
AFM and Fluorescence Microscopy of Single Cells with Simultaneous Mechanical Stimulation via Electrically Stretchable Substrates

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
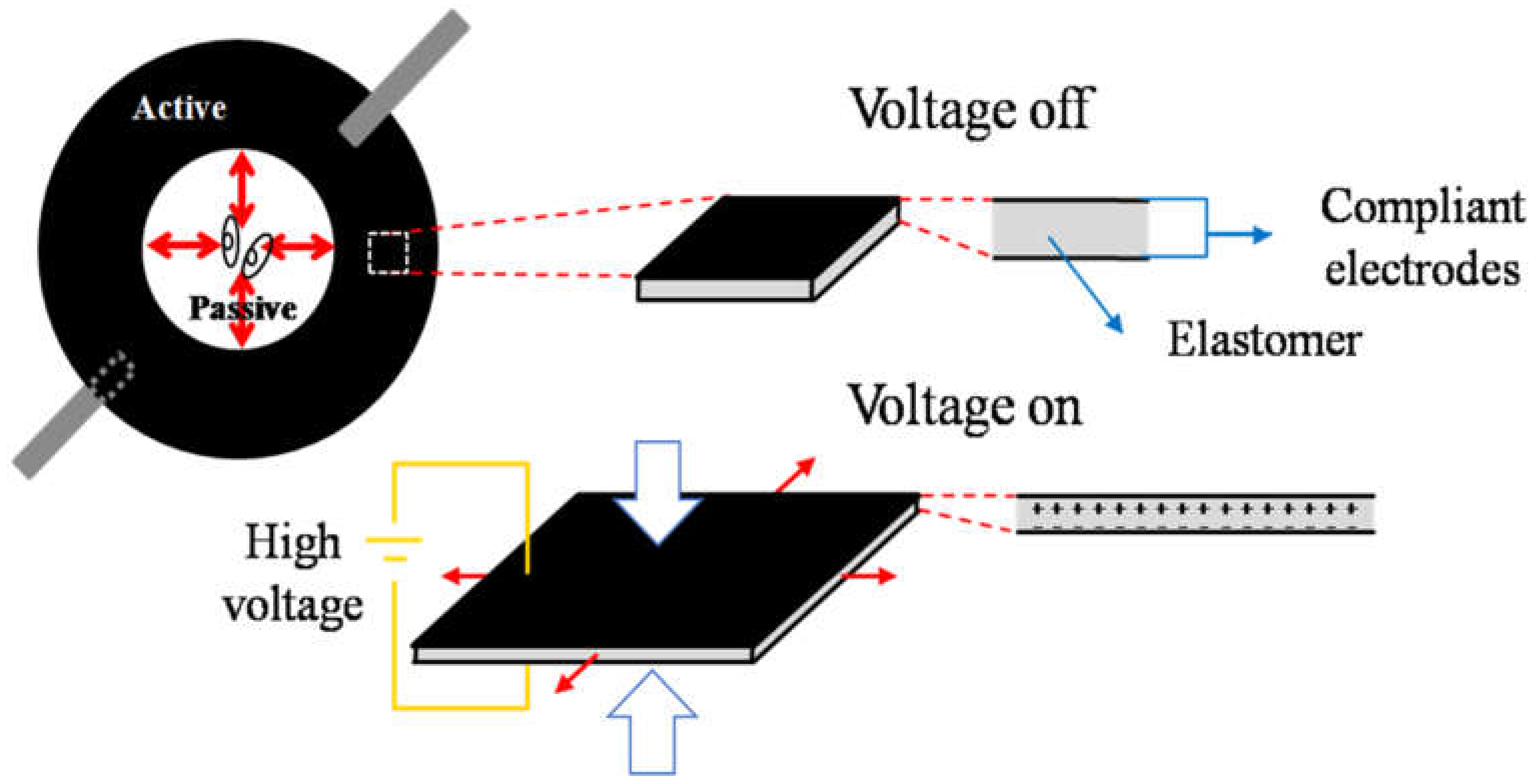
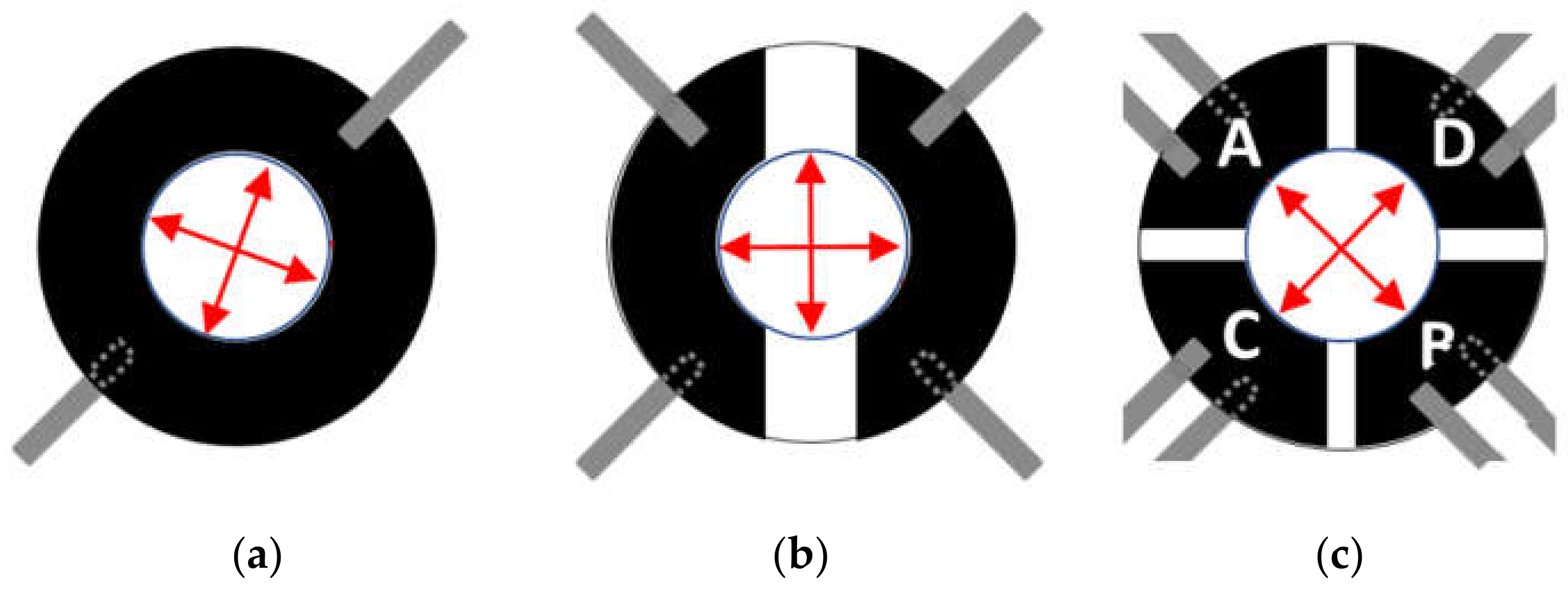
2.1. Cell Stretcher Device Working Principle
2.2. Cell Stretcher Device Fabrication
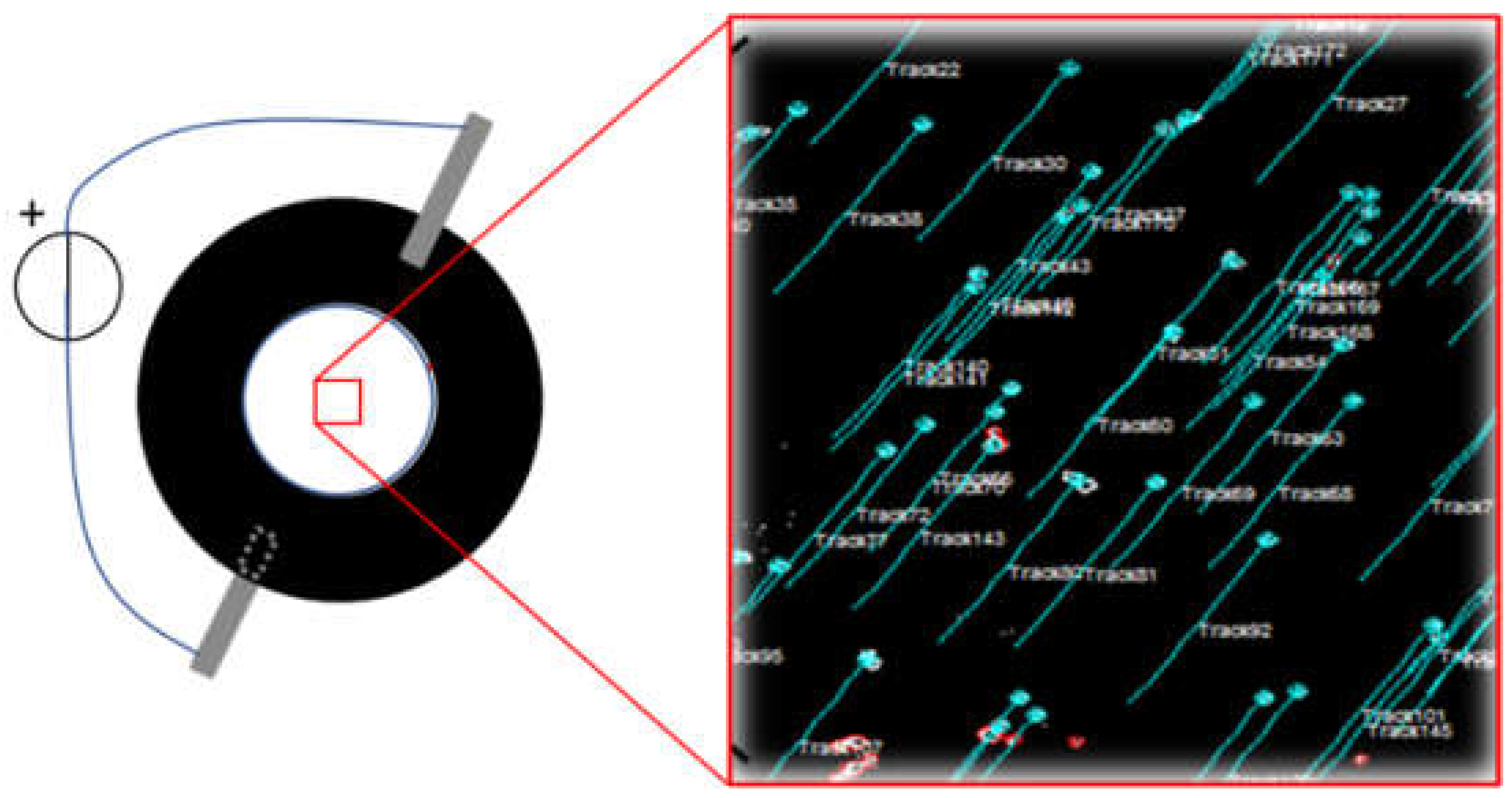
2.3. Strain Calibration
2.4. Cell Culture
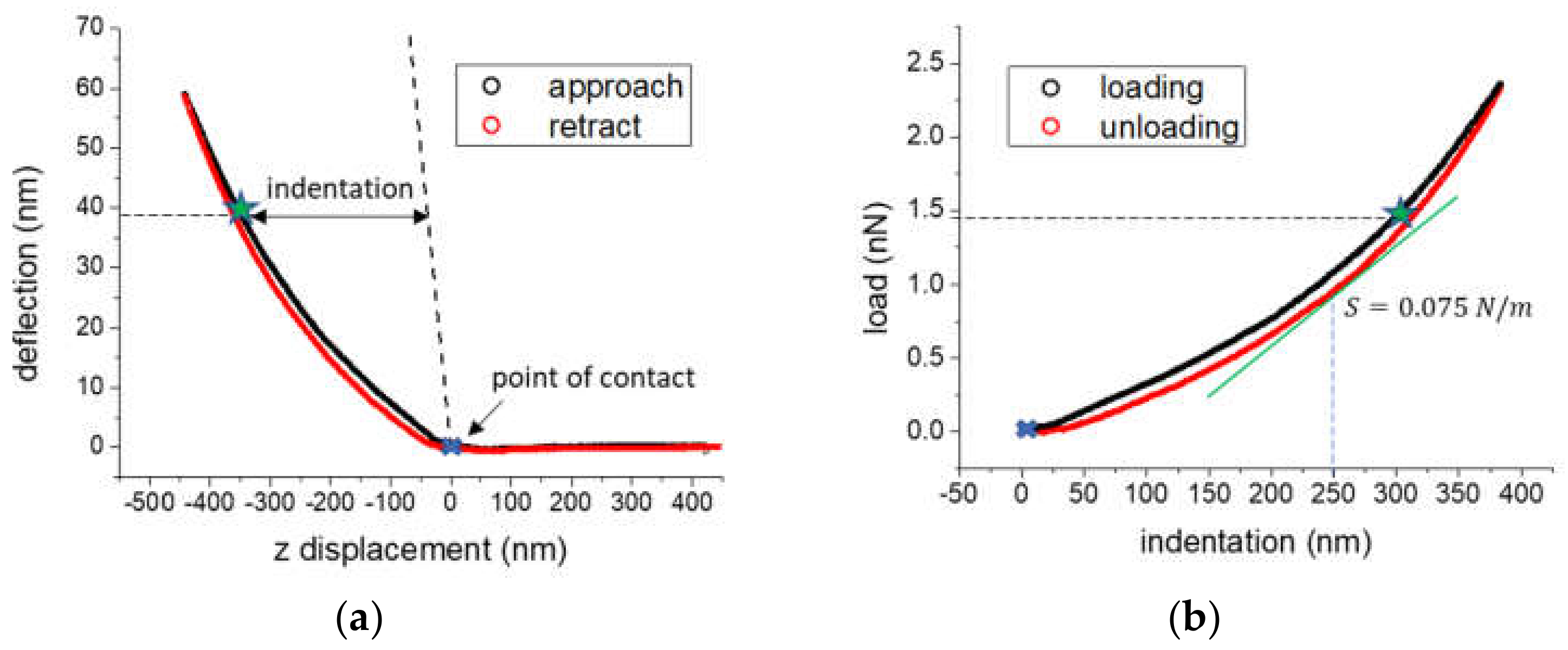
2.5. Atomic Force Microscopy (AFM)
3. Results and Discussion
3.1. Cell Stretcher Device Performance
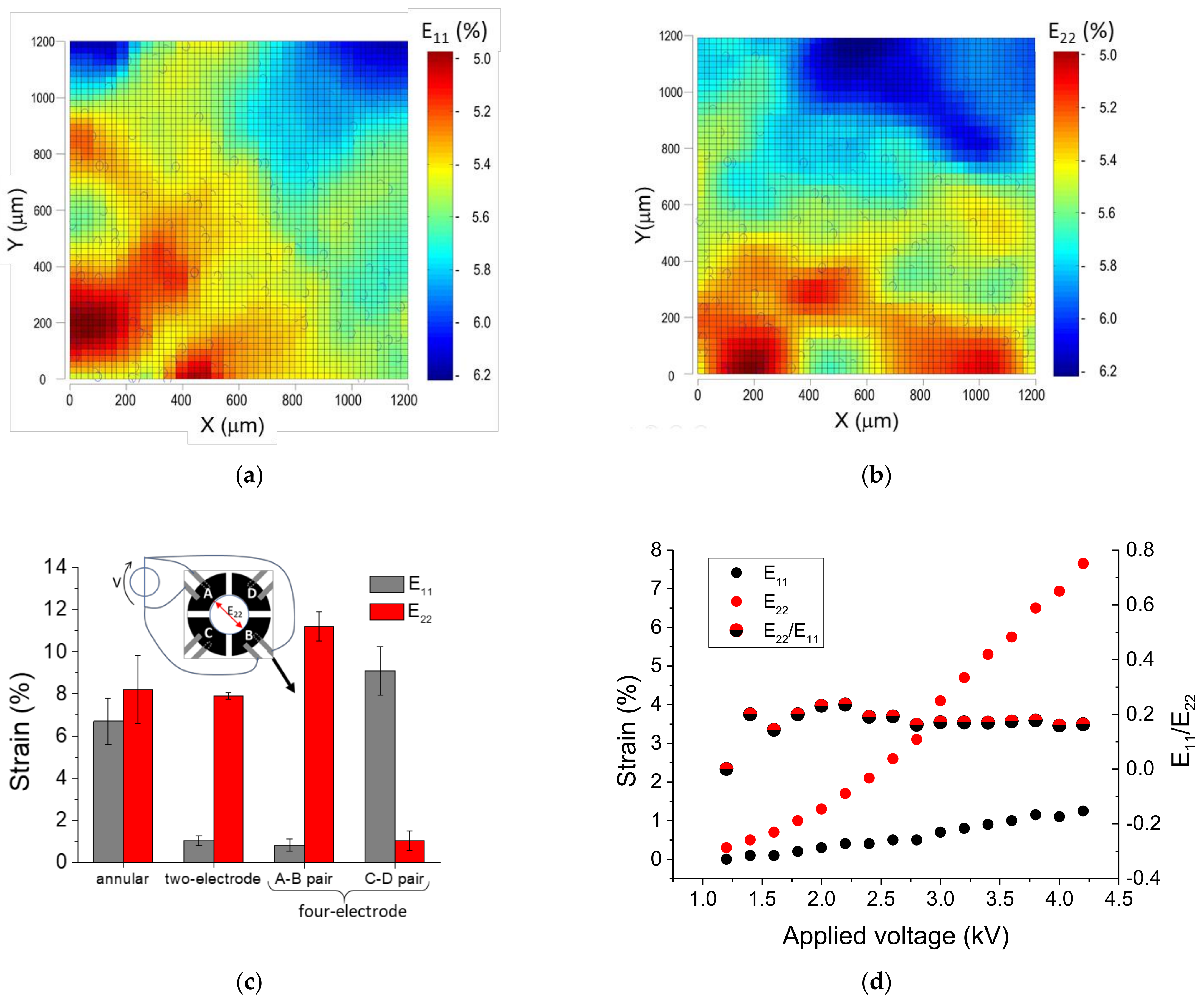
3.1.1. Static Strain Response
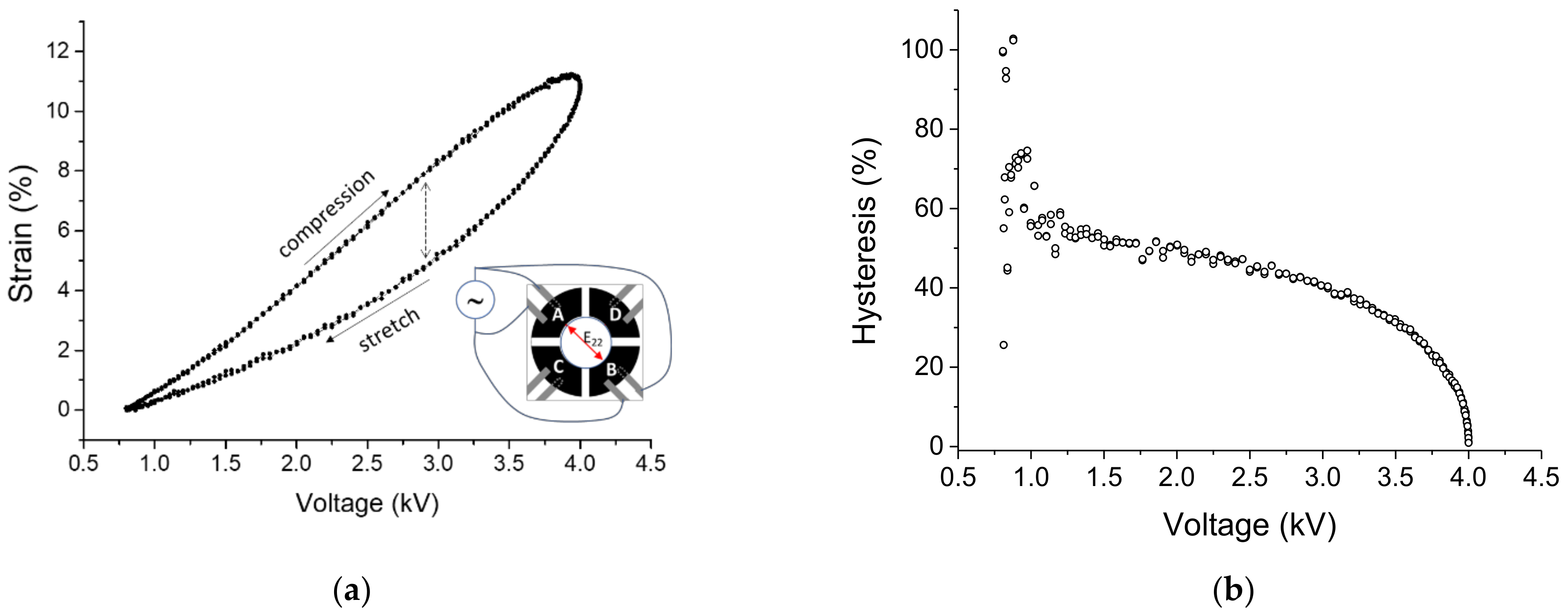
3.1.2. Dynamic Strain Response
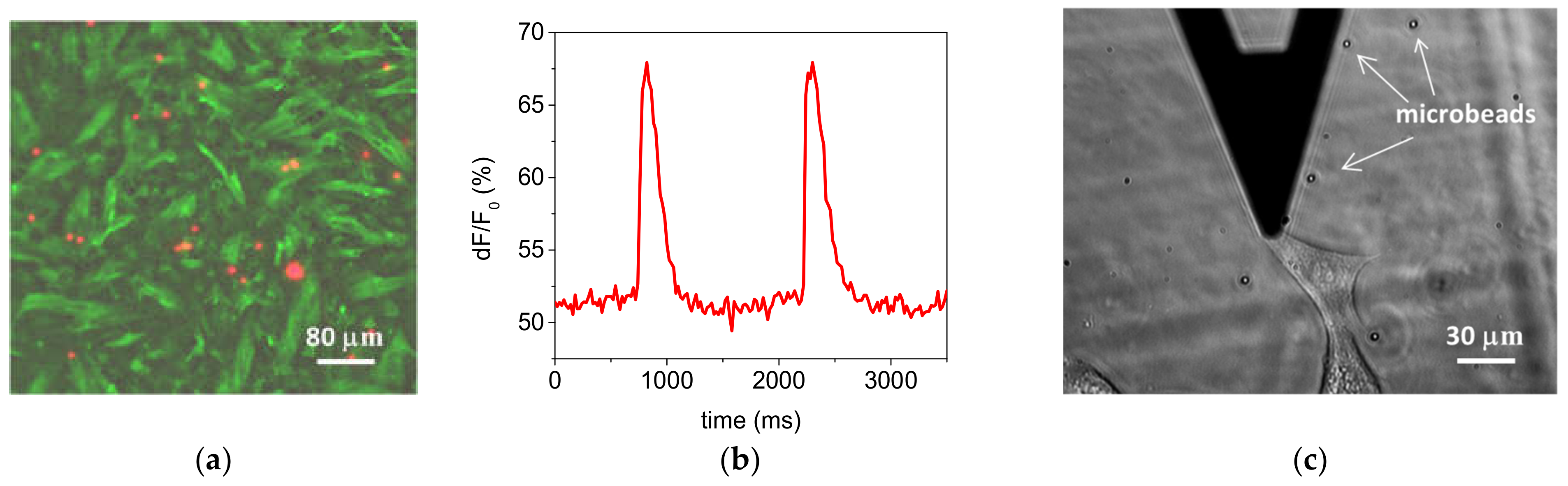
3.1.3. Optical Performance and Biocompatibility
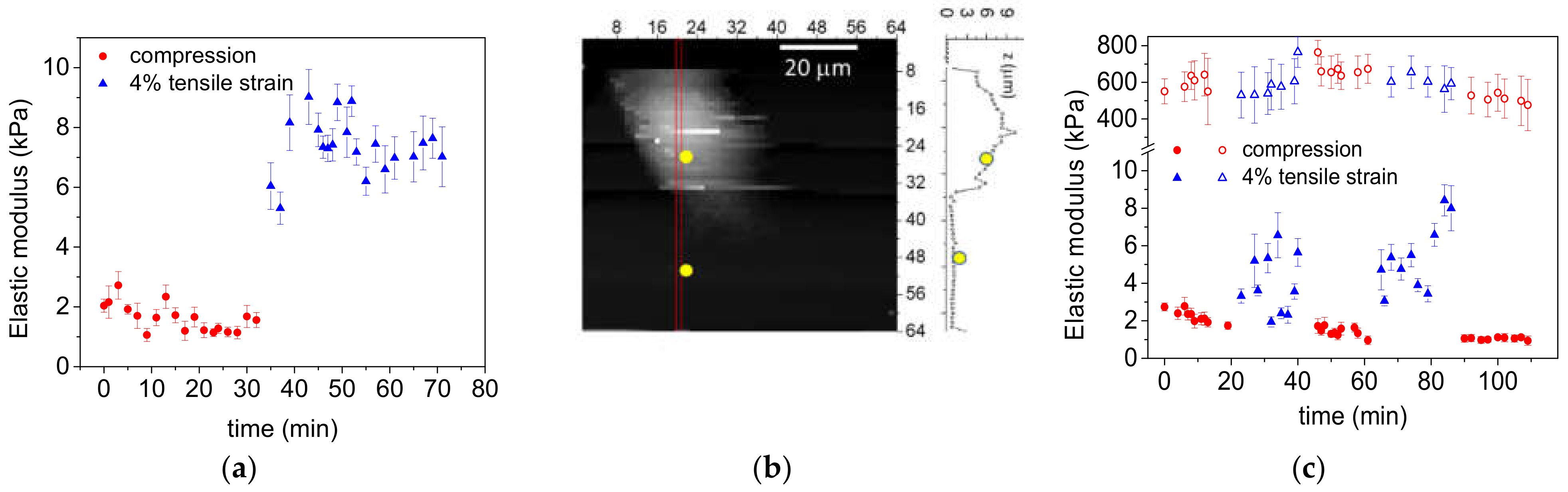
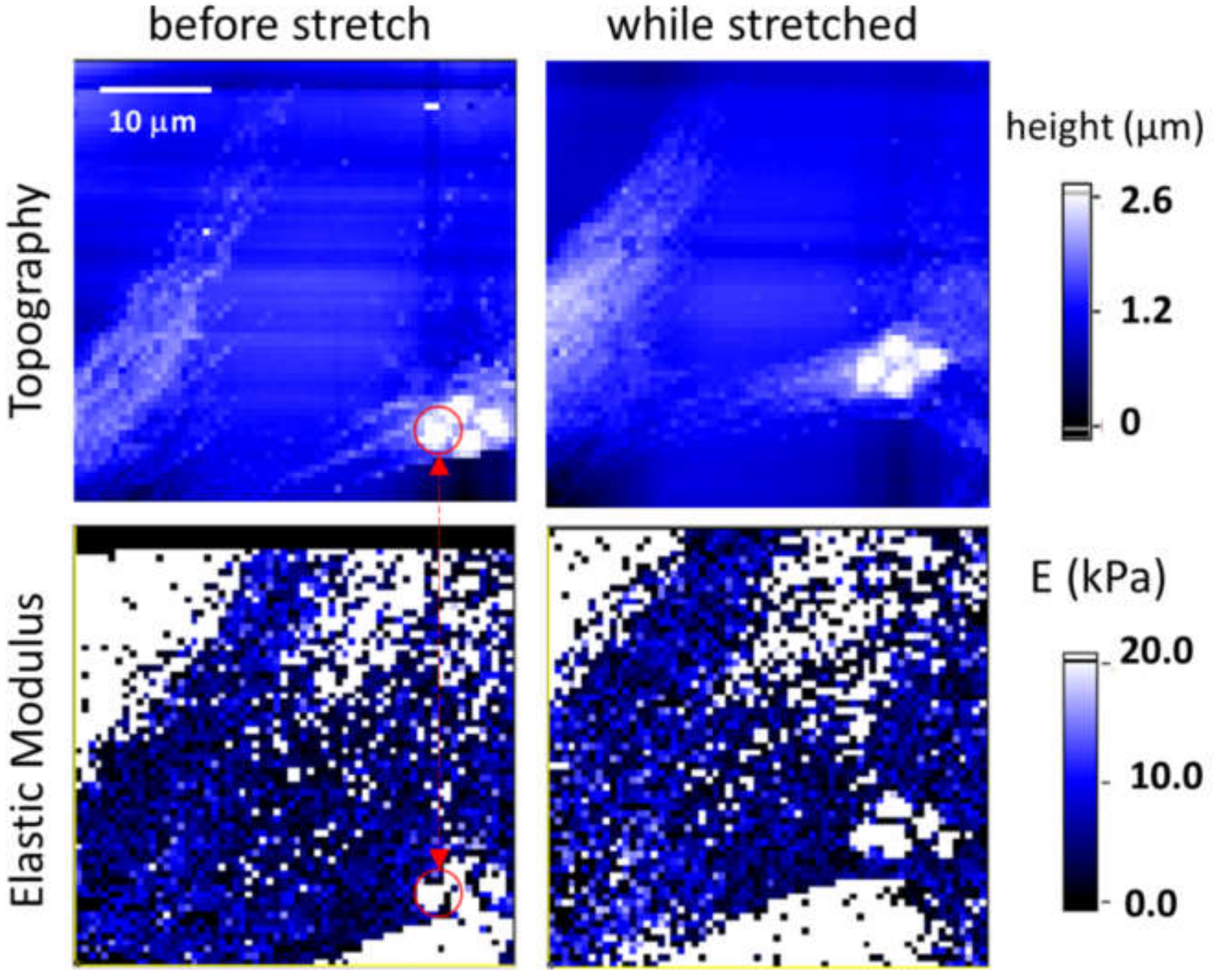
3.2. Cell Elasticity Response to Strain
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pelrine, R.; Kornbluh, R.; Pei, Q.; Joseph, J. High-Speed Electrically Actuated Elastomers with Strain Greater Than 100%. Science 2000, 287, 836–839. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, K.; Mizutani, T.; Tamura, K.; Takeda, K.; Haga, H.; Kawabata, K. The Number of Cyclic Stretch Regulates Cellular Elasticity in C2C12 Myoblasts. CellBio 2012, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, K.; Sato, N.; Obinata, T. Dynamic Reorientation of Cultured Cells and Stress Fibers under Mechanical Stress from Periodic Stretching. Exp. Cell Res. 2001, 268, 104–114. [Google Scholar] [CrossRef]
- Brown, T.D. Techniques for Mechanical Stimulation of Cells in Vitro: A Review. J. Biomech. 2000, 33, 3–14. [Google Scholar] [CrossRef]
- Davis, C.A.; Zambrano, S.; Anumolu, P.; Allen, A.C.B.; Sonoqui, L.; Moreno, M.R. Device-Based in Vitro Techniques for Mechanical Stimulation of Vascular Cells: A Review. J. Biomech. Eng. 2015, 137, 040801. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Porter, C.; Crocker, J.; Reich, D. Dissecting Fat-Tailed Fluctuations in the Cytoskeleton with Active Micropost Arrays. Proc. Natl. Acad. Sci. USA 2019, 116, 201900963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Pak, K.W.; Park, J.; Levchenko, A.; Sun, Y. Microengineered Platforms for Cell Mechanobiology. Annu. Rev. Biomed. Eng. 2009, 11, 203–233. [Google Scholar] [CrossRef] [Green Version]
- Carpi, F.; de Rossi, D.; Kornbluh, R.; Pelrine, R. Dielectric Elastomers as Electromechanical Transducers; Elsevier: Amsterdam, The Netherlands, 2008; ISBN 9780080474885. [Google Scholar]
- Carpi, F.; Frediani, G.; Turco, S.; de Rossi, D. Bioinspired Tunable Lens with Muscle-Like Electroactive Elastomers. Adv. Funct. Mater. 2011, 21, 4152–4158. [Google Scholar] [CrossRef]
- Carpi, F.; Kornbluh, R.; Sommer-Larsen, P.; Alici, G. Electroactive Polymer Actuators as Artificial Muscles: Are They Ready for Bioinspired Applications? Bioinspir. Biomim. 2011, 6, 045006. [Google Scholar] [CrossRef]
- Brochu, P.; Pei, Q. Advances in Dielectric Elastomers for Actuators and Artificial Muscles. Macromol. Rapid Commun. 2010, 31, 10–36. [Google Scholar] [CrossRef]
- Kim, D.-U.; Lee, S.; Chang, S.-H. Dynamic Cell Culture Device Using Electroactive Polymer Actuators with Composite Electrodes to Transfer In-Plane Mechanical Strain to Cells. Int. J. Precis. Eng. Manuf. Green Technol. 2021, 8, 969–980. [Google Scholar] [CrossRef]
- Costa, J.; Ghilardi, M.; Mamone, V.; Ferrari, V.; Busfield, J.J.C.; Ahluwalia, A.; Carpi, F. Bioreactor With Electrically Deformable Curved Membranes for Mechanical Stimulation of Cell Cultures. Front. Bioeng. Biotechnol. 2020, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Li, Z.; Zou, J.; Cheng, J.; Jiang, K.; Liu, C.; Gu, G.; Tao, W.; Song, J. Mechanical Effect on Gene Transfection Based on Dielectric Elastomer Actuator. ACS Appl. Bio Mater. 2020, 3, 2617–2625. [Google Scholar] [CrossRef]
- Janmey, P.A.; McCulloch, C.A. Cell Mechanics: Integrating Cell Responses to Mechanical Stimuli. Annu. Rev. Biomed. Eng. 2007, 9, 1–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopalan, S.M.; Flaim, C.; Bhatia, S.N.; Hoshijima, M.; Knoell, R.; Chien, K.R.; Omens, J.H.; McCulloch, A.D. Anisotropic Stretch-Induced Hypertrophy in Neonatal Ventricular Myocytes Micropatterned on Deformable Elastomers. Biotechnol. Bioeng. 2003, 81, 578–587. [Google Scholar] [CrossRef]
- Prosser, B.L.; Khairallah, R.J.; Ziman, A.P.; Ward, C.W.; Lederer, W.J. X-ROS Signaling in the Heart and Skeletal Muscle: Stretch-Dependent Local ROS Regulates [Ca2+]i. J. Mol. Cell. Cardiol. 2013, 58, 172–181. [Google Scholar] [CrossRef] [Green Version]
- Prosser, B.L.; Ward, C.W.; Lederer, W.J. X-ROS Signalling Is Enhanced and Graded by Cyclic Cardiomyocyte Stretch. Cardiovasc. Res. 2013, 98, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.; Vu, D.K.; Steinmann, P. A Comprehensive Characterization of the Electro-Mechanically Coupled Properties of VHB 4910 Polymer. Arch. Appl. Mech. 2015, 85, 523–537. [Google Scholar] [CrossRef]
- Mehnert, M.; Hossain, M.; Steinmann, P. Experimental and Numerical Investigations of the Electro-Viscoelastic Behavior of VHB 4905TM. Eur. J. Mech. A/Solids 2019, 77, 103797. [Google Scholar] [CrossRef]
- Kofod, G. The Static Actuation of Dielectric Elastomer Actuators: How Does Pre-Stretch Improve Actuation? J. Phys. D Appl. Phys. 2008, 41. [Google Scholar] [CrossRef]
- Palchesko, R.N.; Zhang, L.; Sun, Y.; Feinberg, A.W. Development of Polydimethylsiloxane Substrates with Tunable Elastic Modulus to Study Cell Mechanobiology in Muscle and Nerve. PLoS ONE 2012, 7, 51499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dipalo, M.; Melle, G.; Lovato, L.; Jacassi, A.; Santoro, F.; Caprettini, V.; Schirato, A.; Alabastri, A.; Garoli, D.; Bruno, G.; et al. Plasmonic Meta-Electrodes Allow Intracellular Recordings at Network Level on High-Density CMOS-Multi-Electrode Arrays. Nat. Nanotechnol. 2018, 13, 965–971. [Google Scholar] [CrossRef]
- Cogollo, J.F.S.; Tedesco, M.; Martinoia, S.; Raiteri, R. A New Integrated System Combining Atomic Force Microscopy and Micro-Electrode Array for Measuring the Mechanical Properties of Living Cardiac Myocytes. Biomed. Microdevices 2011, 13. [Google Scholar] [CrossRef]
- Spanu, A.; Lai, S.; Cosseddu, P.; Tedesco, M.; Martinoia, S.; Bonfiglio, A. An Organic Transistor-Based System for Reference-Less Electrophysiological Monitoring of Excitable Cells. Sci. Rep. 2015, 5, 8807. [Google Scholar] [CrossRef] [Green Version]
- Hutter, J.L.; Bechhoefer, J. Calibration of Atomic—Force Microscope Tips. Rev. Sci. Instrum. 1993, 64, 1868–1873. [Google Scholar] [CrossRef] [Green Version]
- Oliver, W.C.; Pharr, G.M. An Improved Technique for Determining Hardness and Elastic Modulus Using Load and Displacement Sensing Indentation Experiments. J. Mater. Res. 1992, 7, 1564–1583. [Google Scholar] [CrossRef]
- Vanlandingham, M.; Villarrubia, J.; Guthrie, W.; Meyers, G. Nanoindentation of Polymers: An Overview. Macromol. Symp. 2001, 167. [Google Scholar] [CrossRef]
- Ahrens, D.; Rubner, W.; Springer, R.; Hampe, N.; Gehlen, J.; Magin, T.M.; Hoffmann, B.; Merkel, R. A Combined AFM and Lateral Stretch Device Enables Microindentation Analyses of Living Cells at High Strains. Methods Protoc. 2019, 2, 43. [Google Scholar] [CrossRef] [Green Version]
- Quinlan, A.M.; Sierad, L.N.; Capulli, A.K.; Firstenberg, L.E.; Billiar, K.L. Combining Dynamic Stretch and Tunable Stiffness to Probe Cell Mechanobiology in Vitro. PLoS ONE 2011, 6, e23272. [Google Scholar] [CrossRef]
- Kang, H.; Wong, D.S.H.; Yan, X.; Jung, H.J.; Kim, S.; Lin, S.; Wei, K.; Li, G.; Dravid, V.P.; Bian, L. Remote Control of Multimodal Nanoscale Ligand Oscillations Regulates Stem Cell Adhesion and Differentiation. ACS Nano 2017, 11, 9636–9649. [Google Scholar] [CrossRef]
- Tremblay, D.; Andrzejewski, L.; Leclerc, A.; Pelling, A.E. Actin and Microtubules Play Distinct Roles in Governing the Anisotropic Deformation of Cell Nuclei in Response to Substrate Strain. Cytoskeleton 2013, 70, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Hecht, E.; Knittel, P.; Felder, E.; Dietl, P.; Mizaikoff, B.; Kranz, C. Combining Atomic Force-Fluorescence Microscopy with a Stretching Device for Analyzing Mechanotransduction Processes in Living Cells. Analyst 2012, 137, 5208–5214. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becerra, N.; Salis, B.; Tedesco, M.; Moreno Flores, S.; Vena, P.; Raiteri, R. AFM and Fluorescence Microscopy of Single Cells with Simultaneous Mechanical Stimulation via Electrically Stretchable Substrates. Materials 2021, 14, 4131. https://0-doi-org.brum.beds.ac.uk/10.3390/ma14154131
Becerra N, Salis B, Tedesco M, Moreno Flores S, Vena P, Raiteri R. AFM and Fluorescence Microscopy of Single Cells with Simultaneous Mechanical Stimulation via Electrically Stretchable Substrates. Materials. 2021; 14(15):4131. https://0-doi-org.brum.beds.ac.uk/10.3390/ma14154131
Chicago/Turabian StyleBecerra, Natalia, Barbara Salis, Mariateresa Tedesco, Susana Moreno Flores, Pasquale Vena, and Roberto Raiteri. 2021. "AFM and Fluorescence Microscopy of Single Cells with Simultaneous Mechanical Stimulation via Electrically Stretchable Substrates" Materials 14, no. 15: 4131. https://0-doi-org.brum.beds.ac.uk/10.3390/ma14154131