
Effects of Stepwise Temperature Shifts in Anaerobic Digestion for Treating Municipal Wastewater Sludge: A Genomic Study


Abstract
:1. Introduction
2. Materials and Methods
2.1. Inoculum and Substrates
2.2. Experimental Procedures
2.3. Deoxyribonucleic Acid (DNA) Extraction and Sequencing
2.4. Microbial Diversity Analysis
3. Results
3.1. Biogas Production under Shifted-Up Temperature
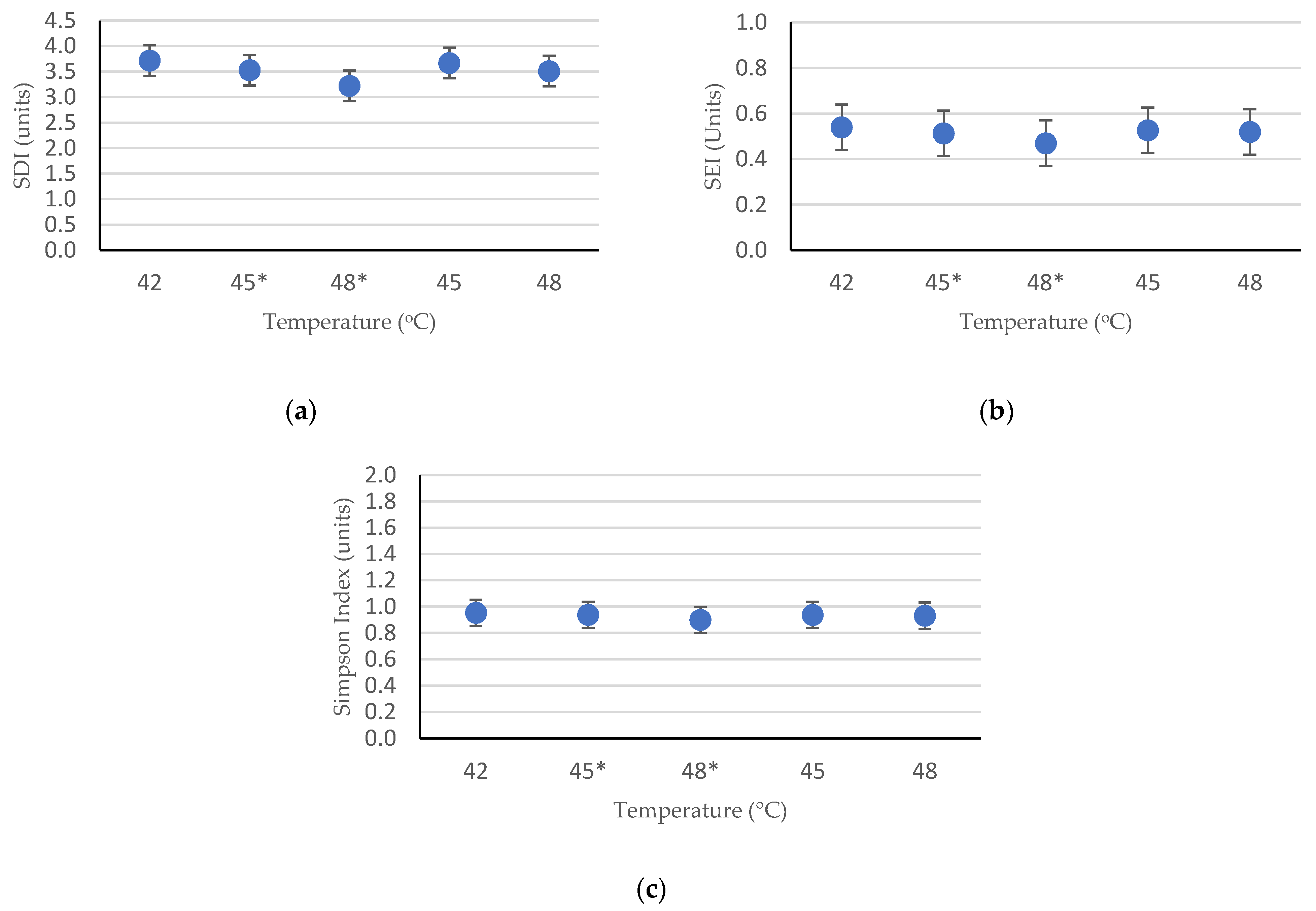
3.2. Alpha Diversity Analysis
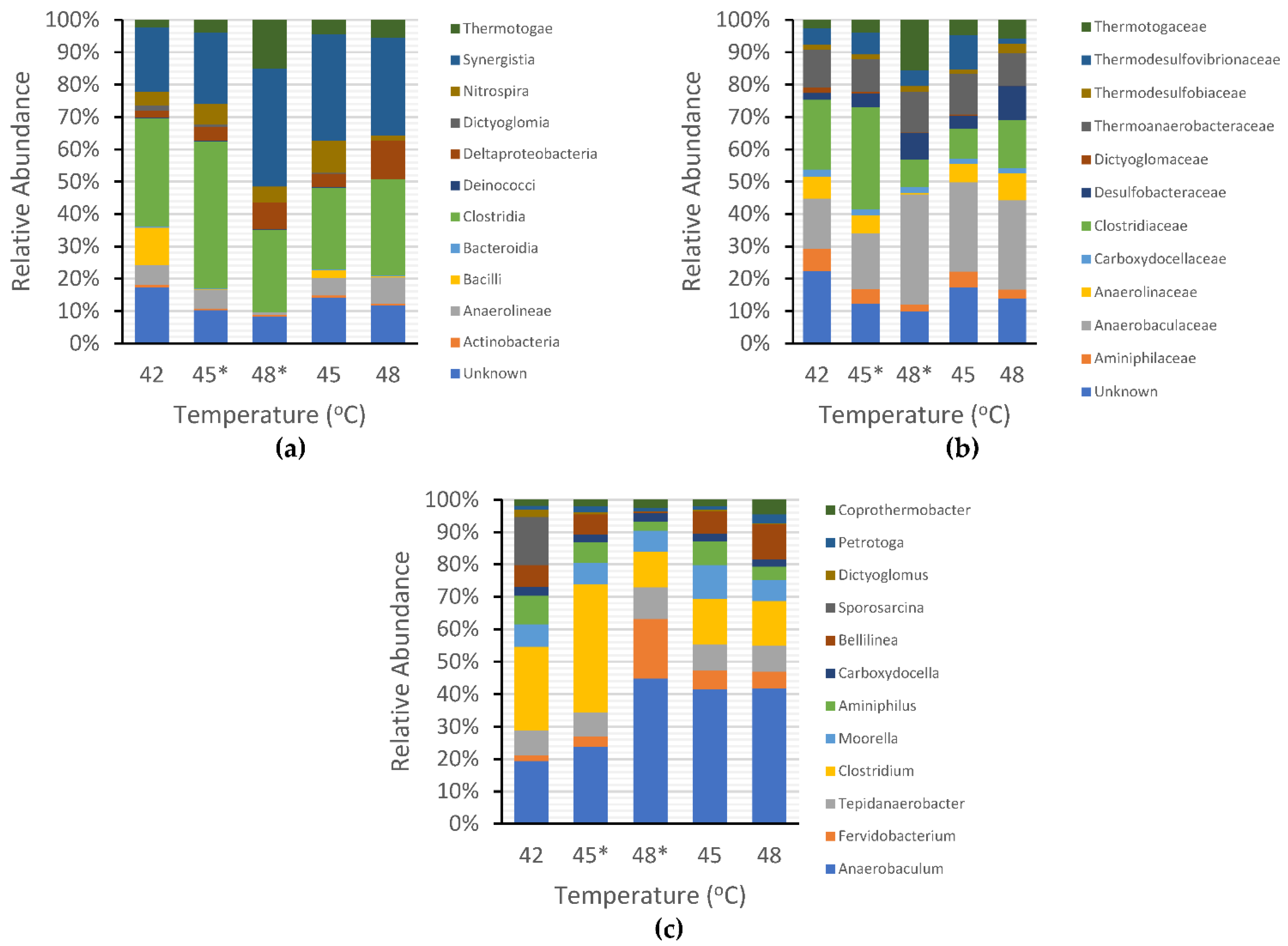
3.3. Microbial Community Structure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nika, C.E.; Gusmaroli, L.; Ghafourian, M.; Atanasova, N.; Buttiglieri, G.; Katsou, E. Nature-Based Solutions as Enablers of Circularity in Water Systems: A Review on Assessment Methodologies, Tools and Indicators. Water Res. 2020, 183, 115988. [Google Scholar] [CrossRef] [PubMed]
- Cayetano, R.D.A.; Kim, G.B.; Park, J.H.; Lee, M.J.; Kim, S.H. Anaerobic Digestion of Waste Activated Sludge Using Dynamic Membrane at Varying Substrate Concentration Reveals New Insight towards Methanogenic Pathway and Biofilm Formation. Chem. Eng. J. 2021, 423, 130249. [Google Scholar] [CrossRef]
- Mateo-Sagasta, J.; Raschid-Sally, L.; Thebo, A. Global Wastewater and Sludge Production, Treatment and Use. In Wastewater: Economic Asset in An Urbanizing World; Springer: Berlin, Germany, 2015; ISBN 9789401795456. [Google Scholar]
- Świerczek, L.; Cieślik, B.M.; Konieczka, P. The Potential of Raw Sewage Sludge in Construction Industry—A Review. J. Clean. Prod. 2018, 200, 342–356. [Google Scholar] [CrossRef]
- Nazari, L.; Sarathy, S.; Santoro, D.; Ho, D.; Ray, M.B.; Xu, C.C. 3—Recent Advances in Energy Recovery from Wastewater Sludge. In Direct Thermochemical Liquefaction for Energy Applications; Woodhead Publishing: Thorston, UK, 2018; ISBN 9780081010259. [Google Scholar]
- Singh, V.; Phuleria, H.C.; Chandel, M.K. Estimation of Energy Recovery Potential of Sewage Sludge in India: Waste to Watt Approach. J. Clean. Prod. 2020, 276, 122538. [Google Scholar] [CrossRef]
- Yan, P.; Qin, R.C.; Guo, J.S.; Yu, Q.; Li, Z.; Chen, Y.P.; Shen, Y.; Fang, F. Net-Zero-Energy Model for Sustainable Wastewater Treatment. Environ. Sci. Technol. 2017, 51, 1017–1023. [Google Scholar] [CrossRef]
- Haron, R.; Mat, R.; Tuan Abdullah, T.A.; Rahman, R.A. Overview on Utilization of Biodiesel By-Product for Biohydrogen Production. J. Clean. Prod. 2018, 172, 314–324. [Google Scholar] [CrossRef]
- Young, M.N.; Marcus, A.K.; Rittmann, B.E. A Combined Activated Sludge Anaerobic Digestion Model (CASADM) to Understand the Role of Anaerobic Sludge Recycling in Wastewater Treatment Plant Performance. Bioresour. Technol. 2013, 136, 196–204. [Google Scholar] [CrossRef] [Green Version]
- Campo, G.; Cerutti, A.; Zanetti, M.; Scibilia, G.; Lorenzi, E.; Ruffino, B. Enhancement of Waste Activated Sludge (WAS) Anaerobic Digestion by Means of Pre- and Intermediate Treatments. Technical and Economic Analysis at a Full-Scale WWTP. J. Environ. Manag. 2018, 216, 372–382. [Google Scholar] [CrossRef]
- Hanum, F.; Yuan, L.C.; Kamahara, H.; Aziz, H.A.; Atsuta, Y.; Yamada, T.; Daimon, H. Treatment of Sewage Sludge Using Anaerobic Digestion in Malaysia: Current State and Challenges. Front. Energy Res. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Panepinto, D.; Riggio, V.A.; Campo, G.; Cerutti, A.; Comoglio, C.; Zanetti, M.C. Analysis of Two Treatment Technologies for Coffee Roasting Matrixes: Combustion and Anaerobic Digestion. Clean Technol. Environ. Policy 2019, 21, 685–694. [Google Scholar] [CrossRef]
- Pramanik, S.K.; Suja, F.B.; Zain, S.M.; Pramanik, B.K. The Anaerobic Digestion Process of Biogas Production from Food Waste: Prospects and Constraints. Bioresour. Technol. Rep. 2019, 8, 100310. [Google Scholar] [CrossRef]
- Silva, F.M.S.; Mahler, C.F.; Oliveira, L.B.; Bassin, J.P. Hydrogen and Methane Production in a Two-Stage Anaerobic Digestion System by Co-Digestion of Food Waste, Sewage Sludge and Glycerol. Waste Manag. 2018, 76, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Maragkaki, A.E.; Fountoulakis, M.; Kyriakou, A.; Lasaridi, K.; Manios, T. Boosting Biogas Production from Sewage Sludge by Adding Small Amount of Agro-Industrial by-Products and Food Waste Residues. Waste Manag. 2018, 71, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.; Chattopadhyay, P.; Shome, A.; Banerjee, S.N.; Chakraborty, A.K.; Mathew, A.K.; Chaudhury, S. An Overview of Physico-Chemical Mechanisms of Biogas Production by Microbial Communities: A Step towards Sustainable Waste Management. 3 Biotech 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirmasoumi, S.; Ebrahimi, S.; Saray, R.K. Enhancement of Biogas Production from Sewage Sludge in a Wastewater Treatment Plant: Evaluation of Pretreatment Techniques and Co-Digestion under Mesophilic and Thermophilic Conditions. Energy 2018, 157, 707–717. [Google Scholar] [CrossRef]
- Kasinski, S. Mesophilic and Thermophilic Anaerobic Digestion of Organic Fraction Separated during Mechanical Heat Treatment of Municipal Waste. Appl. Sci. 2020, 10, 2412. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, K.; Azuma, Y.; Kosaka, T.; Yakushi, T.; Hoshida, H.; Akada, R.; Yamada, M. Genomic Analyses of Thermotolerant Microorganisms Used for High-Temperature Fermentations. Biosci. Biotechnol. Biochem. 2016, 80, 655–668. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Ge, X.; Zhang, W.; Wang, L.; Yu, Z.; Li, Y. Methanol Production from Biogas with a Thermotolerant Methanotrophic Consortium Isolated from an Anaerobic Digestion System. Energy Fuels 2017, 31, 2970–2975. [Google Scholar] [CrossRef]
- Moonmangmee, D.; Adachi, O.; Ano, Y.; Shinagawa, E.; Toyama, H.; Theeragool, G.; Lotong, N.; Matsushita, K. Isolation and Characterization of Thermotolerant Gluconobacter Strains Catalyzing Oxidative Fermentation at Higher Temperatures. Biosci. Biotechnol. Biochem. 2000, 64, 2306–2315. [Google Scholar] [CrossRef] [Green Version]
- Limtong, S.; Sringiew, C.; Yongmanitchai, W. Production of Fuel Ethanol at High Temperature from Sugar Cane Juice by a Newly Isolated Kluyveromyces Marxianus. Bioresour. Technol. 2007, 98, 3367–3374. [Google Scholar] [CrossRef]
- Manaia, C.M.; Moore, E.R.B. Pseudomonas Thermotolerans Sp. Nov., a Thermotolerant Species of the Genus Pseudomonas Sensu Stricto. Int. J. Syst. Evol. Microbiol. 2002, 52, 2203–2209. [Google Scholar] [CrossRef] [PubMed]
- Ndoye, B.; Lebecque, S.; Dubois-Dauphin, R.; Tounkara, L.; Guiro, A.T.; Kere, C.; Diawara, B.; Thonart, P. Thermoresistant Properties of Acetic Acids Bacteria Isolated from Tropical Products of Sub-Saharan Africa and Destined to Industrial Vinegar. Enzyme Microb. Technol. 2006, 39, 916–923. [Google Scholar] [CrossRef]
- Suksong, W.; Kongjan, P.; Prasertsan, P.; O-Thong, S. Thermotolerant Cellulolytic Clostridiaceae and Lachnospiraceae Rich Consortium Enhanced Biogas Production from Oil Palm Empty Fruit Bunches by Solid-State Anaerobic Digestion. Bioresour. Technol. 2019, 291, 121851. [Google Scholar] [CrossRef] [PubMed]
- Łukajtis, R.; Hołowacz, I.; Kucharska, K.; Glinka, M.; Rybarczyk, P.; Przyjazny, A.; Kamiński, M. Hydrogen Production from Biomass Using Dark Fermentation. Renew. Sustain. Energy Rev. 2018, 91, 665–694. [Google Scholar] [CrossRef]
- Scherhag, P.; Ackermann, J.U. Removal of Sugars in Wastewater from Food Production through Heterotrophic Growth of Galdieria Sulphuraria. Eng. Life Sci. 2021, 21, 233–241. [Google Scholar] [CrossRef]
- Yoon, Y.M.; Kim, S.H.; Shin, K.S.; Kim, C.H. Effects of Substrate to Inoculum Ratio on the Biochemical Methane Potential of Piggery Slaughterhouse Wastes. Asian-Australas. J. Anim. Sci. 2014, 27, 600–607. [Google Scholar] [CrossRef]
- Filer, J.; Ding, H.H.; Chang, S. Biochemical Methane Potential (BMP) Assay Method for Anaerobic Digestion Research. Water 2019, 11, 921. [Google Scholar] [CrossRef] [Green Version]
- Raposo, F.; De La Rubia, M.A.; Fernández-Cegrí, V.; Borja, R. Anaerobic Digestion of Solid Organic Substrates in Batch Mode: An Overview Relating to Methane Yields and Experimental Procedures. Renew. Sustain. Energy Rev. 2012, 16, 861–877. [Google Scholar] [CrossRef]
- Raposo, F.; Banks, C.J.; Siegert, I.; Heaven, S.; Borja, R. Influence of Inoculum to Substrate Ratio on the Biochemical Methane Potential of Maize in Batch Tests. Process Biochem. 2006, 41, 1444–1450. [Google Scholar] [CrossRef]
- Liu, Y. Bioenergetic Interpretation on the S0/X0 Ratio in Substrate-Sufficient Batch Culture. Water Res. 1996, 30, 2766–2770. [Google Scholar] [CrossRef]
- Wang, B. Factors That Influence the Biochemical Methane Potential (BMP) Test: Steps towards the Standardisation of BMP Test; Lund University: Lund, Sweden, 2016; ISBN 9789174224368. [Google Scholar]
- Ma, X.; Jiang, T.; Chang, J.; Tang, Q.; Luo, T.; Cui, Z. Effect of Substrate to Inoculum Ratio on Biogas Production and Microbial Community during Hemi-Solid-State Batch Anaerobic Co-Digestion of Rape Straw and Dairy Manure. Appl. Biochem. Biotechnol. 2019, 189, 884–902. [Google Scholar] [CrossRef] [PubMed]
- Dixon, P.J.; Ergas, S.J.; Mihelcic, J.R.; Hobbs, S.R. Effect of Substrate to Inoculum Ratio on Bioenergy Recovery from Food Waste, Yard Waste, and Biosolids by High Solids Anaerobic Digestion. Environ. Eng. Sci. 2019, 36, 1459–1465. [Google Scholar] [CrossRef] [Green Version]
- Bolzonella, D.; Pavan, P.; Battistoni, P.; Cecchi, F. Mesophilic Anaerobic Digestion of Waste Activated Sludge: Influence of the Solid Retention Time in the Wastewater Treatment Process. Process Biochem. 2005, 40, 1453–1460. [Google Scholar] [CrossRef]
- Ramdial, P.K.; Bastian, B.C.; Goodlad, J.; McGrath, J.A.; Lazar, A. Specialized Techniques in Dermatopathology. In McKee’s Pathology of the Skin; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Tucker, C.M.; Cadotte, M.W.; Carvalho, S.B.; Jonathan Davies, T.; Ferrier, S.; Fritz, S.A.; Grenyer, R.; Helmus, M.R.; Jin, L.S.; Mooers, A.O.; et al. A Guide to Phylogenetic Metrics for Conservation, Community Ecology and Macroecology. Biol. Rev. 2017, 92, 698–715. [Google Scholar] [CrossRef]
- Wei, C.H.; Harb, M.; Amy, G.; Hong, P.Y.; Leiknes, T.O. Sustainable Organic Loading Rate and Energy Recovery Potential of Mesophilic Anaerobic Membrane Bioreactor for Municipal Wastewater Treatment. Bioresour. Technol. 2014, 166, 326–334. [Google Scholar] [CrossRef]
- Tetteh, E.K.; Rathilal, S. Biophotocatalytic Reduction of CO2 in Anaerobic Biogas Produced from Wastewater Treatment Using an Integrated System. Catalysts 2022, 12, 76. [Google Scholar] [CrossRef]
- El-Gendy, N.S.; Nassar, H.N. Biosynthesized Magnetite Nanoparticles as an Environmental Opulence and Sustainable Wastewater Treatment. Sci. Total Environ. 2021, 774, 145610. [Google Scholar] [CrossRef]
- González, J.; Sánchez, M.; Gómez, X. Enhancing Anaerobic Digestion: The Effect of Carbon Conductive Materials. C 2018, 4, 59. [Google Scholar] [CrossRef] [Green Version]
- Kouzi, A.I.; Puranen, M.; Kontro, M.H. Evaluation of the Factors Limiting Biogas Production in Full-Scale Processes and Increasing the Biogas Production Efficiency. Environ. Sci. Pollut. Res. 2020, 27, 28155–28168. [Google Scholar] [CrossRef]
- Figeac, N.; Trably, E.; Bernet, N.; Delgenès, J.P.; Escudié, R. Temperature and Inoculum Origin Influence the Performance of Ex-Situ Biological Hydrogen Methanation. Molecules 2020, 25, 5665. [Google Scholar] [CrossRef]
- Zabranska, J.; Pokorna, D. Bioconversion of Carbon Dioxide to Methane Using Hydrogen and Hydrogenotrophic Methanogens. Biotechnol. Adv. 2018, 36, 707–720. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Xu, J.; Liu, Z.; Huang, Z.; Zhao, M.; Shi, W.; Ruan, W. Optimal the Ex-Situ Biogas Biological Upgrading to Biomethane and Its Combined Application with the Anaerobic Digestion Stage. Energy Sources Part A Recover. Util. Environ. Eff. 2021, 43, 2147–2159. [Google Scholar] [CrossRef]
- Guneratnam, A.J.; Ahern, E.; FitzGerald, J.A.; Jackson, S.A.; Xia, A.; Dobson, A.D.W.; Murphy, J.D. Study of the Performance of a Thermophilic Biological Methanation System. Bioresour. Technol. 2017, 225, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.R.; Fornero, J.J.; Stark, R.; Mets, L.; Angenent, L.T. A Single-Culture Bioprocess of Methanothermobacter Thermautotrophicus to Upgrade Digester Biogas by CO2-to-CH4 Conversion with H2. Archaea 2013, 2013, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bassani, I.; Kougias, P.G.; Treu, L.; Porté, H.; Campanaro, S.; Angelidaki, I. Optimization of Hydrogen Dispersion in Thermophilic Up-Flow Reactors for Ex Situ Biogas Upgrading. Bioresour. Technol. 2017, 234, 310–319. [Google Scholar] [CrossRef]
- Porté, H.; Kougias, P.G.; Alfaro, N.; Treu, L.; Campanaro, S.; Angelidaki, I. Process Performance and Microbial Community Structure in Thermophilic Trickling Biofilter Reactors for Biogas Upgrading. Sci. Total Environ. 2019, 655, 529–538. [Google Scholar] [CrossRef]
- Westerholm, M.; Isaksson, S.; Karlsson Lindsjö, O.; Schnürer, A. Microbial Community Adaptability to Altered Temperature Conditions Determines the Potential for Process Optimisation in Biogas Production. Appl. Energy 2018, 226, 838–848. [Google Scholar] [CrossRef]
- Wang, S.; Ma, F.; Ma, W.; Wang, P.; Zhao, G.; Lu, X. Influence of Temperature on Biogas Production Efficiency and Microbial Community in a Two-Phase Anaerobic Digestion System. Water 2019, 11, 133. [Google Scholar] [CrossRef] [Green Version]
- Hao, L.; Michaelsen, T.Y.; Singleton, C.M.; Dottorini, G.; Kirkegaard, R.H.; Albertsen, M.; Nielsen, P.H.; Dueholm, M.S. Novel Syntrophic Bacteria in Full-Scale Anaerobic Digesters Revealed by Genome-Centric Metatranscriptomics. ISME J. 2020, 14, 906–918. [Google Scholar] [CrossRef]
- Alves, J.I.; van Gelder, A.H.; Alves, M.M.; Sousa, D.Z.; Plugge, C.M. Moorella Stamsii Sp. Nov., a New Anaerobic Thermophilic Hydrogenogenic Carboxydotroph Isolated from Digester Sludge. Int. J. Syst. Evol. Microbiol. 2013, 63, 4072–4076. [Google Scholar] [CrossRef] [Green Version]
- McCartney, D.M.; Oleszkiewicz, J.A. Competition between Methanogens and Sulfate Reducers: Effect of COD:Sulfate Ratio and Acclimation. Water Environ. Res. 1993, 65, 655–664. [Google Scholar] [CrossRef]
- Conrad, R. Contribution of Hydrogen to Methane Production and Control of Hydrogen Concentrations in Methanogenic Soils and Sediments. FEMS Microbiol. Ecol. 2006, 28, 193–202. [Google Scholar] [CrossRef]
- Habtewold, J.; Gordon, R.; Sokolov, V.; VanderZaag, A.; Wagner-Riddle, C.; Dunfield, K. Reduction in Methane Emissions from Acidified Dairy Slurry Is Related to Inhibition of Methanosarcina Species. Front. Microbiol. 2018, 9, 2806. [Google Scholar] [CrossRef] [PubMed]
- Demirel, B.; Scherer, P. The Roles of Acetotrophic and Hydrogenotrophic Methanogens during Anaerobic Conversion of Biomass to Methane: A Review. Rev. Environ. Sci. Biotechnol. 2008, 7, 173–190. [Google Scholar] [CrossRef]
- Ma, T.T.; Liu, L.Y.; Rui, J.P.; Yuan, Q.; Feng, D.S.; Zhou, Z.; Dai, L.R.; Zeng, W.Q.; Zhang, H.; Cheng, L. Coexistence and Competition of Sulfate-Reducing and Methanogenic Populations in an Anaerobic Hexadecane-Degrading Culture. Biotechnol. Biofuels 2017, 10, 207. [Google Scholar] [CrossRef] [Green Version]
- Mei, R.; Nobu, M.K.; Narihiro, T.; Kuroda, K.; Muñoz Sierra, J.; Wu, Z.; Ye, L.; Lee, P.K.H.; Lee, P.H.; van Lier, J.B.; et al. Operation-Driven Heterogeneity and Overlooked Feed-Associated Populations in Global Anaerobic Digester Microbiome. Water Res. 2017, 124, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Wang, C.; Sun, F.; Zhu, W.; Wu, W. A Comparison of Microbial Characteristics between the Thermophilic and Mesophilic Anaerobic Digesters Exposed to Elevated Food Waste Loadings. Bioresour. Technol. 2014, 152, 420–428. [Google Scholar] [CrossRef]
- Odirile, P.T.; Marumoloa, P.M.; Manali, A.; Gikas, P. Anaerobic Digestion for Biogas Production from Municipal Sewage Sludge: A Comparative Study between Fine Mesh Sieved Primary Sludge and Sedimented Primary Sludge. Water 2021, 13, 3532. [Google Scholar] [CrossRef]
- Shu, C.H.; Jaiswal, R.; Kuo, M.-d.; Yu, B.H. Enhancing Methane Production in a Two-Stage Anaerobic Digestion of Spent Mushroom Substrate and Chicken Manure via Activation of Sludge, Optimization of Temperature, and C/N Ratio. Front. Environ. Sci. 2022, 9, 1–11. [Google Scholar] [CrossRef]
- Beale, D.J.; Karpe, A.V.; McLeod, J.D.; Gondalia, S.V.; Muster, T.H.; Othman, M.Z.; Palombo, E.A.; Joshi, D. An “omics” Approach towards the Characterisation of Laboratory Scale Anaerobic Digesters Treating Municipal Sewage Sludge. Water Res. 2016, 88, 346–357. [Google Scholar] [CrossRef]
- Moestedt, J.; Nilsson Påledal, S.; Schnürer, A. The Effect of Substrate and Operational Parameters on the Abundance of Sulphate-Reducing Bacteria in Industrial Anaerobic Biogas Digesters. Bioresour. Technol. 2013, 132, 327–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziembińska-Buczyńska, A.; Banach, A.; Bacza, T.; Pieczykolan, M. Diversity and Variability of Methanogens during the Shift from Mesophilic to Thermohilic Conditions While Biogas Production. World J. Microbiol. Biotechnol. 2014, 30, 3047–3053. [Google Scholar] [CrossRef] [PubMed]
- Ahring, B.K. Methanogenesis in Thermophilic Biogas Reactors. Antonie Van Leeuwenhoek 1995, 67, 91–102. [Google Scholar] [CrossRef]
- Boušková, A.; Dohányos, M.; Schmidt, J.E.; Angelidaki, I. Strategies for Changing Temperature from Mesophilic to Thermophilic Conditions in Anaerobic CSTR Reactors Treating Sewage Sludge. Water Res. 2005, 39, 1481–1488. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, W.; Eddy, C. Wastewater Engineering Treatment and Reuse; McGraw Hill: New York, NY, USA, 2003. [Google Scholar]
- Al-mashhadani, M.K.H.; Wilkinson, S.J.; Zimmerman, W.B. Carbon Dioxide Rich Microbubble Acceleration of Biogas Production in Anaerobic Digestion. Chem. Eng. Sci. 2016, 156, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; He, P.; Lin, Y.; Shao, L.; Lü, F. A Methanogenic Consortium Was Active and Exhibited Long-Term Survival in an Extremely Acidified Thermophilic Bioreactor. Front. Microbiol. 2019, 10, 2757. [Google Scholar] [CrossRef]
- Hao, L.P.; Lü, F.; Li, L.; Shao, L.M.; He, P.J. Shift of Pathways during Initiation of Thermophilic Methanogenesis at Different Initial PH. Bioresour. Technol. 2012, 126, 418–424. [Google Scholar] [CrossRef]
- Tezel, U.; Tandukar, M.; Hajaya, M.G.; Pavlostathis, S.G. Transition of Municipal Sludge Anaerobic Digestion from Mesophilic to Thermophilic and Long-Term Performance Evaluation. Bioresour. Technol. 2014, 170, 385–394. [Google Scholar] [CrossRef]
- Tian, Z.; Zhang, Y.; Li, Y.; Chi, Y.; Yang, M. Rapid Establishment of Thermophilic Anaerobic Microbial Community during the One-Step Startup of Thermophilic Anaerobic Digestion from a Mesophilic Digester. Water Res. 2015, 69, 9–19. [Google Scholar] [CrossRef]
- Kim, J.; Lee, C. Response of a Continuous Anaerobic Digester to Temperature Transitions: A Critical Range for Restructuring the Microbial Community Structure and Function. Water Res. 2016, 89, 241–251. [Google Scholar] [CrossRef]
- van den Brand, T.P.H.; Roest, K.; Brdjanovic, D.; Chen, G.H.; van Loosdrecht, M.C.M. Influence of Acetate and Propionate on Sulphate-Reducing Bacteria Activity. J. Appl. Microbiol. 2014, 117, 1839–1847. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Anaerobic Sludge | Units |
|---|---|---|
| pH | 7.09 | pH = −log10[a(H+)] |
| Total Solid (TS) | 8000 | mg/L |
| Volatile Solid (VS) | 3000 | mg/L |
| Fixed Solid (FS) | 5000 | mg/L |
| VS/TS ratio | 0.37 | - |
| Temperature (°C) | pH | Total Suspended Solids (TSS) in mg·L−1 | Volatile Suspended Solids (VSS) in mg·L−1 | VSS.TSS−1 | Chemical Oxygen Demand (COD) in mg·L−1 |
|---|---|---|---|---|---|
| 42 °C | 7.64 | 7300 | 4675 | 0.64 | 498.72 |
| 45 °C | 8.20 | 6860 | 4105 | 0.60 | 1911.76 |
| 48 °C | 8.33 | 6880 | 3465 | 0.50 | 3690.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sudiartha, G.A.W.; Imai, T.; Hung, Y.-T. Effects of Stepwise Temperature Shifts in Anaerobic Digestion for Treating Municipal Wastewater Sludge: A Genomic Study. Int. J. Environ. Res. Public Health 2022, 19, 5728. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19095728
Sudiartha GAW, Imai T, Hung Y-T. Effects of Stepwise Temperature Shifts in Anaerobic Digestion for Treating Municipal Wastewater Sludge: A Genomic Study. International Journal of Environmental Research and Public Health. 2022; 19(9):5728. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19095728
Chicago/Turabian StyleSudiartha, Gede Adi Wiguna, Tsuyoshi Imai, and Yung-Tse Hung. 2022. "Effects of Stepwise Temperature Shifts in Anaerobic Digestion for Treating Municipal Wastewater Sludge: A Genomic Study" International Journal of Environmental Research and Public Health 19, no. 9: 5728. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19095728