Gestational Pesticide Exposure and Child Respiratory Health

 , and
, and
Abstract
:1. Introduction
2. Methods
2.1. Pesticide Exposure Assessment
2.2. Paraoxonase (PON1) Genotype Assessment
2.3. Respiratory Outcomes
2.4. Covariates
2.5. Data Analysis
3. Results
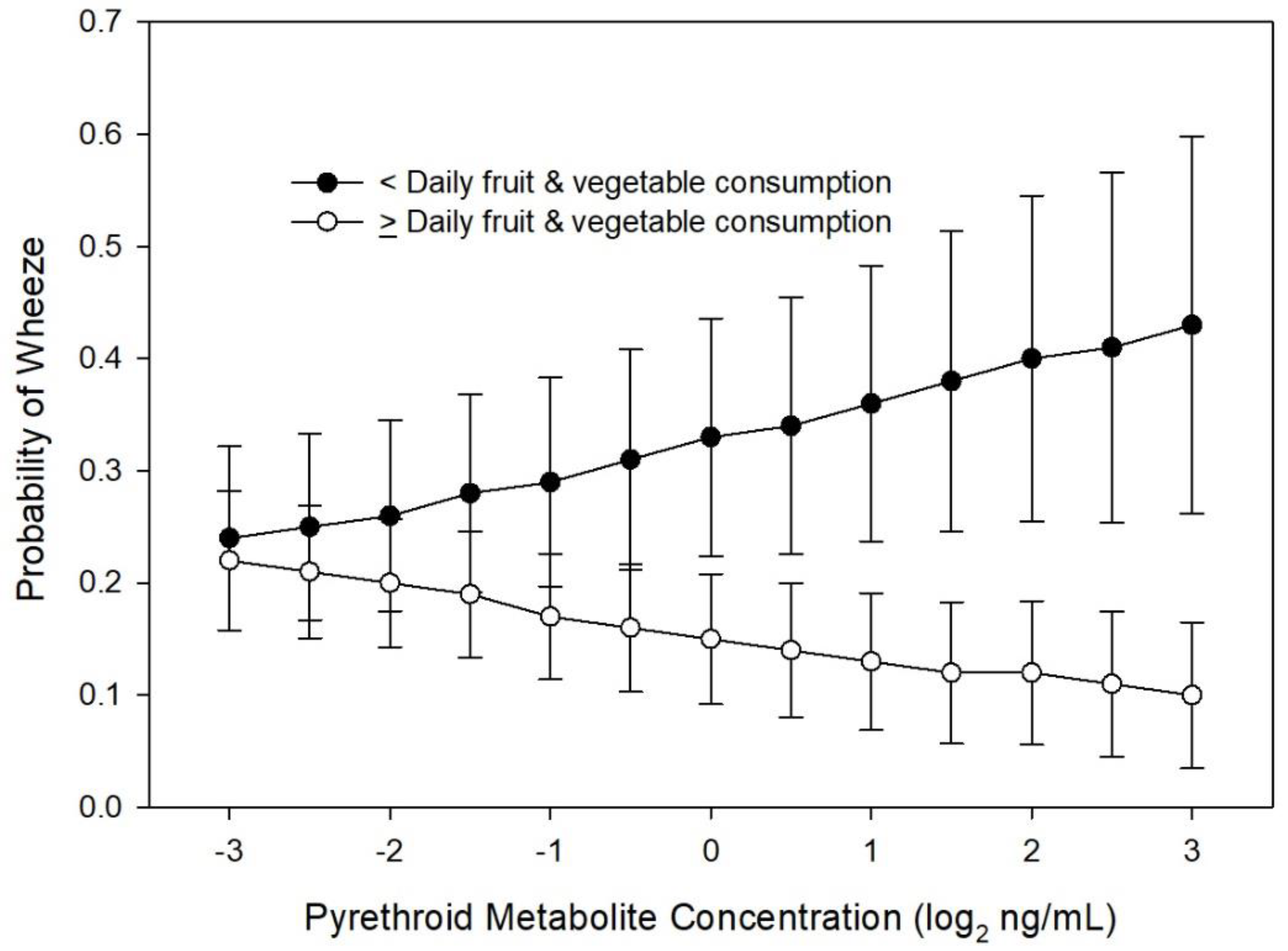
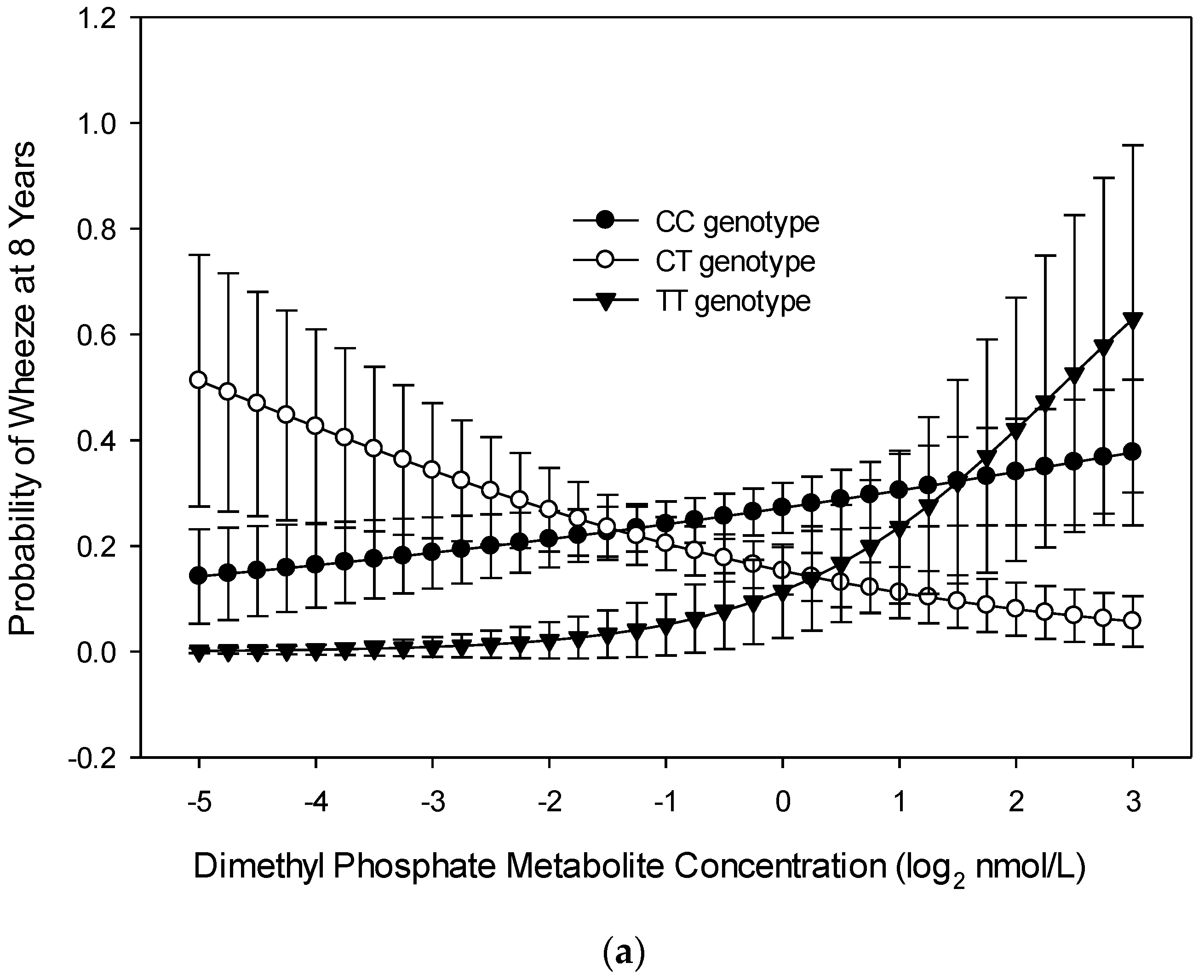
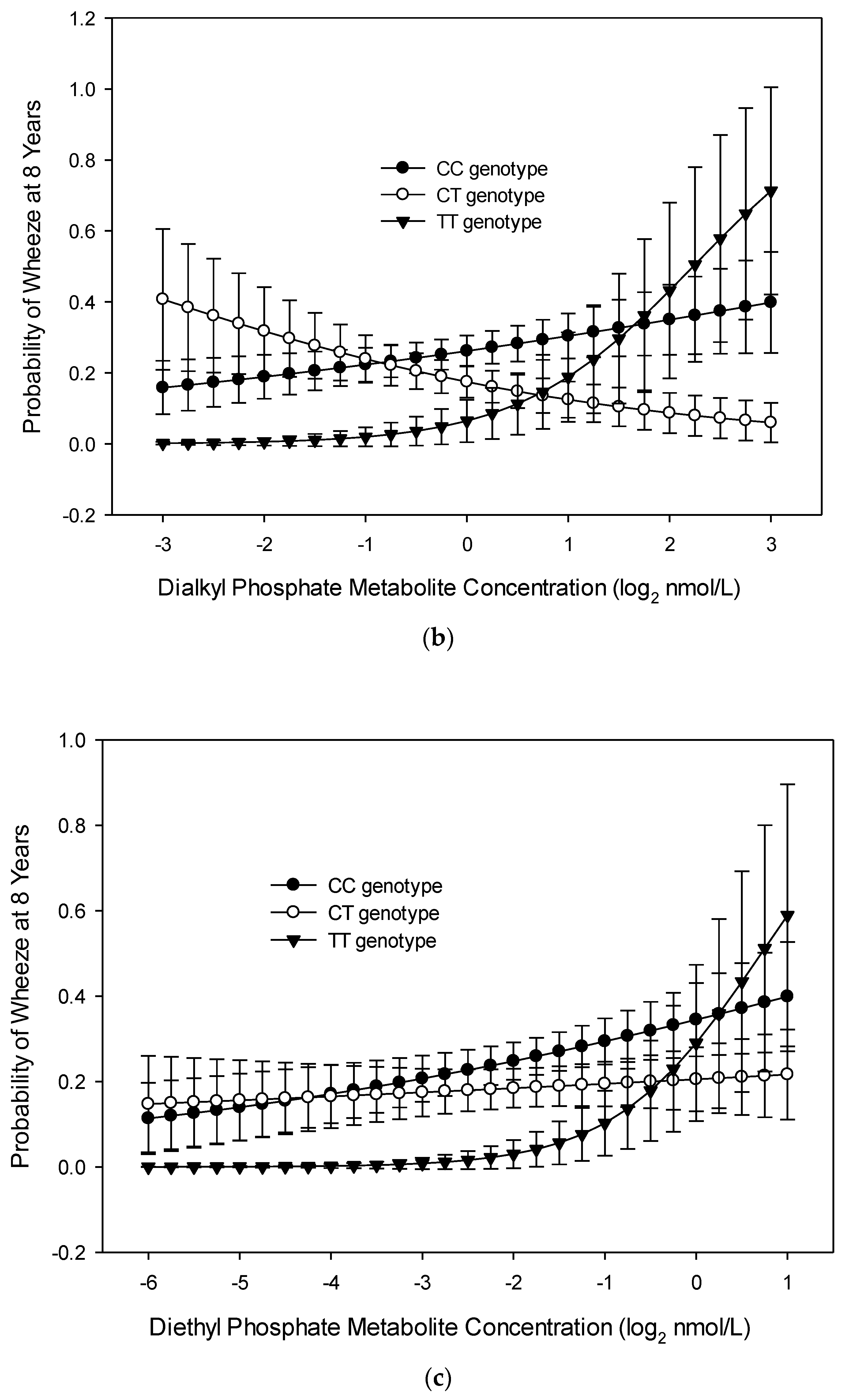
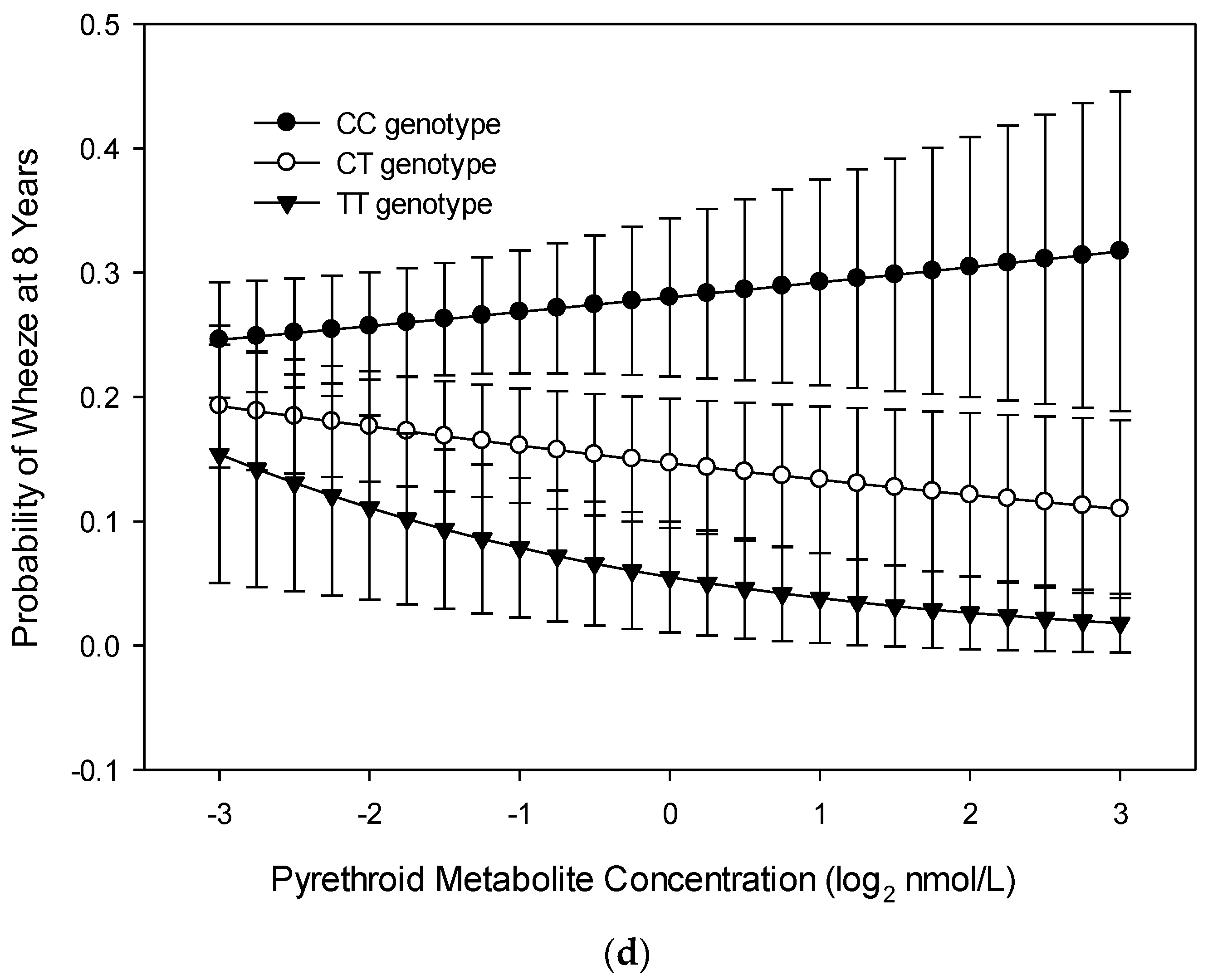
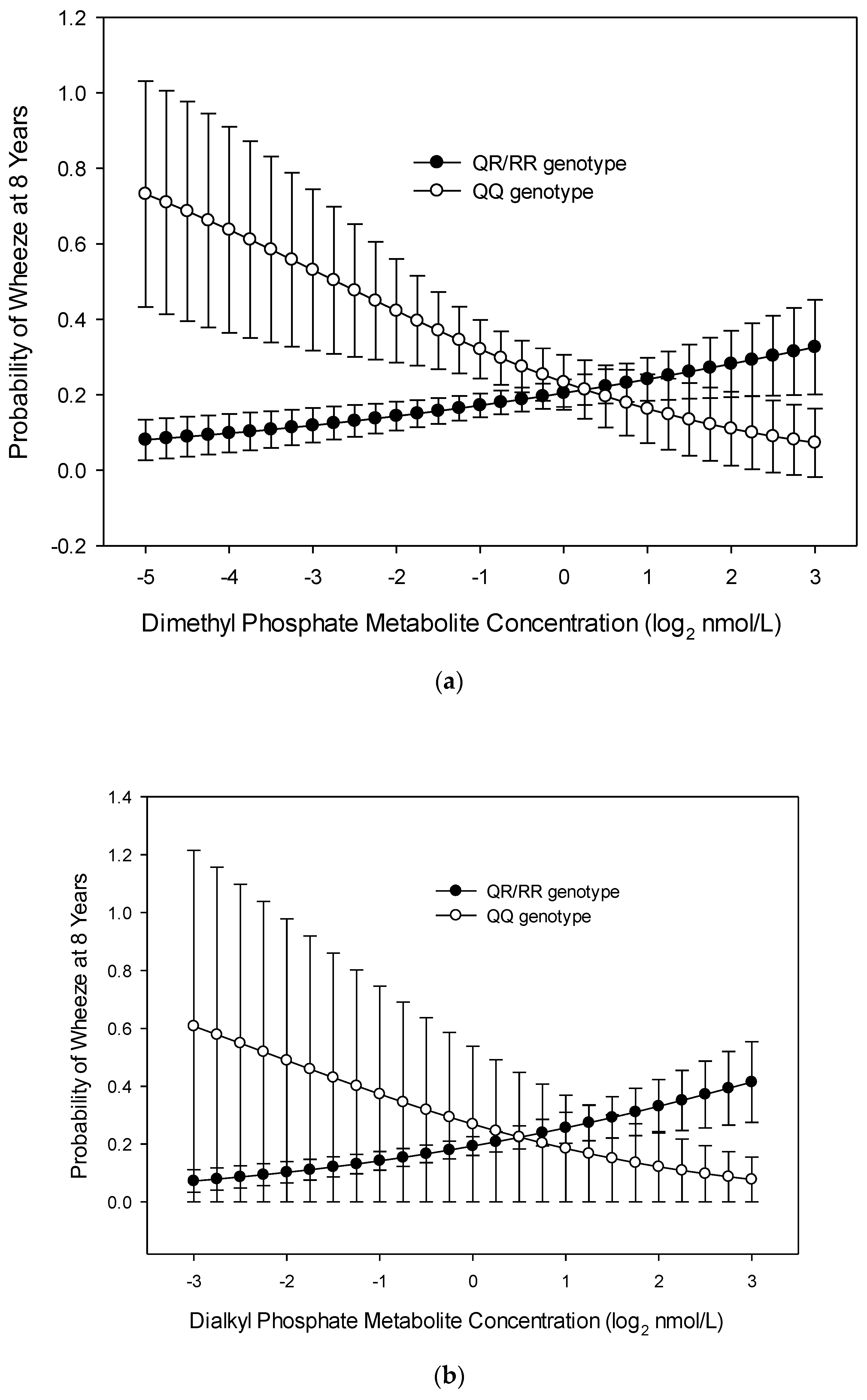
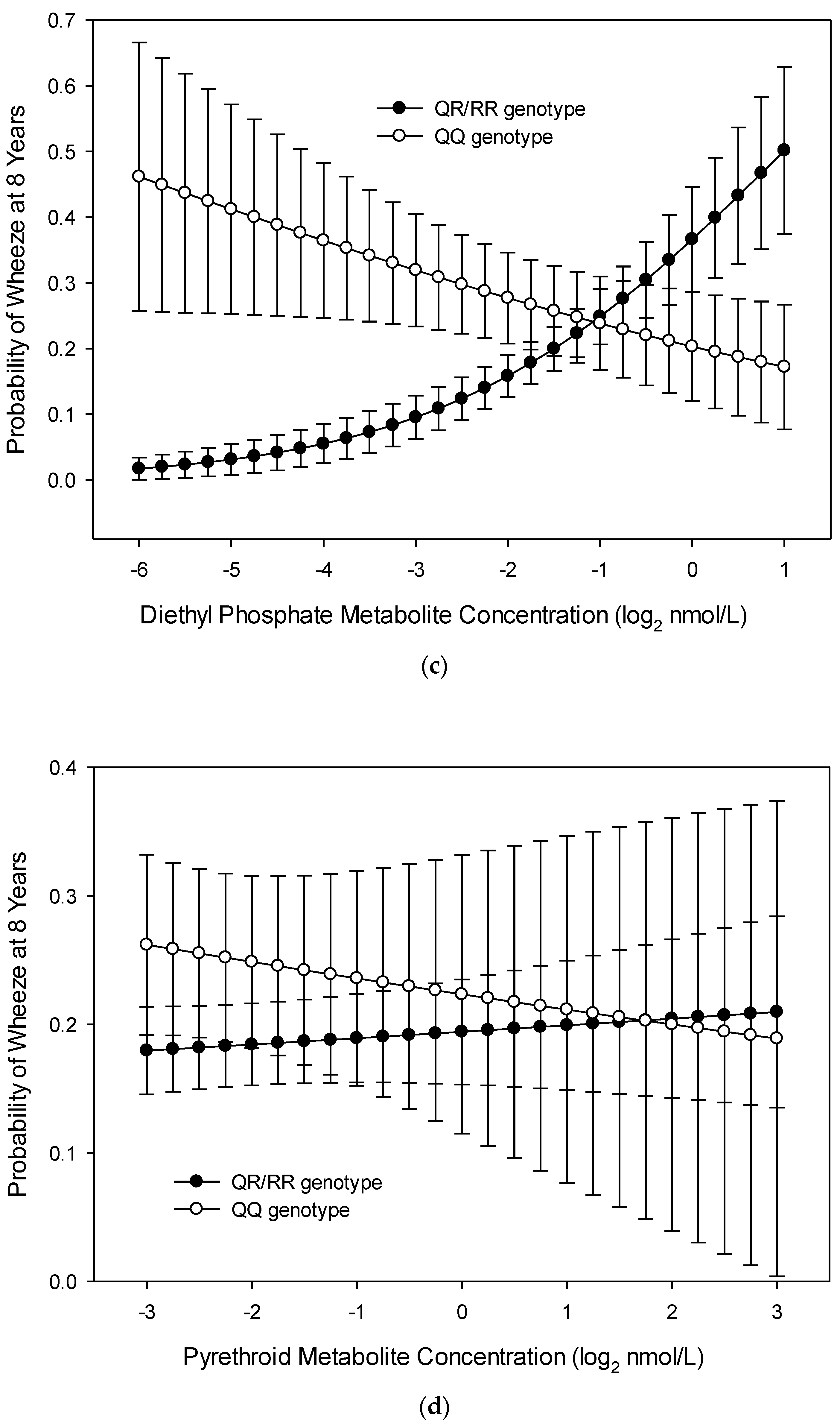
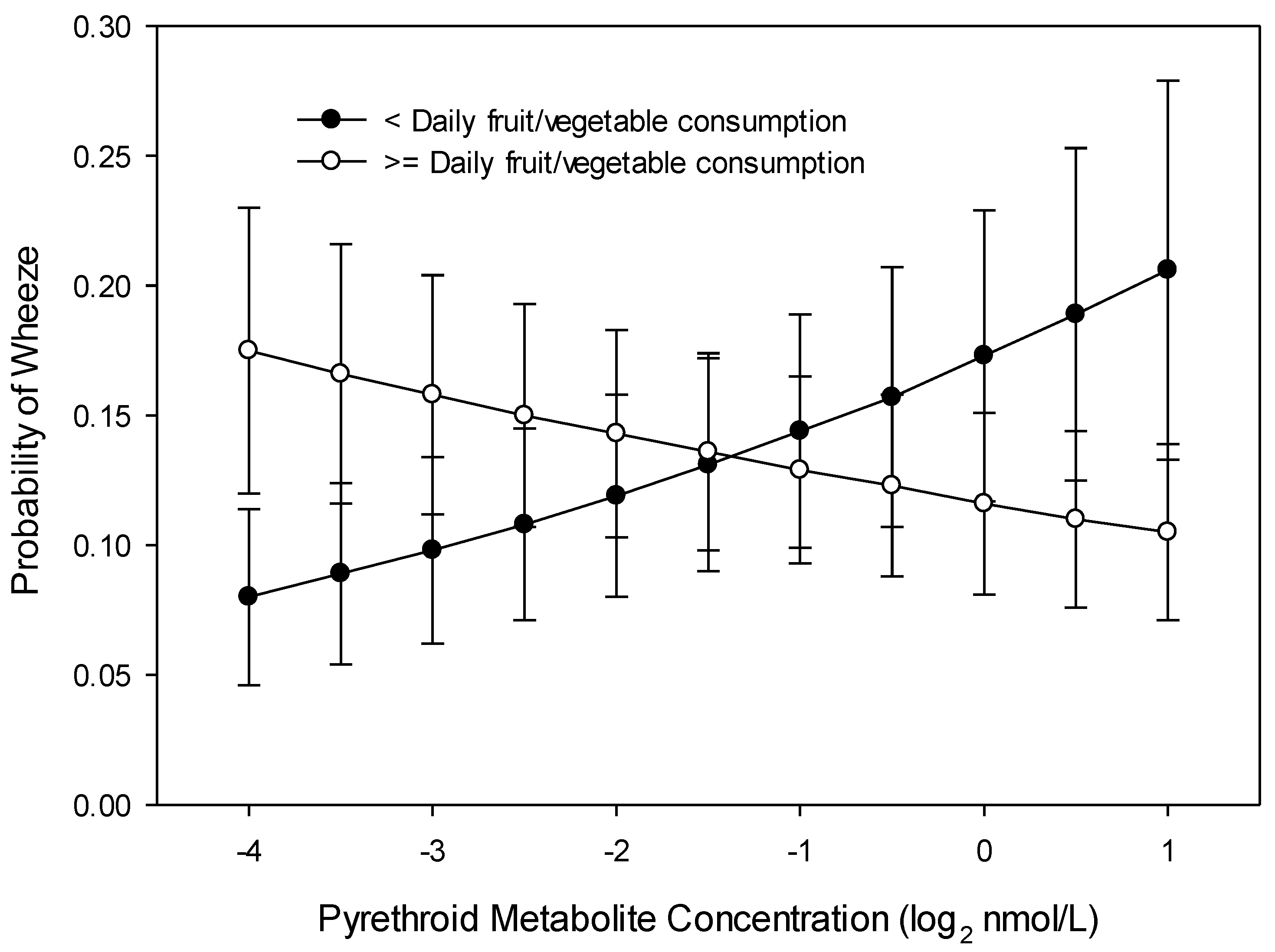
3.1. Wheeze at Age 8 Years
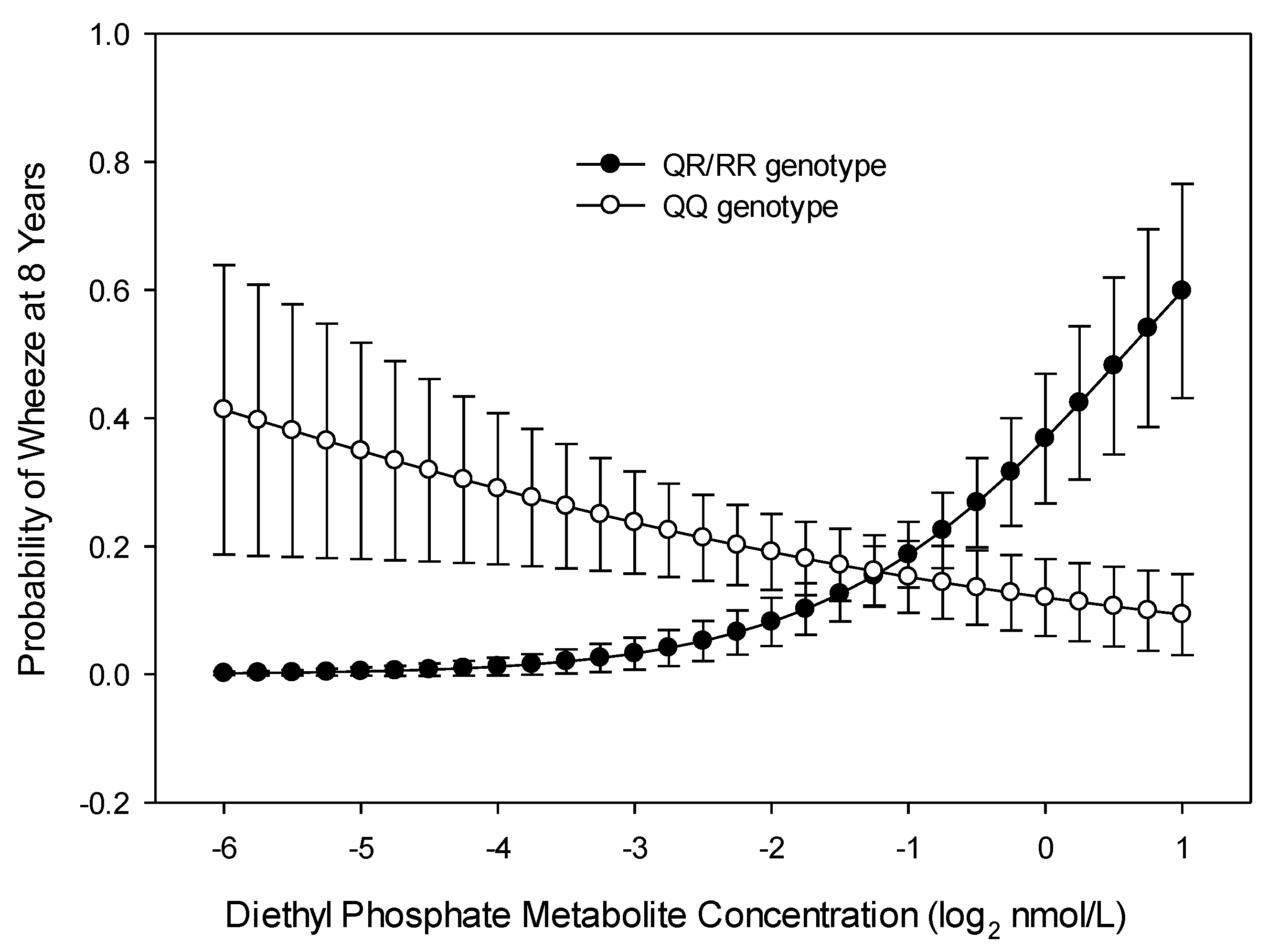
3.2. Wheeze across Childhood (Repeated Measures over 8 Years)
3.3. FEV1
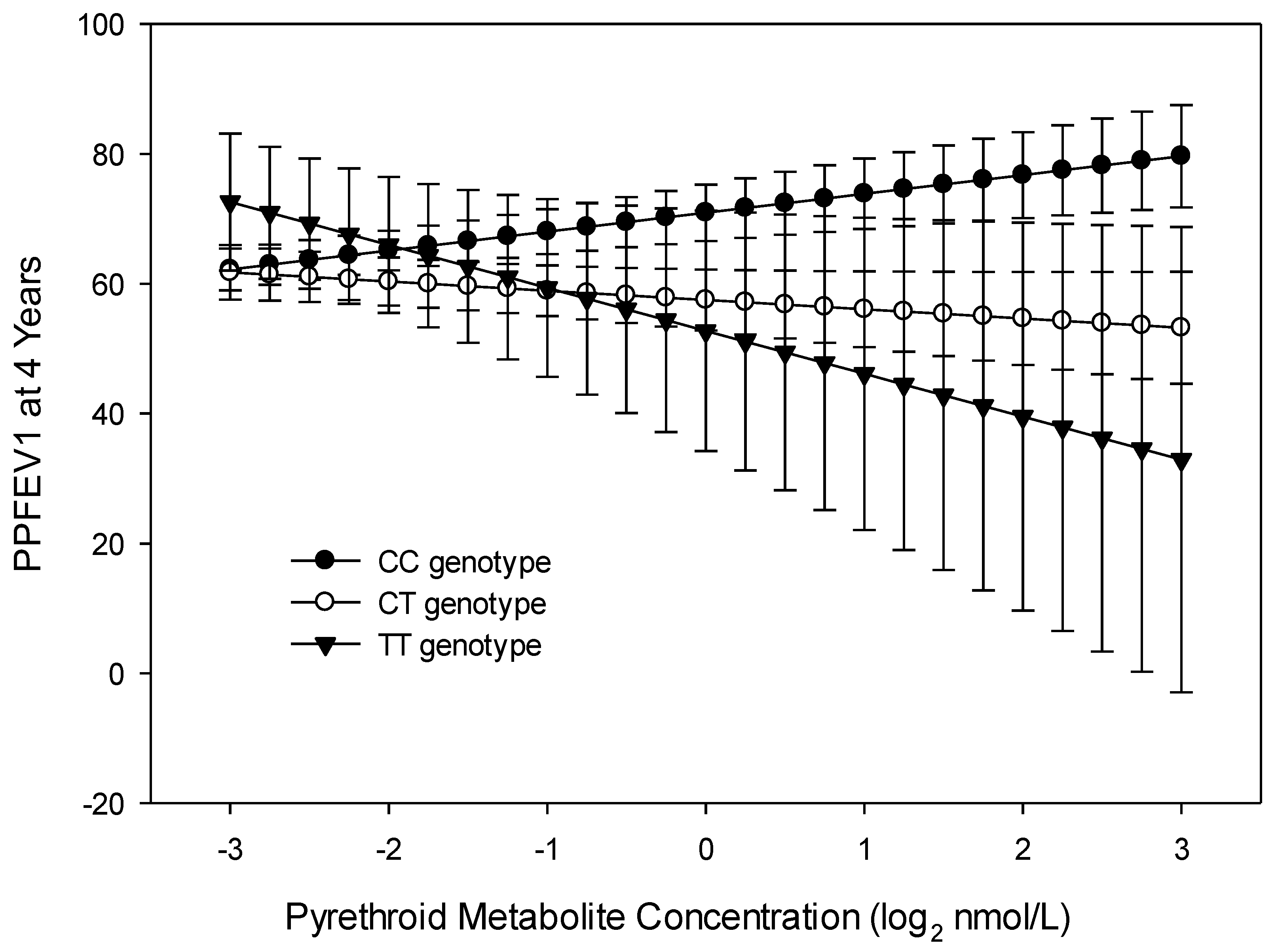
3.4. Trajectory Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- National Center for Health Statistics. National Health and Nutrition Examintation Survey. 2017. Available online: https://www.cdc.gov/nchs/nhanes/index.htm (accessed on 28 August 2020).
- Cicutto, L.; To, T.; Murphy, S. A randomized controlled trial of a public health nurse-delivered asthma program to elementary schools. J. Sch. Health 2013, 83, 876–884. [Google Scholar] [CrossRef] [PubMed]
- National Center for Environmental Health. Asthma’s Impact on Children and Adolescents; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2005.
- Tai, T.; Bame, S.I. Cost-benefit analysis of childhood asthma management through school-based clinic programs. J. Community Health 2011, 36, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.P.; Lahewala, S.; Arora, S.; Agnihotri, K.; Panaich, S.S.; Secord, E.; Levine, D. Asthma: Hospitalization Trends and Predictors of In-Hospital Mortality and Hospitalization Costs in the USA (2001–2010). Int. Arch. Allergy Immunol. 2015, 168, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.M.; Barros, L.; Simoes, F. Health-related quality of life in paediatric asthma: Children’s and parents’ perspectives. Psychol. Health Med. 2015, 20, 940–954. [Google Scholar] [CrossRef]
- Wang, L.Y.; Zhong, Y.; Wheeler, L. Direct and indirect costs of asthma in school-age children. Prev. Chronic Dis. 2005, 2, A11. [Google Scholar]
- Moorman, J.E.; Akinbami, L.J.; Bailey, C.M.; Zahran, H.S.; King, M.E.; Johnson, C.A.; Liu, X. National surveillance of asthma: United States, 2001–2010. Vital Health Stat. 2012, 35, 1–58. [Google Scholar]
- Perla, M.E.; Rue, T.; Cheadle, A.; Krieger, J.; Karr, C.J. Population-based comparison of biomarker concentrations for chemicals of concern among Latino-American and non-Hispanic white children. J. Immigr. Minor. Health 2015, 17, 802–819. [Google Scholar] [CrossRef]
- Raanan, R.; Harley, K.G.; Balmes, J.R.; Bradman, A.; Lipsett, M.; Eskenazi, B. Early-life exposure to organophosphate pesticides and pediatric respiratory symptoms in the CHAMACOS cohort. Environ. Health Perspect. 2015, 123, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Nembhard, W.N.; Kan, H.; Becker, A.; Talbott, E.O. Residential pesticide use is associated with children’s respiratory symptoms. J. Occup. Environ. Med. 2012, 54, 1281–1287. [Google Scholar] [CrossRef]
- Ye, M.; Beach, J.; Martin, J.W.; Senthilselvan, A. Occupational pesticide exposures and respiratory health. Int. J. Environ. Res. Public Health 2013, 10, 6442–6471. [Google Scholar] [CrossRef]
- Gascon, M.; Sunyer, J.; Casas, M.; Martínez, D.; Ballester, F.; Basterechea, M.; Bonde, J.P.; Chatzi, L.; Chevrier, C.; Esplugues, A. Prenatal exposure to DDE and PCB 153 and respiratory health in early childhood: A meta-analysis. Epidemiology 2014, 25, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Gascon, M.; Sunyer, J.; Martínez, D.; Guerra, S.; Lavi, I.; Torrent, M.; Vrijheid, M. Persistent organic pollutants and children’s respiratory health: The role of cytokines and inflammatory biomarkers. Environ. Int. 2014, 69, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Sunyer, J.; Garcia-Esteban, R.; Alvarez, M.; Guxens, M.; Goñi, F.; Basterrechea, M.; Vrijheid, M.; Guerra, S.; Antó, J. DDE in mothers’ blood during pregnancy and lower respiratory tract infections in their infants. Epidemiology 2010, 21, 729–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Jung, K.H.; Horton, M.K.; Camann, D.E.; Liu, X.; Reardon, A.M.; Perzanowski, M.S.; Zhang, H.; Perera, F.P.; Whyatt, R.M.; et al. Prenatal exposure to pesticide ingredient piperonyl butoxide and childhood cough in an urban cohort. Environ. Int. 2012, 48, 156–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, L.G.; Richter, R.J.; Wan-Fen, L.; Cole, T.; Guizzetti, M.; Furlong, C.E. Paraoxonase (PON1) as a biomarker of susceptibility for organophosphate toxicity. Biomarkers 2003, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Harley, K.G.; Engel, S.M.; Vedar, M.G.; Eskenazi, B.; Whyatt, R.M.; Lanphear, B.P.; Bradman, A.; Rauh, V.A.; Yolton, K.; Hornung, R.W.; et al. Prenatal Exposure to Organophosphorous Pesticides and Fetal Growth: Pooled Results from Four Longitudinal Birth Cohort Studies. Environ. Health Perspect. 2016, 124, 1084–1092. [Google Scholar] [CrossRef]
- Harley, K.G.; Huen, K.; Schall, R.A.; Holland, N.T.; Bradman, A.; Barr, D.B.; Eskenazi, B. Association of organophosphate pesticide exposure and paraoxonase with birth outcome in Mexican-American women. PLoS ONE 2011, 6, e23923. [Google Scholar] [CrossRef] [Green Version]
- Naksen, W.; Prapamontol, T.; Mangklabruks, A.; Chantara, S.; Thavornyutikarn, P.; Srinual, N.; Panuwet, P.; Ryan, P.B.; Riederer, A.M.; Barr, D.B. Associations of maternal organophosphate pesticide exposure and PON1 activity with birth outcomes in SAWASDEE birth cohort, Thailand. Environ. Res. 2015, 142, 288–296. [Google Scholar] [CrossRef] [Green Version]
- Rauch, S.A.; Braun, J.M.; Barr, D.B.; Calafat, A.M.; Khoury, J.; Montesano, M.A.; Yolton, K.; Lanphear, B.P. Associations of Prenatal Exposure to Organophosphate Pesticide Metabolites with Gestational Age and Birth Weight. Environ. Health Perspect. 2012, 120, 1055–1060. [Google Scholar] [CrossRef] [Green Version]
- Banhela, N.; Naidoo, P.; Naidoo, S. Association between pesticide exposure and paraoxonase-1 (PON1) polymorphisms, and neurobehavioural outcomes in children: A systematic review. Syst. Rev. 2020, 9, 109. [Google Scholar] [CrossRef]
- Romieu, I.; Barraza-Villarreal, A.; Escamilla-Núñez, C.; Texcalac-Sangrador, J.L.; Hernandez-Cadena, L.; Díaz-Sánchez, D.; De Batlle, J.; Del Rio-Navarro, B.E. Dietary intake, lung function and airway inflammation in Mexico City school children exposed to air pollutants. Respir. Res. 2009, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, B.; Berthon, B.S.; Wark, P.; Wood, L.G. Effects of Fruit and Vegetable Consumption on Risk of Asthma, Wheezing and Immune Responses: A Systematic Review and Meta-Analysis. Nutrients 2017, 9, 341. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.M.; Kalloo, G.; Chen, A.; Dietrich, K.N.; Liddy-Hicks, S.; Morgan, S.; Xu, Y.; Yolton, K.; Lanphear, B.P. Cohort Profile: The Health Outcomes and Measures of the Environment (HOME) study. Int. J. Epidemiol. 2016, 46, 24. [Google Scholar] [CrossRef] [Green Version]
- Braun, J.M.; Chen, A.; Romano, M.E.; Calafat, A.M.; Webster, G.M.; Yolton, K.; Lanphear, B.P. Prenatal perfluoroalkyl substance exposure and child adiposity at 8 years of age: The HOME study. Obesity 2016, 24, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yolton, K.; Xu, Y.; Sucharew, H.; Succop, P.; Altaye, M.; Popelar, A.; Montesano, M.A.; Calafat, A.M.; Khoury, J.C. Impact of low-level gestational exposure to organophosphate pesticides on neurobehavior in early infancy: A prospective study. Environ. Health 2013, 12, 79. [Google Scholar] [CrossRef] [Green Version]
- Bravo, R.; Caltabiano, L.M.; Weerasekera, G.; Whitehead, R.D.; Fernandez, C.; Needham, L.L.; Bradman, A.; Barr, D.B. Measurement of dialkyl phosphate metabolites of organophosphorus pesticides in human urine using lyophilization with gas chromatography-tandem mass spectrometry and isotope dilution quantification. J. Expo. Sci. Environ. Epidemiol. 2004, 14, 249–259. [Google Scholar] [CrossRef] [Green Version]
- National Center for Health Statistics. Plan and operation of the Third National Health and Nutrition Examination Survey, 1988–1994. Series 1: Programs and collection procedures. Vital Health Stat. 1994, 32, 1–407. [Google Scholar]
- Miller, M.R.; Hankinson, J.A.T.S.; Brusasco, V.; Burgos, F.; Casaburi, R.; Coates, A.; Crapo, R.; Enright, P.; Van Der Grinten, C.P.M.; Gustafsson, P.; et al. Standardisation of spirometry. Eur. Respir. J. 2005, 26, 319–338. [Google Scholar] [CrossRef] [Green Version]
- Spanier, A.J.; Kahn, R.S.; Kunselman, A.R.; Schaefer, E.W.; Hornung, R.; Xu, Y.; Calafat, A.M.; Lanphear, B.P. Bisphenol a exposure and the development of wheeze and lung function in children through age 5 years. JAMA Pediatr. 2014, 168, 1131–1137. [Google Scholar] [CrossRef]
- Bernert, J.T., Jr.; McGuffey, J.E.; Morrison, M.A.; Pirkle, J.L. Comparison of serum and salivary cotinine measurements by a sensitive high-performance liquid chromatography-tandem mass spectrometry method as an indicator of exposure to tobacco smoke among smokers and nonsmokers. J. Anal. Toxicol. 2000, 24, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Bernert, J.T.; Jacob, P., 3rd; Holiday, D.B.; Benowitz, N.L.; Sosnoff, C.S.; Doig, M.V.; Feyerabend, C.; Aldous, K.M.; Sharifi, M.; Kellogg, M.D.; et al. Interlaboratory comparability of serum cotinine measurements at smoker and nonsmoker concentration levels: A round-robin study. Nicotine Tob. Res. 2009, 11, 1458–1466. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.M.; Yolton, K.; Dietrich, K.N.; Hornung, R.; Ye, X.; Calafat, A.M.; Lanphear, B.P. Prenatal bisphenol A exposure and early childhood behavior. Environ. Health Perspect. 2009, 117, 1945–1952. [Google Scholar] [CrossRef] [PubMed]
- Nagin, D.S.; Jones, B.L.; Passos, V.L.; Tremblay, R.E. Group-based multi-trajectory modeling. Stat. Methods Med. Res. 2018, 27, 2015–2023. [Google Scholar] [CrossRef]
- Donauer, S.; Altaye, M.; Xu, Y.; Sucharew, H.; Succop, P.; Calafat, A.M.; Khoury, J.C.; Lanphear, B.; Yolton, K. An Observational Study to Evaluate Associations Between Low-Level Gestational Exposure to Organophosphate Pesticides and Cognition During Early Childhood. Am. J. Epidemiol. 2016, 184, 410–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, C.L.; Al-Batanony, M.; Ismail, A.A.; Abdel-Rasoul, G.; Hendy, O.; Olson, J.R.; Rohlman, D.S.; Bonner, M.R. Chlorpyrifos exposure and respiratory health among adolescent agricultural workers. Int. J. Environ. Res. Public Health 2014, 11, 13117–13129. [Google Scholar] [CrossRef] [PubMed]
- Seo, T.; Pahwa, P.; McDuffie, H.H.; Nakada, N.; Goto, S.; Ghosh, S.; Nakagawa, K. Interactive Effect of Paraoxonase-1 Q192R Polymorphism and Smoking History on the Lung Function Decline in Grain Workers. Ann. Epidemiol. 2008, 18, 330–334. [Google Scholar] [CrossRef]
- Martinez, F.D.; Wright, A.L.; Taussig, L.M.; Holberg, C.J.; Halonen, M.; Morgan, W.J. Asthma and wheezing in the first six years of life. The Group Health Medical Associates. N. Engl. J. Med. 1995, 332, 133–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garfitt, S.J.; Jones, K.; Mason, H.J.; Cocker, J. Exposure to the organophosphate diazinon: Data from a human volunteer study with oral and dermal doses. Toxicol. Lett. 2002, 134, 105–113. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wheeze (n = 205) | No Wheeze (n = 162) | All (n = 367) | ||||
|---|---|---|---|---|---|---|
| n | % | n | % | na | % | |
| Mother | ||||||
| Race | ||||||
| White, non-Hispanic | 135 | 57.4 | 100 | 42.6 | 235 | 64.2 |
| Others | 69 | 52.7 | 62 | 47.3 | 131 | 35.8 |
| Education at Delivery | ||||||
| Bachelor’s degree | 4 | 100.0 | 0 | 0.0 | 4 | 2.1 |
| Some college or 2 year degree | 59 | 54.1 | 50 | 45.9 | 109 | 56.8 |
| Some high School (Grades 9–12) | 39 | 49.4 | 40 | 50.6 | 79 | 41.1 |
| Insurance at Delivery | ||||||
| Private | 150 | 56.4 | 116 | 43.6 | 266 | 72.7 |
| Public/uninsured | 54 | 54.0 | 46 | 46.0 | 100 | 27.3 |
| Marital Status at Delivery | ||||||
| Married | 133 | 55.6 | 106 | 44.4 | 239 | 65.3 |
| Not Married | 69 | 54.3 | 58 | 45.7 | 127 | 34.7 |
| Employment at Delivery | ||||||
| No | 37 | 54.4 | 31 | 45.6 | 68 | 18.6 |
| Yes | 167 | 56.0 | 131 | 44.0 | 298 | 81.4 |
| Household Income at Delivery | ||||||
| Less than USD 20,000 | 44 | 55.7 | 35 | 44.3 | 79 | 21.5 |
| USD 20,000–USD 40,000 | 37 | 61.7 | 23 | 38.3 | 60 | 16.3 |
| Greater than USD 40,000 | 124 | 54.4 | 104 | 45.6 | 228 | 62.1 |
| Fruit/Veg Consumption During Pregnancy | ||||||
| Less than daily | 38 | 52.1 | 35 | 47.9 | 73 | 19.9 |
| Daily | 166 | 56.7 | 127 | 43.3 | 293 | 80.1 |
| Parity | ||||||
| 0 | 86 | 53.1 | 76 | 46.9 | 162 | 44.3 |
| 1 | 67 | 57.3 | 50 | 42.7 | 117 | 32.0 |
| >1 | 51 | 58.6 | 36 | 41.4 | 87 | 23.8 |
| Age at Delivery (y, mean ± SD) | 29.3 +/− 5.8 | 29.8 +/− 5.8 | 29.3 +/− 5.8 | |||
| Maternal Cotinine Levels at 16W (ng/mL mean ± SD) | 11.6 +/− 54.7 | 7.7 +/− 27.1 | 9.5 +/− 41.8 | |||
| PON1−108 | ||||||
| CC b | 81 | 57.9 | 59 | 42.1 | 140 | 46.1 |
| CT | 67 | 53.6 | 58 | 46.4 | 125 | 41.1 |
| TT | 19 | 48.7 | 20 | 51.3 | 39 | 12.8 |
| PON1192 | ||||||
| 64 | 57.7 | 47 | 42.3 | 111 | 34.8 | |
| QR | 75 | 56.8 | 57 | 43.2 | 132 | 41.4 |
| RR | 36 | 47.4 | 40 | 52.6 | 76 | 23.8 |
| Child | ||||||
| Race | ||||||
| White, non-Hispanic | 128 | 56.6 | 98 | 43.4 | 226 | 61.7 |
| Others | 76 | 54.3 | 64 | 45.7 | 140 | 38.3 |
| Sex | ||||||
| Female | 109 | 54.8 | 90 | 45.2 | 199 | 54.4 |
| Male | 95 | 56.9 | 72 | 43.1 | 167 | 45.6 |
| Birth weight (g, mean ± SD) | 3390.5 ± 644.7 | 3381.1 ± 569.2 | 3359.8 ± 627.4 | |||
| Birth length (cm, mean ± SD) | 50.9 ± 3.1 | 51.0 ± 2.7 | 50.8 ± 3.0 | |||
| All (n = 367) | Non-white (n = 131) | White (n = 235) | Difference between Non-White and White | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Geometric Mean | Min | Max | Geometric Mean | Min | Max | Geometric Mean | p-value a | |
| 3PBA 16W (ng/mL) | 0.00 | 41.7 | 0.4 | 0.03 | 41.7 | 0.5 | 0.02 | 31.0 | 0.3 | 0.005 |
| 3PBA 26W (ng/mL) | 0.00 | 37.4 | 0.3 | 0.03 | 32.2 | 0.4 | 0.02 | 37.4 | 0.3 | 0.002 |
| DE 16W (nmol/L) | 0.01 | 383.7 | 47.9 | 0.09 | 289.4 | 10.6 | 0.07 | 383.7 | 9.8 | 0.677 |
| DE 26W (nmol/L) | 0.10 | 594.8 | 28.7 | 0.08 | 594.8 | 6.2 | 0.05 | 231.0 | 5.8 | 0.773 |
| DM 16W (nmol/L) | 0.10 | 6945.9 | 45.2 | 0.38 | 4908.5 | 43.1 | 0.10 | 6945.8 | 46.6 | 0.673 |
| DM 26W (nmol/L) | 0.10 | 7299.3 | 33.0 | 0.72 | 7299.3 | 46.4 | 0.09 | 1606.5 | 26.5 | 0.003 |
| DAP 16W (nmol/L) | 1.30 | 7021.3 | 72.6 | 1.64 | 4921.8 | 72.6 | 1.26 | 7021.3 | 72.5 | 0.991 |
| DAP 26W (nmol/L) | 0.30 | 7894.1 | 53.2 | 1.38 | 7894.0 | 69.5 | 0.34 | 1739.1 | 44.8 | 0.008 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilden, R.; Friedmann, E.; Holmes, K.; Yolton, K.; Xu, Y.; Lanphear, B.; Chen, A.; Braun, J.; Spanier, A. Gestational Pesticide Exposure and Child Respiratory Health. Int. J. Environ. Res. Public Health 2020, 17, 7165. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17197165
Gilden R, Friedmann E, Holmes K, Yolton K, Xu Y, Lanphear B, Chen A, Braun J, Spanier A. Gestational Pesticide Exposure and Child Respiratory Health. International Journal of Environmental Research and Public Health. 2020; 17(19):7165. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17197165
Chicago/Turabian StyleGilden, Robyn, Erika Friedmann, Katie Holmes, Kimberly Yolton, Yingying Xu, Bruce Lanphear, Aimin Chen, Joseph Braun, and Adam Spanier. 2020. "Gestational Pesticide Exposure and Child Respiratory Health" International Journal of Environmental Research and Public Health 17, no. 19: 7165. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17197165