
Risky Early Family Environment and Genetic Associations with Adult Metabolic Dysregulation

Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Procedures
2.2.1. Pre-Quarantine
2.2.2. During Quarantine
2.3. Measures
2.4. Overview of Analyses
3. Results
3.1. Correlation Analyses
3.2. Regression Analyses
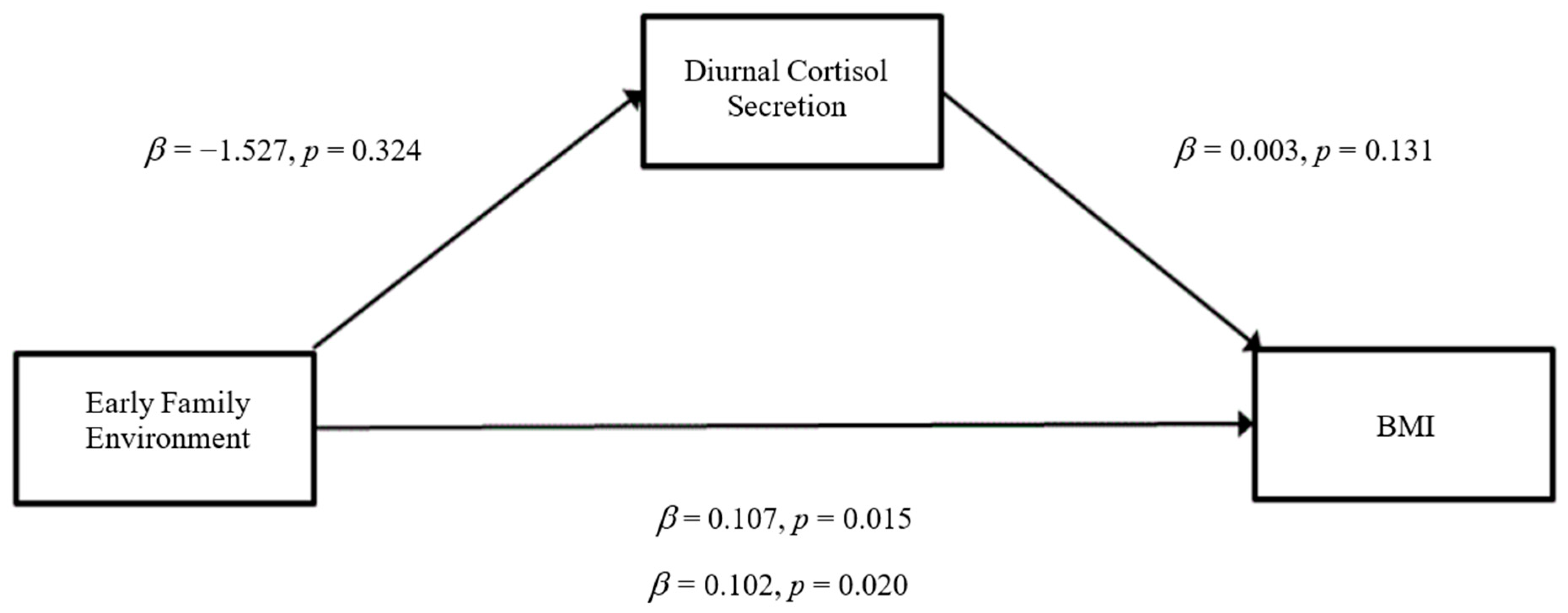
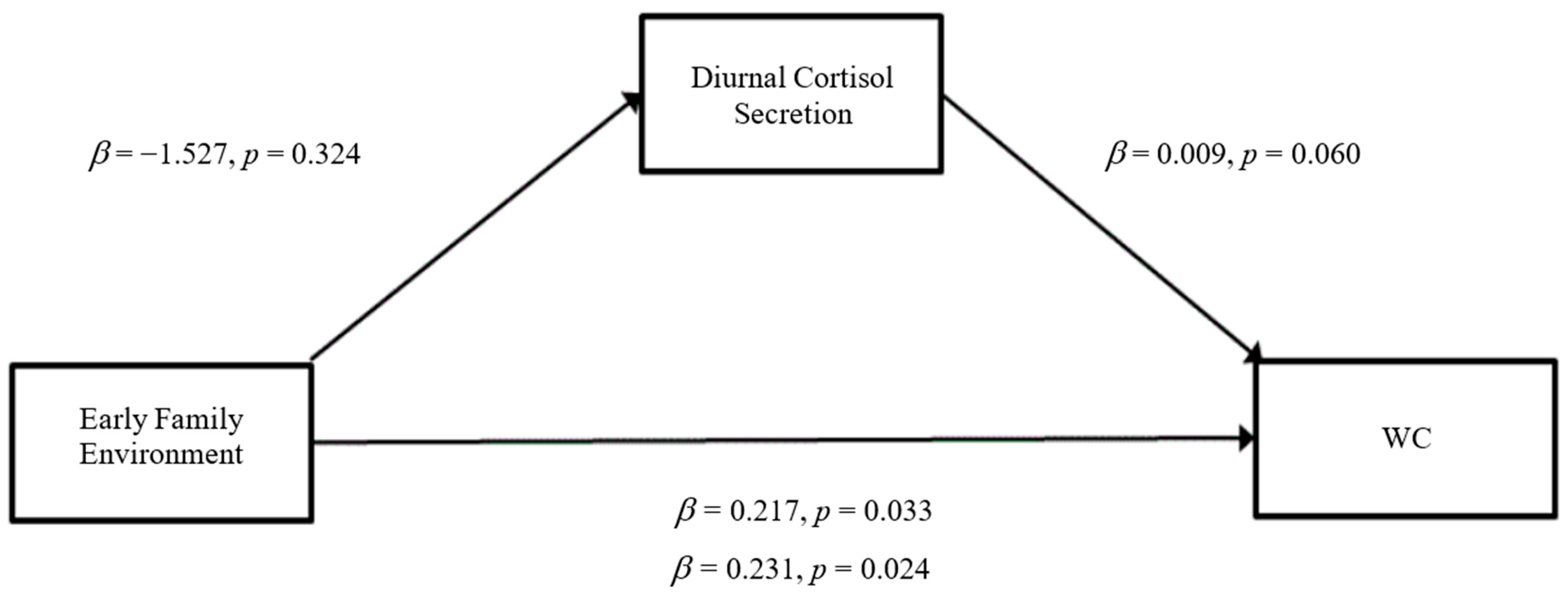
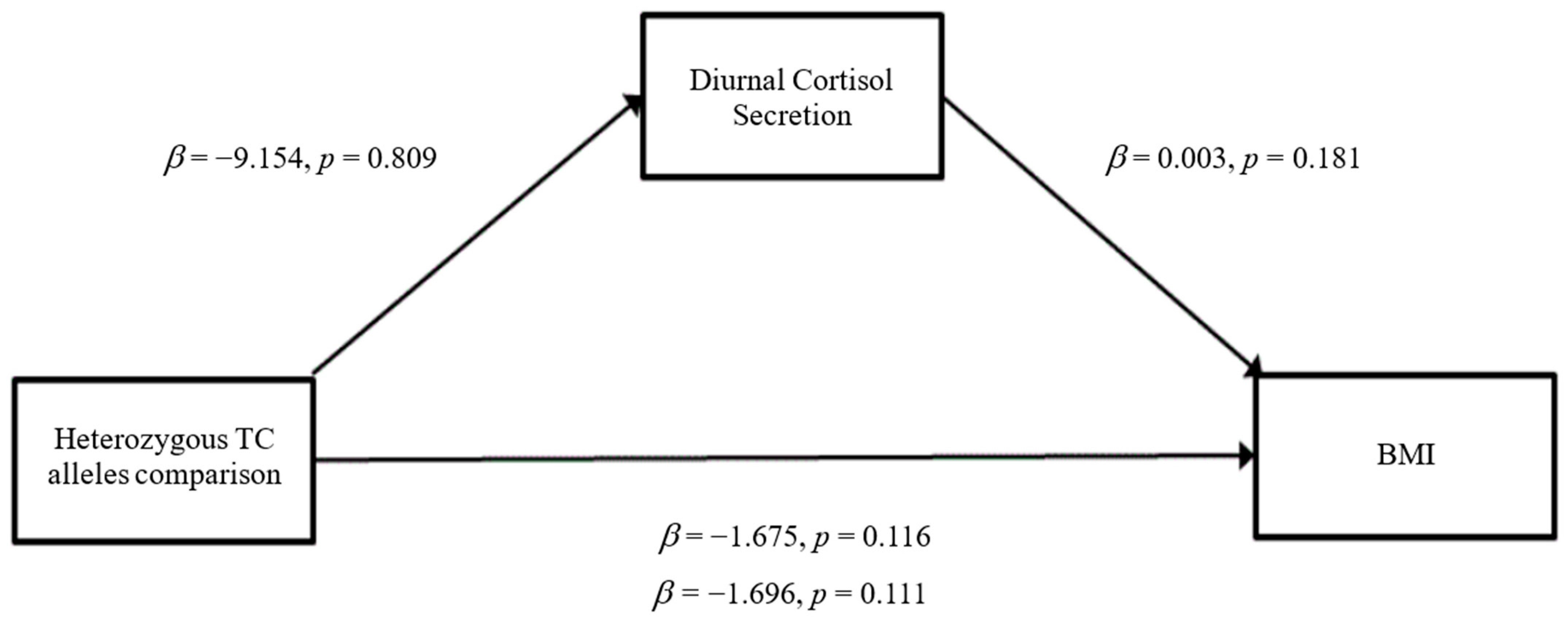
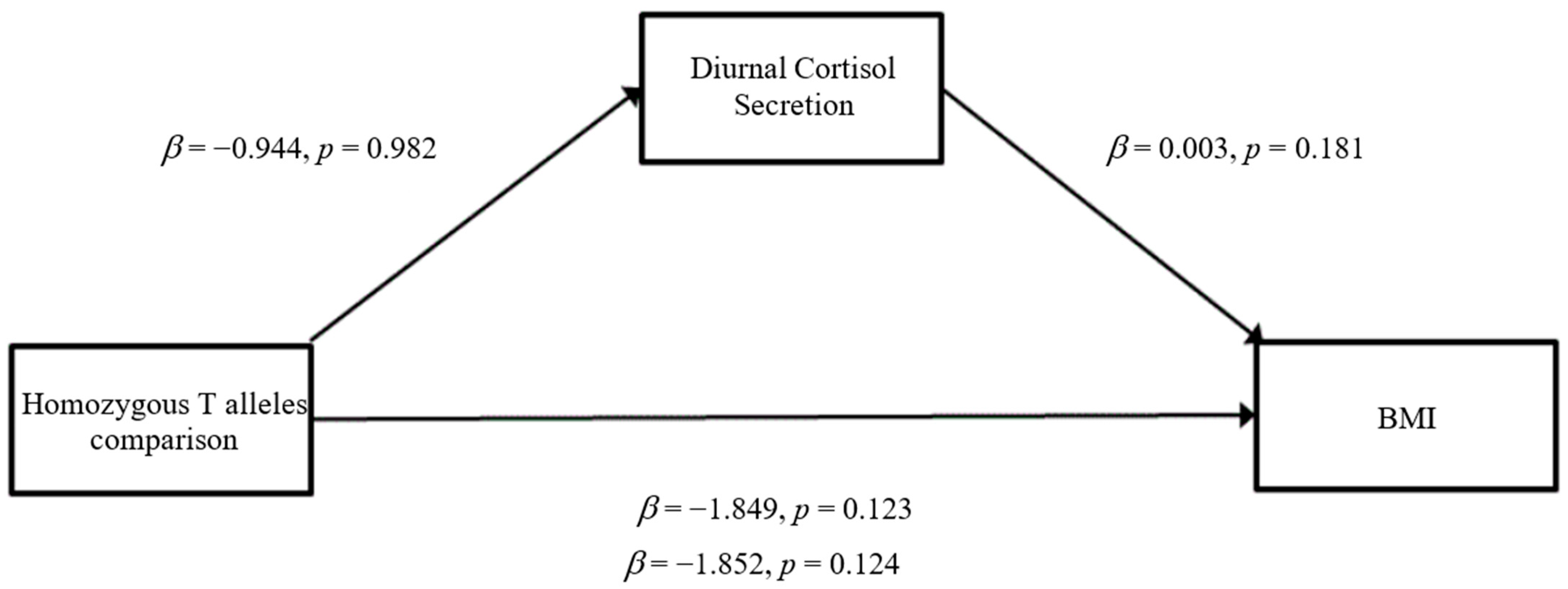
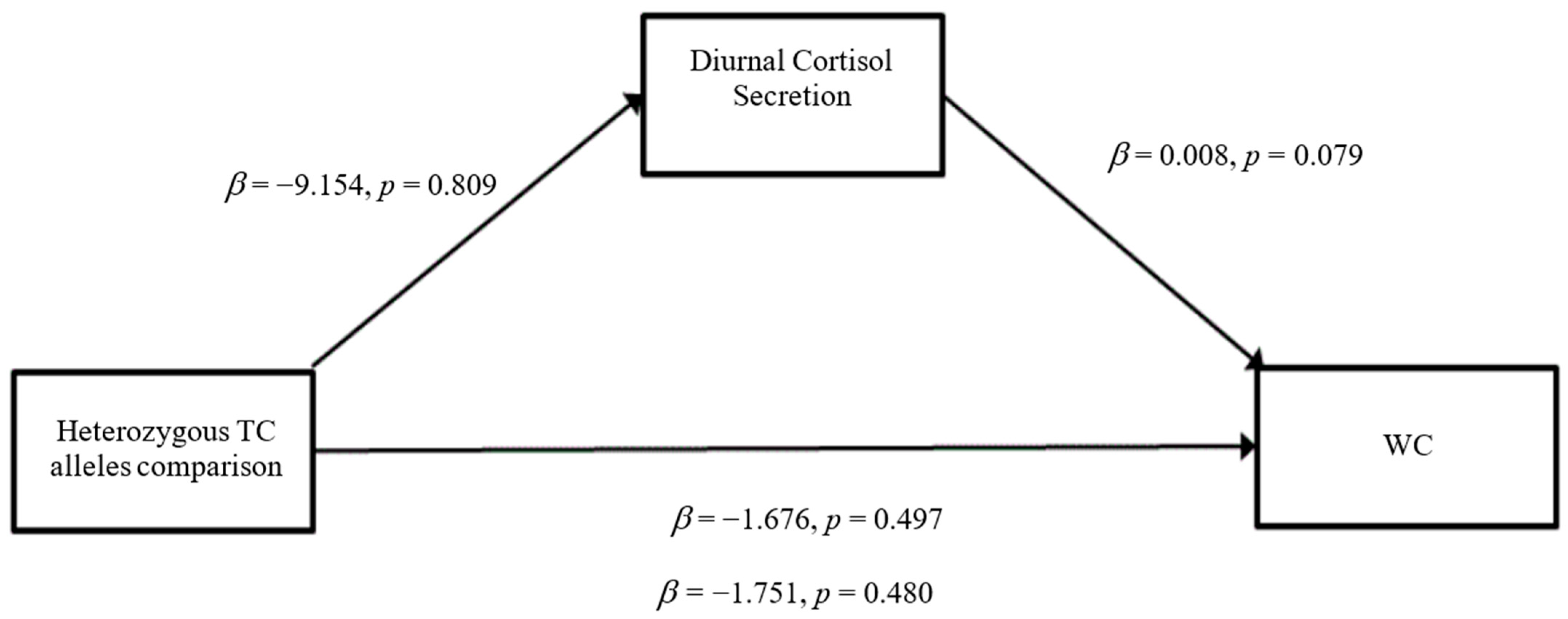
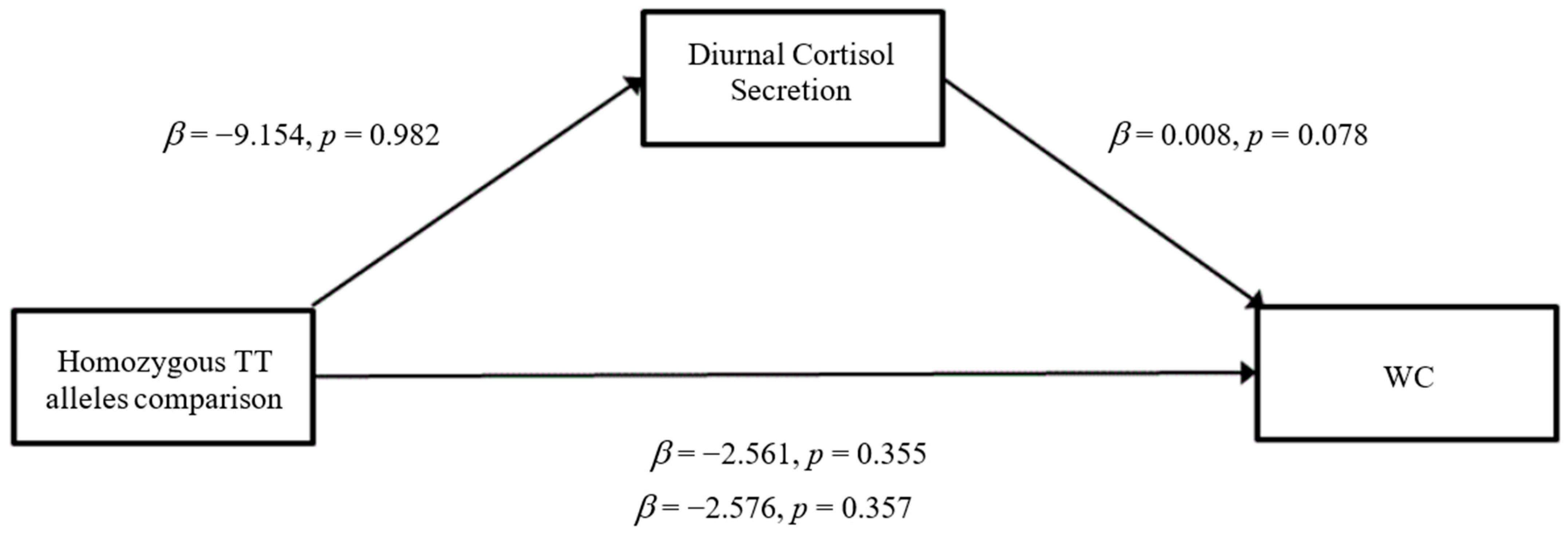
3.3. Mediation Analyses
4. Discussion
Limitations and Future Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Stierman, B.; Afful, J.; Carroll, M.D.; Chen, T.; Davy, O.; Fink, S.; Fryar, C.D.; Gu, Q.; Hales, C.M.; Hughes, J.P.; et al. National Health and Nutrition Examination Survey 2017–March 2020 Prepandemic Data Files Development of Files and Prevalence Estimates for Selected Health Outcomes. Natl. Health Stat. Rep. 2021, 158, 1–21. [Google Scholar] [CrossRef]
- Flegal, K.M.; Kit, B.K.; Orpana, H.; Graubard, B.I. Association of all-cause mortality with overweight and obesity using standard body mass index categories: A systematic review and meta-analysis. JAMA 2013, 309, 71–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, S.G.; Dunn, W.B.; Banerjee, M.; Brown, M.; Broadhurst, D.I.; Goodacre, R.; Cooper, G.J.S.; Kell, D.B.; Cruickshank, J.K. Evidence that multiple defects in lipid regulation occur before hyperglycemia during the prodrome of type-2 diabetes. PLoS ONE 2014, 9, e103217. [Google Scholar]
- Hursting, S.D.; Dunlap, S.M. Obesity, metabolic dysregulation, and cancer: A growing concern and an inflammatory (and microenvironmental) issue. Ann. N. Y. Acad. Sci. 2012, 1271, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Cawley, J.; Meyerhoefer, C. The medical care costs of obesity: An instrumental variables approach. J. Health Econ. 2012, 31, 219–230. [Google Scholar] [CrossRef]
- Danese, A.; Tan, M. Childhood maltreatment and obesity: Systematic review and meta-analysis. Mol. Psychiatry 2014, 19, 544–554. [Google Scholar] [CrossRef]
- Soares, A.L.G.; Matijasevich, A.; Menezes, A.; Assunção, M.C.; Wehrmeister, F.C.; Howe, L.D.; Gonçalves, H. Adverse Childhood Experiences (ACEs) and Adiposity in Adolescents: A Cross-Cohort Comparison. Obesity 2018, 26, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Bentley, T.; Widom, C.S. A 30-year follow-up of the effects of child abuse and neglect on obesity in adulthood. Obesity 2009, 17, 1900–1905. [Google Scholar] [CrossRef]
- Chen, E.; Brody, G.H.; Miller, G.E. Childhood close family relationships and health. Am. Psychol. 2017, 72, 555–566. [Google Scholar] [CrossRef]
- Gu, Z.; Li, D.; He, H.; Wang, J.; Hu, X.; Zhang, P.; Hong, Y.; Liu, B.; Zhang, L.; Ji, G. Body mass index, waist circumference, and waist-to-height ratio for prediction of multiple metabolic risk factors in Chinese elderly population. Sci. Rep. 2018, 8, 385. [Google Scholar] [CrossRef] [Green Version]
- Van Pelt, R.E.; Evans, E.M.; Schechtman, K.B.; Ehsani, A.A.; Kohrt, W.M. Waist circumference vs body mass index for prediction of disease risk in postmenopausal women. IJO 2001, 25, 1183. [Google Scholar] [CrossRef] [PubMed]
- Lehman, B.J.; Taylor, S.E.; Kiefe, C.I.; Seeman, T.E. Relation of childhood socioeconomic status and family environment to adult metabolic functioning in the CARDIA study. Psychosom. Med. 2005, 67, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Weidner, G.; Hutt, J.; Connor, S.L.; Mendell, N.R. Family stress and coronary risk in children. Psychosom. Med. 1992, 54, 471–479. [Google Scholar] [CrossRef]
- Repetti, R.L.; Taylor, S.E.; Seeman, T.E. Risky families: Family social environments and the mental and physical health of offspring. Psychol. Bull. 2002, 128, 330. [Google Scholar] [CrossRef]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. DCNS 2006, 8, 383. [Google Scholar] [CrossRef]
- McEwen, B.S.; Seeman, T. Protective and damaging effects of mediators of stress. Elaborating and testing the concepts of allostasis and allostatic load. Ann. N. Y. Acad. Sci. 1999, 896, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.E.; Chen, E.; Parker, K.J. Psychological stress in childhood and susceptibility to the chronic diseases of aging: Moving toward a model of behavioral and biological mechanisms. Psychol. Bull. 2011, 137, 959. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.F.; Arbel, R.; Shapiro, L.S.; Han, S.C.; Margolin, G. Does the cortisol awakening response link childhood adversity to adult BMI? Health Psychol. 2018, 37, 526. [Google Scholar] [CrossRef]
- Mutch, D.M.; Clément, K. Unraveling the genetics of human obesity. PLoS Genet. 2006, 2, e188. [Google Scholar] [CrossRef] [Green Version]
- Kao, S.L.; Chong, S.S.; Lee, C.G. The role of single nucleotide polymorphisms (SNPs) in understanding complex disorders and pharmacogenomics. Ann. Acad. Med. Singap. 2000, 29, 376–382. [Google Scholar]
- Jorgensen, T.J.; Ruczinski, I.; Kessing, B.; Smith, M.W.; Shugart, Y.Y.; Alberg, A.J. Hypothesis-driven candidate gene association studies: Practical design and analytical considerations. Am. J. Epidemiol. 2009, 170, 986–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patnala, R.; Clements, J.; Batra, J. Candidate gene association studies: A comprehensive guide to useful in silico tools. BMC Genet. 2013, 14, 39. [Google Scholar] [CrossRef] [PubMed]
- Tabor, H.K.; Risch, N.J.; Myers, R.M. Candidate-gene approaches for studying complex genetic traits: Practical considerations. Nat. Rev. Genet. 2002, 3, 391. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Padmanabhan, S.; Wolfarth, B.; Fuku, N.; Lucia, A.; Ahmetov, I.I.; Cieszczyk, P.; Collins, M.; Eynon, N.; Klissouras, V.; et al. Genomics of elite sporting performance: What little we know and necessary advances. In Advances in Genetics; Academic Press: Cambridge, MA, USA, 2013; Volume 84, pp. 123–149. [Google Scholar]
- Zhu, M.; Zhao, S. Candidate gene identification approach: Progress and challenges. Int. J. Biol. Sci. 2007, 3, 420–427. [Google Scholar] [CrossRef] [Green Version]
- Rosmond, R. Association studies of genetic polymorphisms in central obesity: A critical review. IJO 2003, 27, 1141. [Google Scholar] [CrossRef] [Green Version]
- Björntorp, P.E.R. Body fat distribution, insulin resistance, and metabolic diseases. Nutrition 1997, 13, 795–803. [Google Scholar] [CrossRef]
- Rosmond, R.; Bouchard, C.; Björntorp, P. Allelic variants in the GABA A α6 receptor subunit gene (GABRA6) is associated with abdominal obesity and cortisol secretion. IJO 2002, 26, 938. [Google Scholar]
- Hardy, O.T.; Czech, M.P.; Corvera, S. What causes the insulin resistance underlying obesity? Curr. Opin. Endocrinol. Diabetes Obes. 2012, 19, 81. [Google Scholar] [CrossRef] [Green Version]
- Merrick, M.T.; Ford, D.C.; Ports, K.A.; Guinn, A.S.; Chen, J.; Klevens, J.; Metzler, M.; Jones, C.M.; Simon, T.R.; Daniel, V.M.; et al. Vital Signs: Estimated Proportion of Adult Health Problems Attributable to Adverse Childhood Experiences and Implications for Prevention—25 States, 2015–2017. MMWR 2019, 68, 999–1005. [Google Scholar] [CrossRef] [Green Version]
- Kessler, R.C.; Davis, C.G.; Kendler, K.S. Childhood adversity and adult psychiatric disorder in the US National Comorbidity Survey. Psychol. Med. 1997, 27, 1101–1119. [Google Scholar] [CrossRef]
- Shastry, B.S. SNP alleles in human disease and evolution. J. Hum. Genet. 2002, 47, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, W.J.; Casselbrant, M.L.; Li-Korotky, H.S.; Cullen Doyle, A.P.; Lo, C.Y.; Turner, R.; Cohen, S. The interleukin 6− 174 C/C genotype predicts greater rhinovirus illness. J. Infect. Dis. 2010, 201, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Janicki-Deverts, D.; Cohen, S.; Turner, R.B.; Doyle, W.J. Basal salivary cortisol secretion and susceptibility to upper respiratory infection. Brain Behav. Immun. 2016, 53, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.E.; Lerner, J.S.; Sage, R.M.; Lehman, B.J.; Seeman, T.E. Early environment, emotions, responses to stress, and health. J. Pers. 2004, 72, 1365–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, S.E.; Way, B.M.; Welch, W.T.; Hilmert, C.J.; Lehman, B.J.; Eisenberger, N.I. Early family environment, current adversity, the serotonin transporter promoter polymorphism, and depressive symptomatology. Biol. Psychiatry 2006, 60, 671–676. [Google Scholar] [CrossRef]
- Nayman, S.; Jones, E.J.; Smyth, J.M.; Schreier, H.M.C. Associations of childhood and adult adversity with daily experiences in adulthood. Stress Health 2021, 38, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Dressendörfer, R.A.; Kirschbaum, C.; Rohde, W.; Stahl, F.; Strasburger, C.J. Synthesis of a cortisol-biotin conjugate and evaluation as a tracer in an immunoassay for salivary cortisol measurement. J. Steroid Biochem. Mol. Biol. 1992, 43, 683–692. [Google Scholar] [CrossRef]
- Chin, B.; Murphy, M.L.; Janicki-Deverts, D.; Cohen, S. Marital status as a predictor of diurnal salivary cortisol levels and slopes in a community sample of healthy adults. Psychoneuroendocrinology 2017, 78, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge: New York, NY, USA, 1988. [Google Scholar]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G* Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Qiu, W. Powermediation: Power/Sample Size Calculation for Mediation Analysis [Computer Software Manual]. (R Package Version 0.1.3). 2015. Available online: http://CRAN.R-project.org/package=powerMediation (accessed on 3 March 2020).
- Vittinghoff, E.; Sen, Ś.; McCulloch, C.E. Sample size calculations for evaluating mediation. Stat. Med. 2009, 28, 541–557. [Google Scholar] [CrossRef]
- Hayes, A.F. Introduction to Mediation, Moderation, and Conditional Process Analysis: A Regression-Based Approach; Guilford Press: New York, NY, USA, 2013. [Google Scholar]
- Björntorp, P.; Holm, G.; Rosmond, R. Hypothalamic arousal, insulin resistance and type 2 diabetes mellitus. Diabet. Med. 1999, 16, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Vimaleswaran, K.S.; Tachmazidou, I.; Zhao, J.H.; Hirschhorn, J.N.; Dudbridge, F.; Loos, R.J. Candidate genes for obesity-susceptibility show enriched association within a large genome-wide association study for BMI. Hum. Mol. Genet. 2012, 21, 4537–4542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alghamdi, J.; Padmanabhan, S. Fundamentals of complex trait genetics and association studies. In Handbook of Pharmacogenomics and Stratified Medicine; Padmanabhan, S., Ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 235–257. [Google Scholar]
- Champaneri, S.; Xu, X.; Carnethon, M.R.; Bertoni, A.G.; Seeman, T.; DeSantis, A.S.; Roux, A.D.; Shrager, S.; Golden, S.H. Diurnal salivary cortisol is associated with body mass index and waist circumference: The Multiethnic Study of Atherosclerosis. Obesity 2013, 21, E56–E63. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Chandola, T.; Brunner, E.; Kivimaki, M. A nonlinear relationship of generalized and central obesity with diurnal cortisol secretion in the Whitehall II study. J. Clin. Endocr. 2010, 95, 4415–4423. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| 1. Age (years) | −0.114 | −0.007 | 0.332 ** | 0.342 ** | −0.108 | 0.055 | |
| 2. Risky Early Family Environment | −0.168 * | 0.108 | 0.083 | 0.306 ** | −0.080 | ||
| 3. GABRA6 | −0.118 | −0.099 | 0.121 | 0.068 | |||
| 4. Body Mass Index (kg/m2) | 0.894 ** | −0.112 | −0.082 | ||||
| 5. Waist Circumference (cm) | −0.111 | −0.017 | |||||
| 6. Depressed Affect | −0.153 * | ||||||
| 7. Diurnal Cortisol (nmol/L) | |||||||
| Mean | 30.13 | 28.23 | 7.28 | 27.46 | 88.93 | 0.957 | 3.70 |
| Standard Deviation | 10.85 | 10.24 | 1.82 | 6.49 | 15.72 | 1.07 | 0.181 |
| Outcome Variable BMI | β (SE) | t | p-Value |
|---|---|---|---|
| Model 1: Risky Early Family Environment | 0.174 (0.042) | 2.637 | 0.01 |
| Age | 0.307 (0.038) | 4.814 | 0.00 |
| Race | −0.160 (0.875) | −2.519 | 0.01 |
| Sex | 0.151 (0.823) | 2.406 | 0.02 |
| Depressed Affect | 0.139 (0.399) | −2.118 | 0.04 |
| Model 2: GABRA6 | −0.066 (1.374) | −1.051 | 0.30 |
| Age | 0.413 (0.090) | 6.606 | 0.00 |
| Race | −0.144 (2.091) | −2.270 | 0.02 |
| Sex | −0.098 (1.961) | −1.574 | 0.12 |
| Depressed Affect | −0.041 (0.912) | −0.657 | 0.51 |
| Outcome Variable WC | |||
| Model 1: Risky Early Family Environment | 0.150 (0.099) | 2.335 | 0.02 |
| Age | 0.420 (0.090) | 6.778 | 0.00 |
| Race | −0.134 (2.063) | −2.163 | 0.03 |
| Sex | −0.102 (1.939) | −1.672 | 0.10 |
| Depressed Affect | −0.101 (0.941) | −1.570 | 0.12 |
| Model 2: GABRA6 | −0.105 (0.587) | −1.622 | 0.11 |
| Age | 0.295 (0.038) | 4.582 | 0.00 |
| Race | −0.165 (0.893) | −2.524 | 0.01 |
| Sex | 0.158 (0.837) | 2.473 | 0.01 |
| Depressed Affect | −0.072 (0.389) | −1.117 | 0.27 |
| Outcome Variable BMI | β (SE) | t | p |
|---|---|---|---|
| Risky Early Family Environment | 0.166 (0.042) | 2.532 | 0.01 |
| GABRA6 | −0.125 (0.936) | −1.968 | 0.05 |
| Age | 0.308 (0.038) | 4.869 | 0.00 |
| Race | 0.142 (0.879) | 2.223 | 0.03 |
| Sex | 0.164 (0.822) | 2.615 | 0.00 |
| Depressed Affect | −0.120 (0.401) | −1.807 | 0.07 |
| Outcome Variable WC | |||
| Risky Early Family Environment | 0.145 (0.099) | 2.260 | 0.03 |
| GABRA6 | −0.076 (2.217) | −1.215 | 0.23 |
| Age | 0.421 (0.090) | 4.921 | 0.00 |
| Race | 0.123 (2.082) | 1.966 | 0.05 |
| Sex | −0.094 (1.94) | −1.539 | 0.13 |
| Depressed Affect | −0.089 (0.951) | −1.368 | 0.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huizar, Y.P.; Cundiff, J.M.; Schmidt, A.T.; Cribbet, M.R. Risky Early Family Environment and Genetic Associations with Adult Metabolic Dysregulation. Int. J. Environ. Res. Public Health 2022, 19, 14032. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph192114032
Huizar YP, Cundiff JM, Schmidt AT, Cribbet MR. Risky Early Family Environment and Genetic Associations with Adult Metabolic Dysregulation. International Journal of Environmental Research and Public Health. 2022; 19(21):14032. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph192114032
Chicago/Turabian StyleHuizar, Yazmine P., Jenny M. Cundiff, Adam T. Schmidt, and Matthew R. Cribbet. 2022. "Risky Early Family Environment and Genetic Associations with Adult Metabolic Dysregulation" International Journal of Environmental Research and Public Health 19, no. 21: 14032. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph192114032