures of in vitro Arabidopsis thaliana leaves. Acta Bot. Boreali Occident. Sin. 2005, 25, 1362–1367. [Google Scholar]
- Yin, R.; Lin, X.G.; Wang, S.G.; Zhang, H.Y. Effect of DBP/DEHP in vegetable planted soil on the quality of capsicum fruit. Chemosphere 2003, 50, 801–805. [Google Scholar] [CrossRef]
- Yin, R.; Lin, X.G.; Wang, S.G.; Zhang, H.Y.; Chen, R.R.; Wang, J.H. Influence of DBP/DEHP pollution in soil on vegetable quality. J. Agro Environ. Sci. 2004, 23, 1–5. [Google Scholar]
- Mo, C.H.; Cai, Q.Y.; Tang, S.R.; Zeng, Q.Y.; Wu, Q.T. Polycyclic aromatic hydrocarbons and phthalic acid esters in vegetables from nine farms of the Pearl River Delta, South China. Arch. Environ. Contam. Toxicol. 2009, 56, 181–189. [Google Scholar] [CrossRef]
- Chen, H.L.; Yao, J.; Wang, F. Soil microbial and enzyme properties as affected by long-term exposure to phthalate esters. Adv. Mater. Res. 2013, 726, 3653–3656. [Google Scholar] [CrossRef]
- Ma, T.T.; Zhou, W.; Chen, L.K.; Wu, L.H.; Christie, P.; Liu, W.X. Toxicity of phthalate esters to lettuce (Lactuca sativa) and the soil microbial community under different soil conditions. PLoS ONE 2018, 13, e0208111. [Google Scholar] [CrossRef]
- Gao, J.; Chen, B.Q. Effects of PAEs on soil microbial activity and catalase activity. J. Soil Water Conserv. 2008, 22, 166–169. [Google Scholar]
- Gao, J. Dynamic effects of PAEs on soil urease and phosphatase. Agric. Sci. Technol. 2010, 11, 189–192. [Google Scholar]
- Chen, H.; Zhuang, R.; Yao, J.; Wang, F.; Qian, Y. A comparative study on the impact of phthalate esters on soil microbial activity. Bull. Environ. Contam. Toxicol. 2013, 91, 217–223. [Google Scholar] [CrossRef]
- Xu, W.; You, Y.; Wang, Z.; Chen, W.; Zeng, J.; Zhao, X.; Su, Y. Dibutyl phthalate alters the metabolic pathways of microbes in black soils. Sci. Rep. 2018, 8, 2605–2617. [Google Scholar] [CrossRef]
- Walker, G.S. Analysis of soils in a forensic context: Comparison of some current and future options. In Criminal and Environmental Soil Forensics; Springer: Dordrecht, The Netherlands, 2009; pp. 397–409. [Google Scholar]
- Fitzpatrick, R.W. Soil: Forensic Analysis. In Wiley Encyclopedia of Forensic Science; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2009. [Google Scholar]
- Dawson, L.A.; Hillier, S. Measurement of soil characteristics for forensic applications. Surf. Interface Anal. 2010, 42, 363–377. [Google Scholar] [CrossRef]
- Hamilton, H.C.; Strickland, M.S.; Wickings, K.; Bradford, M.A.; Fierer, N. Surveying soil faunal communities using a direct molecular approach. Soil Biol. Biochem. 2009, 41, 1311–1314. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Polme, S.; Koljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, N.M.; Wei, Y.; Zhang, J.; Yu, D.; Tong, J. Characterization and Dynamic Shift of Microbial Communities during Start-Up, Overloading and Steady-State in an Anaerobic Membrane Bioreactor. Int. J. Environ. Res. Public Health 2018, 15, 1399. [Google Scholar] [CrossRef] [PubMed]
- Clements, K.D.; Angert, E.R.; Montgomery, W.L.; Choat, J.H. Intestinal microbiota in fishes: What’s known and what’s not. Mol. Ecol. 2014, 23, 1891–1898. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.H.; Roh, S.W.; Whon, T.W.; Jung, M.J.; Kim, M.S.; Park, D.S.; Yoon, C.; Nam, Y.D.; Kim, Y.J.; Choi, J.H.; et al. Insect gut bacterial diversity determined by environmental habitat, diet, developmental stage, and phylogeny of host. Appl. Environ. Microbiol. 2014, 80, 5254–5264. [Google Scholar] [CrossRef] [PubMed]
- Young, J.M.; Weyrich, L.S.; Cooper, A. Forensic soil DNA analysis using high-throughput sequencing: A comparison of four molecular markers. Forensic Sci. Int. Genet. 2014, 13, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.Y.; Liu, W.X.; Wu, L.H.; Hu, P.J.; Ma, T.T.; Luo, Y.M.; Christie, P. Modulation of the efficiency of trace metal phytoremediation by Sedum plumbizincicola by microbial community structure and function. Plant Soil 2017, 421, 1–15. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, Y.; Qian, P.; Gu, J.D. Optimization of phthalic acid batch biodegradation and the use of modified Richards model for modeling degradation. Int. Biodeterior. Biodegrad. 2004, 53, 57–63. [Google Scholar] [CrossRef]
- Lertsirisopon, R.; Soda, S.; Sei, K.; Ike, M. Abiotic degradation of four phthalic acid esters in aqueous phase under natural sunlight irradiation. J. Environ. Sci. 2009, 21, 285–290. [Google Scholar] [CrossRef]
- Chang, B.V.; Yang, C.M.; Cheng, C.H.; Yuan, S.Y. Biodegradation of phthalate esters by two bacteria strains. Chemosphere 2004, 55, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.; Shi, J.; Qin, L.; Feng, M.; Cheng, D.; Wang, T.; Zhang, X. Toxicity evaluation of 4,4’-di-CDPS and 4,4’-di-CDE on green algae Scenedesmus obliquus: Growth inhibition, change in pigment content, and oxidative stress. Environ. Sci. Pollut. Res. 2018, 25, 15630–15640. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Xie, P.; Sheng, X.M.; Wu, Z.B.; Xie, L.Q. Toxicity of cypermethrin on growth, pigments, and superoxide dismutase of Scenedesmus obliquus. Ecotoxicol. Environ. Saf. 2005, 60, 188–192. [Google Scholar]
- Liao, C.S.; Nishikawa, Y.; Shih, Y.T. Characterization of di-n-butyl phthalate phytoremediation by garden lettuce (lactuca sativa l. var. longifolia) through kinetics and proteome analysis. Sustainability 2019, 11, 1625. [Google Scholar]
- Yuan, L.; Liu, X.G.; Tang, Y. The comparison of free radical scavenging capacity of carotenoids. Pack. Food Mach. 2015, 33, 7–11. [Google Scholar]
- Rupani, P.F.; Embrandiri, A.; Ibrahim, M.H.; Ghole, V.; Lee, C.T.; Abbaspour, M. Effects of different vermicompost extracts of palm oil mill effluent and palm-pressed fiber mixture on seed germination of mung bean and its relative toxicity. Environ. Sci. Pollut. Res. 2018, 25, 35805–35810. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.T.; Christie, P.; Luo, Y.M.; Teng, Y. Physiological and antioxidant responses of germinating mung bean seedlings to phthalate esters in soil. Pedosphere 2014, 24, 107–115. [Google Scholar] [CrossRef]
- Li, Y.J.; Yuan, B.Z.; Bie, Z.L.; Kang, Y. Effect of drip irrigation criteria on yield and quality of muskmelon grown in greenhouse conditions. Agric. Water Manag. 2012, 109, 30–35. [Google Scholar] [CrossRef]
- Trawiński, J.; Skibiński, R. Studies on photodegradation process of psychotropic drugs: A review. Environ. Sci. Pollut. Res. 2016, 24, 1152–1199. [Google Scholar] [CrossRef]
- Pandey, S.; Parvez, S.; Sayeed, I.; Haque, R.; Bin-Hafeez, B.; Raisuddin, S. Biomarkers of oxidative stress: A comparative study of river Yamuna fish Wallago attu (Bl & Schn). Sci. Total Environ. 2003, 309, 105–115. [Google Scholar]
- Liu, H.R.; Song, H.X.; Liu, D.P.; Guan, C.Y.; Liu, Q.; Chen, S.Y. Dynamics changes of soluble sugar and free amino acid contents in stem and leaf of different oilseed rape varieties. Acta Agric. Boreali Occident. Sin. 2007, 16, 123–126. [Google Scholar]
- Luo, L.X.; Sun, T.H.; Jin, Y.H. Accumulation of superoxide radical in wheat leaves under cadmium stress. Actaentiae Circumstantiae 1998, 18, 495–499. [Google Scholar]
- Niepceron, M.; Martinlaurent, F.; Crampon, M.; Portetkoltalo, F.; Akpavinceslas, M.; Legras, M.; Bru, D.; Bureau, F.; Bodilis, J. Gamma-proteobacteria as a potential bioindicator of a multiple contamination by polycyclic aromatic hydrocarbons (PAHs) in agricultural soils. Environ. Pollut. 2013, 180, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Yasin, F. Actinobacterial isolates from tin tailings and forest soil for bioremediation of heavy metals. J. Trop. For. Sci. 2014, 26, 153–162. [Google Scholar]
- Amoroso, M.J.; Abate, C.M. Bioremediation of copper, chromium and cadmium by actinomycetes from contaminated soil. In Bio-Geo Interactions in Metal-Contaminated Soils; Springer: Berlin/Heidelberg, Germany, 2012; Volume 31, pp. 349–364. [Google Scholar]
- Kumar, A.; Bisht, B.S.; Joshi, V.D. Bioremediation potential of three acclimated bacteria with reference to heavy metal removal from waste. Int. J. Environ. Sci. 2011, 2, 896–908. [Google Scholar]
- Lin, Y.; Wang, X.; Wang, B.; Mohamad, O.; Wei, G. Bioaccumulation characterization of zinc and cadmium by Streptomyces zinciresistens, a novel actinomycete. Ecotoxicol. Environ. Saf. 2012, 77, 7–17. [Google Scholar] [CrossRef]
- Alvarez, V.M.; Marques, J.M.; Korenblum, E.; Seldin, L. Comparative bioremediation of crude oil-amended tropical soil microcosms by natural attenuation, bioaugmentation or bioenrichment. Appl. Environ. Soil Sci. 2011, 2011, 1–10. [Google Scholar] [CrossRef]
- Fuentes, M.S.; Sáez, J.M.; Benimeli, C.S.; Amoroso, M.J. Lindane biodegradation by defined consortia of indigenous streptomyces, strains. Water Air Soil Pollut. 2011, 222, 217–231. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Goebel, B.M.; Pace, N.R. Impact of culture-independent studies on the emerging phylogenetic view of bacterial diversity. J. Bacteriol. 1998, 180, 4765–4774. [Google Scholar]
- Barns, S.M.; Takala, S.L.; Kuske, C.R. Wide distribution and diversity of members of the bacterial kingdom Acidobacterium in the environment. Appl. Environ. Microbiol. 1999, 65, 1731–1737. [Google Scholar]
- Lee, S.H.; Ka, J.O.; Cho, J.C. Members of the phylum Acidobacteria are dominant and metabolically active in rhizosphere soil. FEMS Microbiol. Lett. 2010, 285, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Radeva, G.; Kenarova, A.; Bachvarova, V.; Flemming, K.; Selenska-Pobell, S. Bacterial diversity at abandoned uranium mining and milling sites in bulgaria as revealed by 16s rRNA genetic diversity study. Water Air Soil Pollut. 2013, 224, 1748. [Google Scholar] [CrossRef]
- Chen, R.; Zhao, L.; Sun, X.; Qu, J.; Lu, P.; Shen, W. Microbial community structure in petroleum polluted soil at north of Shaanxi. Environ. Pollut. Control 2017, 11, 1181–1185. [Google Scholar]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed]
- Vierheilig, J.; Farnleitner, A.H.; Kollanur, D.; Blöschl, G.; Reischer, G.H. High abundance of genetic bacteroidetes markers for total fecal pollution in pristine alpine soils suggests lack in specificity for feces. J. Microbiol. Methods 2012, 88, 433–435. [Google Scholar] [CrossRef] [PubMed]
- Beghelli, F.G.D.S.; Frascareli, D.; Pompêo, M.L.M.; Moschini-Carlos, V. Trophic state evolution over 15 years in a tropical reservoir with low nitrogen concentrations and cyanobacteria predominance. Water Air Soil Pollut. 2016, 227, 1–15. [Google Scholar]
- Dorokhova, M.F.; Kosheleva, N.E.; Terskaya, E.V. Algae and cyanobacteria in soils of Moscow. Am. J. Plant Sci. 2015, 6, 2461–2471. [Google Scholar] [CrossRef]
- Singh, R.N. Role of Blue-Green Algae in Nitrogen Economy of Indian Agriculture; Indian Council of Agricultural Research: New Delhi, India, 1961. [Google Scholar]
- Li, A.; Qu, Y.Y.; Pi, W.Q.; Zhou, J.T.; Gai, Z.H.; Xu, P. Metabolic characterization and genes for the conversion of biphenyl in Dyella ginsengisoli La-4. Biotechnol. Bioeng. 2012, 109, 609–613. [Google Scholar] [CrossRef]
- Li, A.; Qu, Y.Y.; Zhou, J.; Fang, M. Characterization of a newly isolated biphenyl-degrading bacterium, Dyella ginsengisoli La-4. Appl. Biochem. Biotechnol. 2009, 159, 687–695. [Google Scholar] [CrossRef]
- Salles, J.F.; van Veen, J.A.; van Elsas, J.D. Multivariate analyses of Burkholderia species in soil: Effect of crop and land use history. Appl. Environ. Microbiol. 2004, 70, 4012. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.K.; Mishra, B.M.; Tripathi, P. Salinity stress responses in the plant growth promoting rhizobacteria, Azospirillum spp. J. Biosci. 1998, 23, 463–471. [Google Scholar] [CrossRef]
- Yildirim, E.; Turan, M.; Donmez, M.F. Mitigation of salt stress in radish (Raphanus sativus L.) by plant growth promoting rhizobacteria. Roum. Biotechnol. Lett. 2008, 13, 3933–3943. [Google Scholar]
- Upadhyay, S.K.; Singh, D.P.; Saikia, R. Genetic diversity of plant growth promoting rhizobacteria isolated from rhizospheric soil of wheat under saline condition. Curr. Microbiol. 2009, 59, 489–496. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Controlled Variables | Day 0 | Day 30 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PAE Con. (mg kg−1) | SOM Content (%) | Water Content (% Max WHC) | pH | PAE Conc. (mg kg−1) | SOM Content (%) | Water Content (% Max WHC) | pH | PAEs Conc. (mg kg−1) | SOM Content (%) | Water Content (% Max WHC) | pH | ||
| Group 1 | S1 | 0 Control | 2.0 | 80 | 7.0 | - | 1.9 ± 0.3 | 82 ± 2 | 6.5 ± 0.4 | - | 1.6 ± 0.2 | 85 ± 3 | 6.7 ± 0.3 |
| S2 | 20 DBP | 19.8 ± 0.4 | 1.8 ± 0.3 | 77 ± 5 | 7.0 ± 0.2 | 15.6 ± 0.5 | 1.5 ± 0.3 | 83 ± 5 | 7.2 ± 0.3 | ||||
| S3 | 20 DEHP | 20.3 ± 0.2 | 1.9 ± 0.3 | 79 ± 4 | 6.9 ± 0.2 | 17.2 ± 0.4 | 1.7 ± 0.2 | 85 ± 4 | 7.1 ± 0.2 | ||||
| Group 2 | S4 | 0 Control | 8.5 | - | 2.0 ± 0.2 | 76 ± 3 | 8.2 ± 0.2 | - | 1.7 ± 0.2 | 86 ± 2 | 8.3 ± 0.1 | ||
| S5 | 20 DBP | 19.1 ± 0.5 | 2.0 ± 0.4 | 83 ± 4 | 8.5 ± 0.3 | 14.3 ± 0.4 | 1.8 ± 0.4 | 86 ± 3 | 8.4 ± 0.2 | ||||
| S6 | 20 DEHP | 19.7 ± 0.4 | 2.0 ± 0.3 | 79 ± 5 | 8.2 ± 0.2 | 15.5 ± 0.5 | 1.7 ± 0.3 | 83 ± 3 | 8.4 ± 0.3 | ||||
| Group 3 | S7 | 0 Control | 4.0 | 60 | 7.0 | - | 3.6 ± 0.2 | 63 ± 5 | 7.1 ± 0.3 | - | 3.3 ± 0.4 | 58 ± 3 | 7.3 ± 0.4 |
| S8 | 20 DBP | 18.7 ± 0.5 | 3.9 ± 0.4 | 58 ± 3 | 7.1 ± 0.2 | 15.9 ± 0.3 | 3.6 ± 0.2 | 56 ± 5 | 7.2 ± 0.3 | ||||
| S9 | 20 DEHP | 19.9± 0.3 | 4.2 ± 0.3 | 56 ± 4 | 6.8 ± 0.3 | 16.8 ± 0.3 | 4.0 ± 0.3 | 55 ± 3 | 6.9 ± 0.3 | ||||
| Group 4 | S10 | 0 Control | 8.5 | - | 4.4 ± 0.2 | 59 ± 2 | 8.0 ± 0.2 | - | 4.1 ± 0.3 | 56 ± 2 | 8.2 ± 0.3 | ||
| S11 | 20 DBP | 19.3 ± 0.6 | 3.9 ± 0.3 | 62 ± 4 | 8.1 ± 0.2 | 14.6 ± 0.3 | 3.5 ± 0.2 | 55 ± 4 | 8.4 ± 0.4 | ||||
| S12 | 20 DEHP | 20.5 ± 0.3 | 3.7 ± 0.4 | 60 ± 2 | 8.8 ± 0.4 | 16.8 ± 0.5 | 3.4 ± 0.2 | 56 ± 3 | 8.9 ± 0.3 | ||||
| Treatment | Ace | Chao | Shannon | Simpson | Coverage | |
|---|---|---|---|---|---|---|
| Group 1 | S1 | 5596 | 4344 | 7.10 | 0.0016 | 0.80 |
| S2 | 4854 ** | 3664 ** | 6.84 ** | 0.0022 ** | 0.83 | |
| S3 | 4585 ** | 3422 ** | 6.64 ** | 0.0033 ** | 0.87 | |
| Group 2 | S4 | 4258 | 3263 | 6.75 | 0.0028 | 0.84 |
| S5 | 4042 * | 3019 * | 6.53 | 0.0033 ** | 0.86 | |
| S6 | 4183 | 3202 | 6.41 | 0.0052 ** | 0.88 | |
| Group 3 | S7 | 5331 | 4163 | 7.06 | 0.0026 | 0.83 |
| S8 | 4218 ** | 3368 ** | 6.69 * | 0.0035 ** | 0.88 | |
| S9 | 4571 ** | 3588 ** | 6.36 ** | 0.0106 ** | 0.89 | |
| Group 4 | S10 | 2517 | 1982 | 4.77 | 0.0934 | 0.92 |
| S11 | 2228 ** | 1722 ** | 4.62 | 0.0398 ** | 0.95 | |
| S12 | 1985 ** | 1505 ** | 3.03 ** | 0.3249 ** | 0.96 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, T.; Liu, L.; Zhou, W.; Chen, L.; Christie, P. Effects of Phthalate Esters on Ipomoea aquatica Forsk. Seedlings and the Soil Microbial Community Structure under Different Soil Conditions. Int. J. Environ. Res. Public Health 2019, 16, 3489. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16183489
Ma T, Liu L, Zhou W, Chen L, Christie P. Effects of Phthalate Esters on Ipomoea aquatica Forsk. Seedlings and the Soil Microbial Community Structure under Different Soil Conditions. International Journal of Environmental Research and Public Health. 2019; 16(18):3489. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16183489
Chicago/Turabian StyleMa, Tingting, Linwei Liu, Wei Zhou, Like Chen, and Peter Christie. 2019. "Effects of Phthalate Esters on Ipomoea aquatica Forsk. Seedlings and the Soil Microbial Community Structure under Different Soil Conditions" International Journal of Environmental Research and Public Health 16, no. 18: 3489. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16183489