
Impact of Cross-Linked Hyaluronic Acid on Osteogenic Differentiation of SAOS-2 Cells in an Air-Lift Model
 , and
, and
Abstract
:1. Introduction
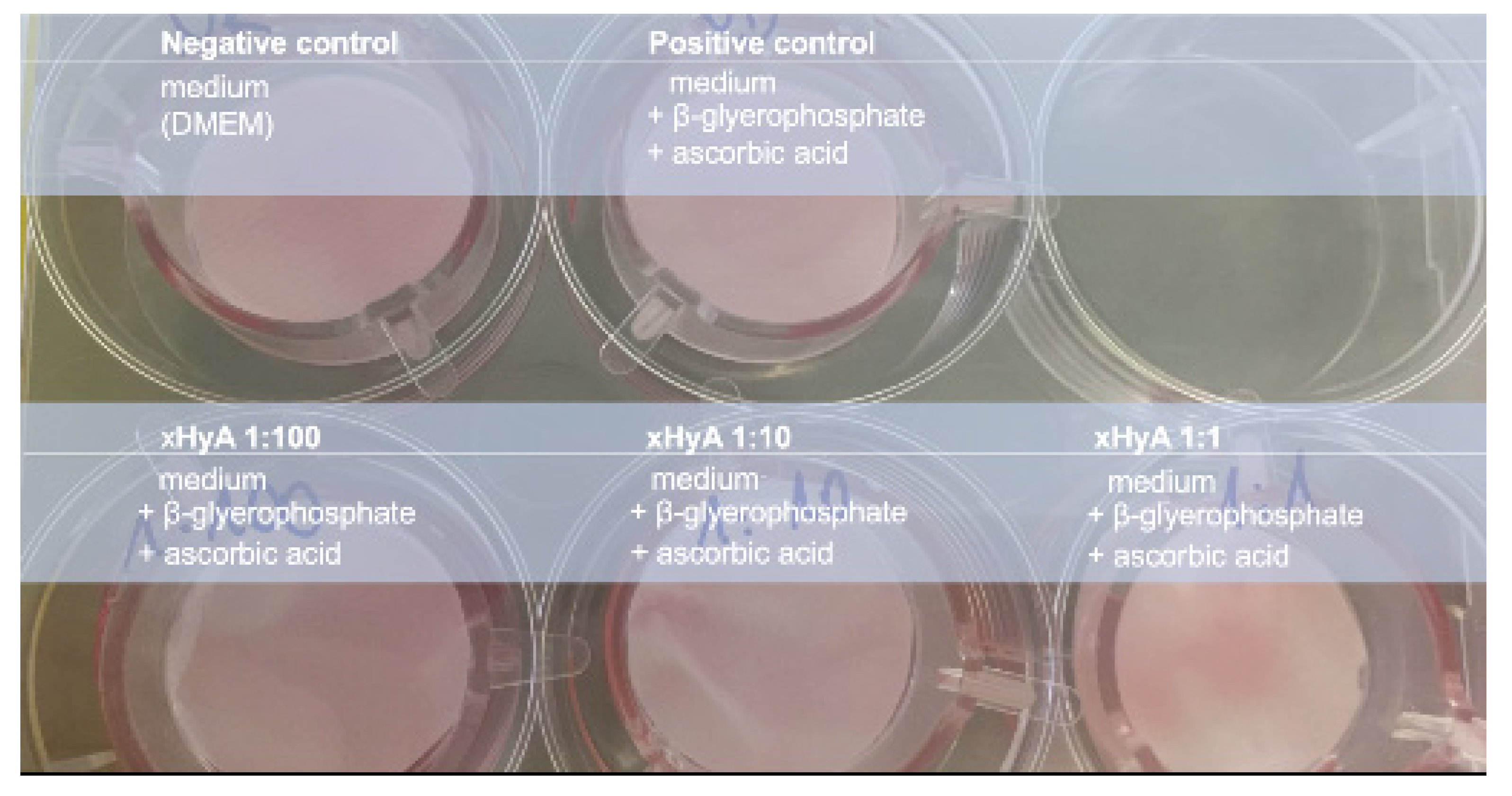
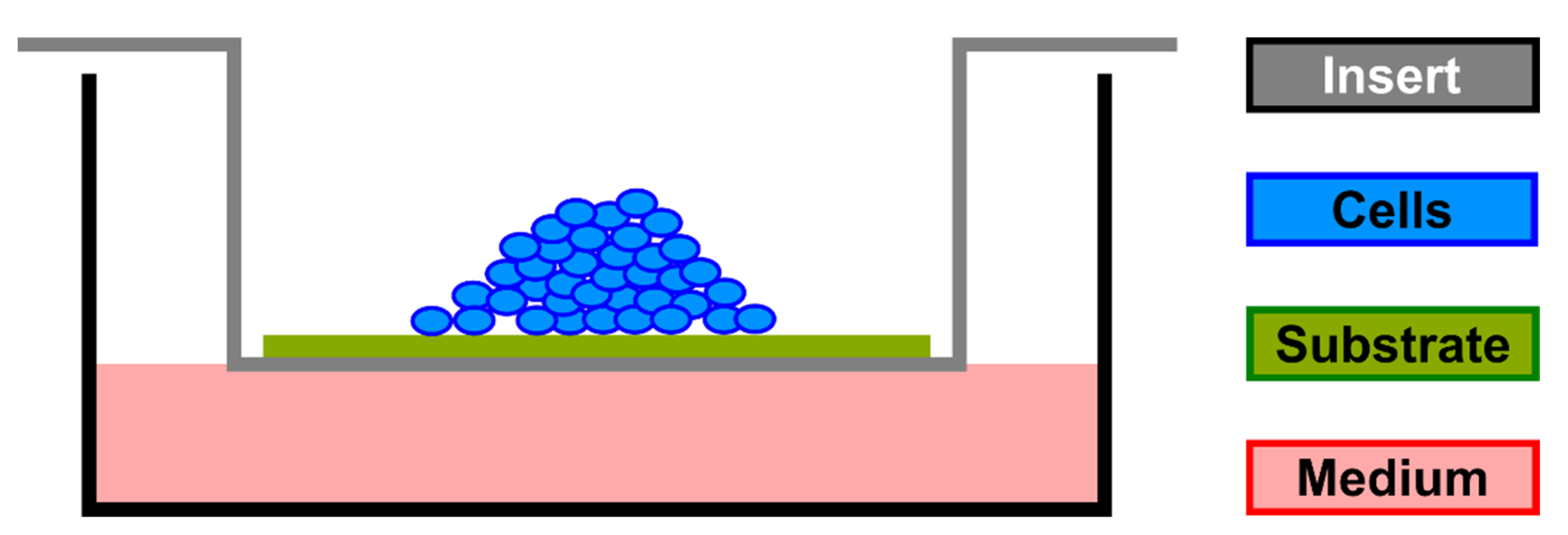
2. Materials and Methods
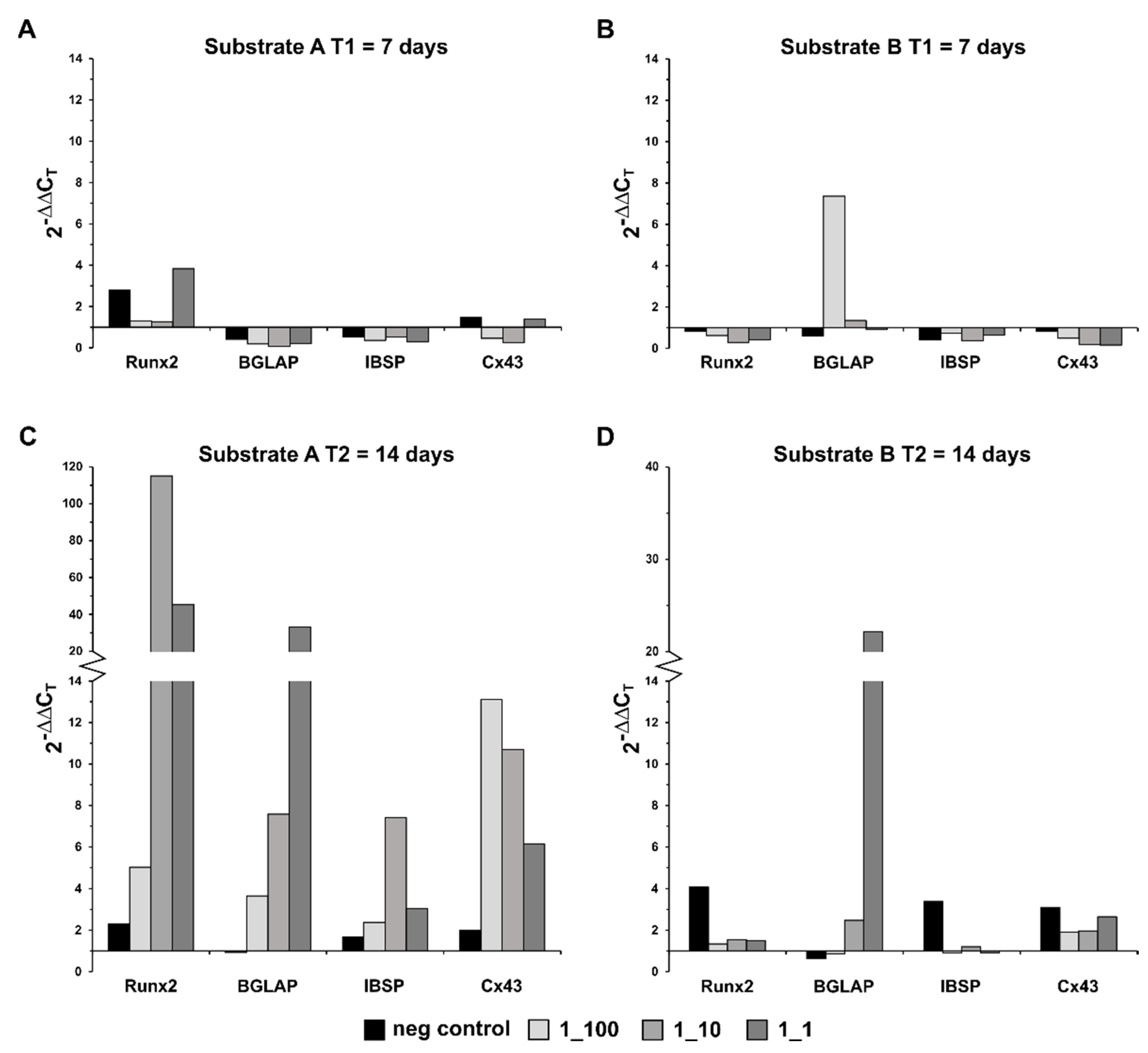
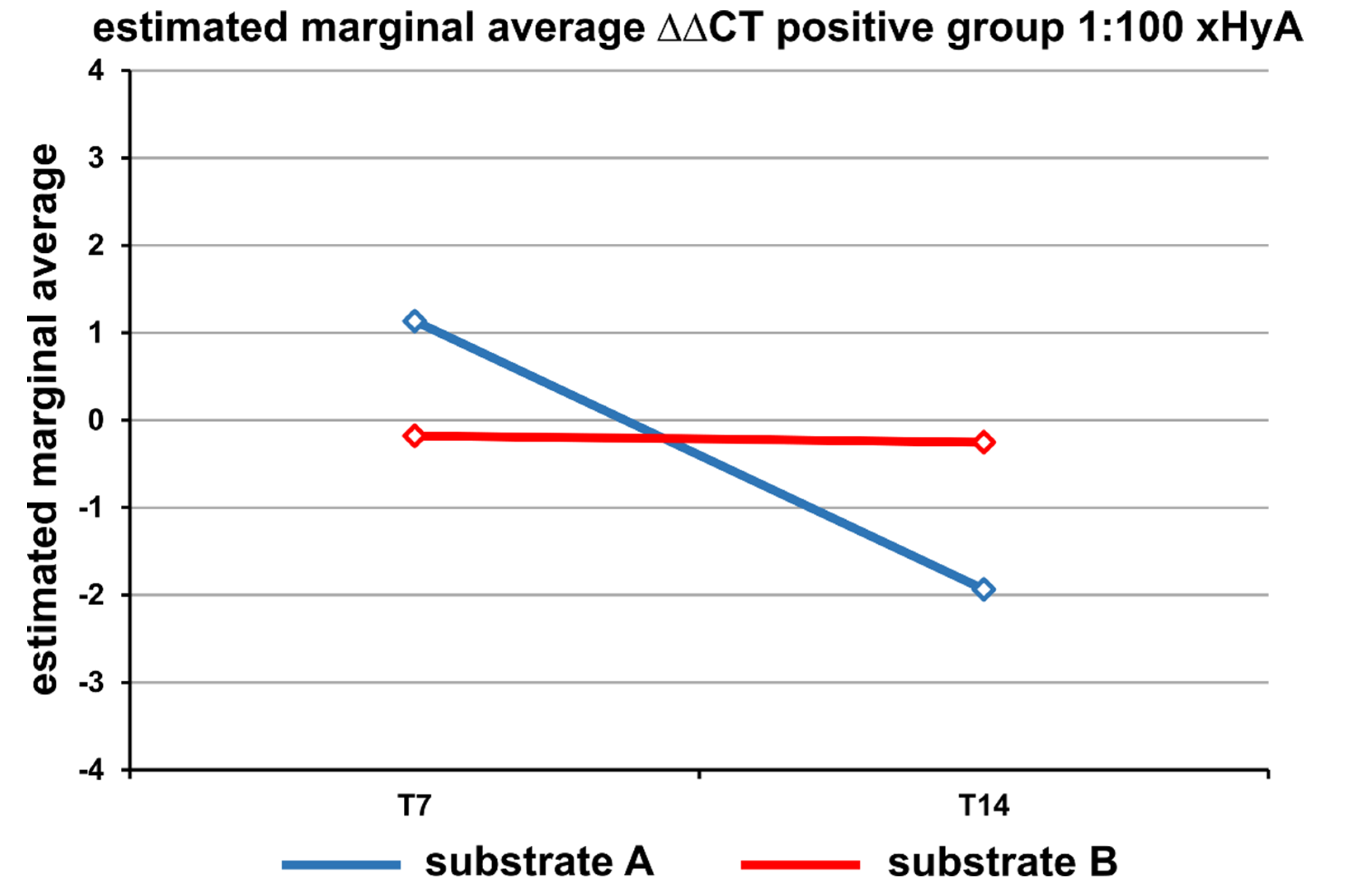
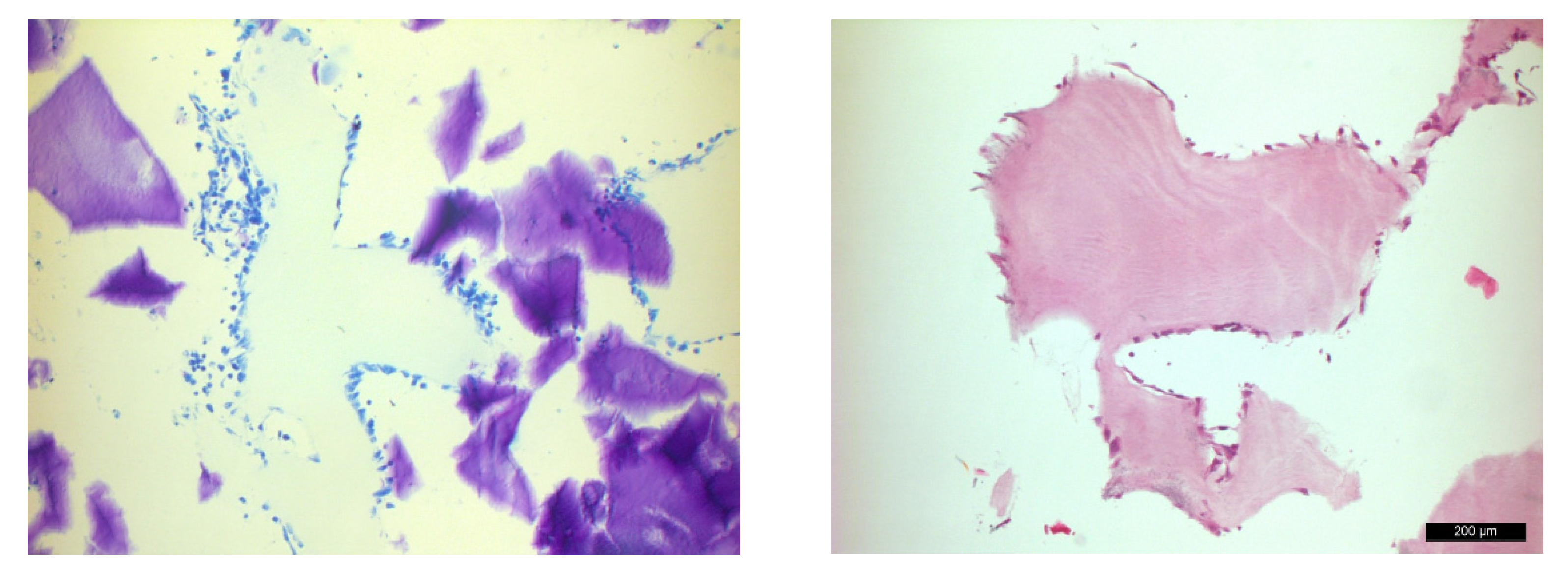
3. Results
Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aarden, E.M.; Wassenaar, A.M.; Alblas, M.J.; Nijweide, P.J. Immunocytochemical demonstration of extracellular matrix proteins in isolated osteocytes. Histochem. Cell Biol. 1996, 106, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Delmas, P.D.; Eastell, R.; Garnero, P.; Seibel, M.J.; Stepan, J.; Committee of Scientific Advisors of the International Osteoporosis Foundation. The use of biochemical markers of bone turnover in osteoporosis. Committee of Scientific Advisors of the International Osteoporosis Foundation. Osteoporos. Int. 2000, 11 (Suppl. 6), S2–S17. [Google Scholar] [CrossRef]
- Ducy, P.; Desbois, C.; Boyce, B.; Pinero, G.; Story, B.; Dunstan, C.; Smith, E.; Bonadio, J.; Goldstein, S.; Gundberg, C.; et al. Increased bone formation in osteocalcin-deficient mice. Nature 1996, 382, 448–452. [Google Scholar] [CrossRef]
- Poser, J.W.; Price, P.A. A method for decarboxylation of gamma-carboxyglutamic acid in proteins. Properties of the decarboxylated gamma-carboxyglutamic acid protein from calf bone. J. Biol. Chem. 1979, 254, 431–436. [Google Scholar] [CrossRef]
- Fisher, L.W.; McBride, O.W.; Termine, J.D.; Young, M.F. Human bone sialoprotein. Deduced protein sequence and chromosomal localization. J. Biol. Chem. 1990, 265, 2347–2351. [Google Scholar] [CrossRef]
- Kerr, J.M.; Fisher, L.W.; Termine, J.D.; Wang, M.G.; McBride, O.W.; Young, M.F. The human bone sialoprotein gene (IBSP): Genomic localization and characterization. Genomics 1993, 17, 408–415. [Google Scholar] [CrossRef]
- Oldberg, A.; Franzen, A.; Heinegard, D. The primary structure of a cell-binding bone sialoprotein. J. Biol. Chem. 1988, 263, 19430–19432. [Google Scholar] [CrossRef]
- Sodek, J.; Chen, J.; Nagata, T.; Kasugai, S.; Todescan, R., Jr.; Li, I.W.; Kim, R.H. Regulation of osteopontin expression in osteoblasts. Ann. N. Y. Acad. Sci. 1995, 760, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Sasaki, Y.; Ogata, Y. Calcium hydroxide regulates bone sialoprotein gene transcription in human osteoblast-like Saos2 cells. J. Oral Sci. 2011, 53, 77–86. [Google Scholar] [CrossRef]
- Ishikawa, M.; Yamada, Y. The Role of Pannexin 3 in Bone Biology. J. Dent. Res. 2017, 96, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Kobayashi, Y.; Koide, M.; Uehara, S.; Okamoto, M.; Ishihara, A.; Kayama, T.; Saito, M.; Marumo, K. The Regulation of Bone Metabolism and Disorders by Wnt Signaling. Int. J. Mol. Sci. 2019, 20, 5525. [Google Scholar] [CrossRef] [PubMed]
- Shapira, L.; Halabi, A. Behavior of two osteoblast-like cell lines cultured on machined or rough titanium surfaces. Clin. Oral Implant. Res. 2009, 20, 50–55. [Google Scholar] [CrossRef]
- Friedmann, A.; Dehnhardt, J.; Kleber, B.M.; Bernimoulin, J.P. Cytobiocompatibility of collagen and ePTFE membranes on osteoblast-like cells in vitro. J. Biomed. Mater. Res. A 2008, 86, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, B.; Wachtel, H.C.; Somogyi, H.; Merker, H.J.; Bernimoulin, J.P. Bone formation by rat calvarial cells grown at high density in organoid culture. Cell Differ. Dev. 1988, 25, 145–154. [Google Scholar] [CrossRef]
- Lin, W.; Liu, Z.; Kampf, N.; Klein, J. The Role of Hyaluronic Acid in Cartilage Boundary Lubrication. Cells 2020, 9, 1606. [Google Scholar] [CrossRef] [PubMed]
- Necas, J.; Bartosikova, L.; Brauner, P.; Kolar, J. Hyaluronic acid (hyaluronan): A review. Vet. Med. 2008, 8, 397–411. [Google Scholar] [CrossRef]
- Dahiya, P.; Kamal, R. Hyaluronic Acid: A boon in periodontal therapy. N. Am. J. Med. Sci. 2013, 5, 309–315. [Google Scholar] [CrossRef]
- Aslan, M.; Simsek, G.; Dayi, E. The effect of hyaluronic acid-supplemented bone graft in bone healing: Experimental study in rabbits. J. Biomater. Appl. 2006, 20, 209–220. [Google Scholar] [CrossRef]
- Kang, J.H.; Kim, Y.Y.; Chang, J.Y.; Kho, H.S. Influences of hyaluronic acid on the anticandidal activities of lysozyme and the peroxidase system. Oral Dis. 2011, 17, 577–583. [Google Scholar] [CrossRef]
- Sasaki, T.; Watanabe, C. Stimulation of osteoinduction in bone wound healing by high-molecular hyaluronic acid. Bone 1995, 16, 9–15. [Google Scholar] [CrossRef]
- Carlson, G.A.; Dragoo, J.L.; Samimi, B.; Bruckner, D.A.; Bernard, G.W.; Hedrick, M.; Benhaim, P. Bacteriostatic properties of biomatrices against common orthopaedic pathogens. Biochem. Biophys. Res. Commun. 2004, 321, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Pirnazar, P.; Wolinsky, L.; Nachnani, S.; Haake, S.; Pilloni, A.; Bernard, G.W. Bacteriostatic effects of hyaluronic acid. J. Periodontol. 1999, 70, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Deed, R.; Rooney, P.; Kumar, P.; Norton, J.D.; Smith, J.; Freemont, A.J.; Kumar, S. Early-response gene signalling is induced by angiogenic oligosaccharides of hyaluronan in endothelial cells. Inhibition by non-angiogenic, high-molecular-weight hyaluronan. Int. J. Cancer 1997, 71, 251–256. [Google Scholar] [CrossRef]
- de Brito Bezerra, B.; Mendes Brazao, M.A.; de Campos, M.L.; Casati, M.Z.; Sallum, E.A.; Sallum, A.W. Association of hyaluronic acid with a collagen scaffold may improve bone healing in critical-size bone defects. Clin. Oral Implant. Res. 2012, 23, 938–942. [Google Scholar] [CrossRef]
- Kawano, M.; Ariyoshi, W.; Iwanaga, K.; Okinaga, T.; Habu, M.; Yoshioka, I.; Tominaga, K.; Nishihara, T. Mechanism involved in enhancement of osteoblast differentiation by hyaluronic acid. Biochem. Biophys. Res. Commun. 2011, 405, 575–580. [Google Scholar] [CrossRef]
- Mendes, R.M.; Silva, G.A.; Lima, M.F.; Calliari, M.V.; Almeida, A.P.; Alves, J.B.; Ferreira, A.J. Sodium hyaluronate accelerates the healing process in tooth sockets of rats. Arch. Oral Biol. 2008, 53, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Asparuhova, M.B.; Chappuis, V.; Stahli, A.; Buser, D.; Sculean, A. Role of hyaluronan in regulating self-renewal and osteogenic differentiation of mesenchymal stromal cells and pre-osteoblasts. Clin. Oral Investig. 2020, 24, 3923–3937. [Google Scholar] [CrossRef]
- Zhai, P.; Peng, X.; Li, B.; Liu, Y.; Sun, H.; Li, X. The application of hyaluronic acid in bone regeneration. Int. J. Biol. Macromol. 2020, 151, 1224–1239. [Google Scholar] [CrossRef]
- Eliezer, M.; Sculean, A.; Miron, R.J.; Nemcovsky, C.; Weinberg, E.; Weinreb, M.; Zoabi, H.; Bosshardt, D.D.; Fujioka-Kobayashi, M.; Moses, O. Hyaluronic acid slows down collagen membrane degradation in uncontrolled diabetic rats. J. Periodontal. Res. 2019, 54, 644–652. [Google Scholar] [CrossRef]
- Fujioka-Kobayashi, M.; Muller, H.D.; Mueller, A.; Lussi, A.; Sculean, A.; Schmidlin, P.R.; Miron, R.J. In vitro effects of hyaluronic acid on human periodontal ligament cells. BMC Oral Health 2017, 17, 44. [Google Scholar] [CrossRef] [Green Version]
- Lodes, N.; Seidensticker, K.; Perniss, A.; Nietzer, S.; Oberwinkler, H.; May, T.; Walles, T.; Hebestreit, H.; Hackenberg, S.; Steinke, M. Investigation on Ciliary Functionality of Different Airway Epithelial Cell Lines in Three-Dimensional Cell Culture. Tissue Eng. Part A 2020, 26, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Mallick, S.P.; Rastogi, A.; Tripathi, S.; Srivastava, P. Strategies on process engineering of chondrocyte culture for cartilage tissue regeneration. Bioprocess Biosyst. Eng. 2017, 40, 601–610. [Google Scholar] [CrossRef]
- Seo, A.; Kitagawa, N.; Matsuura, T.; Sato, H.; Inai, T. Formation of keratinocyte multilayers on filters under airlifted or submerged culture conditions in medium containing calcium, ascorbic acid, and keratinocyte growth factor. Histochem. Cell Biol. 2016, 146, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Pischon, N.; Zimmermann, B.; Bernimoulin, J.P.; Hagewald, S. Effects of an enamel matrix derivative on human osteoblasts and PDL cells grown in organoid cultures. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2006, 102, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Hagewald, S.; Pischon, N.; Jawor, P.; Bernimoulin, J.P.; Zimmermann, B. Effects of enamel matrix derivative on proliferation and differentiation of primary osteoblasts. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2004, 98, 243–249. [Google Scholar] [CrossRef]
- Leboy, P.S.; Beresford, J.N.; Devlin, C.; Owen, M.E. Dexamethasone induction of osteoblast mRNAs in rat marrow stromal cell cultures. J. Cell. Physiol. 1991, 146, 370–378. [Google Scholar] [CrossRef]
- Smith, E.; Redman, R.A.; Logg, C.R.; Coetzee, G.A.; Kasahara, N.; Frenkel, B. Glucocorticoids inhibit developmental stage-specific osteoblast cell cycle. Dissociation of cyclin A-cyclin-dependent kinase 2 from E2F4-p130 complexes. J. Biol. Chem. 2000, 275, 19992–20001. [Google Scholar] [CrossRef]
- Boskey, A.L.; Roy, R. Cell culture systems for studies of bone and tooth mineralization. Chem. Rev. 2008, 108, 4716–4733. [Google Scholar] [CrossRef]
- Park, J.B. The effects of dexamethasone, ascorbic acid, and beta-glycerophosphate on osteoblastic differentiation by regulating estrogen receptor and osteopontin expression. J. Surg. Res. 2012, 173, 99–104. [Google Scholar] [CrossRef]
- Eliezer, M.; Sculean, A.; Miron, R.J.; Nemcovsky, C.; Bosshardt, D.D.; Fujioka-Kobayashi, M.; Weinreb, M.; Moses, O. Cross-linked hyaluronic acid slows down collagen membrane resorption in diabetic rats through reducing the number of macrophages. Clin. Oral Investig. 2022, 26, 2401–2411. [Google Scholar] [CrossRef]
- Toloue, S.M.; Chesnoiu-Matei, I.; Blanchard, S.B. A clinical and histomorphometric study of calcium sulfate compared with freeze-dried bone allograft for alveolar ridge preservation. J. Periodontol. 2012, 83, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Klinger, A.; Asad, R.; Shapira, L.; Zubery, Y. In vivo degradation of collagen barrier membranes exposed to the oral cavity. Clin. Oral Implant. Res. 2010, 21, 873–876. [Google Scholar]
- Boyan, B.D.; Bonewald, L.F.; Paschalis, E.P.; Lohmann, C.H.; Rosser, J.; Cochran, D.L.; Dean, D.D.; Schwartz, Z.; Boskey, A.L. Osteoblast-mediated mineral deposition in culture is dependent on surface microtopography. Calcif. Tissue Int. 2002, 71, 519–529. [Google Scholar] [CrossRef]
- Masaki, C.; Schneider, G.B.; Zaharias, R.; Seabold, D.; Stanford, C. Effects of implant surface microtopography on osteoblast gene expression. Clin. Oral Implant. Res. 2005, 16, 650–656. [Google Scholar] [CrossRef]
- Papaioannou, K.A.; Markopoulou, C.E.; Gioni, V.; Mamalis, A.A.; Vayouraki, H.N.; Kletsas, D.; Vrotsos, I.A. Attachment and proliferation of human osteoblast-like cells on guided bone regeneration (GBR) membranes in the absence or presence of nicotine: An in vitro study. Int. J. Oral Maxillofac. Implant. 2011, 26, 509–519. [Google Scholar]
- Takata, T.; Wang, H.L.; Miyauchi, M. Migration of osteoblastic cells on various guided bone regeneration membranes. Clin. Oral Implant. Res. 2001, 12, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Friedmann, A.; Fickl, S.; Fischer, K.R.; Dalloul, M.; Goetz, W.; Kauffmann, F. Horizontal Augmentation of Chronic Mandibular Defects by the Guided Bone Regeneration Approach: A Randomized Study in Dogs. Materials 2021, 15, 238. [Google Scholar] [CrossRef]
- Zhao, N.; Wang, X.; Qin, L.; Zhai, M.; Yuan, J.; Chen, J.; Li, D. Effect of hyaluronic acid in bone formation and its applications in dentistry. J. Biomed. Mater. Res. A 2016, 104, 1560–1569. [Google Scholar] [CrossRef]
- Asparuhova, M.B.; Kiryak, D.; Eliezer, M.; Mihov, D.; Sculean, A. Activity of two hyaluronan preparations on primary human oral fibroblasts. J. Periodontal. Res. 2018, 54, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Kyyak, S.; Pabst, A.; Heimes, D.; Kammerer, P.W. The Influence of Hyaluronic Acid Biofunctionalization of a Bovine Bone Substitute on Osteoblast Activity In Vitro. Materials 2021, 14, 2885. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer | Sequence (5′ to 3′) |
|---|---|---|
| Cx43 | forward | CCT TCT TGC TGA TCC AGT GGT AC |
| reverse | ACC AAG GAC ACC AVV AGC AT | |
| BGLAP | forward | TTC TTT CCT CTT CCC CTT G |
| reverse | CCT CTT CTG GAG TTT ATT TGG | |
| IBSP | forward | GGA GAC TTC AAA TGA AGG AG |
| reverse | CAG AAA GTG TGG TAT TCT CAG | |
| Runx2 | forward | CCA ACC CAC GAA TGC ACT ATC |
| reverse | TAG TGA GTG GTG GCG GAC ATA C | |
| GAPDH | forward | TGC ACC ACC AAC TGC TTA GC |
| reverse | GGC ATG GAC TGT GGT CAT GAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nobis, B.; Ostermann, T.; Weiler, J.; Dittmar, T.; Friedmann, A. Impact of Cross-Linked Hyaluronic Acid on Osteogenic Differentiation of SAOS-2 Cells in an Air-Lift Model. Materials 2022, 15, 6528. https://0-doi-org.brum.beds.ac.uk/10.3390/ma15196528
Nobis B, Ostermann T, Weiler J, Dittmar T, Friedmann A. Impact of Cross-Linked Hyaluronic Acid on Osteogenic Differentiation of SAOS-2 Cells in an Air-Lift Model. Materials. 2022; 15(19):6528. https://0-doi-org.brum.beds.ac.uk/10.3390/ma15196528
Chicago/Turabian StyleNobis, Bianca, Thomas Ostermann, Julian Weiler, Thomas Dittmar, and Anton Friedmann. 2022. "Impact of Cross-Linked Hyaluronic Acid on Osteogenic Differentiation of SAOS-2 Cells in an Air-Lift Model" Materials 15, no. 19: 6528. https://0-doi-org.brum.beds.ac.uk/10.3390/ma15196528