
Vitamin D and Swimming Exercise Prevent Obesity in Rats under a High-Fat Diet via Targeting FATP4 and TLR4 in the Liver and Adipose Tissue
 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Grouping
2.2. Ethical Statement
2.3. Diets
2.4. Experimental Design
2.4.1. Groups
2.4.2. Swimming Exercise
2.5. Measurements of Obesity-Associated Parameters
Body Weight Gain
2.6. Sample Collection
2.7. Adiposity Index
2.8. Biochemical Analysis
2.8.1. Lipid Profile
2.8.2. Liver Enzymes
2.8.3. Inflammatory Markers
2.8.4. Hormones
Adipokines
Insulin
2.9. Quantitative Realtime PCR of FATP4 in Hepatic and Adipose Tissue
2.10. Histological Analysis
2.10.1. Histopathological Study of Liver Tissue Sections
2.10.2. Histopathological Study of Adipose Tissue Sections
2.10.3. Immunohistochemical Analysis
2.10.4. Immunohistochemical Evaluation
2.11. Statistical Analysis
3. Results
3.1. Measures of Adiposity
3.2. Lipid Profile
3.3. Liver Enzymes Levels
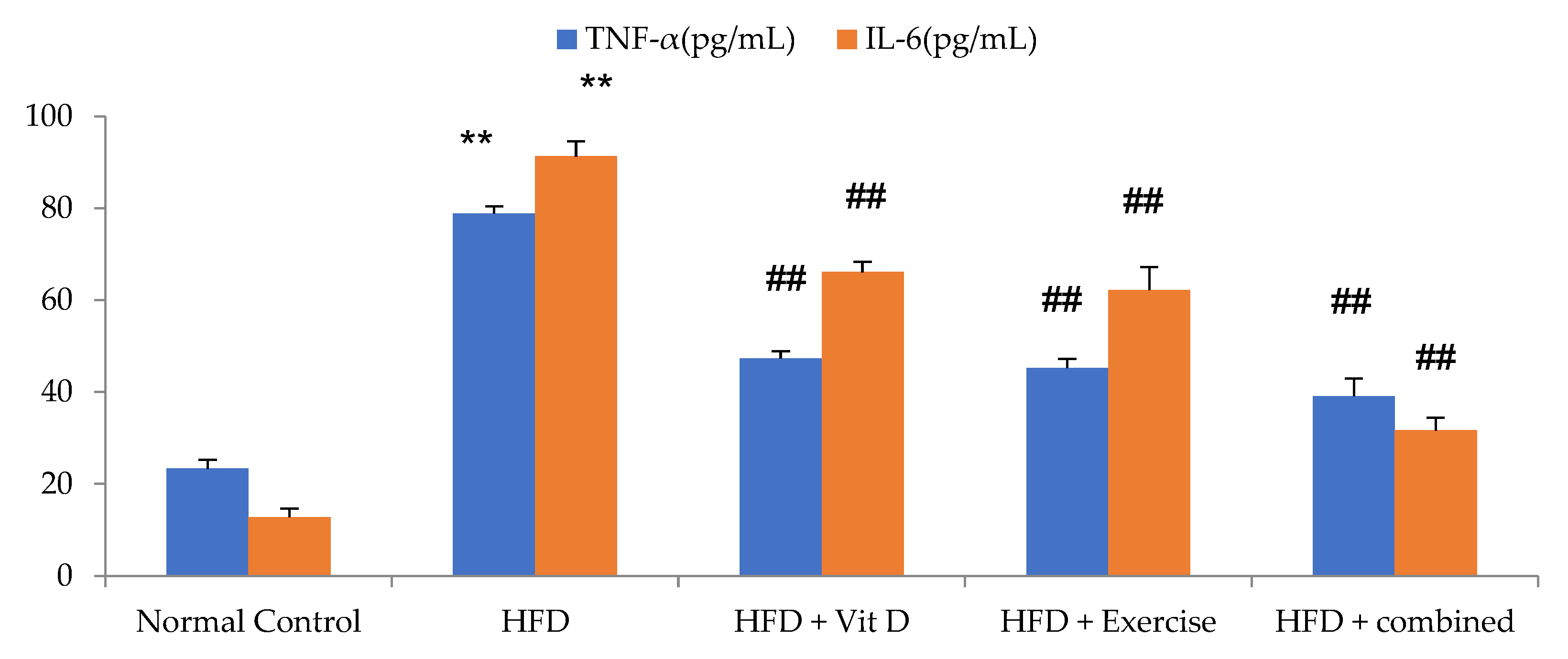
3.4. Inflammatory Markers Levels
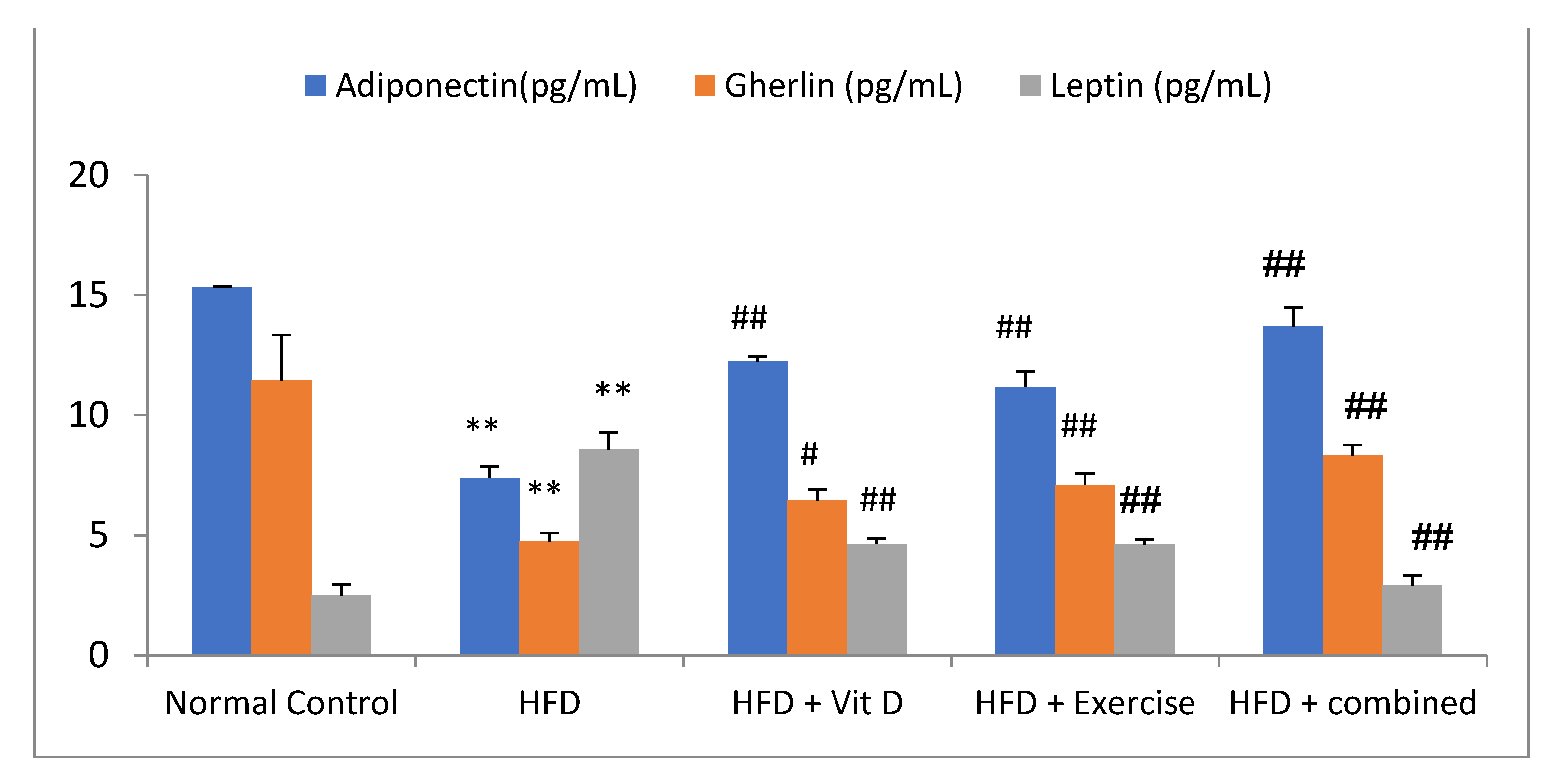
3.5. Adipokines Levels
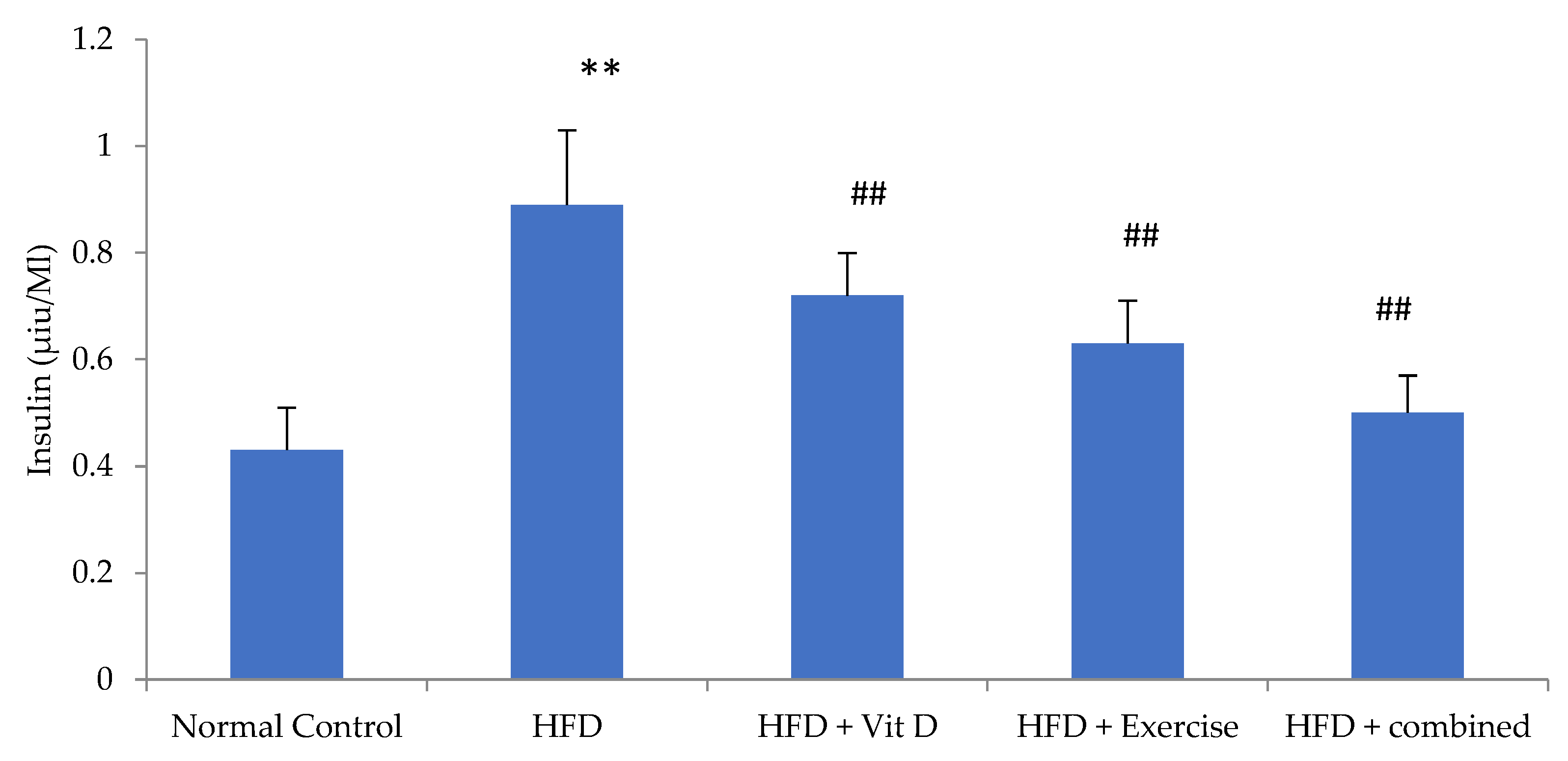
3.6. Insulin Level
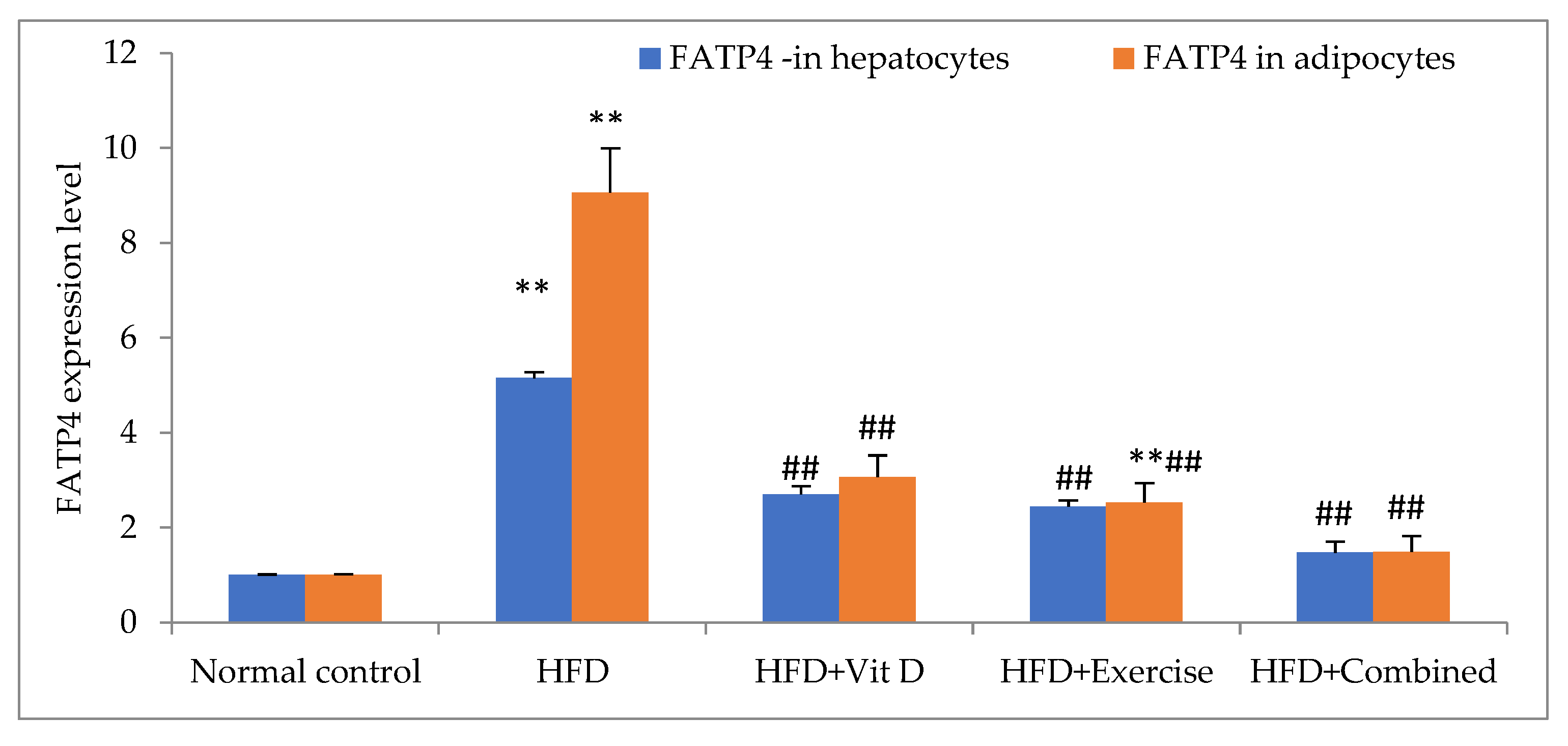
3.7. Expression of FATP4 in Tissues
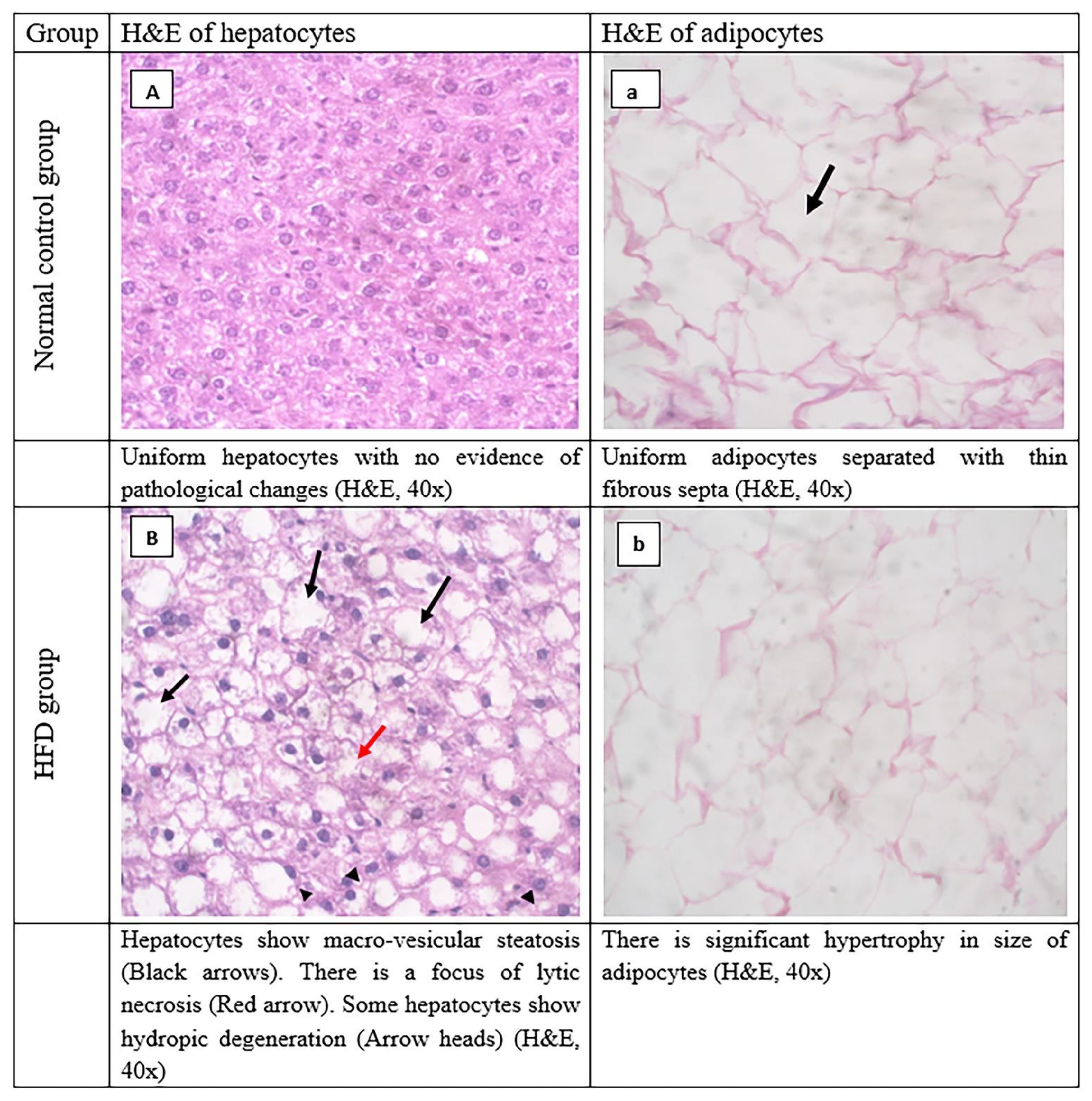
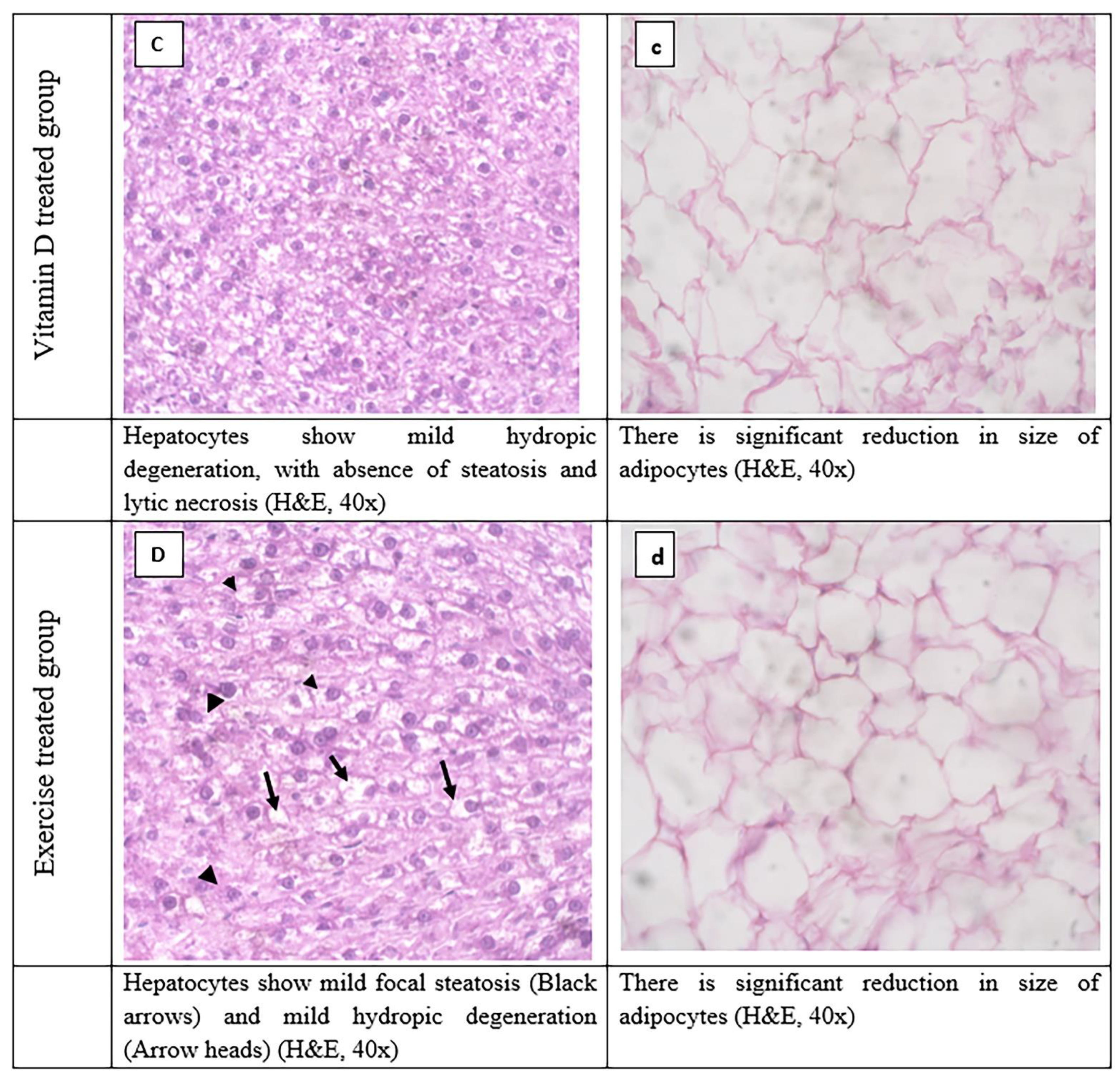
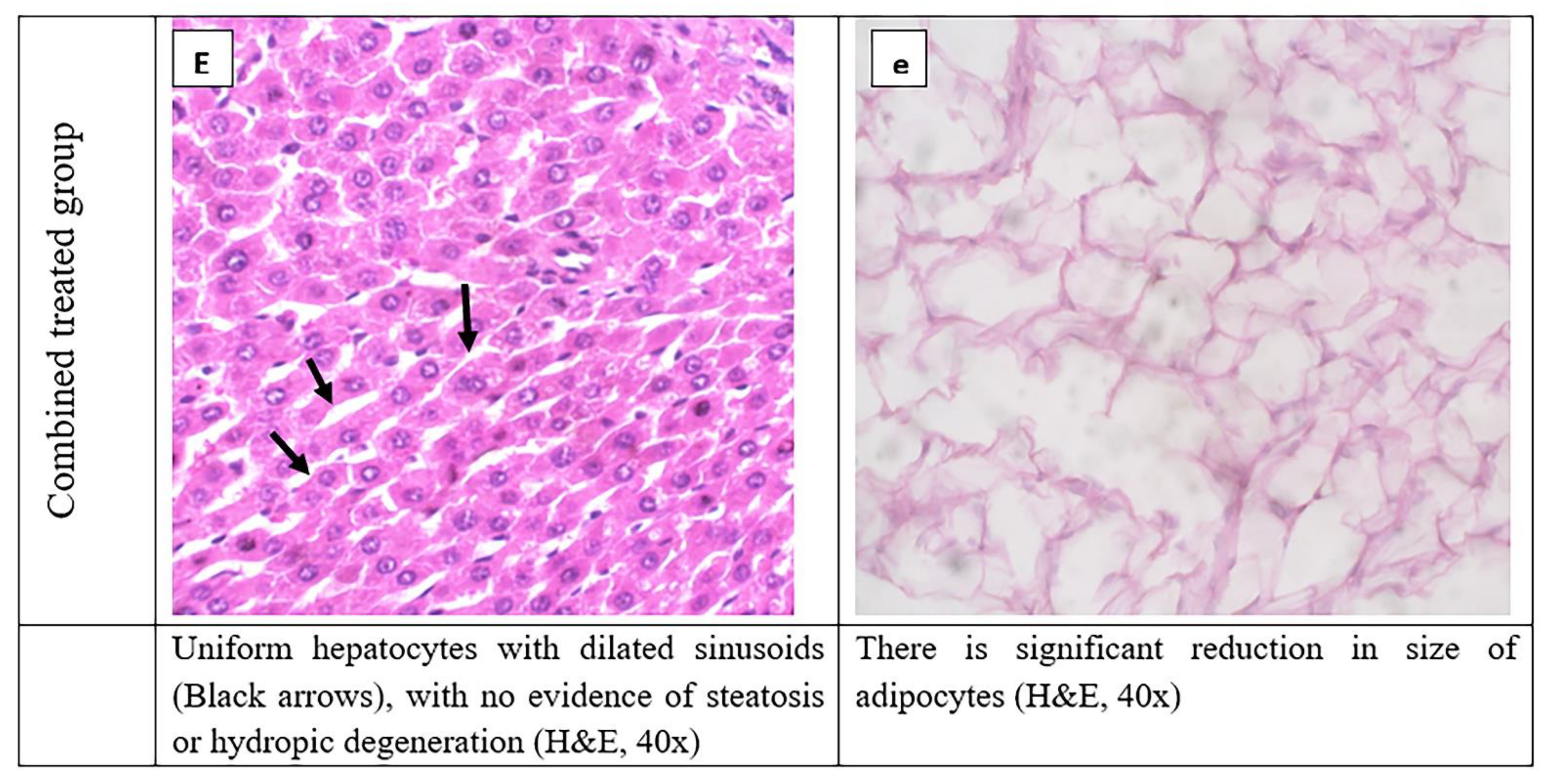
3.8. Histopathological Evaluation of Adipose Tissue and Liver Tissues Sections
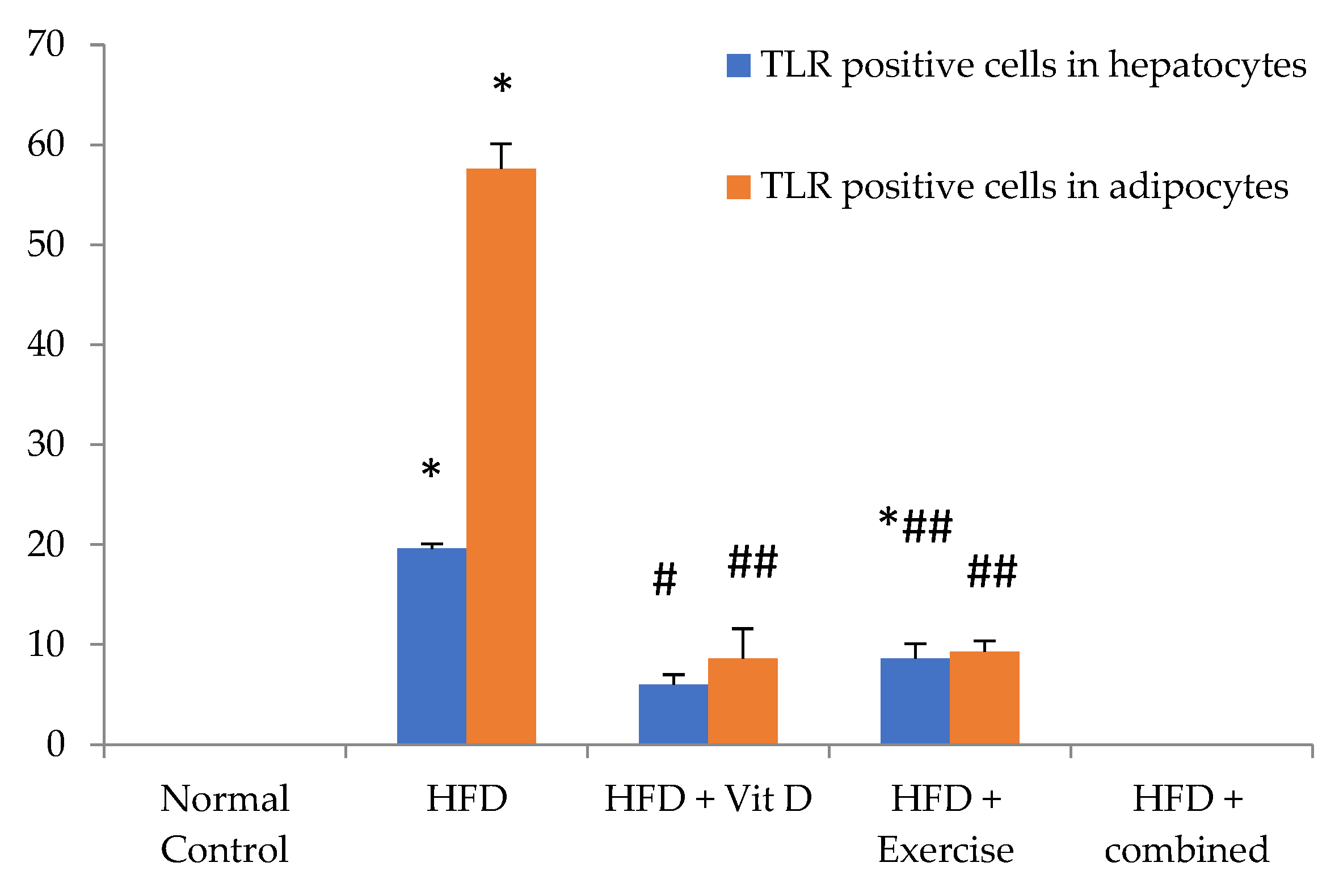
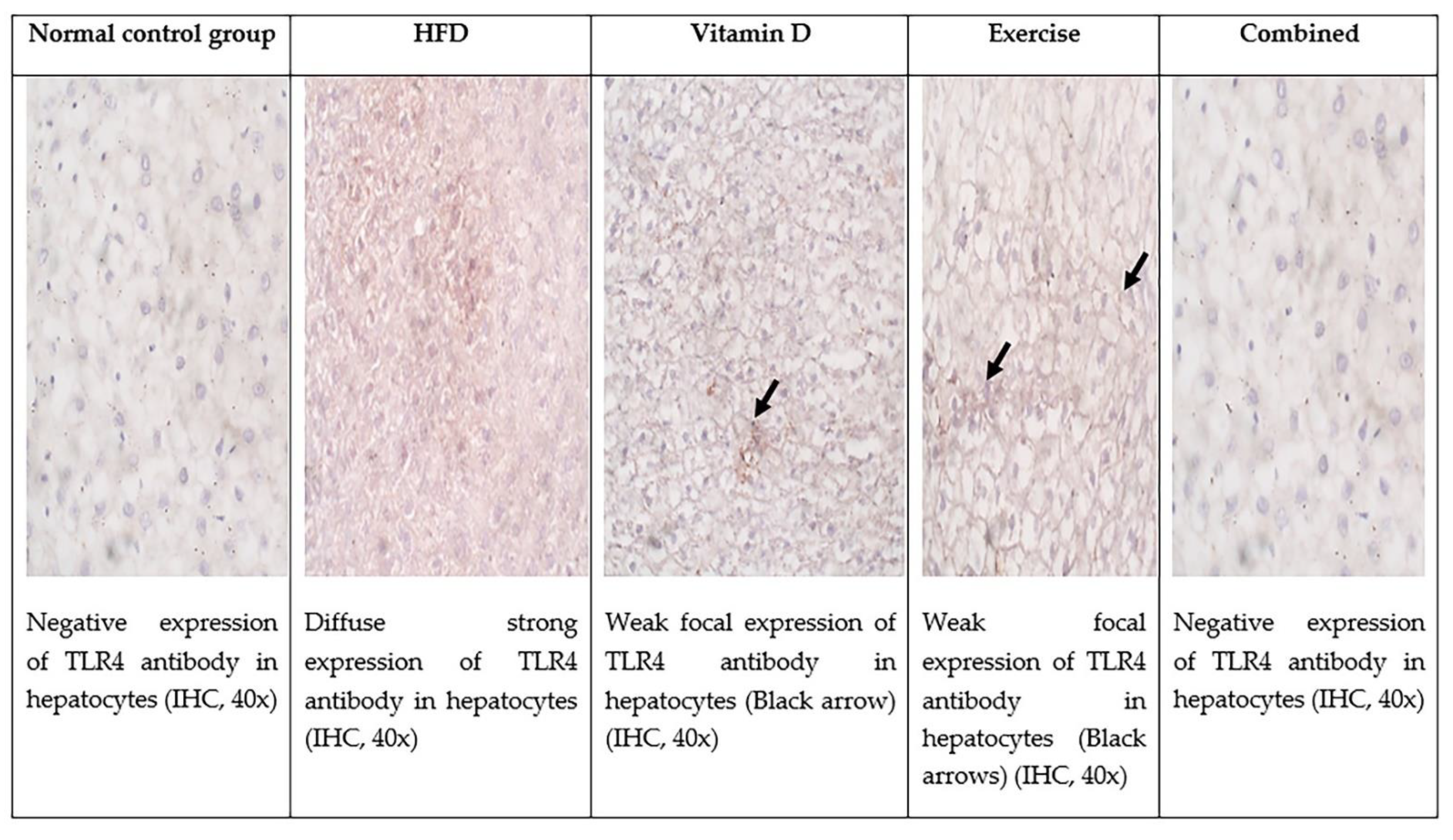
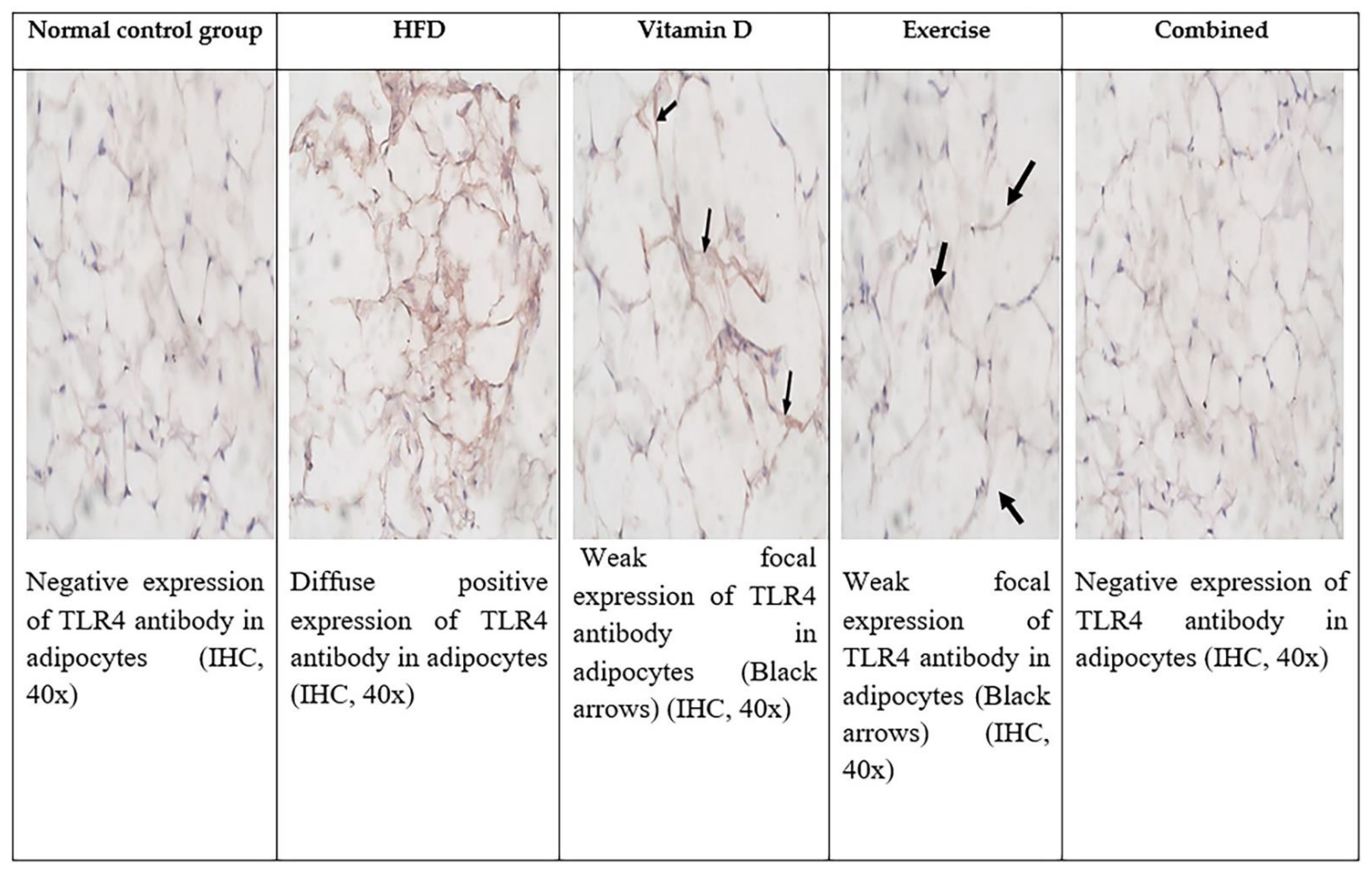
3.9. Immunohistochemical Evaluation of Adipose Tissue and Liver Tissues Sections
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karnafel, W.; Możejko-Pastewka, B. Obesity and the risk of type 2 diabetes mellitus and certain types of cancer. Clin. Diabetol. 2015, 4, 163–171. [Google Scholar]
- Johnson, C.B.; Davis, M.K.; Law, A.; Sulpher, J. Shared risk factors for cardiovascular disease and cancer: Implications for preventive health and clinical care in oncology patients. Can. J. Cardiol. 2016, 32, 900–907. [Google Scholar] [CrossRef]
- Caleyachetty, R.; Thomas, G.N.; Toulis, K.A.; Mohammed, N.; Gokhale, K.M.; Balachandran, K.; Nirantharakumar, K. Metabolically healthy obese and incident cardiovascular disease events among 3.5 million men and women. J. Am. Coll. Cardiol. 2017, 70, 1429–1437. [Google Scholar] [CrossRef]
- Vilarrasa, N.; Maravall, J.; Estepa, A.; Sánchez, R.; Masdevall, C.; Navarro, M.; Alía, P.; Soler, J.; Gómez, J. Low 25-hydroxyvitamin D concentrations in obese women: Their clinical significance and relationship with anthropometric and body composition variables. J. Endocrinol. Investig. 2007, 30, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Saied, E.M.; El-Maradny, Y.A.; Osman, A.A.; Darwish, A.M.G.; Abo Nahas, H.H.; Niedbała, G.; Piekutowska, M.; Abdel-Rahman, M.A.; Balbool, B.A.; Abdel-Azeem, A.M. A Comprehensive Review about the Molecular Structure of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Insights into Natural Products against COVID-19. Pharmaceutics 2021, 13, 1759. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrova, K.; Mozaffarian, D.; Pischon, T. Addressing the perfect storm: Biomarkers in obesity and pathophysiology of cardiometabolic risk. Clin. Chem. Lab. Med. 2018, 64, 142–153. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Choi, J.-W.; Yun, J.W.; Chung, I.-S.; Cho, H.C.; Song, S.-E.; Im, S.-S.; Song, D.-K. Proteomics approach to identify serum biomarkers associated with the progression of diabetes in Korean patients with abdominal obesity. PLoS ONE 2019, 14, e0222032. [Google Scholar] [CrossRef] [PubMed]
- Timotin, A.; Cinato, M.; Boal, F.; Dejean, S.; Anesia, R.; Arnaut, O.; Lagente, C.; Roncalli, J.; Desmoulin, F.; Tronchere, H. Differential protein profiling as a potential multi-marker approach for obese patients with heart failure: A retrospective study. Sci. Rep. 2018, 8, 7894. [Google Scholar] [CrossRef] [Green Version]
- Foster-Schubert, K.E.; Cummings, D.E. Emerging therapeutic strategies for obesity. Endocr. Rev. 2006, 27, 779–793. [Google Scholar] [CrossRef] [Green Version]
- Ragab, S.M.; Abd Elghaffar, S.K.; El-Metwally, T.H.; Badr, G.; Mahmoud, M.H.; Omar, H.M. Effect of a high fat, high sucrose diet on the promotion of non-alcoholic fatty liver disease in male rats: The ameliorative role of three natural compounds. Lipids Health Dis. 2015, 14, 83. [Google Scholar] [CrossRef] [Green Version]
- Afifi, N.A.; Ramadan, A.; Erian, E.Y.; Saleh, D.O.; Sedik, A.A.; Badawi, M.; El Hotaby, W. Trigonelline attenuates hepatic complications and molecular alterations in high-fat high-fructose diet-induced insulin resistance in rats. Can. J. Physiol. Pharmacol. 2017, 95, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, S.A.M.; Shedid, E.S.; Saied, E.M.; Jassbi, A.R.; Jamebozorgi, F.H.; Rateb, M.E.; Du, M.; Abdel-Daim, M.M.; Kai, G.-Y.; Al-Hammady, M.A.M.; et al. Cyanobacteria—From the Oceans to the Potential Biotechnological and Biomedical Applications. Marine Drugs 2021, 19, 241. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Han, H.J.; Kim, A.H.; Choi, J.Y.; Cho, S.J.; Park, Y.B.; Jung, U.J.; Choi, M.S. d-Allulose supplementation normalized the body weight and fat-pad mass in diet-induced obese mice via the regulation of lipid metabolism under isocaloric fed condition. Mol. Nutr. Food Res. 2016, 60, 1695–1706. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, M.N.; Fadl, M.A.; Wahab, H.A.H.A.; Hashem, R.M.H.; Faid, R.A.A.M.; Mousa, N.A.; Khalil, M.F. Effect of a Training Program Accompanied by a Suggested Diet on Some Physiological Variables and Regulating Blood Sugar Level in Type II Diabetics. Int. J. Hum. Mov. Sports Sci 2022, 10, 54–65. [Google Scholar] [CrossRef]
- Wortsman, J.; Matsuoka, L.Y.; Chen, T.C.; Lu, Z.; Holick, M.F. Decreased bioavailability of vitamin D in obesity. Am. J. Clin. Nutr. 2000, 72, 690–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalaby, M.N.; Sakoury, M.M.A.; Akl, H.F.M.; Hassan, R.H.A.; Ababtain, H.A.S.; Alghamdi, A. Effect of Physical Exertion on the Effect of Physical Exertion on the Concentration of Copper and Blood Pressure in Athletesn the Concentration of Copper and Blood Pressure in Athletes. Pedagogy phys. cult. sports 2022, 26, 260–264. [Google Scholar] [CrossRef]
- Yao, Y.; Zhu, L.; He, L.; Duan, Y.; Liang, W.; Nie, Z.; Jin, Y.; Wu, X.; Fang, Y. A meta-analysis of the relationship between vitamin D deficiency and obesity. Int. J. Clin. Exp. Med. 2015, 8, 14977. [Google Scholar]
- Shalaby, M.N.; Sakoury, M.M.A.; Abdi, E.; Elgamal, S.; Elrkbwey, S.; Ramadan, W.; Taiar, R. The Impact of Resistance Training on Gene Expression of IGF1 and Athletes’ Physiological Parameters. Open Access Maced J Med Sci 2021, 9, 934–940. [Google Scholar] [CrossRef]
- Mohamed, D.I.; Alaa El-Din Aly El-Waseef, D.; Nabih, E.S.; El-Kharashi, O.A.; Abd El-Kareem, H.F.; Abo Nahas, H.H.; Abdel-Wahab, B.A.; Helmy, Y.A.; Alshawwa, S.Z.; Saied, E.M. Acetylsalicylic Acid Suppresses Alcoholism-Induced Cognitive Impairment Associated with Atorvastatin Intake by Targeting Cerebral MiRNA155 and NLRP3: In Vivo, and In Silico Study. Pharmaceutics 2022, 14, 529. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-S.; Kim, J.-C.; Kim, J.-S.; Kim, S.H. Effects of swimming exercise on serum irisin and bone FNDC5 in rat models of high-fat diet-induced osteoporosis. J. Sport. Sci. Med. 2019, 18, 596. [Google Scholar]
- Levin, B.E.; Dunn-Meynell, A.A. Defense of body weight depends on dietary composition and palatability in rats with diet-induced obesity. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2002, 282, R46–R54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul Kadir, N.A.A.; Rahmat, A.; Jaafar, H.Z. Protective effects of tamarillo (Cyphomandra betacea) extract against high fat diet induced obesity in Sprague-Dawley rats. J. Obes. 2015, 2015, 846041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orhan, C.; Tuzcu, M.; Deeh Defo, P.B.; Sahin, N.; Ojalvo, S.P.; Sylla, S.; Komorowski, J.R.; Sahin, K. Effects of a Novel Magnesium Complex on Metabolic and Cognitive Functions and the Expression of Synapse-Associated Proteins in Rats Fed a High-Fat Diet. Biol. Trace Elem. Res. 2022, 200, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, D.I.; Abou-Bakr, D.A.; Ezzat, S.F.; El-Kareem, H.F.A.; Nahas, H.H.A.; Saad, H.A.; Mehana, A.E.; Saied, E.M. Vitamin D3 Prevents the Deleterious Effects of Testicular Torsion on Testis by Targeting MiRNA-145 and ADAM17: In Silico and In Vivo Study. Pharmaceuticals 2021, 14, 1222. [Google Scholar] [CrossRef] [PubMed]
- Ciolac, E.G.; Greve, J.M.D.A. Exercise-induced improvements in cardiorespiratory fitness and heart rate response to exercise are impaired in overweight/obese postmenopausal women. Clinics 2011, 66, 583–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eleawa, S.; Sakr, H.F. Effect of exercise and orlistat therapy in rat model of obesity induced with high fat diet. Med. J. Cairo Univ. 2013, 81, 59–67. [Google Scholar]
- He, C.; Cheng, D.; Peng, C.; Li, Y.; Zhu, Y.; Lu, N. High-fat diet induces dysbiosis of gastric microbiota prior to gut microbiota in association with metabolic disorders in mice. Front. Microbiol. 2018, 9, 639. [Google Scholar] [CrossRef] [Green Version]
- Fossati, P.; Prencipe, L. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem. 1982, 28, 2077–2080. [Google Scholar] [CrossRef]
- Allain, C.C.; Poon, L.S.; Chan, C.S.; Richmond, W.; Fu, P.C. Enzymatic determination of total serum cholesterol. Clin. Chem. 1974, 20, 470–475. [Google Scholar]
- Burstein, M.; Scholnick, H.; Morfin, R.J. Rapid method for the isolation of lipoproteins from human serum by precipitation with polyanions. J. Lipid Res. 1970, 11, 583–595. [Google Scholar] [CrossRef]
- Wieland, H.; Seidel, D. A simple specific method for precipitation of low density lipoproteins. J. Lipid Res. 1983, 24, 904–909. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, D.I.; Ezzat, S.F.; Elayat, W.M.; El-Kharashi, O.A.; El-Kareem, H.F.A.; Nahas, H.H.A.; Abdel-Wahab, B.A.; Alshawwa, S.Z.; Saleh, A.; Helmy, Y.A.; et al. Hepatoprotective Role of Carvedilol against Ischemic Hepatitis Associated with Acute Heart Failure via Targeting MiRNA-17 and Mitochondrial Dynamics-Related Proteins: An In Vivo and In Silico Study. Pharmaceuticals 2022, 15, 832. [Google Scholar] [CrossRef] [PubMed]
- Ilić, I.R.; Stojanović, N.M.; Radulović, N.S.; Živković, V.V.; Randjelović, P.J.; Petrović, A.S.; Božić, M.; Ilić, R.S. The Quantitative ER immunohistochemical analysis in breast cancer: Detecting the 3+ 0, 4+ 0, and 5+ 0 Allred score cases. Medicina 2019, 55, 461. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Xu, H.; Yu, X.; Wu, Y.; Sui, D. Metformin ameliorates diabetic nephropathy in a rat model of low-dose streptozotocin-induced diabetes. Exp. Ther. Med. 2017, 14, 383–390. [Google Scholar] [CrossRef]
- Petrie, J.R.; Guzik, T.J.; Touyz, R.M. Diabetes, hypertension, and cardiovascular disease: Clinical insights and vascular mechanisms. Can. J. Cardiol. 2018, 34, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Kaur, J. A comprehensive review on metabolic syndrome. Cardiol. Res. Pract. 2014, 2014, 943162. [Google Scholar] [CrossRef]
- Hruby, A.; Hu, F.B. The epidemiology of obesity: A big picture. Pharmacoeconomics 2015, 33, 673–689. [Google Scholar] [CrossRef]
- Öner-İyidoğan, Y.; Kocak, H.; Seyidhanoğlu, M.; Gürdöl, F.; Gülçubuk, A.; Yildirim, F.; Çevik, A.; Uysal, M. Curcumin prevents liver fat accumulation and serum fetuin-A increase in rats fed a high-fat diet. J. Physiol. Biochem. 2013, 69, 677–686. [Google Scholar] [CrossRef]
- Lobo, S.; Wiczer, B.M.; Smith, A.J.; Hall, A.M.; Bernlohr, D.A. Fatty acid metabolism in adipocytes: Functional analysis of fatty acid transport proteins 1 and 4. J. Lipid Res. 2007, 48, 609–620. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Herrmann, T.; Seeßle, J.; Liebisch, G.; Merle, U.; Stremmel, W.; Chamulitrat, W. Role of fatty acid transport protein 4 in metabolic tissues: Insights into obesity and fatty liver disease. Biosci. Rep. 2022, 42, BSR20211854. [Google Scholar] [CrossRef] [PubMed]
- Engin, A.B. What is lipotoxicity? Obes. Lipotoxicity 2017, 960, 197–220. [Google Scholar]
- Abdelmegeed, M.A.; Esterle, A.; Chilian, W.M. Eicosanoids in Metabolic Syndrome. Immunopharmacology 2013, 66, 157. [Google Scholar]
- Herrmann, T.; Buchkremer, F.; Gosch, I.; Hall, A.M.; Bernlohr, D.A.; Stremmel, W. Mouse fatty acid transport protein 4 (FATP4): Characterization of the gene and functional assessment as a very long chain acyl-CoA synthetase. Gene 2001, 270, 31–40. [Google Scholar] [CrossRef]
- Araújo, J.R.; Tomas, J.; Brenner, C.; Sansonetti, P. Impact of high-fat diet on the intestinal microbiota and small intestinal physiology before and after the onset of obesity. Biochimie 2017, 141, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Ehehalt, R.; Sparla, R.; Kulaksiz, H.; Herrmann, T.; Füllekrug, J.; Stremmel, W. Uptake of long chain fatty acids is regulated by dynamic interaction of FAT/CD36 with cholesterol/sphingolipid enriched microdomains (lipid rafts). BMC Cell Biol. 2008, 9, 45. [Google Scholar] [CrossRef] [Green Version]
- Gimeno, R.E.; Ortegon, A.M.; Patel, S.; Punreddy, S.; Ge, P.; Sun, Y.; Lodish, H.F.; Stahl, A. Characterization of a heart-specific fatty acid transport protein. J. Biol. Chem. 2003, 278, 16039–16044. [Google Scholar] [CrossRef]
- Jia, Z.; Moulson, C.L.; Pei, Z.; Miner, J.H.; Watkins, P.A. Fatty acid transport protein 4 is the principal very long chain fatty acyl-CoA synthetase in skin fibroblasts. J. Biol. Chem. 2007, 282, 20573–20583. [Google Scholar] [CrossRef] [Green Version]
- Stahl, A.; Hirsch, D.J.; Gimeno, R.E.; Punreddy, S.; Ge, P.; Watson, N.; Patel, S.; Kotler, M.; Raimondi, A.; Tartaglia, L.A. Identification of the major intestinal fatty acid transport protein. J. Mol. Cell 1999, 4, 299–308. [Google Scholar] [CrossRef]
- Milger, K.; Herrmann, T.; Becker, C.; Gotthardt, D.; Zickwolf, J.; Ehehalt, R.; Watkins, P.A.; Stremmel, W.; Füllekrug, J. Cellular uptake of fatty acids driven by the ER-localized acyl-CoA synthetase FATP4. J. Cell Sci. 2006, 119, 4678–4688. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Pei, Z.; Maiguel, D.; Toomer, C.J.; Watkins, P.A. The fatty acid transport protein (FATP) family: Very long chain acyl-CoA synthetases or solute carriers? J. Mol. Neurosci. 2007, 33, 25–31. [Google Scholar] [CrossRef]
- Digel, M.; Ehehalt, R.; Stremmel, W.; Füllekrug, J. Acyl-CoA synthetases: Fatty acid uptake and metabolic channeling. Mol. Cell Biochem. 2009, 326, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.A. New insights into the roles of proteins and lipids in membrane transport of fatty acids. Prostaglandins Leukot. Essent. Fat. Acids 2007, 77, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Gertow, K.; Pietiläinen, K.H.; Yki-Järvinen, H.; Kaprio, J.; Rissanen, A.; Eriksson, P.; Hamsten, A.; Fisher, R. Expression of fatty-acid-handling proteins in human adipose tissue in relation to obesity and insulin resistance. Diabetologia 2004, 47, 1118–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, A.J.; Chen, D.F. The expression and the significance of L-FABP and FATP4 in the development of nonalcoholic fatty liver disease in rats. Zhonghua Gan Zang Bing Za Zhi = Zhonghua Ganzangbing Zazhi = Chin. J. Hepatol. 2005, 13, 776–779. [Google Scholar]
- Krammer, J.; Digel, M.; Ehehalt, F.; Stremmel, W.; Füllekrug, J.; Ehehalt, R. Overexpression of CD36 and acyl-CoA synthetases FATP2, FATP4 and ACSL1 increases fatty acid uptake in human hepatoma cells. Int. J. Med. Sci. 2011, 8, 599. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Zhou, J.; Song, S.-s.; Kong, W.; Shi, Y.-C.; Chen, L.-L.; Zeng, T.-S. TLR4/AP-1-targeted anti-inflammatory intervention attenuates insulin sensitivity and liver steatosis. Mediat. Inflamm. 2020, 2020, 2960517. [Google Scholar] [CrossRef]
- Guo, J.; Friedman, S.L.J.F. Toll-like receptor 4 signaling in liver injury and hepatic fibrogenesis. Fibrogenesis Tissue Repair 2010, 3, 21. [Google Scholar] [CrossRef]
- Rogero, M.M.; Calder, P.C.J.N. Obesity, inflammation, toll-like receptor 4 and fatty acids. Nutrients 2018, 10, 432. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-J.; Choi, Y.; Choi, Y.-H.; Park, T. Obesity activates toll-like receptor-mediated proinflammatory signaling cascades in the adipose tissue of mice. J. Nutr. Biochem. 2012, 23, 113–122. [Google Scholar] [CrossRef]
- Miura, K.; Ohnishi, H.J. Role of gut microbiota and Toll-like receptors in nonalcoholic fatty liver disease. World J. Gastroenterol. WJG 2014, 20, 7381. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, M.M.; Biscaia, P.B.; Brunoski, J.; Ribeiro, R.A.; Franco, G.C.N.; Scomparin, D.X. Vitamin D supplementation decreases visceral adiposity and normalizes leptinemia and circulating TNF-α levels in western diet-fed obese rats. Life Sci. 2021, 278, 119550. [Google Scholar] [CrossRef] [PubMed]
- Pramono, A.; Jocken, J.W.; Essers, Y.P.; Goossens, G.H.; Blaak, E.E. Vitamin D and tissue-specific insulin sensitivity in humans with overweight/obesity. J. Clin. Endocrinol. Metab. 2019, 104, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Mousa, A.; Naderpoor, N.; Wilson, K.; Plebanski, M.; de Courten, M.P.; Scragg, R.; de Courten, B. Vitamin D supplementation increases adipokine concentrations in overweight or obese adults. Eur. J. Nutr. 2020, 59, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Lotfi-Dizaji, L.; Mahboob, S.; Aliashrafi, S.; Vaghef-Mehrabany, E.; Ebrahimi-Mameghani, M.; Morovati, A. Effect of vitamin D supplementation along with weight loss diet on meta-inflammation and fat mass in obese subjects with vitamin D deficiency: A double-blind placebo-controlled randomized clinical trial. Clin. Endocrinol. 2019, 90, 94–101. [Google Scholar] [CrossRef] [Green Version]
- Guareschi, Z.M.; Valcanaia, A.C.; Ceglarek, V.M.; Hotz, P.; Amaral, B.K.; de Souza, D.W.; de Souza, T.A.; Nardelli, T.; Ferreira, T.R.; Leite, N.C. The effect of chronic oral vitamin D supplementation on adiposity and insulin secretion in hypothalamic obese rats. Br. J. Nutr. 2019, 121, 1334–1344. [Google Scholar] [CrossRef]
- Vranić, L.; Mikolašević, I.; Milić, S.J.M. Vitamin D deficiency: Consequence or cause of obesity? J. Med. 2019, 55, 541. [Google Scholar] [CrossRef] [Green Version]
- Refaat, B.; Abdelghany, A.H.; Ahmad, J.; Abdalla, O.M.; Elshopakey, G.E.; Idris, S.; El-Boshy, M.J.B. Vitamin D3 enhances the effects of omega-3 oils against metabolic dysfunction-associated fatty liver disease in rat. BioFactors 2022, 48, 498–513. [Google Scholar] [CrossRef]
- Yin, Y.; Yu, Z.; Xia, M.; Luo, X.; Lu, X.; Ling, W. Vitamin D attenuates high fat diet–induced hepatic steatosis in rats by modulating lipid metabolism. J. Eur. J. Clin. Investig. 2012, 42, 1189–1196. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, M.; Xu, W.; Zhang, H.; Qian, W.; Li, X.; Cheng, X. Active vitamin D supplementation alleviates initiation and progression of nonalcoholic fatty liver disease by repressing the p53 pathway. Life Sci. 2020, 241, 117086. [Google Scholar] [CrossRef]
- Al-Ghamdi, H.A.; Al Fayez, F.F.; Bima, A.I.; Khawaji, T.M.; Elsamanoudy, A.Z. Study of cellular senescence and vitamin D deficiency in nonalcoholic fatty liver disease and the potential protective effect of vitamin D supplementation. J. Clin. Exp. Hepatol. 2021, 11, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Long, Q.; Chen, F.; Zhang, T.; Wang, W.J.C. Active vitamin D impedes the progression of non-alcoholic fatty liver disease by inhibiting cell senescence in a rat model. Clin. Res. Hepatol. Gastroenterol. 2020, 44, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Jordy, A.B.; Kraakman, M.J.; Gardner, T.; Estevez, E.; Kammoun, H.L.; Weir, J.M.; Kiens, B.; Meikle, P.J.; Febbraio, M.A.; Henstridge, D.C. Analysis of the liver lipidome reveals insights into the protective effect of exercise on high-fat diet-induced hepatosteatosis in mice. Am. J. Physiol.-Endocrinol. Metab. 2015, 308, E778–E791. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, J.; Jordy, A.B.; Sjøberg, K.A.; Füllekrug, J.; Stahl, A.; Nybo, L.; Kiens, B. Enhanced fatty acid oxidation and FATP4 protein expression after endurance exercise training in human skeletal muscle. PLoS ONE 2012, 7, e29391. [Google Scholar] [CrossRef] [Green Version]
- Chistyakov, D.V.; Azbukina, N.V.; Lopachev, A.V.; Kulichenkova, K.N.; Astakhova, A.A.; Sergeeva, M.G. Rosiglitazone as a modulator of TLR4 and TLR3 signaling pathways in rat primary neurons and astrocytes. J. Int. J. Mol. Sci. 2018, 19, 113. [Google Scholar] [CrossRef] [Green Version]
- Hassan, N.F.; Nada, S.A.; Hassan, A.; El-Ansary, M.R.; Al-Shorbagy, M.Y.; Abdelsalam, R.M. Saroglitazar deactivates the hepatic LPS/TLR4 signaling pathway and ameliorates adipocyte dysfunction in rats with high-fat emulsion/LPS model-induced non-alcoholic steatohepatitis. Inflammation 2019, 42, 1056–1070. [Google Scholar] [CrossRef]
- Roth, C.L.; Elfers, C.T.; Figlewicz, D.P.; Melhorn, S.J.; Morton, G.J.; Hoofnagle, A.; Yeh, M.M.; Nelson, J.E.; Kowdley, K.V. Vitamin D deficiency in obese rats exacerbates nonalcoholic fatty liver disease and increases hepatic resistin and Toll-like receptor activation. Hepatology 2012, 55, 1103–1111. [Google Scholar] [CrossRef]
- Jahn, D.; Dorbath, D.; Kircher, S.; Nier, A.; Bergheim, I.; Lenaerts, K.; Hermanns, H.M.; Geier, A. Beneficial effects of vitamin D treatment in an obese mouse model of non-alcoholic steatohepatitis. Nutrients 2019, 11, 77. [Google Scholar] [CrossRef]
- Dickie, L.J.; Church, L.D.; Coulthard, L.R.; Mathews, R.J.; Emery, P.; McDermott, M.F. Vitamin D3 down-regulates intracellular Toll-like receptor 9 expression and Toll-like receptor 9-induced IL-6 production in human monocytes. Rheumatology 2010, 49, 1466–1471. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.G.; Carvalho, B.M.; Tobar, N.; Ropelle, E.R.; Pauli, J.R.; Bagarolli, R.A.; Guadagnini, D.; Carvalheira, J.B.; Saad, M.J. Physical exercise reduces circulating lipopolysaccharide and TLR4 activation and improves insulin signaling in tissues of DIO rats. Diabetes 2011, 60, 784–796. [Google Scholar] [CrossRef] [Green Version]
- Kawanishi, N.; Yano, H.; Yokogawa, Y.; Suzuki, K. Exercise training inhibits inflammation in adipose tissue via both suppression of macrophage infiltration and acceleration of phenotypic switching from M1 to M2 macrophages in high-fat-diet-induced obese mice. Exerc. Immunol. Rev. 2010, 16, 105–118. [Google Scholar] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| FATP4 | Forward: 5′GCACTCTCACTTCCTCTTGC3′ Reverse: 5′GCAGGAAAGGAACAATGCCA3′ |
| GAPDH | Forward: 5′CCCCATTTGATGTTAGCGGG3′ Reverse: 5′AACGACCCCTTCATTGACCT3′ |
| Group Variable (Mean ± SD) | Normal Control | HFD | Vitamin D | Exercise Treated | Combined |
|---|---|---|---|---|---|
| Initial weight (gm) | 232.5 ± 19.48 | 210 ± 22.36 | 219.17 ± 23.54 | 224.17 ± 18 | 219 ± 22.74 |
| Final weight (gm) | 249.16 ± 18.28 | 418.57 ± 31.72 ** | 363.33 ± 13.29 # | 339 ± 30 ## | 312 ± 13.4 *## |
| Body size (cm) | 22.83 ± 1.72 | 27 ± 1.15 ** | 26.16 ± 1.33 | 24.33 ± 1.03 # | 25.8 ± 0.84 ** |
| Fat weight(gm) | 0.5 ± 0.5 | 20.57 ± 5.44 ** | 11.42 ± 2.53 ## | 9.08 ± 1.02 ## | 9.70 ± 0.84 ## |
| Adiposity Index | 0.21 ± 0.22 | 4.9 ± 1.27 ** | 3.16 ± 0.78 ## | 2.70 ± 0.30 ## | 3.11 ± 0.24 ## |
| Lee’s obesity index | 0.27 ± 0.02 | 0.28 ± 0.02 | 0.27 ± 0.01 | 0.29 ± 0.01 | 0.26 ± 0.02 |
| Group Variable (Mean ± SD) | Normal Control | HFD | Vitamin D | Exercise | Combined |
|---|---|---|---|---|---|
| Total cholesterol (mg/dL) | 56.70 ± 6.01 | 231.40 ± 33.28 ** | 115.83 ± 21.54 ## | 97.50 ± 15.41 ## | 64.29 ± 10.18 ## |
| HDL (mg/dL) | 52.50 ± 9.35 | 17.14 ± 3.93 ** | 29.16 ± 3.76 # | 32 ± 3.76 # | 40 ± 10.81 *## |
| LDL (mg/dL) | 59.90 ± 12.50 | 149.14 ± 8.89 ** | 122.83 ± 3.59 ## | 112.97 ± 4.95 ## | 93.71 ± 5.6 **## |
| VLDL (mg/dL) | 29.50 ± 3.72 | 47.43 ± 3.10 ** | 38 ± 1.87 ## | 35.17 ± 1.10 ## | 31.50 ± 3.35 ## |
| TG (mg/dL) | 140.50 ± 5.71 | 207.85 ± 9.11 ** | 190 ± 2.82 ## | 177.67 ± 2.80 ## | 167.14 ± 4.80 ##** |
| Group Variable (Mean ± SD) | Normal Control | HFD | Vitamin D | Exercise | Combined |
|---|---|---|---|---|---|
| ALT (mg/dL) | 19.8 ± 5.80 | 31.4 ± 5.20 ** | 26.80 ± 2.20 | 21.20 ± 1.92 ## | 18 ± 3.80 ## |
| AST (mg/dL) | 25.60 ± 7.20 | 39.80 ± 1.50 ** | 31.40 ± 3.00 # | 27.6 ± 2.40 ## | 20.8 ± 2.77 ## |
| Group Variable (Mean ± SD) | Normal Control | HFD | Vitamin D | Exercise | Combined |
|---|---|---|---|---|---|
| Size of adipocytes (mm2) | 647.75 ± 70.9 | 950.7 ± 154.6 * | 634 ± 22.6 ## | 465.7 ± 186.8 ## | 436.2 ± 91.8 *## |
| Number of adipocytes | 57.2 ± 6 | 62.2 ± 6.8 | 51.5 ± 1.2 ## | 53 ± 2.4 ## | 47.5 ± 2 **## |
| Group Variable (Mean ± SD) | Normal Control | HFD | Vitamin D | Exercise | Combined |
|---|---|---|---|---|---|
| Liver tissue Grading | 0 | 1 (One focus of lytic necrosis) Marked fatty change, Marked hydropic degeneration | 0 No fatty changesMild hydropic degeneration | 0 Mild fatty change, Mild hydropic degeneration | 0 |
| Group Variable (Mean ± SD) | Normal Control | HFD | Vitamin D | Exercise | Combined |
|---|---|---|---|---|---|
| TLR positive cells in hepatocytes | 0 | 19.6 ± 0.5 ** | 6 ± 1 ## | 8.6 ± 1.5 *## | 0 ## |
| Staining intensity | 0 | Moderate Score 2 | Weak Score 1 | Weak Score 1 | 0 |
| Total score | 0 | 4 | 2 | 2 | 0 |
| Group variable (mean ± SD) | Normal group | HFD group | Vitamin D treated group | Exercise treated group | Combined treated group |
| TLR positive cells in adipocytes | 0 | 57.6 ± 2.5 ** | 8.6 ± 3 ## | 9.3 ± 1.1 ## | 0 ## |
| Staining intensity | 0 | Strong Score 3 | Weak Score 1 | Weak Score 1 | 0 |
| Total score | 0 | 7 | 2 | 2 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolieb, E.; Maher, S.A.; Shalaby, M.N.; Alsuhaibani, A.M.; Alharthi, A.; Hassan, W.A.; El-Sayed, K. Vitamin D and Swimming Exercise Prevent Obesity in Rats under a High-Fat Diet via Targeting FATP4 and TLR4 in the Liver and Adipose Tissue. Int. J. Environ. Res. Public Health 2022, 19, 13740. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph192113740
Kolieb E, Maher SA, Shalaby MN, Alsuhaibani AM, Alharthi A, Hassan WA, El-Sayed K. Vitamin D and Swimming Exercise Prevent Obesity in Rats under a High-Fat Diet via Targeting FATP4 and TLR4 in the Liver and Adipose Tissue. International Journal of Environmental Research and Public Health. 2022; 19(21):13740. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph192113740
Chicago/Turabian StyleKolieb, Eman, Shymaa Ahmed Maher, Mohammed Nader Shalaby, Amnah Mohammed Alsuhaibani, Afaf Alharthi, Wael A. Hassan, and Karima El-Sayed. 2022. "Vitamin D and Swimming Exercise Prevent Obesity in Rats under a High-Fat Diet via Targeting FATP4 and TLR4 in the Liver and Adipose Tissue" International Journal of Environmental Research and Public Health 19, no. 21: 13740. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph192113740