
CRISPR-Associated (CAS) Effectors Delivery via Microfluidic Cell-Deformation Chip


Abstract
:1. Introduction
2. Limitations of Previous Automated Delivery Technologies
3. Microfluidic Devices for Gene Delivery Systems
4. CRISPR/Cas9 Systems and Designing the sgRNA
5. The other CRISPR/Cas Approaches for Diagnosis
6. Methods of Delivery CRISPR/Cas System Viral Transduction
Non-Viral Delivery
7. Microfluidic Methods and Clinical Applications

{kind=link}
{kind=link}
| Target | Transfection Method | Experiments | Results | Ref. |
|---|---|---|---|---|
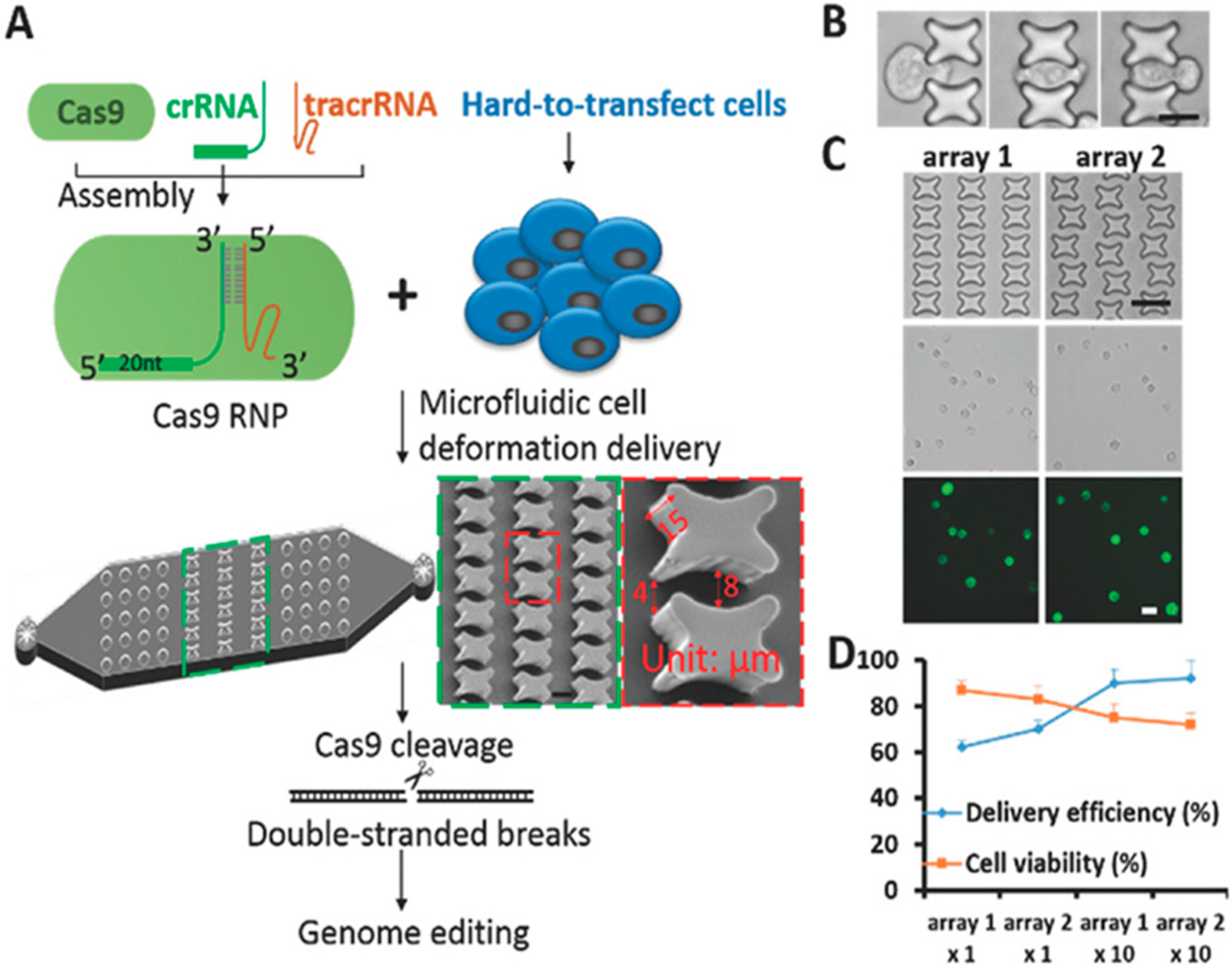
| Cas9/RNPs | Microfluidic Rapid mechanical deformation to shear cells for delivery | The fabrication of the delivery chip and enhanced the transition efficiency of Cas9 RNPs into human primary CD4+ T cells | Knock-in genome amendment in primary T cells PD-1 as an engineering T cells for cancer and tumor immunotherapies | Han, X. et al., 2017 [95] |
| CRISPR/Cas9 | Microfluidic Rapid mechanical deformation of cells to shear cells | The combination of CRISPR-Cas9 screen with chip-based cell deformation sorting highly deformable cells in a high-efficiency method | Identify tumor-inhibiting kinases and loss of CRISPR-Cas9-mediated kinase function for cancer cell deformation and invasive potential | Han, X. et al., 2016 [97] |
| CRISPR/Cas9 | Microfluidic Rapid mechanical deformation of cells to shear cells | Optimizing the physical constriction in a microfluidic setup for delivering ssDNA, siRNAs, and large-sized plasmids into different cell types, adherent and non-adherent cells | Approaching high delivery efficiency of macromolecules into hard-to-transfect lymphoma cells and embryonic stem cells, with maintaining high cell viability | Han, X. et al., 2015 [52] |
| CRISPR/Cas9 | Microfluidic A new droplet-based method along with chemical delivery through lipid vesicles | Digital microfluidics (DMF) for targeting to edit automated the MAPK/ERK pathway, especially RAF1 gene with and without a Raf-1 inhibitor | Decoding the genes related to lung cancer in terms of transfer and deletion efficiency with a standard imaging pipeline | Sinha, H. et al., 2018 [74] |
| CRISPR/Cas9 | Microfluidic A high-throughput electroporation microsystem (electrical force) | Two plasmids encoding green and red fluorescent proteins electroporated into HeLa cells on a 169-microwell array chip | Conducting sgRNA into 293T cells that are expressing the Cas9 nuclease for increasing efficient transfection | Bian, S. et al., 2017 [104] |
| CRISPR/Cas9 | Electroporation treatment & mechanical membrane disruption-based microfluidic | Comparing electroporation with microfluidic membrane deformation technique or “cell squeezing” with in two cell types: HSC & T cells to target PD-1 | Squeezing had minimal aberrant transcriptional responses and T cells edited had a tumor-killing function | Ditommaso, T. et al., 2018 [108] |
| CRISPR/Cas9 | Microfluidic integrated with chemical delivery and inducible CRISPR-Cas construct for detection yeast | Evaluation DNA damage or genomic instability behave in induction with genotoxic chemicals (MMS, HU) or a single double-strand break using induced CRISPR-Cas9 expression | A device for long-term imaging of yeast cells under stable or varied media conditions Permitting acquisition of high-resolution images from cells fixing in a two dimensional imaging plane | Schmidt, G.W. et al., 2017 [119] |
| CRISPR/Cas9 | A centrifugal microfluidic platform with fluorescence-based sedimentation nuclease assay for detection | Multiple Cas9 species and a dCas9 (dead Cas9) integrated into effector domains for detecting of both Cas9 protein levels and function of catalytic nuclease | Improving a manual microfluidic tool for the detection of fluorescence signals and tracing of Cas9 multiple species directly in the bodily fluids | Phaneuf, C. et al., 2019 [25] |
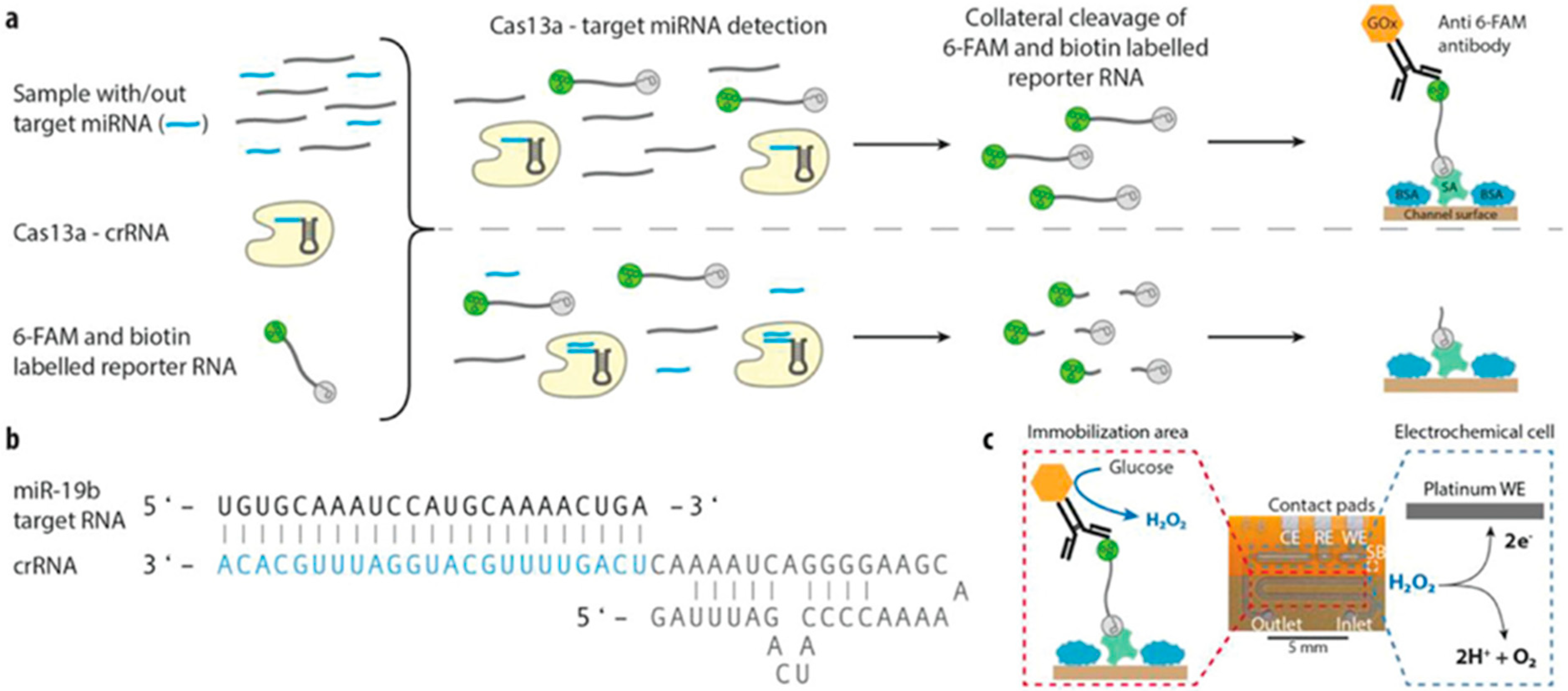
| CRISPR/Cas13a | Microfluidic integrated electrochemical biosensor for the on-site detection | Combining the CRISPR technology with a microfluidic equipped with the DFR technology (dry film photo-resist) to evaluate miRNA levels | Permitting the detection of the target miRNAs, including miR-19b and miR-20a as brain tumor markers with high sensitivity method | Brush, R. et al., 2019 [36] |
| CRISPR/Cas13a | Microfluidic integrated a photocleavable capture probe and a sequence-specific barcode fluorescence reporter | Mixing automated Microfluidic, hybridization and measuring nonspecific cleavage products of Cas13a by a custom integrated fluorometer in small size and in-field diagnosis | Applying an automated point of care system for Ebola RNA detection with RNA-guided RNA endonuclease Cas13a, Achieving within 5 min, a detection limit of 20 pfu/mL of purified Ebola RNA | Qin, P. et al., 2019 [115] |
| CRISPR/Cas12 | Microfluidic An integrated micropillar polydimethylsiloxane Microinjection for viral detection | After modification surface, the reporter probes were firstly immobilized in the enclosed channel, and the CRISPR Cas12a/crRNA complex was injected into the fully enclosed system for non-specifically cleaves the ssDNA reporters | Using the high activity of the CRISPR-Cas12a enzyme and the ability of micropillars (IMPACT chip) to bind more reporter probe or pathogens and the next-generation molecular diagnostic platform for point-of-care applications | Hass, K.N. et al., 2020 [120] |
| CRISPR/Cas12 | Microfluidic integrated electric field gradient using a selective ionic focusing detection (isotachophoresis) | Combining CRISPR-Cas12 system with an electro-kinetic microfluidic technique and loop-mediated isothermal amplification to evaluate viruses | Diagnosing SARS-CoV-2 in 30 min that provide automated purification of target RNA from a nasopharyngeal swab sample | Ramachandran, A. et al., 2020 [118] |
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Han, H.A.; Pang, J.K.S.; Soh, B.-S. Mitigating off-target effects in CRISPR/Cas9-mediated in vivo gene editing. J. Mol. Med. 2020, 98, 615–632. [Google Scholar] [CrossRef] [Green Version]
- Blaese, R.M.; Culver, K.W.; Miller, A.D.; Carter, C.S.; Fleisher, T.; Clerici, M.; Shearer, G.; Chang, L.; Chiang, Y.; Tolstoshev, P.; et al. T Lymphocyte-Directed Gene Therapy for ADA- SCID: Initial Trial Results After 4 Years. Science 1995, 270, 475–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, B.X.; Loh, S.J.H.; Chan, W.K.; Soh, B.S. In Vivo Genome Editing as a Therapeutic Approach. Int. J. Mol. Sci. 2018, 19, 2721. [Google Scholar] [CrossRef] [Green Version]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horii, T.; Arai, Y.; Yamazaki, M.; Morita, S.; Kimura, M.; Itoh, M.; Abe, Y.; Hatada, I. Validation of microinjection methods for generating knockout mice by CRISPR/Cas-mediated genome engineering. Sci. Rep. 2014, 4, 4513. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Yan, C.; Pan, X.; Mahfouz, M.; Wang, J.; Zhu, J.-K.; Shi, Y.; Yan, N. Structural Basis for Sequence-Specific Recognition of DNA by TAL Effectors. Science 2012, 335, 720–723. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Xu, J.; Ge, S.; Lai, L. CRISPR/Cas: Advances, Limitations, and Applications for Precision Cancer Research. Front. Med. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Cyranoski, D. Chinese scientists to pioneer first human CRISPR trial. Nat. Cell Biol. 2016, 535, 476–477. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Glass, Z.; Huang, M.; Chen, Z.-Y.; Xu, Q. Ex vivo cell-based CRISPR/Cas9 genome editing for therapeutic applications. Biomaterials 2020, 234, 119711. [Google Scholar] [CrossRef]
- Li, L.; He, Z.-Y.; Wei, X.-W.; Gao, G.-P.; Wei, Y.-Q. Challenges in CRISPR/CAS9 delivery: Potential roles of non-viral vectors. Hum. Gene Ther. 2015, 26, 452–462. [Google Scholar] [CrossRef]
- Elvira, K.S. Microfluidic technologies for drug discovery and development: Friend or foe? Trends Pharmacol. Sci. 2021. [Google Scholar] [CrossRef]
- Chiu, D.T.; Demello, A.J.; Di Carlo, D.; Doyle, P.S.; Hansen, C.; Maceiczyk, R.M.; Wootton, R.C. Small but Perfectly Formed? Successes, Challenges, and Opportunities for Microfluidics in the Chemical and Biological Sciences. Chem 2017, 2, 201–223. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, S.J.; Issadore, D.; Mitchell, M.J. Microfluidic formulation of nanoparticles for biomedical applications. Biomaterials 2021, 274, 120826. [Google Scholar] [CrossRef]
- Yixiao, L.; Jianzhang, P.A.N.; Qun, F. Research advances of high-throughput cell-based drug screening systems based on microfluidic technique. Chin. J. Chromatogr. 2021, 39, 567. [Google Scholar]
- Levy, S.L.; Craighead, H.G. DNA manipulation, sorting, and mapping in nanofluidic systems. Chem. Soc. Rev. 2010, 39, 1133–1152. [Google Scholar] [CrossRef]
- Ottino, J.M.; Wiggins, S. Introduction: Mixing in microfluidics. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2004, 362, 923–935. [Google Scholar] [CrossRef]
- Balbino, T.A.; Azzoni, A.R.; De La Torre, L.G. Microfluidic devices for continuous production of pDNA/cationic liposome complexes for gene delivery and vaccine therapy. Colloids Surf. B Biointerfaces 2013, 111, 203–210. [Google Scholar] [CrossRef]
- Laohakunakorn, N.; Lavickova, B.; Swank, Z.; Laurent, J.; Maerkl, S.J. Steady-state cell-free gene expression with microfluidic chemostats. In Synthetic Gene Circuits; Springer: Berlin/Heidelberg, Germany, 2021; pp. 189–203. [Google Scholar]
- Ahmadi, S.; Rabiee, N.; Bagherzadeh, M.; Karimi, M. Microfluidic devices for gene delivery systems. Biomed. Appl. Microfluid. Devices 2021, 187–208. [Google Scholar] [CrossRef]
- Kim, J.; Hwang, I.; Britain, D.; Chung, T.D.; Sun, Y.; Kim, D.-H. Microfluidic approaches for gene delivery and gene therapy. Lab. Chip 2011, 11, 3941–3948. [Google Scholar] [CrossRef]
- Niculescu, A.-G.; Chircov, C.; Bîrcă, A.; Grumezescu, A. Fabrication and Applications of Microfluidic Devices: A Review. Int. J. Mol. Sci. 2021, 22, 2011. [Google Scholar] [CrossRef]
- Rautela, I.; Uniyal, P.; Thapliyal, P.; Chauhan, N.; Sinha, V.B.; Sharma, M.D. An extensive review to facilitate under-standing of CRISPR technology as a gene editing possibility for enhanced therapeutic applications. Gene 2021, 785, 145615. [Google Scholar] [CrossRef]
- Zhang, S.; Shen, J.; Li, D.; Cheng, Y. Strategies in the delivery of Cas9 ribonucleoprotein for CRISPR/Cas9 genome editing. Theranostics 2021, 11, 614–648. [Google Scholar] [CrossRef]
- Leibowitz, M.L.; Papathanasiou, S.; Doerfler, P.A.; Blaine, L.J.; Sun, L.; Yao, Y.; Zhang, C.-Z.; Weiss, M.J.; Pellman, D. Chromothripsis as an on-target consequence of CRISPR–Cas9 genome editing. Nat. Genet. 2021, 1–11. [Google Scholar] [CrossRef]
- Phaneuf, C.R.; Seamon, K.J.; Eckles, T.P.; Sinha, A.; Schoeniger, J.S.; Harmon, B.; Meagher, R.J.; Abhyankar, V.V.; Koh, C.-Y. Ultrasensitive multi-species de-tection of CRISPR-Cas9 by a portable centrifugal microfluidic platform. Anal. Methods 2019, 11, 559–565. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary classification of CRISPR–Cas systems: A burst of class 2 and derived variants. Nat. Rev. Microbiol. 2019, 18, 67–83. [Google Scholar] [CrossRef]
- Yang, H.; Ren, S.; Yu, S.; Pan, H.; Li, T.; Ge, S.; Zhang, J.; Xia, N. Methods Favoring Homology-Directed Repair Choice in Response to CRISPR/Cas9 Induced-Double Strand Breaks. Int. J. Mol. Sci. 2020, 21, 6461. [Google Scholar] [CrossRef]
- Doudna, J.A.; Sontheimer, E.J. The Use of CRISPR/cas9, ZFNs, TALENs in Generating Site-Specific Genome Alterations; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Qasim, W.; Zhan, H.; Samarasinghe, S.; Adams, S.; Amrolia, P.; Stafford, S.; Butler, K.; Rivat, C.; Wright, G.; Somana, K.; et al. Molecular remission of infant B-ALL after infusion of universal TALEN gene-edited CAR T cells. Sci. Transl. Med. 2017, 9, eaaj2013. [Google Scholar] [CrossRef]
- Zhang, B. CRISPR/Cas gene therapy. J. Cell. Physiol. 2021, 236, 2459–2481. [Google Scholar] [CrossRef]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; et al. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef] [Green Version]
- Rusk, N. Spotlight on Cas12. Nat. Chem. Biol. 2019, 16, 215. [Google Scholar] [CrossRef]
- East-Seletsky, A.; O’Connell, M.R.; Knight, S.C.; Burstein, D.; Cate, J.H.D.; Tjian, R.; Doudna, J.A. Two distinct RNase activities of CRISPR-C2c2 enable guide-RNA processing and RNA detection. Nat. Cell Biol. 2016, 538, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Cui, B.; Kiga, K.; Aiba, Y.; Tan, X.-E.; Sato’O, Y.; Kawauchi, M.; Boonsiri, T.; Thitiananpakorn, K.; Taki, Y.; et al. Composition and Diversity of CRISPR-Cas13a Systems in the Genus Leptotrichia. Front. Microbiol. 2019, 10, 2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freije, C.A.; Myhrvold, C.; Boehm, C.K.; Lin, A.E.; Welch, N.L.; Carter, A.; Metsky, H.C.; Luo, C.; Abudayyeh, O.O.; Gootenberg, J.S.; et al. Programmable inhibition and detection of RNA viruses using Cas13. Mol. Cell 2019, 76, 826–837. [Google Scholar] [CrossRef] [Green Version]
- Bruch, R.; Baaske, J.; Chatelle, C.; Meirich, M.; Madlener, S.; Weber, W.; Dincer, C.; Urban, G. CRISPR/Cas13a-powered electro-chemical microfluidic biosensor for nucleic acid amplification-free miRNA diagnostics. Adv Mater. 2019, 31, 1905311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-X.; Li, M.; Lee, C.M.; Chakraborty, S.; Kim, H.-W.; Bao, G.; Leong, K.W. CRISPR/Cas9-based genome editing for disease modeling and therapy: Challenges and opportunities for non-viral delivery. Chem Rev. 2017, 117, 9874–9906. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Keller, B.; Makalou, N.; Sutton, R.E. Systematic Determination of the Packaging Limit of Lentiviral Vectors. Hum. Gene Ther. 2001, 12, 1893–1905. [Google Scholar] [CrossRef]
- Chen, X.; Gonçalves, M.A.F.V. Engineered viruses as genome editing devices. Mol. Ther. 2016, 24, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Esvelt, K.M.; Mali, P.; Braff, J.L.; Moosburner, M.; Yaung, S.J.; Church, G.M. Orthogonal Cas9 proteins for RNA-guided gene regulation and editing. Nat. Methods 2013, 10, 1116–1121. [Google Scholar] [CrossRef] [Green Version]
- Fajrial, A.K.; He, Q.Q.; Wirusanti, N.I.; Slansky, J.E.; Ding, X. A review of emerging physical transfection methods for CRISPR/Cas9-mediated gene editing. Theranostics 2020, 10, 5532–5549. [Google Scholar] [CrossRef]
- Ran, F.A.; Cong, L.; Yan, W.X.; Scott, D.A.; Gootenberg, J.S.; Kriz, A.J.; Zetsche, B.; Shalem, O.; Wu, X.; Makarova, K.S.; et al. In vivo genome editing using Staphylococcus aureus Cas9. Nature 2015, 520, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-Y.; Hsieh, H.-Y.; Pitt, W.G.; Huang, C.-Y.; Tseng, I.-C.; Yeh, C.-K.; Wei, K.-C.; Liu, H.-L. Focused ultrasound-induced blood-brain barrier opening for non-viral, non-invasive, and targeted gene delivery. J. Control. Release 2015, 212, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-D.; Huang, L. Non-viral is superior to viral gene delivery. J. Control. Release 2007, 123, 181–183. [Google Scholar] [CrossRef]
- Kim, S.; Kim, D.; Cho, S.W.; Kim, J.; Kim, J.-S. Highly efficient RNA-guided genome editing in human cells via de-livery of purified Cas9 ribonucleoproteins. Genome Res. 2014, 24, 1012–1019. [Google Scholar] [CrossRef] [Green Version]
- Glass, Z.; Lee, M.; Li, Y.; Xu, Q. Engineering the Delivery System for CRISPR-Based Genome Editing. Trends Biotechnol. 2018, 36, 173–185. [Google Scholar] [CrossRef]
- Miller, J.B.; Zhang, S.; Kos, P.; Xiong, H.; Zhou, K.; Perelman, S.S.; Zhu, H.; Siegwart, D.J. Non-Viral CRISPR/Cas Gene Editing In Vitro and In Vivo Enabled by Synthetic Nanoparticle Co-Delivery of Cas9 mRNA and sgRNA. Angew Chem. Int. Ed. 2017, 56, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.; Norville, J.E.; Church, G.M. RNA-Guided Human Genome Engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuma, T.; Nishikawa, A.; Kume, S.; Chayama, K.; Yamamoto, T. Multiplex genome engineering in human cells using all-in-one CRISPR/Cas9 vector system. Sci. Rep. 2015, 4, 5400. [Google Scholar] [CrossRef] [Green Version]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Liu, Z.; Jo, M.C.; Zhang, K.; Li, Y.; Zeng, Z.; Li, N.; Zu, Y.; Qin, L. CRISPR-Cas9 delivery to hard-to-transfect cells via membrane deformation. Sci. Adv. 2015, 1, e1500454. [Google Scholar] [CrossRef] [Green Version]
- Perez-Pinera, P.; Kocak, D.D.; Vockley, C.M.; Adler, A.F.; Kabadi, A.M.; Polstein, L.R.; Thakore, P.I.; Glass, K.A.; Ousterout, D.G.; Leong, K.W.; et al. RNA-guided gene activa-tion by CRISPR-Cas9–based transcription factors. Nat. Methods 2013, 10, 973–976. [Google Scholar] [CrossRef] [Green Version]
- Mout, R.; Ray, M.; Tonga, G.Y.; Lee, Y.-W.; Tay, T.; Sasaki, K.; Rotello, V.M. Direct Cytosolic Delivery of CRISPR/Cas9-Ribonucleoprotein for Efficient Gene Editing. ACS Nano. 2017, 11, 2452–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chew, W.L.; Tabebordbar, M.; Cheng, J.K.W.; Mali, P.; Wu, E.Y.; Ng, A.H.M.; Zhu, K.; Wagers, A.J.; Church, G.M. A multifunctional AAV–CRISPR–Cas9 and its host response. Nat. Methods 2016, 13, 868–874. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zuris, J.A.; Meng, F.; Rees, H.; Sun, S.; Deng, P.; Han, Y.; Gao, X.; Pouli, D.; Wu, Q.; et al. Efficient delivery of genome-editing proteins using bioreducible lipid nanoparticles. Proc. Natl. Acad. Sci. USA 2016, 113, 2868–2873. [Google Scholar] [CrossRef] [Green Version]
- Zuris, J.A.; Thompson, D.B.; Shu, Y.; Guilinger, J.P.; Bessen, J.L.; Hu, J.H.; Maeder, M.L.; Joung, K.J.; Chen, Z.-Y.; Liu, D.R. Cationic lipid-mediated delivery of proteins enables efficient protein-based genome editing in vitro and in vivo. Nat. Biotechnol. 2015, 33, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, K.J.; Dorkin, J.R.; Yang, J.H.; Heartlein, M.W.; DeRosa, F.; Mir, F.F.; Fenton, O.S.; Anderson, D.G. Optimization of lipid nanoparticle formulations for mRNA delivery in vivo with fractional factorial and definitive screening designs. Nano. Lett. 2015, 15, 7300–7306. [Google Scholar] [CrossRef]
- Ginn, S.L.; Amaya, A.K.; Alexander, I.E.; Edelstein, M.; Abedi, M.R. Gene therapy clinical trials worldwide to 2017: An update. J. Gene Med. 2018, 20, e3015. [Google Scholar] [CrossRef]
- Li, L.; Wang, H.; Ong, Z.Y.; Xu, K.; Ee, P.L.R.; Zheng, S.; Hedrick, J.L.; Yang, Y.-Y. Polymer- and lipid-based nanoparticle therapeutics for the treatment of liver diseases. Nano. Today 2010, 5, 296–312. [Google Scholar] [CrossRef]
- Steichen, S.D.; Caldorera-Moore, M.; Peppas, N.A. A review of current nanoparticle and targeting moieties for the delivery of cancer therapeutics. Eur. J. Pharm. Sci. 2013, 48, 416–427. [Google Scholar] [CrossRef] [Green Version]
- Rahimi, H.; Salehiabar, M.; Charmi, J.; Barsbay, M.; Ghaffarlou, M.; Razlighi, M.R.; Davaran, S.; Khalilov, R.; Sugiyama, M.; Nosrati, H.; et al. Harnessing nanoparticles for the efficient delivery of the CRISPR/Cas9 system. Nano. Today 2020, 34, 100895. [Google Scholar] [CrossRef]
- Yu, X.; Liang, X.; Xie, H.; Kumar, S.; Ravinder, N.; Potter, J.; de Mollerat du Jeu, X.; Chesnut, J.D. Improved delivery of Cas9 protein/gRNA com-plexes using lipofectamine CRISPRMAX. Biotechnol Lett. 2016, 38, 919–929. [Google Scholar] [CrossRef] [Green Version]
- Stewart, M.P.; Langer, R.; Jensen, K.F. Intracellular Delivery by Membrane Disruption: Mechanisms, Strategies, and Concepts. Chem. Rev. 2018, 118, 7409–7531. [Google Scholar] [CrossRef] [PubMed]
- O’Dea, S.; Annibaldi, V.; Gallagher, L.; Mulholland, J.; Molloy, E.L.; Breen, C.J.; Gilbert, D.S.M.; Maguire, M.; Curry, F.-R. Vector-free intracellular delivery by reversible permeabilization. PLoS ONE 2017, 12, e0174779. [Google Scholar] [CrossRef] [Green Version]
- Meacham, J.M.; Durvasula, K.; Degertekin, F.L.; Fedorov, A.G. Physical methods for intracellular delivery: Practical aspects from laboratory use to industrial-scale processing. J. Lab. Autom. 2014, 19, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Chiappini, C.; De Rosa, E.; Martinez, O.J.; Liu, X.; Steele, J.R.; Stevens, M.M.; Tasciotti, E. Biodegradable silicon nanoneedles delivering nucleic acids intracellularly induce localized in vivo neovascularization. Nat. Mater. 2015, 14, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Fajrial, A.K.; Ding, X. Advanced nanostructures for cell membrane poration. Nanotechnology 2019, 30, 264002. [Google Scholar] [CrossRef] [Green Version]
- Stadtmauer, E.A.; Fraietta, J.A.; Davis, M.M.; Cohen, A.D.; Weber, K.L.; Lancaster, E.; Mangan, P.A.; Kulikovskaya, I.; Gupta, M.; Chen, F.; et al. CRISPR-engineered T cells in patients with refractory cancer. Science 2020, 367, 987. [Google Scholar] [CrossRef]
- Hruscha, A.; Schmid, B. Generation of zebrafish models by CRISPR/Cas9 genome editing. In Neuronal Cell Death; Springer: Berlin/Heidelberg, Germany, 2015; pp. 341–350. [Google Scholar]
- Crispo, M.; Mulet, A.P.; Tesson, L.; Barrera, N.; Cuadro, F.; dos Santos-Neto, P.C.; Nguyen, T.H.; Creneguy, A.; Brusselle, L.; Anegon, I.; et al. Efficient generation of myo-statin knockout sheep using CRISPR/Cas9 technology and microinjection into zygotes. PLoS ONE 2015, 10, e0136690. [Google Scholar]
- Song, K.; Li, G.; Zu, X.; Du, Z.; Liu, L.; Hu, Z. The Fabrication and Application Mechanism of Microfluidic Systems for High Throughput Biomedical Screening: A Review. Micromachines 2020, 11, 297. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Chen, Y.; Tang, H.; Zong, N.; Jiang, X. Microfluidics for Biomedical Analysis. Small Methods 2020, 4. [Google Scholar] [CrossRef]
- Sinha, H.; Quach, A.B.V.; Vo, P.Q.N.; Shih, S.C.C. An automated microfluidic gene-editing platform for deciphering cancer genes. Lab. Chip 2018, 18, 2300–2312. [Google Scholar] [CrossRef]
- Kelly, R.T.; Lin, J.M. Cell analysis on microfluidics. Anal. Bioanal. Chem. 2018, 410, 7825–7826. [Google Scholar] [CrossRef] [PubMed]
- Oshchepkov, M.S.; Kovalenko, L.V.; Kotova, Y.O.; Solov’Eva, I.N.; Bystrova, N.A.; Kochetkov, K.A. Potential of Application of Microfluidic Devices in Preparative Chemistry. INEOS OPEN 2020, 3, 92–98. [Google Scholar] [CrossRef]
- Silverio, V.; de Freitas, S.C. Microfabrication techniques for microfluidic devices. In Complex Fluid-Flows in Microfluidics; Springer: Berlin/Heidelberg, Germany, 2018; pp. 25–51. [Google Scholar]
- Han, J.P.; Sun, J.; Wang, L.; Liu, P.; Zhuang, B.; Zhao, L.; Liu, Y.; Li, C.X. The optimization of electrophoresis on a glass micro-fluidic chip and its application in forensic science. J. Forensic Sci. 2017, 62, 1603–1612. [Google Scholar] [CrossRef]
- Agarwal, A.; Goss, J.A.; Cho, A.; McCain, M.L.; Parker, K.K. Microfluidic heart on a chip for higher throughput pharmacological studies. Lab. Chip 2013, 13, 3599–3608. [Google Scholar] [CrossRef] [Green Version]
- Alrifaiy, A.; Lindahl, O.A.; Ramser, K. Polymer-Based Microfluidic Devices for Pharmacy, Biology and Tissue Engineering. Polymers 2012, 4, 1349–1398. [Google Scholar] [CrossRef]
- Liu, Y.; Hu, K.; Wang, Y. Primary Hepatocytes Cultured on a Fiber-Embedded PDMS Chip to Study Drug Metabolism. Polymers 2017, 9, 215. [Google Scholar] [CrossRef] [Green Version]
- Oderinde, O.; Liu, S.; Li, K.; Kang, M.; Imtiaz, H.; Yao, F.; Fu, G. Multifaceted polymeric materials in three-dimensional processing (3DP) technologies: Current progress and prospects. Polym. Adv. Technol. 2018, 29, 1586–1602. [Google Scholar] [CrossRef]
- Lu, X.; Liu, C.; Hu, G.; Xuan, X. Particle manipulations in non-Newtonian microfluidics: A review. J. Colloid Interface Sci. 2017, 500, 182–201. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Guo, J.; Liang, Z.; Fan, D. Artificial Micro/Nanomachines for Bioapplications: Biochemical Delivery and Diagnostic Sensing. Adv. Funct. Mater. 2018, 28. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Y.; Onck, P.; Toonder, J.D. A concise review of microfluidic particle manipulation methods. Microfluid. Nanofluidics 2020, 24, 1–20. [Google Scholar] [CrossRef]
- Chang, H.-N.; Leroueil, P.R.; Selwa, K.; Gasper, C.J.; Tsuchida, R.E.; Wang, J.J.; McHugh, W.M.; Cornell, T.T.; Baker, J.R.; Goonewardena, S.N. Profiling Inflammatory Responses with Microfluidic Immunoblotting. PLoS ONE 2013, 8, e81889. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Qu, F.; Xia, Y.; Jiang, X. Straightforward and Ultrastable Surface Modification of Microfluidic Chips with Norepinephrine Bitartrate Improves Performance in Immunoassays. Anal. Chem. 2018, 90, 3697–3702. [Google Scholar] [CrossRef]
- Ma, J.; Lee, S.M.-Y.; Yi, C.; Li, C.-W. Controllable synthesis of functional nanoparticles by microfluidic platforms for biomedical applications–A review. Lab. Chip 2017, 17, 209–226. [Google Scholar] [CrossRef] [PubMed]
- Ran, R.; Middelberg, A.P.; Zhao, C.-X. Microfluidic synthesis of multifunctional liposomes for tumour targeting. Colloids Surf. B Biointerfaces 2016, 148, 402–410. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Zhang, W.; Zheng, W.; Cao, F.; Jiang, X. Inkjet-printed barcodes for a rapid and multiplexed paper-based assay compatible with mobile devices. Lab. Chip 2017, 17, 3874–3882. [Google Scholar] [CrossRef]
- Hinderer, S.; Layland, S.L.; Schenke-Layland, K. ECM and ECM-like materials—Biomaterials for applications in regenerative medicine and cancer therapy. Adv. Drug Deliv. Rev. 2016, 97, 260–269. [Google Scholar] [CrossRef]
- Domachuk, P.; Tsioris, K.; Omenetto, F.G.; Kaplan, D.L. Bio-microfluidics: Biomaterials and Biomimetic Designs. Adv. Mater. 2009, 22, 249–260. [Google Scholar] [CrossRef]
- Song, K.; Wang, Z.; Liu, R.; Chen, G.; Liu, L. Microfabrication-Based Three-Dimensional (3-D) Extracellular Matrix Microenvironments for Cancer and Other Diseases. Int. J. Mol. Sci. 2018, 19, 935. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Hamilton, M.; Vasquez, B.; He, M. 3D-printing enabled micro-assembly of a microfluidic electroporation system for 3D tissue engineering. Lab. Chip 2019, 19, 2362–2372. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liu, Z.; Ma, Y.; Zhang, K.; Qin, L. Cas9 Ribonucleoprotein Delivery via Microfluidic Cell-Deformation Chip for Human T-Cell Genome Editing and Immunotherapy. Adv. Biosyst. 2017, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemmi, H.; Takeuchi, O.; Kawai, T.; Kaisho, T.; Sato, S.; Sanjo, H.; Matsumoto, M.; Hoshino, K.; Wagner, H.; Takeda, K.; et al. A Toll-like receptor recognizes bacterial DNA. Nat. Cell Biol. 2000, 408, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liu, Z.; Zhao, L.; Wang, F.; Yu, Y.; Yang, J.; Chen, R.; Qin, L. Microfluidic Cell Deformability Assay for Rapid and Efficient Kinase Screening with the CRISPR-Cas9 System. Angew Chem. 2016, 128, 8703–8707. [Google Scholar] [CrossRef]
- Byun, S.; Son, S.; Amodei, D.; Cermak, N.; Shaw, J.; Kang, J.H.; Hecht, V.C.; Winslow, M.M.; Jacks, T.; Mallicj, P.; et al. Characterizing deformability and surface friction of cancer cells. Proc. Natl. Acad. Sci. USA 2013, 110, 7580–7585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Kai, K.; Choi, D.S.; Iwamoto, T.; Nguyen, Y.H.; Wong, H.; Landis, M.D.; Ueno, N.T.; Chang, J.; Qin, L. Microfluidics separation reveals the stem-cell-like deformability of tumor-initiating cells. Proc. Natl. Acad. Sci. USA 2012, 109, 18707–18712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, M.; Takemoto, T. Electroporation enables the efficient mRNA delivery into the mouse zygotes and facilitates CRISPR/Cas9-based genome editing. Sci. Rep. 2015, 5, 11315. [Google Scholar] [CrossRef] [Green Version]
- Geng, T.; Bao, N.; Sriranganathanw, N.; Li, L.; Lu, C. Genomic DNA Extraction from Cells by Electroporation on an Integrated Microfluidic Platform. Anal. Chem. 2012, 84, 9632–9639. [Google Scholar] [CrossRef] [Green Version]
- Bao, N.; Wang, J.; Lu, C. Microfluidic electroporation for selective release of intracellular molecules at the single-cell level. Electrophoresis 2008, 29, 2939–2944. [Google Scholar] [CrossRef]
- Wang, J.; Bao, N.; Paris, L.L.; Wang, H.-Y.; Geahlen, R.L.; Lu, C. Detection of Kinase Translocation Using Microfluidic Electroporative Flow Cytometry. Anal. Chem. 2008, 80, 1087–1093. [Google Scholar] [CrossRef] [Green Version]
- Bian, S.; Zhou, Y.; Hu, Y.; Cheng, J.; Chen, X.; Xu, Y.; Liu, P. High-throughput in situ cell electroporation microsystem for parallel delivery of single guide RNAs into mammalian cells. Sci. Rep. 2017, 7, srep42512. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Zhang, J.; Bian, S.; Chen, Z.; Hu, Y.; Hu, R.; Li, J.; Cheng, Y.; Zhang, X.; Zhou, Y.; et al. High-throughput superhydrophobic microwell arrays for investigating multifactorial stem cell niches. Lab. Chip 2016, 16, 2996–3006. [Google Scholar] [CrossRef]
- Szeto, G.L.; Van Egeren, D.; Worku, H.; Sharei, A.; Alejandro, B.; Park, C.; Frew, K.; Brefo, M.; Mao, S.; Heimann, M.; et al. Microfluidic squeezing for intracellular antigen loading in polyclonal B-cells as cellular vaccines. Sci Rep. 2015, 5, 10276. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, B.; Juba, B.M.; Vazquez, M.; Kortum, S.W.; Pierce, B.S.; Pacheco, M.; Roberts, L.; Strohbach, J.W.; Jones, L.H.; et al. Microfluidic-Enabled Intracellular Delivery of Membrane Impermeable Inhibitors to Study Target Engagement in Human Primary Cells. ACS Chem. Biol. 2017, 12, 2970–2974. [Google Scholar] [CrossRef]
- DiTommaso, T.; Cole, J.M.; Cassereau, L.; Buggé, J.A.; Hanson, J.L.S.; Bridgen, D.T.; Stokes, B.D.; Loughhead, S.M.; Beutel, B.A.; Kathrin Nussbaum, G.; et al. Cell engineering with micro-fluidic squeezing preserves functionality of primary immune cells in vivo. Proc. Natl. Acad. Sci. USA 2018, 115, E10907–E10914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Koo, T.; Park, S.W.; Kim, D.; Kim, K.; Cho, H.-Y.; Song, D.W.; Lee, K.J.; Jung, M.H.; Kim, S.; et al. In vivo genome editing with a small Cas9 orthologue derived from Campylobacter jejuni. Nat. Commun. 2017, 8, 14500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, M.; Lee, C.; Gasiunas, G.; Davis, T.H.; Cradick, T.; Siksnys, V.; Bao, G.; Cathomen, T.; Mussolino, C. Streptococcus thermophilus CRISPR-Cas9 Systems Enable Specific Editing of the Human Genome. Mol. Ther. 2016, 24, 636–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawluk, A.; Amrani, N.; Zhang, Y.; Garcia, B.; Hidalgo-Reyes, Y.; Lee, J.; Edraki, A.; Shah, M.; Sontheimer, E.J.; Maxwell, K.L.; et al. Naturally Occurring Off-Switches for CRISPR-Cas9. Cell 2016, 167, 1829–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrington, L.B.; Paez-Espino, D.; Staahl, B.T.; Chen, J.S.; Ma, E.; Kyrpides, N.C.; Doudna, J.A. A thermostable Cas9 with increased lifetime in human plasma. Nat. Commun. 2017, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hou, Z.; Zhang, Y.; Propson, N.E.; Howden, S.; Chu, L.-F.; Sontheimer, E.J.; Thomson, J.A. Efficient genome engineering in human pluripotent stem cells using Cas9 from Neisseria meningitidis. Proc. Natl. Acad. Sci. USA 2013, 110, 15644–15649. [Google Scholar] [CrossRef] [Green Version]
- Joung, J.; Ladha, A.; Saito, M.; Segel, M.; Bruneau, R.; Huang, M.W.; Kim, N.-G.; Yu, X.; Li, J.; Walker, B.D.; et al. Point-of-care testing for COVID-19 using SHERLOCK diagnostics. medRxiv 2020. [Google Scholar] [CrossRef]
- Qin, P.; Park, M.; Alfson, K.J.; Tamhankar, M.; Carrion, R.; Patterson, J.L.; Griffiths, A.; He, Q.; Yildiz, A.; Mathies, R.A.; et al. Rapid and Fully Microfluidic Ebola Virus Detection with CRISPR-Cas13a. ACS Sens. 2019, 4, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Servellita, V.; Singh, J.; Miao, X.; Streithorst, J.A.; Granados, A.; Sotomayor-Gonzalez, A.; et al. CRISPR–Cas12-based detection of SARS-CoV-2. Nat. Biotechnol. 2020, 38, 870–874. [Google Scholar] [CrossRef] [Green Version]
- Rogacs, A.; Marshall, L.A.; Santiago, J.G. Purification of nucleic acids using isotachophoresis. J. Chromatogr. A 2014, 1335, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Huyke, D.A.; Sharma, E.; Sahoo, M.K.; Banaei, N.; Pinsky, B.A.; Santiago, J.G. Electric-field-driven micro-fluidics for rapid CRISPR-based diagnostics and its application to detection of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 29518–29525. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, G.W.; Frey, O.; Rudolf, F.; Muzi-Falconi, M.; Brown, G.W. The CellClamper: A Convenient Microfluidic Device for Time-Lapse Imaging of Yeast. Adv. Struct. Saf. Stud. 2017, 1672, 537–555. [Google Scholar] [CrossRef]
- Hass, K.; Bao, M.; He, Q.; Park, M.; Qin, P.; Du, K. Integrated Micropillar Polydimethylsiloxane Accurate CRISPR Detection (IMPACT) System for Rapid Viral DNA Sensing. ACS Omega 2020, 5, 27433–27441. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peyravian, N.; Malekzadeh Kebria, M.; Kiani, J.; Brouki Milan, P.; Mozafari, M. CRISPR-Associated (CAS) Effectors Delivery via Microfluidic Cell-Deformation Chip. Materials 2021, 14, 3164. https://0-doi-org.brum.beds.ac.uk/10.3390/ma14123164
Peyravian N, Malekzadeh Kebria M, Kiani J, Brouki Milan P, Mozafari M. CRISPR-Associated (CAS) Effectors Delivery via Microfluidic Cell-Deformation Chip. Materials. 2021; 14(12):3164. https://0-doi-org.brum.beds.ac.uk/10.3390/ma14123164
Chicago/Turabian StylePeyravian, Noshad, Maziar Malekzadeh Kebria, Jafar Kiani, Peiman Brouki Milan, and Masoud Mozafari. 2021. "CRISPR-Associated (CAS) Effectors Delivery via Microfluidic Cell-Deformation Chip" Materials 14, no. 12: 3164. https://0-doi-org.brum.beds.ac.uk/10.3390/ma14123164