
Detraining Effects of COVID-19 Pandemic on Physical Fitness, Cytokines, C-Reactive Protein and Immunocytes in Men of Various Age Groups



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Measurement Methods
2.2.1. Physical Fitness Measure
Body Composition
Flexibility
Muscle Strength
Muscle Endurance
Cardiorespiratory Endurance
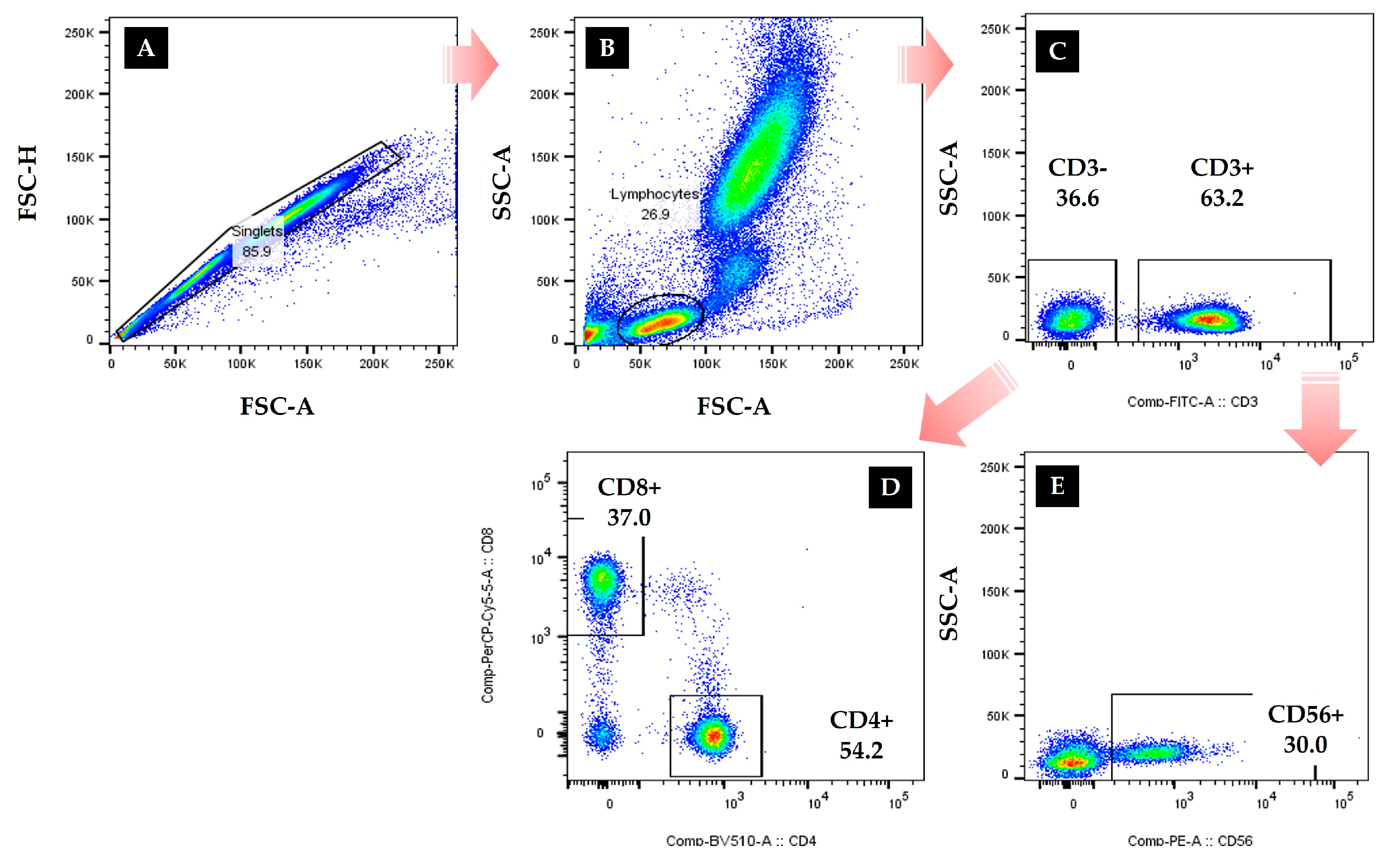
2.2.2. Immunocytes Measures
2.2.3. Cytokines and CRP Measures
2.3. Data Process and Statistical Analyses
3. Results
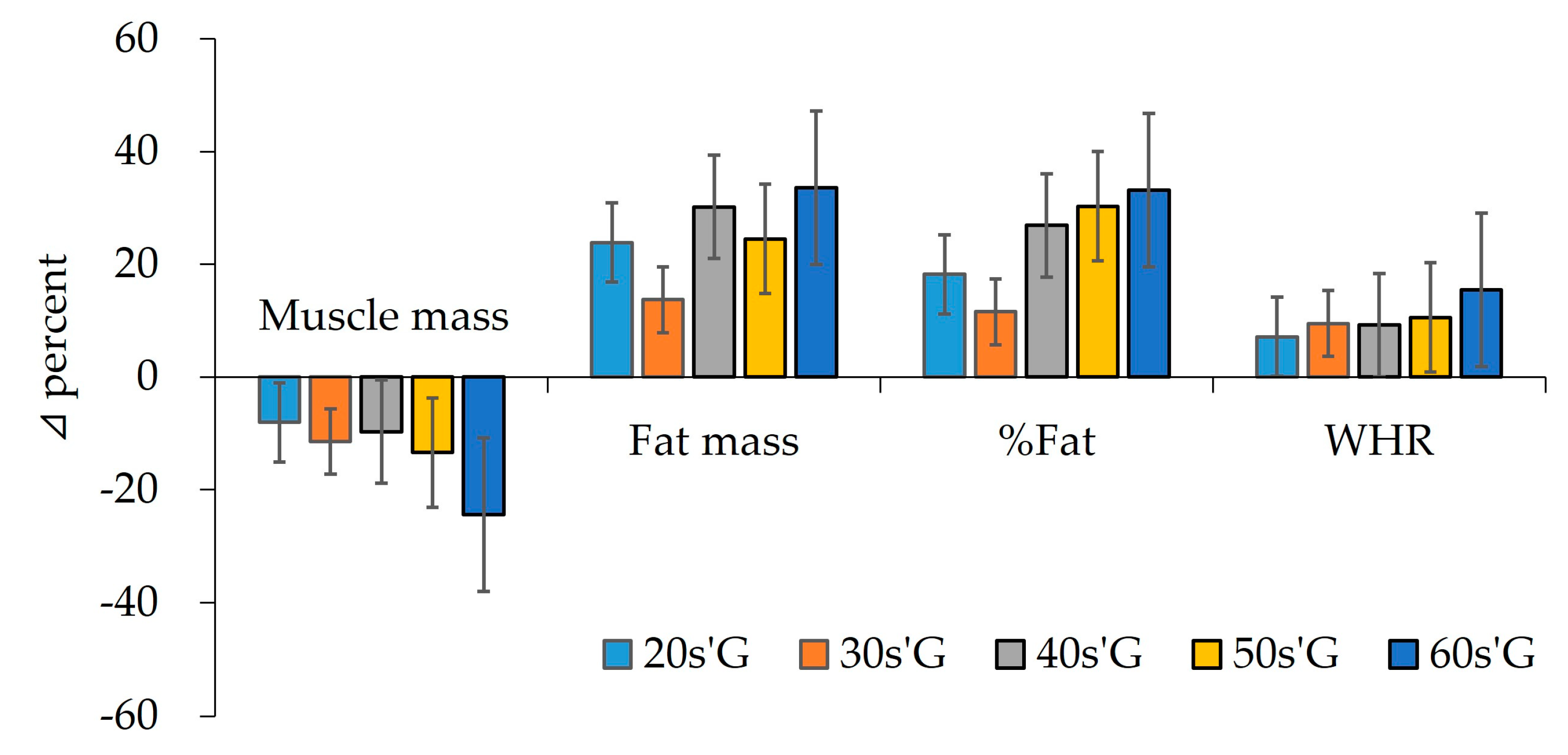
3.1. Analysis of Demographic Variables and Body Composition
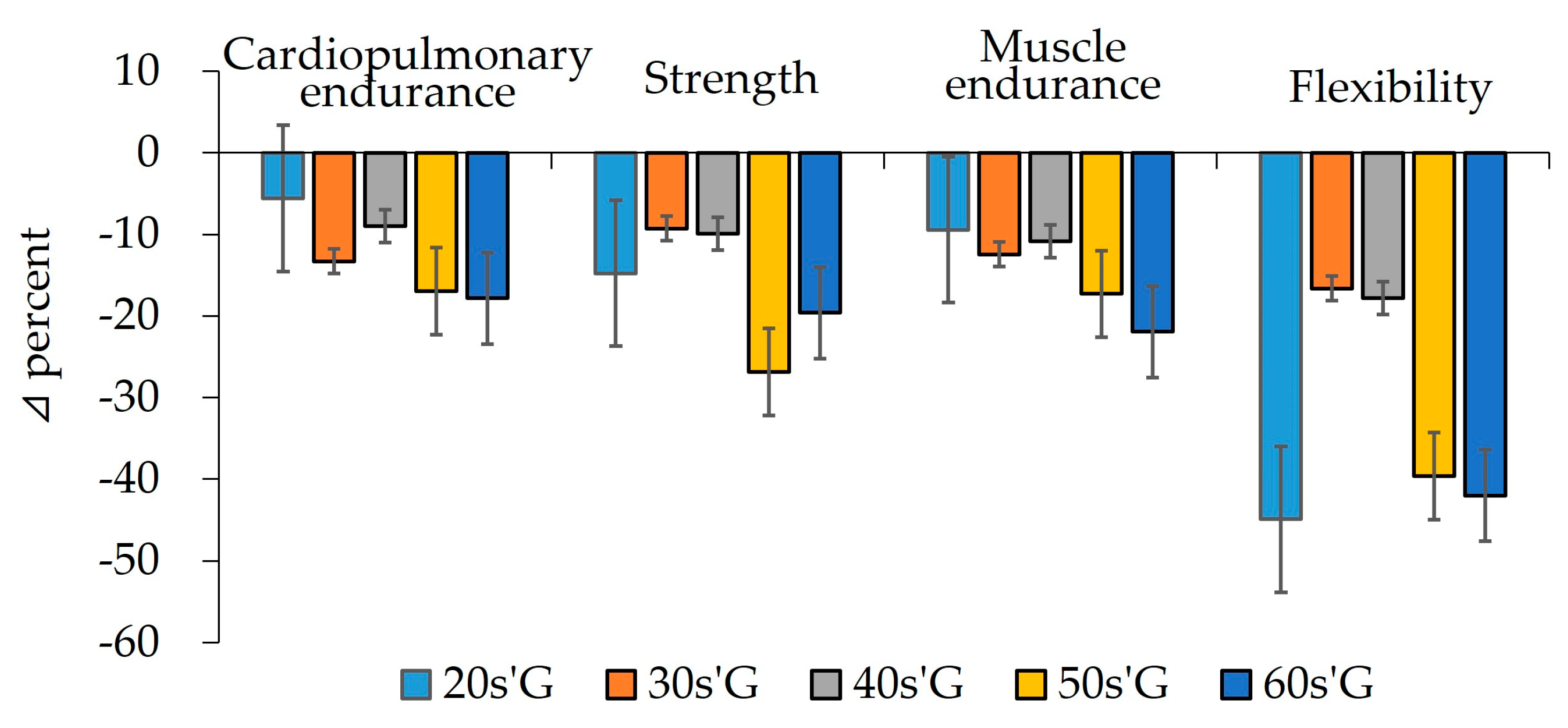
3.2. Comparisons of Physical Fitness
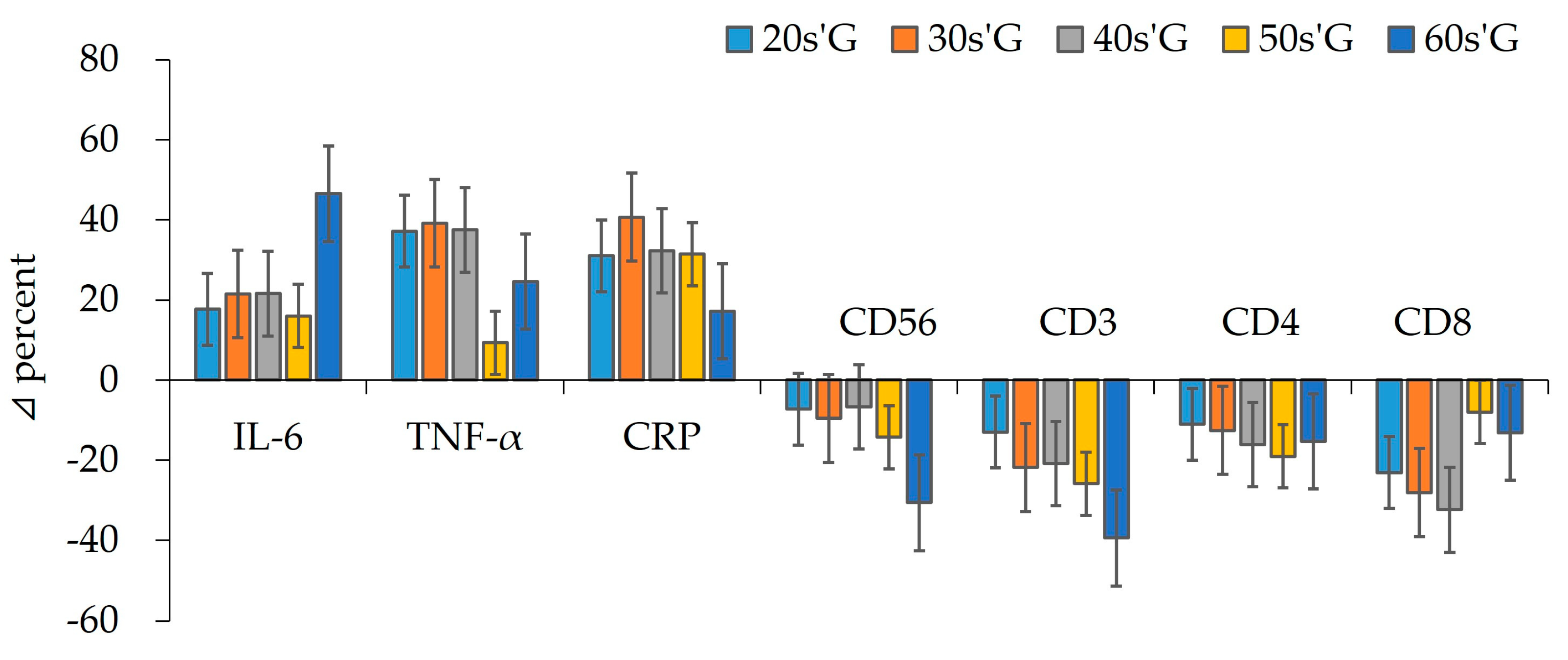
3.3. Comparison Results of Immunocytes
3.4. Comparison Results of Cytokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Viana, R.B.; de Lira, C.A.B. Exergames as coping strategies for anxiety disorders during the COVID-19 quarantine period. Games Health J. 2020, 9, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Lee, K.S.; Heo, S.J.; Jee, Y.S. Effects of high intensity plank exercise on physical fitness and immunocyte function in a middle-aged man: A case report. Medicina 2021, 57, 845. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Heo, S.J.; Hyun, G.S.; Park, S.K.; Cha, J.Y.; Jee, Y.S. Elbow plank exercise improves immunocyte function and physical fitness in an elderly male: A case study. J. Men’s Health 2021, 1–8. [Google Scholar] [CrossRef]
- Finn, O.J. Cancer immunology. N. Engl. J. Med. 2008, 358, 2704–2715. [Google Scholar] [CrossRef]
- Hansson, G.K.; Nilsson, J. Introduction: Atherosclerosis as inflammation: A controversial concept becomes accepted. J. Intern. Med. 2008, 263, 462–463. [Google Scholar] [CrossRef]
- Libby, P. The molecular mechanisms of the thrombotic complications of atherosclerosis. J. Intern. Med. 2008, 263, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Chou, M.Y.; Hartvigsen, K.; Hansen, L.F.; Fogelstrand, L.; Shaw, P.X.; Boullier, A.; Binder, C.J.; Witztum, J.L. Oxidation-specific epitopes are important targets of innate immunity. J. Intern. Med. 2008, 263, 479–488. [Google Scholar] [CrossRef]
- Petersen, A.M.W.; Pedersen, B.K. The anti-inflammatory effect of exercise. J. Appl. Physiol. 2005, 98, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Duramad, P.; Holland, N.T. Biomarkers of immunotoxicity for environmental and public health research. Int. J. Environ. Res. Public Health 2011, 8, 1388–1401. [Google Scholar] [CrossRef] [Green Version]
- D’Andrea Greve, J.M.; Brech, G.C.; Quintana, M.; de Seixas Soares, A.L.; Alonso, A.C. Impacts of COVID-19 on the immune, neuromuscular, and musculoskeletal systems and rehabilitation. Rev. Bras. Med. Esporte 2020, 26, 285–288. [Google Scholar] [CrossRef]
- Descotes, J. Methods of evaluating immunotoxicity. Expert Opin. Drug Metab. Toxicol. 2006, 2, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Descotes, J.; Testud, F. Toxicovigilance: A new approach for the hazard identification and risk assessment of toxicants in human beings. Toxicol. Appl. Pharmacol. 2005, 207, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Park, J.; Yoo, J.; Jee, Y.S. Effect of playing soccer on stress, sociality, and physical fitness in alienated youth: A retrospective study. J. Exerc. Rehabil. 2020, 16, 154–161. [Google Scholar] [CrossRef]
- Jee, Y.S. The efficacy and safety of whole-body electromyostimulation in applying to human body: Based from graded exercise test. J. Exerc. Rehabil. 2018, 14, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Min, S.; Park, S.H.; Yoo, J.; Jee, Y.S. Influence of isometric exercise combined with electromyostimulation on inflammatory cytokine levels, muscle strength, and knee joint function in elderly women with early knee osteoarthritis. Front. Physiol. 2021, 12, 688260. [Google Scholar] [CrossRef]
- Cohen, J. A power primer. Psychol. Bull. 1992, 112, 155–159. [Google Scholar] [CrossRef]
- Costill, D.L.; Fink, W.J.; Hargreaaves, M.; King, D.S.; Thomas, R.; Fielding, R. Metabolic characteristics of skeletal muscle during detraining from competitive swimming. Med. Sci. Sports Exerc. 1985, 17, 339–343. [Google Scholar] [CrossRef]
- Fox, E.L.; Mathews, D.K. The Physiological Basis of Physical Education and Athletics; Saunders College Publishing: Philadelphia, PA, USA, 1981. [Google Scholar]
- Saltin, B.; Rowell, L.B. Functional adaptations to physical activity and inactivity. Fed. Proc. 1980, 39, 1506–1513. [Google Scholar]
- Graves, J.E.; Pollock, M.L.; Leggett, S.H.; Braith, R.W.; Carpenter, D.M.; Bishop, L.E. Effect of reduced training frequency on muscular strength. Int. J. Sports Med. 1988, 9, 316–319. [Google Scholar] [CrossRef]
- Hakkinen, K.; Komi, P.V. Changes in electrical and mechanical behavior of leg extensor muscles during heavy resistance strength training, Scand. J. Sports Sci. 1985, 7, 55–64. [Google Scholar]
- Fleck, S.J. Detraining: Its effects on endurance and strength. Strength Cond. J. 1994, 16, 22–28. [Google Scholar] [CrossRef]
- Houston, M.E.; Bentzen, H.; Larsen, H. Interrelationships between skeletal muscle adaptations and performance as studied by detraining and retraining. Acta Physiol. Scand. 1979, 105, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Ogden, L.G.; Stroebele, N.; Wyatt, H.R.; Catenacci, V.A.; Peters, J.C.; Stuht, J.; Wing, R.R.; Hill, J.O. Cluster analysis of the national weight control registry to identify distinct subgroups maintaining successful weight loss. Obesity 2012, 20, 2039–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutin, B.; Barbeau, P.; Litaker, M.S.; Ferguson, M.; Owens, S. Heart rate variability in obese children: Relations to total body and visceral adiposity, and changes with physical training and detraining. Obes. Res. 2000, 8, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Roffey, D.M.; Budiansky, A.; Coyle, M.J.; Wai, E.K. Obesity and low back pain: Is there a weight of evidence to support a positive relationship? Curr. Obes. Rep. 2013, 2, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Thompson, D.; Karpe, F.; Lafontan, M.; Frayn, K. Physical activity and exercise in the regulation of human adipose tissue physiology. Physiol. Rev. 2012, 92, 157–191. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, L.J.; Barbagallo, M. The cardiometabolic syndrome and sarcopenic obesity in older persons. J. Cardiometab. Syndr. 2007, 2, 183–189. [Google Scholar] [CrossRef]
- Jensen, G.L. Inflammation: Roles in aging and sarcopenia. J. Parenter. Enteral. Nutr. 2008, 32, 656–659. [Google Scholar] [CrossRef]
- Oppenheim, J.J. Cytokines: Past, present, and future. Int. J. Hematol. 2001, 74, 3–8. [Google Scholar] [CrossRef]
- Sharif, K.; Watad, A.; Bragazzi, N.L.; Lichtbroun, M.; Amital, H.; Shoenfeld, Y. Physical activity and autoimmune diseases: Get moving and manage the disease. Autoimmun. Rev. 2018, 17, 53–72. [Google Scholar] [CrossRef]
- Conti, P.; Ronconi, G.; Caraffa, A.L.; Gallenga, C.E.; Ross, R.; Frydas, I.; Kritas, S.K. Induction of proinflammatory cytokines (IL-1 and IL-6) and lung inflammation by Coronavirus-19 (COVI-19 or SARS-CoV-2): Anti-inflammatory strategies. J. Biol. Regul. Homeost. Agents 2020, 34, 11–15. [Google Scholar] [CrossRef]
- Guo, W.; Li, M.; Dong, Y.; Zhou, H.; Zhang, Z.; Tian, C.; Hu, D. Diabetes is a risk factor for the progression and prognosis of COVID-19. Diabetes Metab. Res. Rev. 2020, 36, e3319. [Google Scholar] [CrossRef] [Green Version]
- Morley, J.E.; Silver, A.J. Anorexia in the elderly. Neurobiol. Aging 1988, 9, 9–16. [Google Scholar] [CrossRef]
- Thomas, D.R. Sarcopenia. Clin. Geriatr. Med. 2010, 26, 331–346. [Google Scholar] [CrossRef]
- Rodriguez-Mañas, L.; de Carvalho, I.A.; Bhasin, S.; Bischoff-Ferrari, H.A.; Cesari, M.; Evans, W.; Hare, J.M.; Pahor, M.; Parini, A.; Rolland, Y.; et al. ICFSR task force perspective on biomarkers for sarcopenia and frailty. J. Frailty Aging 2020, 9, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Leeuwenburgh, C.; Gurley, C.M.; Strotman, B.A.; Dupont-Versteegden, E.E. Age-related differences in apoptosis with disuse atrophy in soleus muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1288–R1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, B.K.; Hoffman-Goetz, L. Exercise and the immune system: Regulation, integration, and adaptation. Physiol. Rev. 2000, 80, 1055–1081. [Google Scholar] [CrossRef] [Green Version]
- Pistilli, E.E.; Jackson, J.R.; Always, S.E. Death receptor-associated pro-apoptotic signaling in aged skeletal muscle. Apoptosis 2006, 11, 2115–2126. [Google Scholar] [CrossRef] [PubMed]
- Jee, Y.S. Exercise is an antigen for vaccination: First series of scientific evidence. J. Exerc. Rehabil. 2019, 15, 339–340. [Google Scholar] [CrossRef] [Green Version]
- Jee, Y.S. How much exercise do we need to improve our immune system? Second series of scientific evidence. J. Exerc. Rehabil. 2020, 16, 113–114. [Google Scholar] [CrossRef]
- Jee, Y.S. Influences of acute and/or chronic exercise on human immunity: Third series of scientific evidence. J. Exerc. Rehabil. 2020, 16, 205–206. [Google Scholar] [CrossRef]
- Jee, Y.S. Physical exercise for strengthening innate immunity during COVID-19 pandemic: 4th series of scientific evidence. J. Exerc. Rehabil. 2020, 16, 383–384. [Google Scholar] [CrossRef]
- Jee, Y.S. Acquired immunity and moderate physical exercise: 5th series of scientific evidence. J. Exerc. Rehabil. 2021, 17, 2–3. [Google Scholar] [CrossRef]
- Grant, R.W.; Mariani, R.A.; Vieira, V.J.; Fleshner, M.; Smith, T.P.; Keylock, K.T.; Lowder, T.W.; McAuley, E.; Hu, L.; Chapman-Novakofski, K.; et al. Cardiovascular exercise intervention improves the primary antibody response to keyhole limpet hemocyanin (KLH) in previously sedentary older adults. Brain Behav. Immun. 2008, 22, 923–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keylock, K.T.; Lowder, T.; Leifheit, K.A.; Cook, M.; Mariani, R.A.; Ross, K.; Kim, K.; Chapman-Novakofski, K.; McAuley, E.; Woods, J.A. Higher antibody, but not cell-mediated, responses to vaccination in high physically fit elderly. J. Appl. Physiol. 2007, 102, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Kohut, M.L.; Arntson, B.A.; Lee, W.; Rozeboom, K.; Yoon, K.J.; Cunnick, J.E.; McElhaney, J. Moderate exercise improves antibody response to influenza immunization in older adults. Vaccine 2004, 22, 2298–2306. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.P.; Kennedy, S.L.; Fleshner, M. Influence of age and physical activity on the primary in vivo antibody and T cell-mediated responses in men. J. Appl. Physiol. 2004, 97, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Verkuilen, J.; Rosengren, K.S.; Mariani, R.A.; Reed, M.; Grubisich, S.A.; Woods, J.A. Effects of a Taiji and Qigong intervention on the antibody response to influenza vaccine in older adults. Am. J. Chin. Med. 2007, 35, 597–607. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.S.; Jan, M.S.; Chen, H.I. The effect of chronic and acute exercise on immunity in rats. Int. J. Sports Med. 1993, 14, 86–92. [Google Scholar] [CrossRef]
- Peake, J.; Nosaka, K.K.; Suzuki, K. Characterization of inflammatory responses to eccentric exercise in humans. Exerc. Immunol. Rev. 2005, 11, 64–85. [Google Scholar]
- Infante, J.R.; Peran, F.; Rayo, J.I.; Serrano, J.; Dominguez, M.L.; Garcia, L.; Roldan, A. Levels of immune cells in transcendental meditation practitioners. Int. J. Yoga 2014, 7, 147. [Google Scholar] [CrossRef] [PubMed]
- Rajbhoj, P.H.; Shete, S.U.; Verma, A.; Bhogal, R.S. Effect of yoga module on pro-inflammatory and anti-inflammatory cytokines in industrial workers of lonavla: A randomized controlled trial. J. Clin. Diagn. Res. 2015, 9, CC1–CC5. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2016, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Codella, R.; Chirico, A.; Lucidi, F.; Ferrulli, A.; La Torre, A.; Luzi, L. The immune-modulatory effects of exercise should be favorably harnessed against COVID-19. J. Endocrinol. Investig. 2021, 44, 1119–1122. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | |||||||
|---|---|---|---|---|---|---|---|
| 20s’G (n = 14; 22%) | 30s’G (n = 12; 19%) | 40s’G (n = 12; 19%) | 50s’G (n = 12; 19%) | 60s’G (n = 14; 22%) | X2 | p | |
| Age (y) | 23.29 ± 1.33 | 35.09 ± 2.07 | 44.50 ± 2.28 | 54.92 ± 2.15 | 63.64 ± 1.95 | 59.567 | 0.001 |
| Height (cm) | 174.74 ± 3.93 | 169.62 ± 3.92 | 171.80 ± 4.51 | 176.55 ± 4.05 | 166.36 ± 4.21 | 29.344 | 0.001 |
| Weight (kg) | 74.44 ± 6.97 | 73.05 ± 5.51 | 74.03 ± 9.12 | 78.28 ± 6.82 | 65.29 ± 8.14 | 16.918 | 0.002 |
| Percent fat (%) | 18.34 ± 5.51 | 17.97 ± 3.87 | 19.37 ± 4.70 | 18.54 ± 6.58 | 19.40 ± 7.28 | 1.258 | 0.869 |
| WHR | 0.84 ± 0.03 | 0.84 ± 0.02 | 0.87 ± 0.04 | 0.85 ± 0.04 | 0.84 ± 0.04 | 7.458 | 0.114 |
| Detrained periods (wk) | 45.83 ± 6.32 | 42.52 ± 5.72 | 45.93 ± 5.77 | 44.21 ± 5.24 | 46.25 ± 5.19 | 4.263 | 0.372 |
| 20s’G | 30s’G | 40s’G | 50s’G | 60s’G | F | η2 | ||
|---|---|---|---|---|---|---|---|---|
| Muscle mass | pre | 38.60 ± 7.75 | 47.68 ± 8.86 | 47.43 ± 9.27 | 33.99 ± 8.66 | 33.07 ± 10.02 | 5.545 *** | 0.587 |
| (kg) | post | 35.51 ± 7.48 | 42.23 ± 8.71 | 42.84 ± 9.73 | 29.44 ± 6.18 | 24.99 ± 6.69 | 6.744 *** | 0.683 |
| Z | −1.915 | −2.848 ** | −2.551 * | −1.969 * | −3.235 *** | |||
| Fat mass | pre | 13.71 ± 4.50 | 13.06 ± 2.69 | 14.41 ± 4.03 | 14.65 ± 5.80 | 12.41 ± 3.85 | 0.623 | 0.199 |
| (kg) | post | 16.98 ± 3.17 | 14.85 ± 3.27 | 18.76 ± 6.10 | 18.24 ± 5.69 | 16.58 ± 5.40 | 1.413 | 0.269 |
| Z | −2.731 ** | −1.913 | −2.510 * | −2.668 ** | −3.297 *** | |||
| Percent fat | pre | 18.34 ± 5.51 | 17.97 ± 3.87 | 19.37 ± 4.70 | 18.54 ± 6.58 | 19.40 ± 7.28 | 0.471 | 0.101 |
| (%) | post | 21.68 ± 4.22 | 20.05 ± 4.21 | 24.58 ± 5.65 | 24.16 ± 5.55 | 25.83 ± 8.66 | 1.082 | 0.340 |
| Z | −2.166 * | −1.423 | −2.432 * | −2.667 ** | −2.982 ** | |||
| WHR | pre | 0.84 ± 0.03 | 0.84 ± 0.02 | 0.87 ± 0.04 | 0.85 ± 0.04 | 0.84 ± 0.04 | 2.166 | 0.323 |
| post | 0.90 ± 0.03 | 0.92 ± 0.06 | 0.95 ± 0.11 | 0.94 ± 0.10 | 0.97 ± 0.17 | 0.843 | 0.248 | |
| Z | −3.118 ** | −2.938 ** | −2.654 ** | −2.867 ** | −2.937 ** | |||
| 20s’G | 30s’G | 40s’G | 50s’G | 60s’G | F | η2 | ||
|---|---|---|---|---|---|---|---|---|
| VO2max | pre | 61.37 ± 8.13 | 62.18 ± 7.59 | 55.91 ± 10.61 | 52.52 ± 5.80 | 46.87 ± 7.25 | 0.836 | 0.603 |
| (ml/kg/min) | post | 57.94 ± 6.51 | 53.91 ± 5.45 | 50.89 ± 6.83 | 43.62 ± 3.33 | 38.51 ± 6.02 | 0.519 | 0.793 |
| Z | −2.040 * | −2.756 ** | −2.118 * | −2.667 ** | −3.296 *** | |||
| Grip strength | pre | 42.08 ± 9.35 | 43.94 ± 11.68 | 39.40 ± 11.32 | 40.39 ± 7.49 | 34.16 ± 4.30 | 1.513 | 0.360 |
| (kg) | post | 35.86 ± 7.95 | 39.86 ± 11.32 | 35.48 ± 10.63 | 29.55 ± 3.15 | 27.46 ± 4.13 | 0.659 | 0.272 |
| Z | −3.045 ** | −2.580 ** | −2.432 * | −2.904 ** | −3.235 *** | |||
| Sit-ups | pre | 59.14 ± 8.21 | 57.73 ± 8.57 | 50.75 ± 7.77 | 50.58 ± 8.67 | 49.14 ± 7.05 | 0.785 | 0.477 |
| (reps) | post | 53.57 ± 7.47 | 50.55 ± 10.20 | 45.25 ± 9.18 | 41.83 ± 8.75 | 38.36 ± 6.89 | 0.445 | 0.576 |
| Z | −2.737 ** | −2.849 ** | −1.779 | −2.670 ** | −3.238 *** | |||
| Sit & reach | pre | 15.86 ± 6.32 | 13.72 ± 5.48 | 13.99 ± 7.67 | 12.88 ± 7.16 | 10.24 ± 4.26 | 0.327 | 0.305 |
| (cm) | post | 8.74 ± 4.26 | 11.44 ± 5.28 | 11.50 ± 6.51 | 7.78 ± 4.58 | 5.94 ± 3.24 | 1.934 | 0.420 |
| Z | −2.856 ** | −0.889 | −1.883 | −2.589 ** | −3.109 ** | |||
| 20s’G | 30s’G | 40s’G | 50s’G | 60s’G | F | η2 | ||
|---|---|---|---|---|---|---|---|---|
| Leucocytes | pre | 5.67 ± 11.89 | 5.93 ± 10.82 | 5.41 ± 8.11 | 5.71 ± 6.67 | 5.88 ± 6.95 | 0.814 | 0.201 |
| (×103/μL) | post | 5.96 ± 9.66 | 6.26 ± 10.16 | 6.02 ± 8.85 | 6.20 ± 7.45 | 6.35 ± 6.38 | 0.545 | 0.181 |
| Z | −1.262 | −1.511 | −1.608 | −1.571 | −2.198 * | |||
| NK cell | pre | 30.72 ± 6.18 | 30.58 ± 7.33 | 28.53 ± 4.87 | 25.09 ± 7.57 | 18.55 ± 8.69 | 1.401 | 0.569 |
| (%) | post | 28.48 ± 7.98 | 27.65 ± 6.65 | 26.61 ± 6.30 | 21.51 ± 6.97 | 12.87 ± 5.43 | 3.191* | 0.682 |
| Z | −1.100 | −1.481 | −1.067 | −1.680 | −2.497 * | |||
| CD3 | pre | 66.36 ± 8.91 | 67.09 ± 10.36 | 64.69 ± 19.67 | 64.71 ± 10.07 | 55.66 ± 16.42 | 0.528 | 0.308 |
| (%) | post | 57.74 ± 10.11 | 52.42 ± 16.14 | 51.22 ± 14.46 | 47.94 ± 18.06 | 33.74 ± 14.87 | 0.837 | 0.512 |
| Z | −2.623 ** | −2.524 * | −2.904 ** | −2.666 ** | −3.181 *** | |||
| CD4 | pre | 44.36 ± 6.00 | 42.81 ± 5.27 | 43.17 ± 9.19 | 40.03 ± 9.51 | 34.23 ± 11.03 | 0.514 | 0.418 |
| (%) | post | 39.46 ± 7.22 | 37.42 ± 7.01 | 36.22 ± 11.29 | 32.40 ± 8.74 | 28.99 ± 8.65 | 0.508 | 0.421 |
| Z | −2.299 * | −2.032 * | −1.826 | −2.320 * | −2.032 * | |||
| CD8 | pre | 35.65 ± 7.31 | 34.78 ± 8.52 | 30.88 ± 8.97 | 20.86 ± 6.70 | 18.14 ± 3.81 | 1.484 | 0.729 |
| (%) | post | 27.41 ± 7.73 | 25.01 ± 7.07 | 20.88 ± 6.75 | 19.19 ± 4.64 | 15.75 ± 3.44 | 0.583 | 0.589 |
| Z | −2.570 ** | −2.386 * | −2.070 * | −1.625 | −1.663 | |||
| 20s’G | 30s’G | 40s’G | 50s’G | 60s’G | F | η2 | ||
|---|---|---|---|---|---|---|---|---|
| IL-6 | pre | 11.58 ± 4.55 | 11.45 ± 4.53 | 15.85 ± 4.21 | 18.85 ± 4.97 | 17.77 ± 3.67 | 2.895 * | 0.590 |
| (pg/mL) | post | 13.63 ± 5.02 | 13.91 ± 4.71 | 19.27 ± 3.82 | 21.87 ± 5.40 | 26.04 ± 6.63 | 0.954 | 0.693 |
| Z | −2.040 * | −2.134 * | −2.197 * | −1.257 | −2.982 ** | |||
| TNF-α | pre | 22.68 ± 10.09 | 21.01 ± 5.46 | 22.32 ± 5.24 | 35.92 ± 9.41 | 32.15 ± 6.83 | 4.459 ** | 0.625 |
| (pg/mL) | post | 31.11 ± 12.29 | 29.24 ± 7.37 | 30.69 ± 5.30 | 39.26 ± 9.29 | 40.06 ± 12.00 | 0.963 | 0.438 |
| Z | −2.605 ** | −2.667 ** | −2.824 ** | −0.393 | −2.103 * | |||
| CRP | pre | 26.24 ± 8.81 | 29.59 ± 6.60 | 26.17 ± 5.06 | 36.14 ± 11.44 | 44.86 ± 13.17 | 2.576 * | 0.621 |
| (pg/mL) | post | 34.38 ± 9.78 | 41.63 ± 6.01 | 34.62 ± 6.89 | 47.50 ± 10.33 | 52.58 ± 13.84 | 6.067 *** | 0.608 |
| Z | −2.668 ** | −2.934 ** | −3.059 ** | −2.353 * | −2.166 * | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heo, S.-J.; Park, S.-K.; Jee, Y.-S. Detraining Effects of COVID-19 Pandemic on Physical Fitness, Cytokines, C-Reactive Protein and Immunocytes in Men of Various Age Groups. Int. J. Environ. Res. Public Health 2022, 19, 1845. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19031845
Heo S-J, Park S-K, Jee Y-S. Detraining Effects of COVID-19 Pandemic on Physical Fitness, Cytokines, C-Reactive Protein and Immunocytes in Men of Various Age Groups. International Journal of Environmental Research and Public Health. 2022; 19(3):1845. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19031845
Chicago/Turabian StyleHeo, Seung-Jae, Sang-Kyun Park, and Yong-Seok Jee. 2022. "Detraining Effects of COVID-19 Pandemic on Physical Fitness, Cytokines, C-Reactive Protein and Immunocytes in Men of Various Age Groups" International Journal of Environmental Research and Public Health 19, no. 3: 1845. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19031845