Prenatal Exposure to Gutkha, a Globally Relevant Smokeless Tobacco Product, Induces Hepatic Changes in Adult Mice

 , ,
, ,
Abstract
:1. Introduction
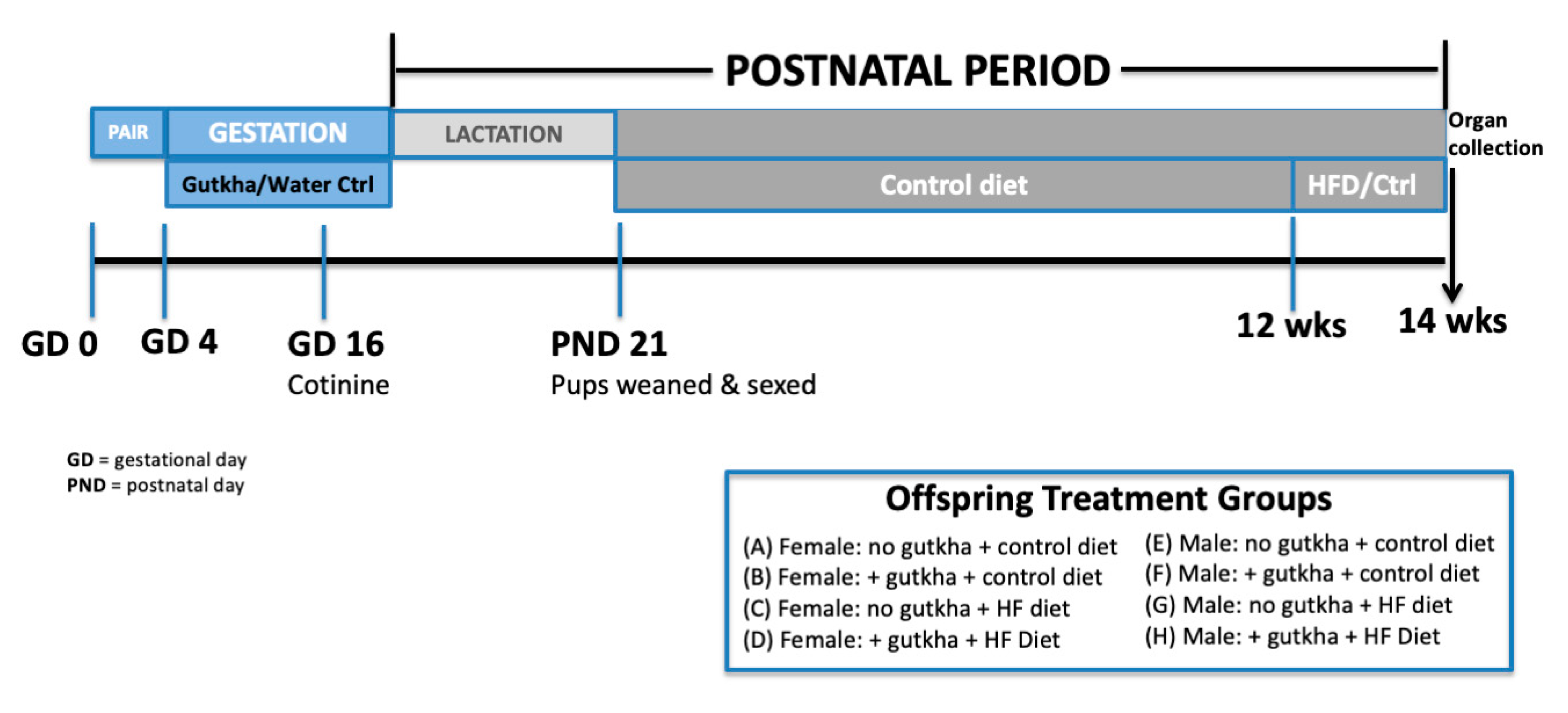
2. Materials and Methods
2.1. Animals
2.2. Breeding and Gutkha Preparation/Exposure
2.3. Cotinine Measurements
2.4. High-Fat Diet
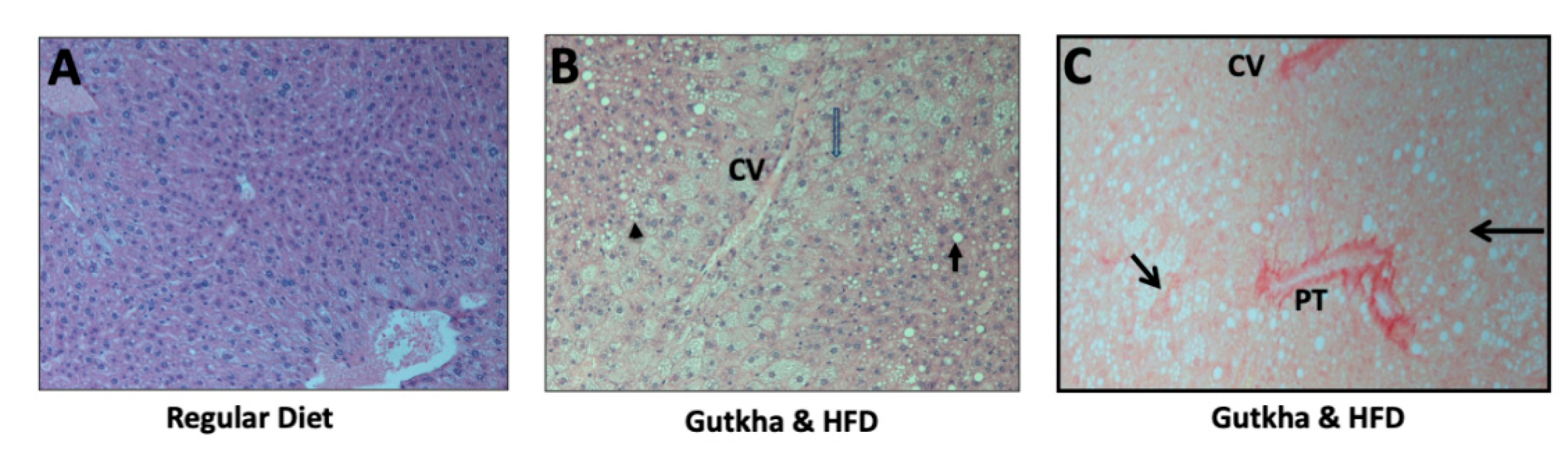
2.5. Histopathology
2.6. Semi-Quantitative RT-PCR
2.7. Plasma Lipid Analyses
2.8. Statistics
3. Results
3.1. Cotinine Measurements
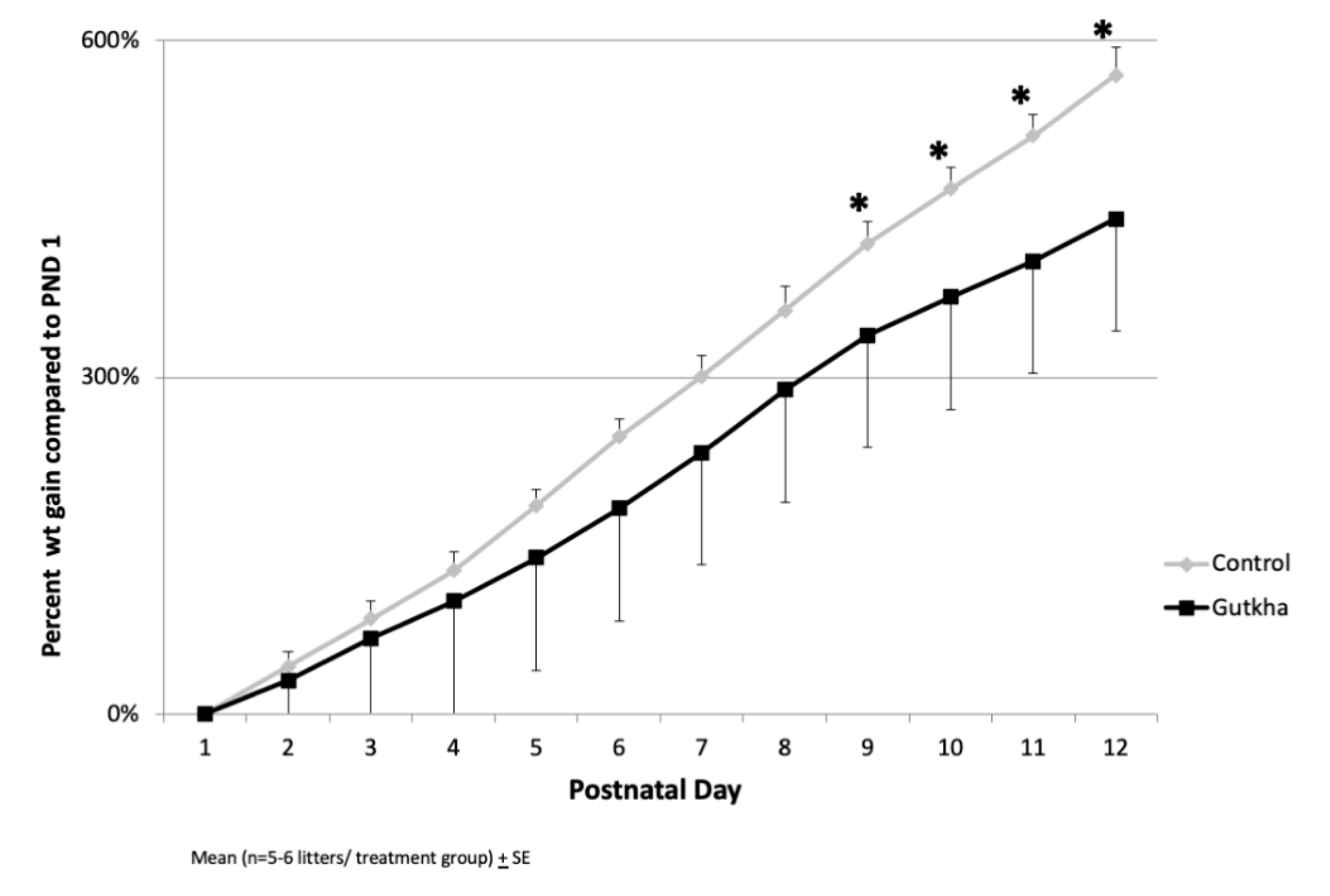
3.2. Gestational Parameters
3.3. Offspring Body and Organ Weights
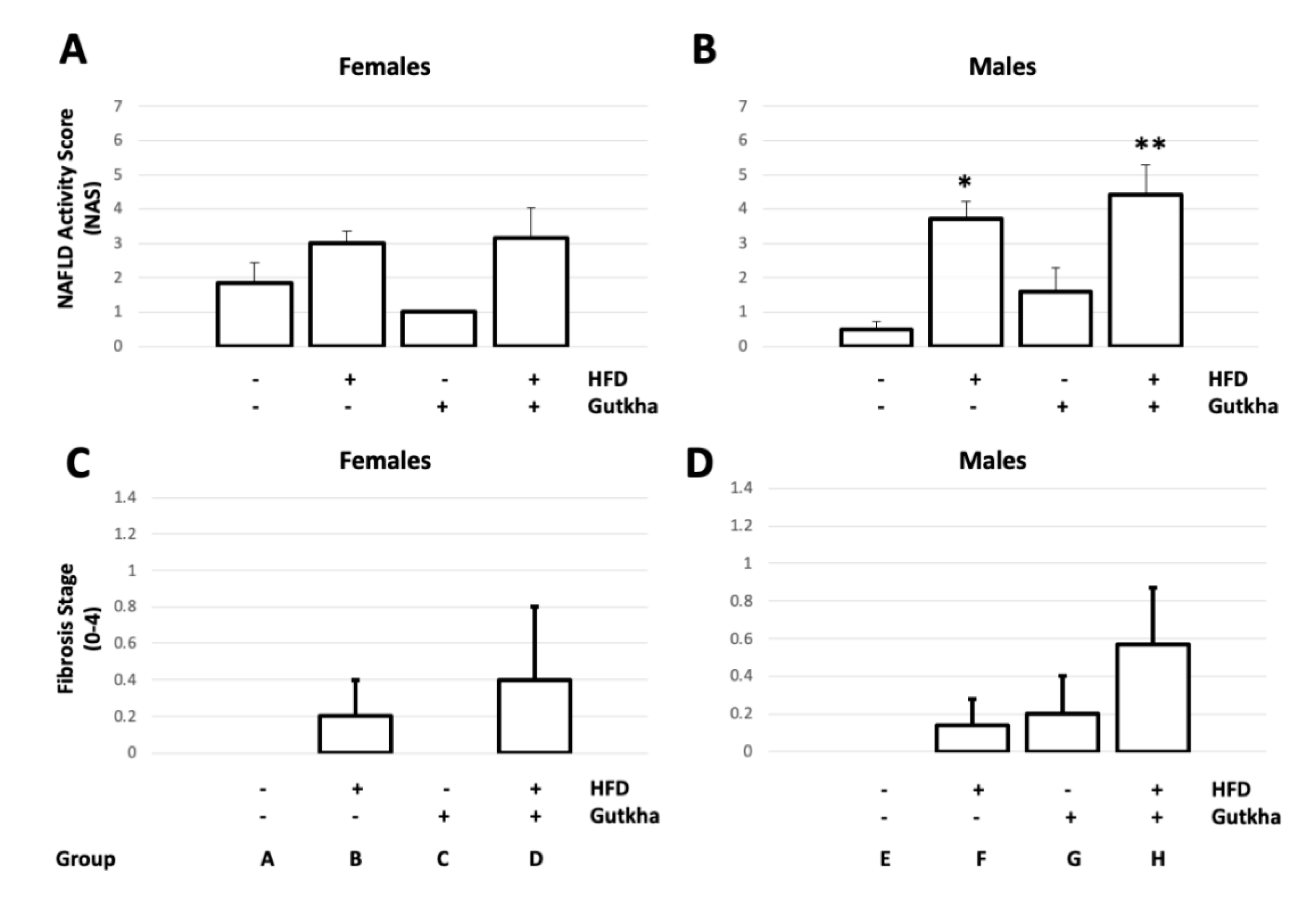
3.4. Adult Offspring Liver Histopathology
3.5. Blood Biochemistry
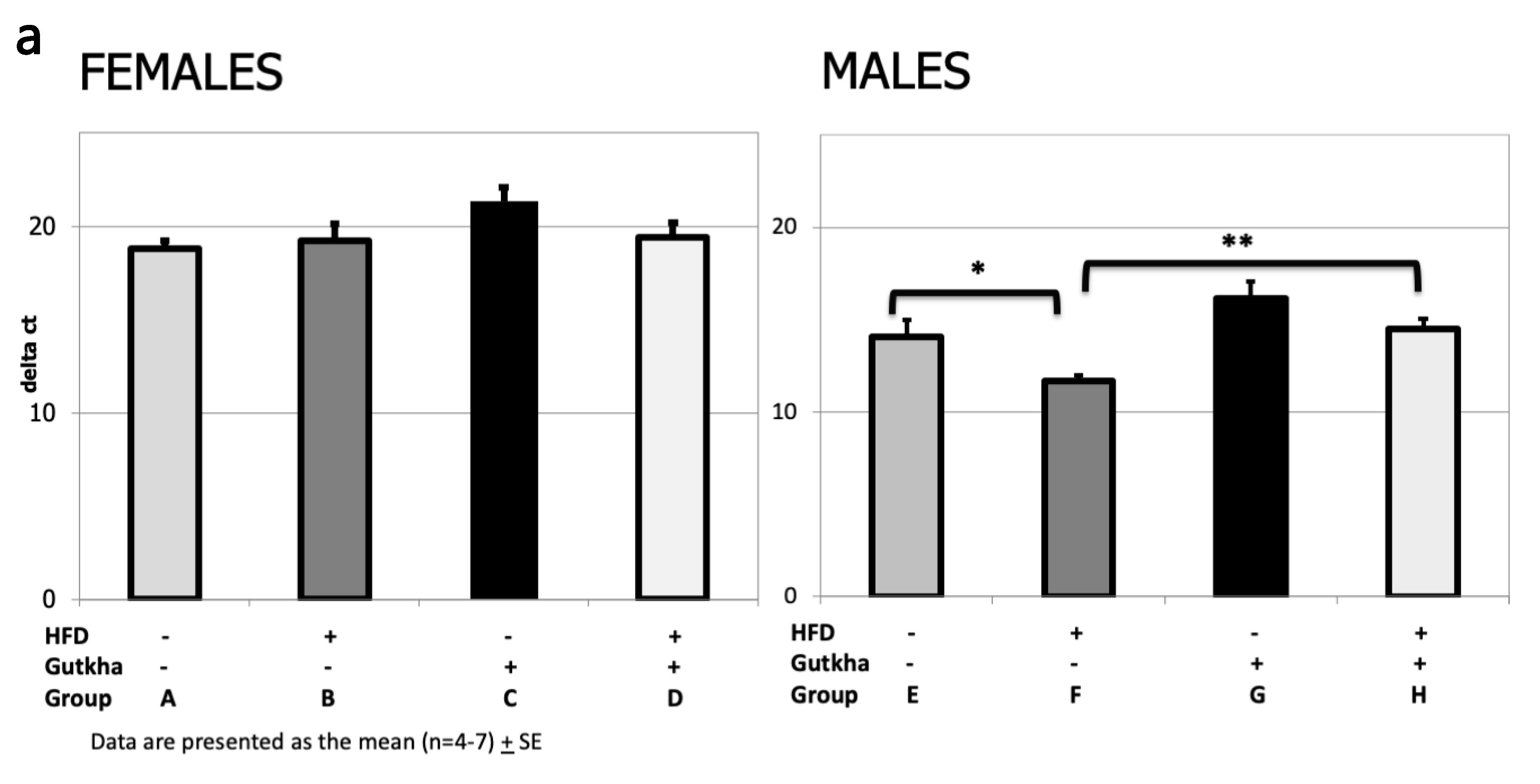
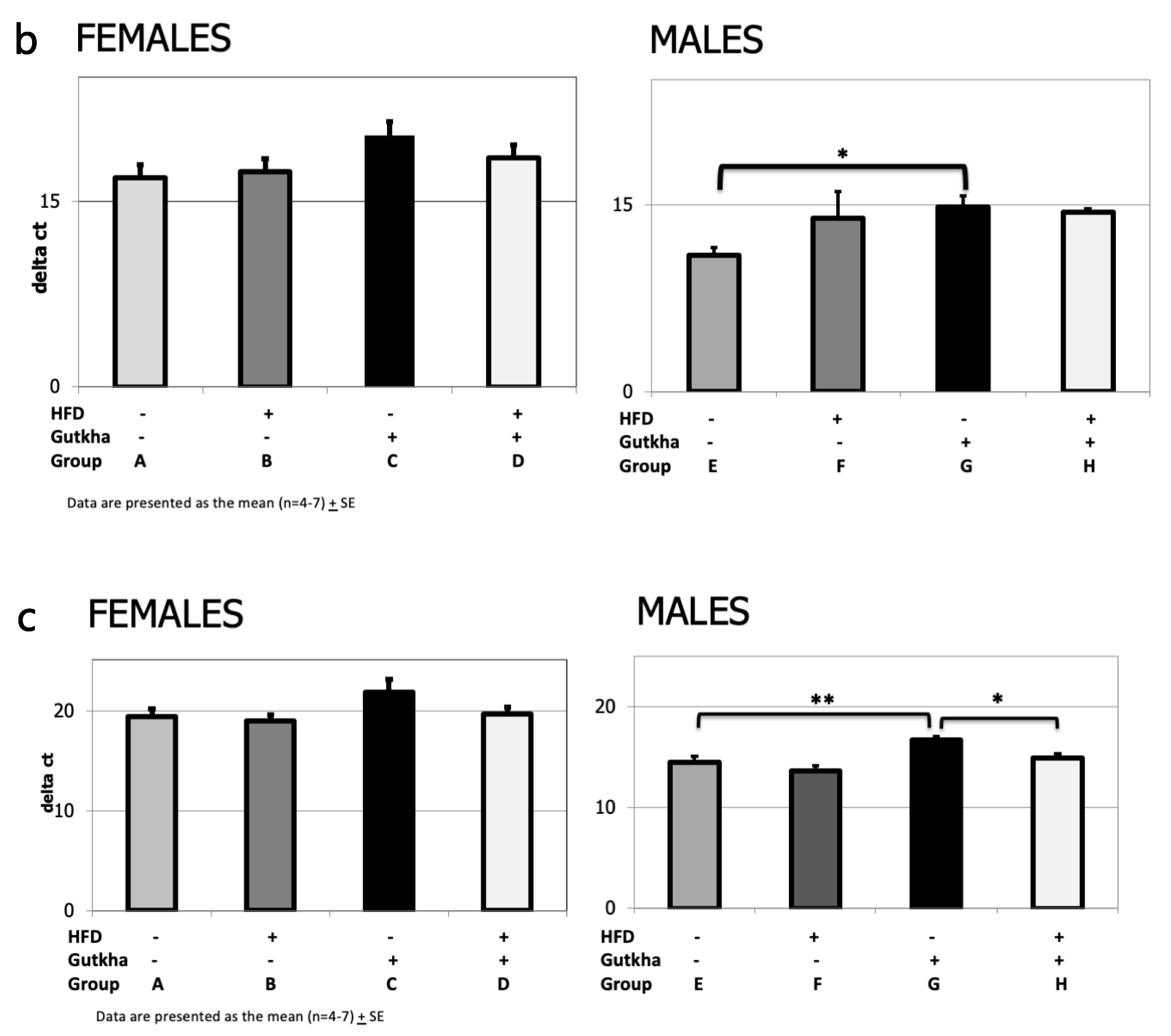
3.6. Pro-Fibrotic Cytokine (IL-6 and IL-1b) and Collagen (Col1A) mRNA Expression
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashi, T.; Friedman, S.L.; Hoshida, Y. Hepatic stellate cells as key target in liver fibrosis. Adv. Drug Deliv. Rev. 2017, 121, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Allina, J.; Grabowski, J.; Doherty-Lyons, S.; Fiel, M.I.; Jackson, C.E.; Zelikoff, J.T.; Odin, J.A. Maternal allergy acts synergistically with cigarette smoke exposure during pregnancy to induce hepatic fibrosis in adult male offspring. J. Immunotoxicol. 2011, 8, 258–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manderski, M.T.; Steinberg, M.B.; Rahi, K.N.; Banerjee, S.C.; Delnevo, C.D. Surveillance of Tobacco Use Among South Asians in the US: Are We Underestimating Prevalence? J. Community Health 2016, 41, 1140–1145. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.N.; Yel, D.; Sin, S.; Khieng, S.; Lopez, J.; Job, J.; Ferry, L.; Knutsen, S. Tobacco use among adults in Cambodia: Evidence for a tobacco epidemic among women. Bull. World Health Organ. 2009, 87, 905–912. [Google Scholar] [CrossRef]
- Singh, P.N.; Eng, C.; Yel, D.; Kheam, T.; Job, J.S.; Kanal, K. Maternal use of cigarettes, pipes, and smokeless tobacco associated with higher infant mortality rates in Cambodia. Asia Pac. J. Public Health 2013, 25, 64S–74S. [Google Scholar] [CrossRef] [Green Version]
- Vismita, G. 90% of Smokeless Tobacco Users Live in South-East Asia; SEAR/PR 1563; WHO S-E Asia: New Delhi, India, 2013. [Google Scholar]
- Nair, S.; Schensul, J.J.; Begum, S.; Begum, S.; Pednekar, M.S.; Oncken, C.; Bilgi, S.M.; Pasi, A.R.; Donta, B. Use of smokeless tobacco by Indian women aged 18–40 years during pregnancy and reproductive years. PLoS ONE 2015, 10, e0119814. [Google Scholar] [CrossRef]
- Barakoti, R.; Ghimire, A.; Pandey, A.R.; Baral, D.D.; Pokharel, P.K. Tobacco Use during Pregnancy and Its Associated Factors in a Mountain District of Eastern Nepal: A Cross-Sectional Questionnaire Survey. Front. Public Health 2017, 6, 129. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.C.; Sreevidya, S. Smokeless tobacco use, birth weight, and gestational age: Population based, prospective cohort study of 1217 women in Mumbai, India. BMJ 2004, 328, 1538. [Google Scholar] [CrossRef] [Green Version]
- El-Zayadi, A.R. Heavy smoking and liver. World J. Gastroenterol. 2006, 12, 6098–6101. [Google Scholar] [CrossRef]
- Mantaka, A.; Koulentaki, M.; Samonakis, D.; Sifaki, S.P.; Voumvouraki, A.; Tzardi, M.; Kouroumalis. Association of smoking with liver fibrosis and mortality in primary biliary cholangitis. Eur. J. Gastroenterol. Hepatol. 2018, 30, 1461–1469. [Google Scholar] [CrossRef] [PubMed]
- Capurso, G.; Lahner, E. The interaction between smoking, alcohol and the gut microbiome. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Mathur, A.K.; Acker, W.B., II; Al-Holou, S.N.; Ehrlichman, L.K.; Lewin, S.A.; Nguyen, C.K.; Peterson, S.F.; Ranney, D.N.; Sell, K.; et al. Effects of smoking on survival for patients with end-stage liver disease. J. Am. Coll. Surg. 2009, 208, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Santiago, S.E.; Huffman, K.J. Postnatal effects of prenatal nicotine exposure on body weight, brain size and cortical connectivity in mice. Neurosci. Res. 2012, 73, 282–291. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Betel-quid and areca-nut chewing and some areca-nut derived nitrosamines. IARC Monogr. Eval. Carcinog. Risks Hum. 2004, 85, 1–334. [Google Scholar] [PubMed]
- McNeill, A.; Bedi, R.; Islam, S.; Alkhatib, M.N.; West, R. Levels of toxins in oral tobacco products in the UK. Tob. Control 2006, 15, 64–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, D.N.; Popovech, M.A.; Gany, F.; Hoffman, C.; Blum, J.L.; Zelikoff, J.T. Toxicity of gutkha, a smokeless tobacco product gone global: Is there more to the toxicity than nicotine? Int. J. Environ. Res. Public Health 2014, 9, 919–933. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Chen, X.; Ward, S.C.; Liu, Y.; Ouedraogo, Y.; Xu, C.; Cederbaum, A.I.; Lu, Y. CYP2A6 is associated with obesity: Studies in human samples and a high fat diet mouse model. Int. J. Obes. 2019, 43, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, Q.; Li, X.; Zhang, X. Experimental nonalcoholic fatty liver disease in mice leads to cytochrome p450 2a5 upregulation through nuclear factor erythroid 2-like 2 translocation. Redox Biol. 2013, 24, 433–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avti, P.K.; Kumar, S.; Pathak, C.M.; Vaiphei, K.; Khanduja, K.L. Smokeless Tobacco Impairs the Antioxidant Defense in Liver, Lung, and Kidney of Rats. Toxicol. Sci. 2006, 89, 547–553. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.H.; Fiel, M.I.; Sun, Q.; Guo, J.; Gordon, R.E.; Chen, L.C.; Friedman, S.L.; Odin, J.A.; Allina, J. Kupffer cell activation by ambient air particulate matter exposure may exacerbate non-alcoholic fatty liver disease. J. Immunotoxicol. 2009, 6, 266–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Blum, J.L.; Edwards, J.R.; Prozialeck, W.C.; Xiong, J.Q.; Zelikoff, J.T. Effects of Maternal Exposure to Cadmium Oxide Nanoparticles During Pregnancy on Maternal and Offspring Kidney Injury Markers Using a Murine Model. J. Toxicol. Environ. Health A 2015, 78, 711–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, S.P.; Conklin, D.J.; Bhatnagar, A.; Bolanowski, D.D.; Lyon, J.; Zelikoff, J.T. Prenatal exposure to cigarette smoke induces diet- and sex-dependent dyslipidemia and weight gain in adult murine offspring. Environ. Health Perspect. 2009, 117, 1042–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunt, E.M.; Kleiner, D.E.; Wilson, L.A.; Belt, P.; Neuschwander-Tetri, B.A. Nonalcoholic fatty liver disease (NAFLD) activity score and the histopathologic Diagnosis of NAFLD: Distinct clinicopathologic meanings. Hepatology 2011, 53, 810–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Gào, X.; Zhang, Y.; Breitling, L.P.; Schöttker, B.; Brenner, H. Associations of self-reported smoking, cotinine levels and epigenetic smoking indicators with oxidative stress among older adults: A population-based study. Eur. J. Epidemiol. 2017, 32, 443–456. [Google Scholar] [CrossRef]
- Gupta, R.; van Dongen, J.; Fu, Y.; Abdellaoui, A.; Tyndale, R.F.; Velagapudi, V.; Boomsma, D.I.; Korhonen, T.; Kaprio, J.; Loukola, A.; et al. Epigenome-wide association study of serum cotinine in current smokers reveals novel genetically driven loci. Clin. Epigenetics 2019, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Nagata, K.; Yamazoe, Y. Genetic polymorphism of human cytochrome p450 involved in drug metabolism. Drug Metab. Pharmacokinet. 2002, 17, 167–189. [Google Scholar] [CrossRef] [PubMed]
- Niaz, K.; Maqbool, F.; Khan, F.; Bahadar, H.; Hassan, F.I.; Abdollahi, M. Smokeless tobacco (paan and gutkha) consumption, prevalence, and contribution to oral cancer. Epidemiol. Health 2017, 39, e2017009. [Google Scholar] [CrossRef] [PubMed]
- Wickstrom, R. Effects of Nicotine during pregnancy: Human and experimental evidence. Curr. Neuropharmacol. 2007, 5, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Fan, F.; McCarthy, D.M.; Zhang, L.; Cannon, E.N.; Spencer, T.J.; Biederman, J.; Bhide, P.G. A prenatal nicotine exposure mouse model of methylphenidate responsive ADHD-associated cognitive phenotypes. Int. J. Dev. Neurosci. 2017, 58, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Quelhas, D.; Kompala, C.; Wittenbrink, B.; Han, Z.; Parker, M.; Shapiro, M.; Downs, S.; Kraemer, K.; Fanzo, J.; Morris, S.; et al. The association between active tobacco use during pregnancy and growth outcomes of children under five years of age: A systematic review and meta-analysis. BMC Public Health 2018, 18, 1372. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, Y.; Friso, S.; Choi, S.W. Epigenetics in non-alcoholic fatty liver disease. Mol. Asp. Med. 2017, 54, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Podrini, C.; Borghesan, M.; Greco, A.; Pazienza, V.; Mazzoccoli, G.; Vinciguerra, M. Redox homeostasis and epigenetics in non-alcoholic fatty liver disease (NAFLD). Curr. Pharm. Des. 2013, 19, 2737–2746. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, H.; Long, W.; Shen, X.; Wu, D.; Song, S.; Sun, Y.M.; Liu, P.X.; Fan, S.; Fan, F.; et al. Sex differences in the toxicity of polyethylene glycol-coated gold nanoparticles in mice. Int. J. Nanomed. 2013, 8, 2409–2419. [Google Scholar] [PubMed] [Green Version]
- Mrdjen, I.; Morse, M.A.; Ruch, R.J.; Knobloch, T.J.; Choudhary, S.; Weghorst, C.M.; Lee, J. Impact of Microcystin-LR on Liver Function Varies by Dose and Sex in Mice. Toxins 2018, 10, 435. [Google Scholar] [CrossRef] [Green Version]
- Thangavel, C.; Boopathi, E.; Shapiro, B.H. Inherent sex-dependent regulation of human hepatic CYP3A5. Br. J. Pharmacol. 2013, 168, 988–1000. [Google Scholar] [CrossRef] [Green Version]
- Uno, Y.; Takata, R.; Kito, G.; Yamazaki, H.; Nakagawa, K.; Nakamura, Y.; Kamataki, T.; Katagiri, T. Sex- and age-dependent gene expression in human liver: An implication for drug-metabolizing enzymes. Drug Metab. Pharmacokinet. 2017, 32, 100–107. [Google Scholar] [CrossRef]
- Fuscoe, J.C.; Vijay, V.; Hanig, J.P.; Han, T.; Ren, L.; Greenhaw, J.J.; Beger, R.D.; Pence, L.M.; Shi, Q. Hepatic Transcript Profiles of Cytochrome P450 Genes Predict Sex Differences in Drug Metabolism. Drug Metab. Dispos. 2020, 48, 447–458. [Google Scholar] [CrossRef]
- Filis, P.; Nagrath, N.; Fraser, M.; Hay, D.C.; Iredale, J.P.; O’Shaughnessy, P.; Fowler, P.A. Maternal Smoking Dysregulates Protein Expression in Second Trimester Human Fetal Livers in a Sex-Specific Manner. J. Clin. Endocrinol. Metab. 2015, 100, E861–E870. [Google Scholar] [CrossRef] [Green Version]
- Banderali, G.; Martelli, A.; Landi, M.; Moretti, F.; Betti, F.; Radaelli, G.; Lassandro, C.; Verduci, E. Short and long term health effects of parental tobacco smoking during pregnancy and lactation: A descriptive review. J. Transl. Med. 2015, 13, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaddoe, V.W.V.; de Jonge, L.L.; van Dam, R.M.; Willett, W.C.; Harris, H.; Stampfer, M.J.; Hu, F.B.; Michels, K.B. Fetal exposure to parental smoking and the risk of type 2 diabetes in adult women. Diabetes Care 2014, 37, 2966–2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haghighi, A.; Schwartz, D.H.; Abrahamowicz, M.; Leonard, G.T.; Perron, M.; Richer, L.; Veillette, S.; Gaudet, D.; Paus, T.; Pausova, Z. Prenatal Exposure to Maternal Cigarette Smoking, Amygdala Volume, and Fat Intake in Adolescence. JAMA Psychiatry 2013, 70, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Dai, M.; Bi, Y.; Xu, M.; Xu, Y.; Li, M.; Wang, T.; Huang, F.; Xu, B.; Zhang, J.; et al. Active Smoking, Passive Smoking, and Risk of Nonalcoholic Fatty Liver Disease (NAFLD): A Population-Based Study in China. J. Epidemiol. 2013, 23, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, L.C.; Azen, C.; Hobel, C.J.; Platt, L.D. Maternal tobacco use and substance abuse: Reported prevalence rates and associations with the delivery of small for gestational age neonates. Obstet. Gynecol. 1993, 81, 396–401. [Google Scholar]
- Mao, C.; Yuan, X.; Cui, Y.; Li, H.; Lv, J.; Feng, X.; Liu, Y.; Chen, L.; Xu, Z. Prenatal exposure to nicotine with associated in utero hypoxia decreased fetal brain muscarinic mRNA in the rat. Brain Res. 2008, 1189, 43–50. [Google Scholar] [CrossRef]
- Gentili, S.; Morrison, J.L.; McMillen, I.C. Intrauterine Growth Restriction and Differential Patterns of Hepatic Growth and Expression of IGF1, PCK2, and HSDL1 mRNA in the Sheep Fetus in Late Gestation. Biol. Reprod. 2009, 6, 1121–1127. [Google Scholar] [CrossRef] [Green Version]
- Thorn, S.R.; Regnault, T.R.; Brown, L.D.; Rozance, P.J.; Keng, J.; Roper, M.; Wilkening, R.B.; Hay, J.W.W.; Friedman, J.E. Intrauterine Growth Restriction Increases Fetal Hepatic Gluconeogenic Capacity and Reduces Messenger Ribonucleic Acid Translation Initiation and Nutrient Sensing in Fetal Liver and Skeletal Muscle. Endocrinology 2009, 150, 3021–3030. [Google Scholar] [CrossRef]
- Kim, S. Overview of Cotinine Cutoff Values for Smoking Status Classification. Int. J. Environ. Res. Public Health 2016, 13, 1236. [Google Scholar] [CrossRef]
- Center for Disease Control (CDC). Biomonitoring Summary: Cotinine, CAS No. 486-56-6; Metabolite of Nicotine (a Component of TOBACCO Smoke). 2017. Available online: https://www.cdc.gov/biomonitoring/Cotinine_BiomonitoringSummary.html (accessed on 15 October 2020).
- Dewar, B.J.; Bradford, B.U.; Thurman, R.G. Nicotine Increases Hepatic Oxygen Uptake in the Isolated Perfused Rat Liver by Inhibiting Glycolysis. J. Pharm. Exp. Ther. 2002, 301, 930–937. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Hanawa, N.; Saberi, B.; Kaplowitz, N. Mechanisms of liver injury. III. Role of glutathione redox status in liver injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G1–G7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, J.K.C.; Zhang, X.; Yu, J. Animal models of non-alcoholic fatty liver disease: Current perspectives and recent advances. J. Pathol. 2017, 241, 36–44. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Sex | Gutkha | Diet | Average Absolute Wt Gain a |
|---|---|---|---|---|
| (grams) | ||||
| A | F | No | CTRL | 0.80* ± 0.52 |
| B | F | Yes | CTRL | 0.33 ± 0.44 |
| C | F | No | HF | 2.83* ± 0.47 |
| D | F | Yes | HF | 1.75 ± 0.88 |
| E | M | No | CTRL | −0.39 ± 0.69 |
| F | M | Yes | CTRL | 0.70* ± 0.52 |
| G | M | No | HF | 0.82* ± 0.67 |
| H | M | Yes | HF | 3.08 ± 0.70 |
| Group | Sex | Gutkha | Diet | ALT | AST | T. Chol | TRIG | HDL | LDL | HDL/LDL Ratio |
|---|---|---|---|---|---|---|---|---|---|---|
| A | F | No | CTRL | 16.6* ± 0.7 | 45.3 ± 5.6 | 115.7 ± 13.5 | 23.0*± 6.8 | 84.5*± 10.0 | 14.7 ± 0.9 | 5.8 ± 0.6 |
| B | F | Yes | CTRL | 23.5* ± 2.6 | 52.1 ± 3.5 | 82.7 ± 9.9 | 8.5*± 1.8 | 54.5*± 4.9 | 12.4 ± 1.0 | 4.9 ± 0.5 |
| C | F | No | HF | 24.0 ± 1.3 | 47.6* ± 3.8 | 109.1 ± 16.0 | 20.4 ± 3.4 | 64.8 ± 5.9 | 18.7 ± 1.6 | 4.3 ± 0.3 |
| D | F | Yes | HF | 25.4 ± 2.3 | 62.5* ± 5.6 | 106.1 ± 3.1 | 19.9 ± 3.1 | 74.3 ± 3.4 | 18.7 ± 1.7 | 4.4 ± 0.4 |
| E | M | No | CTRL | 21.7 ± 2.5 | 32.2 ± 3.4 | 142.2 ± 9.8 | 45.5* ± 7.1 | 102.1 ± 7.6 | 16.1 ± 0.7 | 6.3 ± 0.4 |
| F | M | Yes | CTRL | 20.7 ± 1.8 | 28.0* ± 2.9 | 150.7 ± 14.8 | 23.4* ± 3.1 | 112.2 ± 8.8 | 16.7 ± 2.8 | 7.0* ± 0.5 |
| G | M | No | HF | 27.9 ±1.9 | 41.9 ± 3.3 | 175.1 ± 10.7 | 60.3 ± 8.0 | 124.6 ± 7.1 | 20.8 ± 1.1 | 6.1 ± 0.41 |
| H | M | Yes | HF | 28.3 ±3.0 | 43.3* ± 2.0 | 191.0 ± 8.3 | 48.4 ± 5.7 | 136.5 ± 6.1 | 25.0 ± 0.9 | 5.5* ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doherty Lyons, S.; Blum, J.L.; Hoffman-Budde, C.; Tijerina, P.B.; Fiel, M.I.; J. Conklin, D.; Gany, F.; Odin, J.A.; Zelikoff, J.T. Prenatal Exposure to Gutkha, a Globally Relevant Smokeless Tobacco Product, Induces Hepatic Changes in Adult Mice. Int. J. Environ. Res. Public Health 2020, 17, 7895. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17217895
Doherty Lyons S, Blum JL, Hoffman-Budde C, Tijerina PB, Fiel MI, J. Conklin D, Gany F, Odin JA, Zelikoff JT. Prenatal Exposure to Gutkha, a Globally Relevant Smokeless Tobacco Product, Induces Hepatic Changes in Adult Mice. International Journal of Environmental Research and Public Health. 2020; 17(21):7895. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17217895
Chicago/Turabian StyleDoherty Lyons, Shannon, Jason L. Blum, Carol Hoffman-Budde, Pamela B. Tijerina, M. Isabel Fiel, Daniel J. Conklin, Francesca Gany, Joseph A. Odin, and Judith T. Zelikoff. 2020. "Prenatal Exposure to Gutkha, a Globally Relevant Smokeless Tobacco Product, Induces Hepatic Changes in Adult Mice" International Journal of Environmental Research and Public Health 17, no. 21: 7895. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17217895