
Molecular Characterization, Pathogenicity and Biological Characterization of Colletotrichum Species Associated with Anthracnose of Camellia yuhsienensis Hu in China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Plant Material
2.2. Molecular Characterization
2.3. Morphological Characterization
2.4. Koch’s Postulates Verification
2.5. Pathogenicity Tests
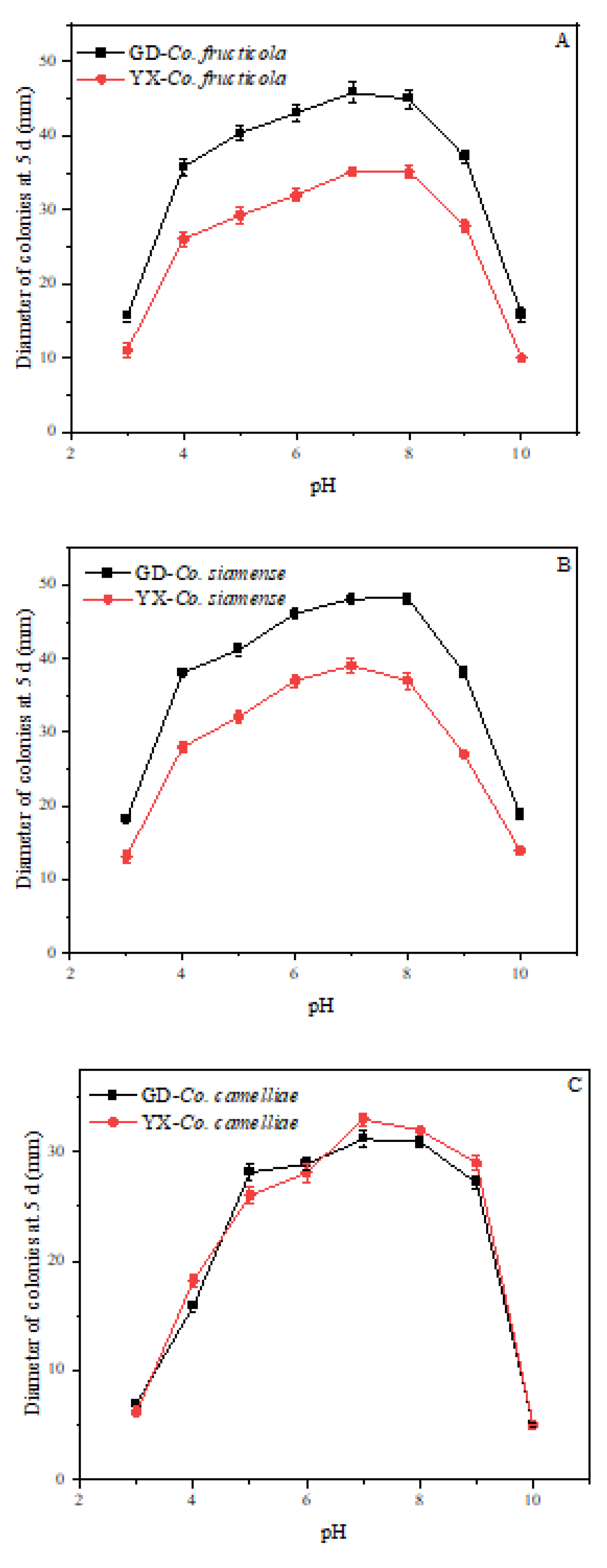
2.6. Effect of Temperature and pH on Mycelial Growth
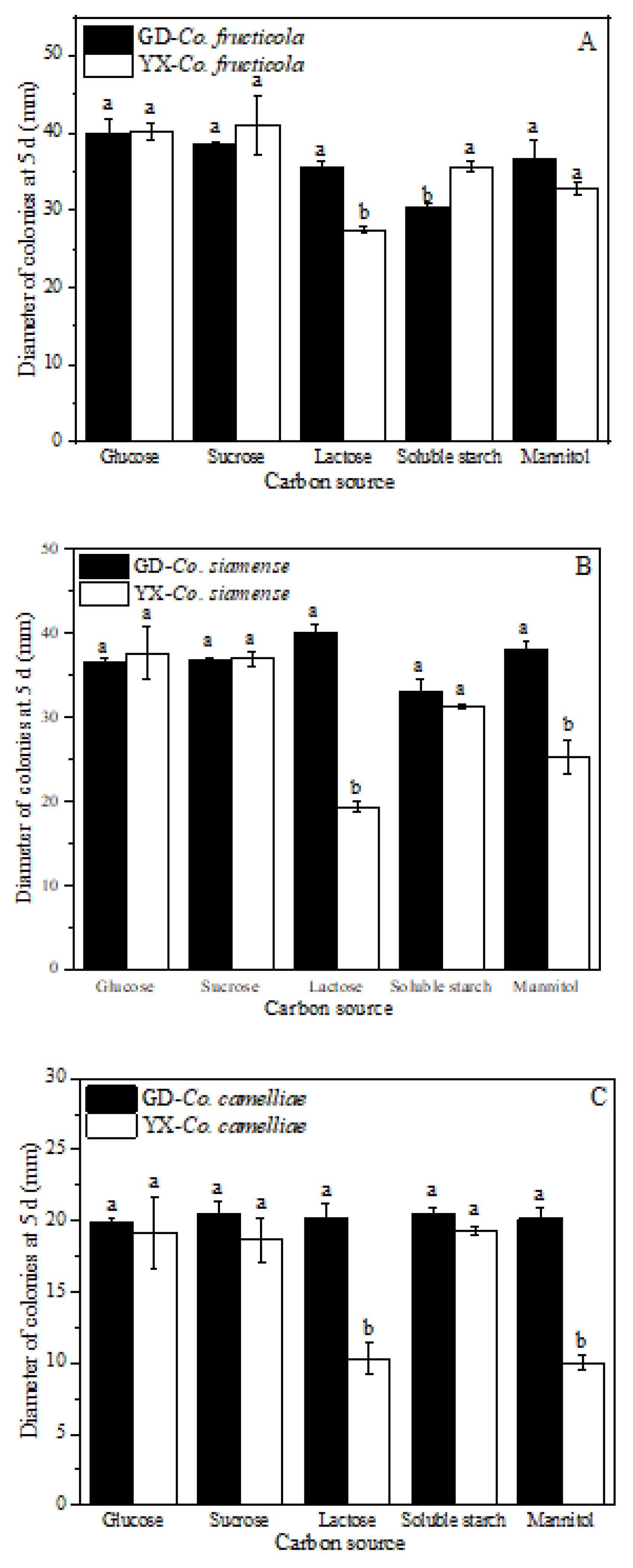
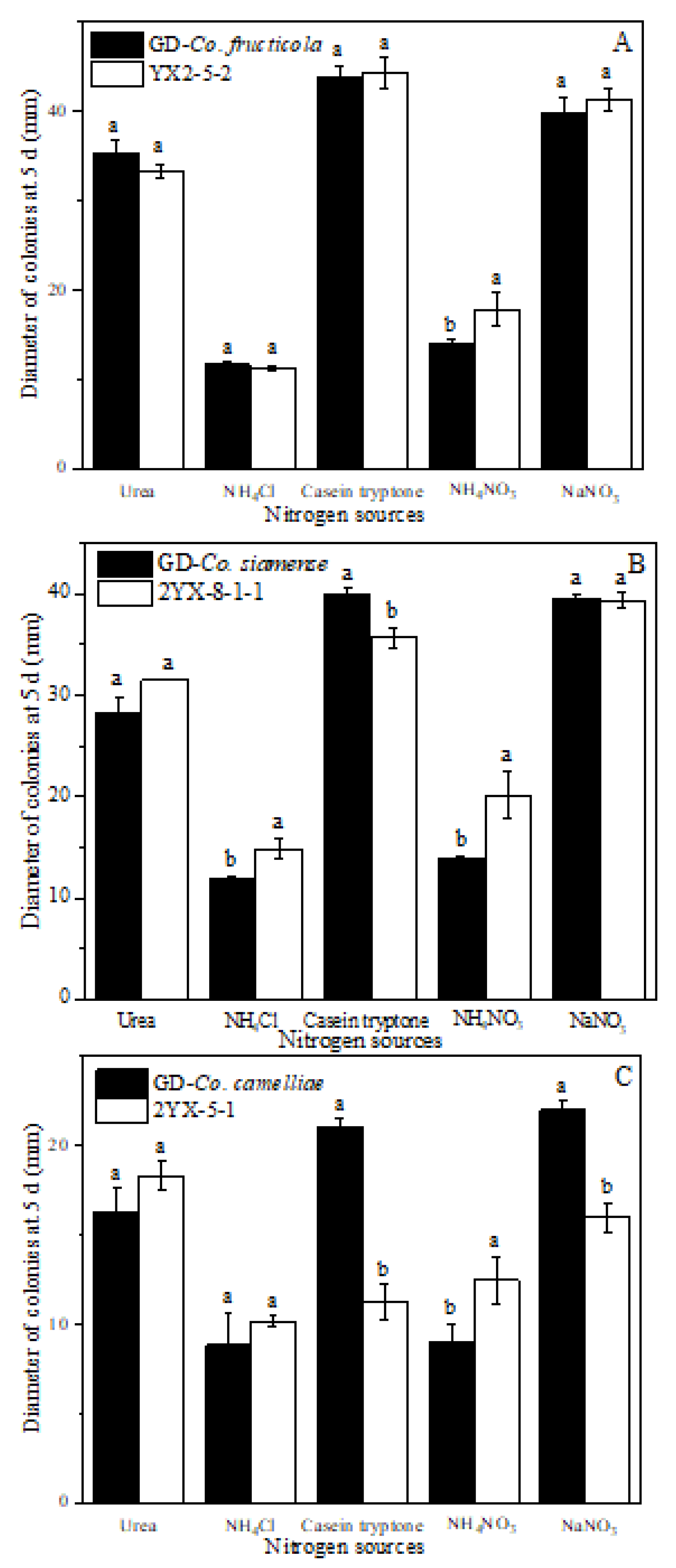
2.7. Effect of Carbon and Nitrogen Sources on Mycelial Growth
3. Results and Discussion
3.1. Symptom Characteristics
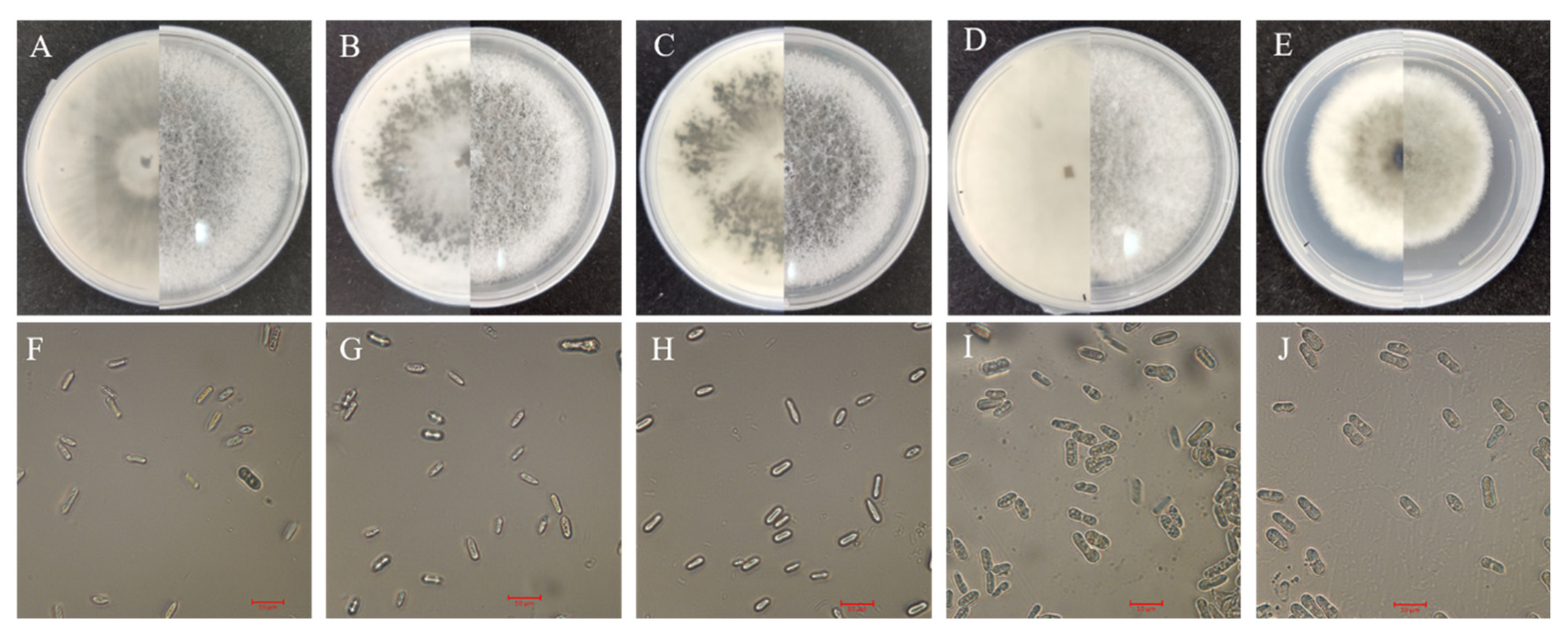
3.2. Cultural and Morphological Characteristics
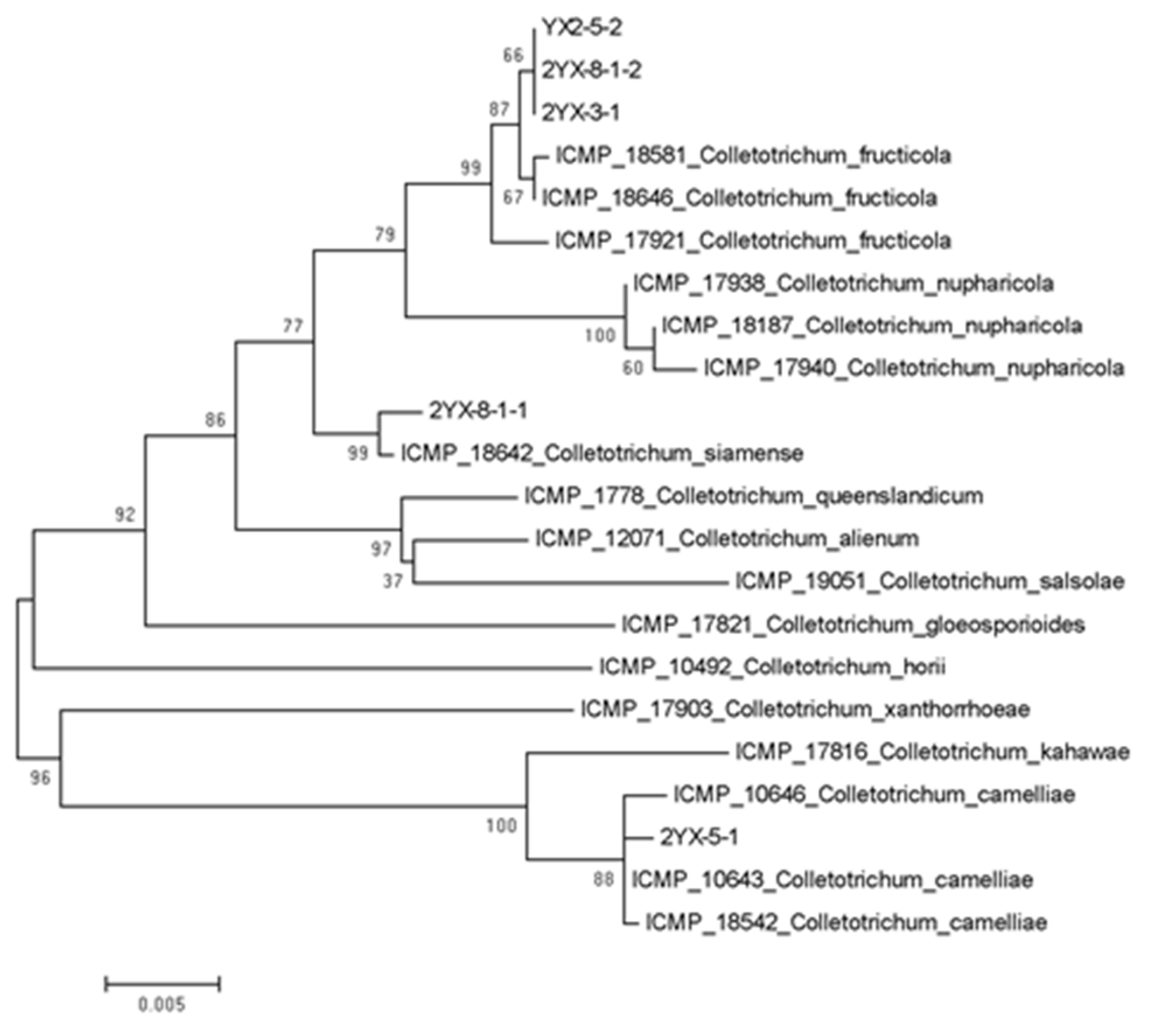
3.3. Phylogenetic Analysis
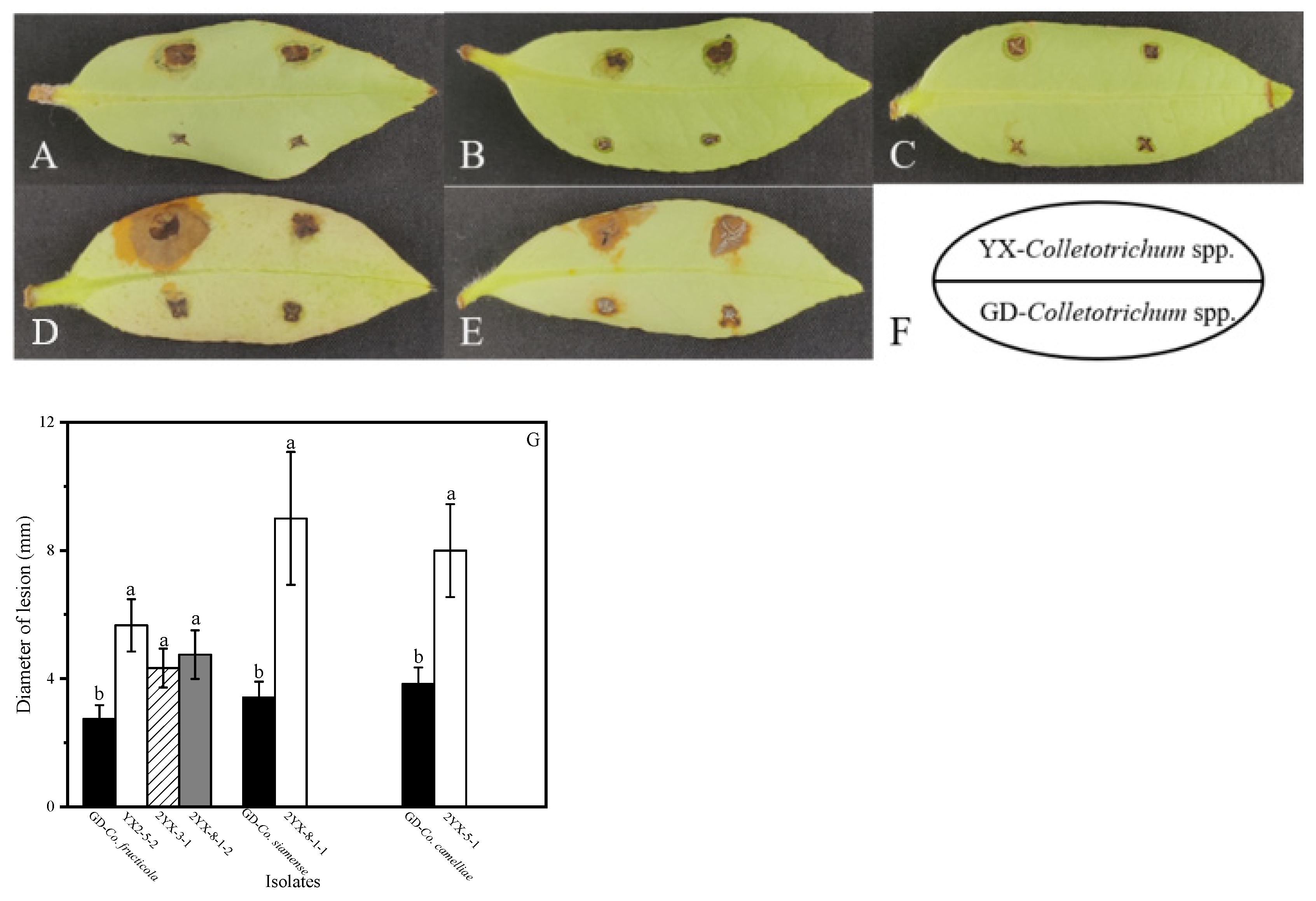
3.4. Pathogenicity Tests
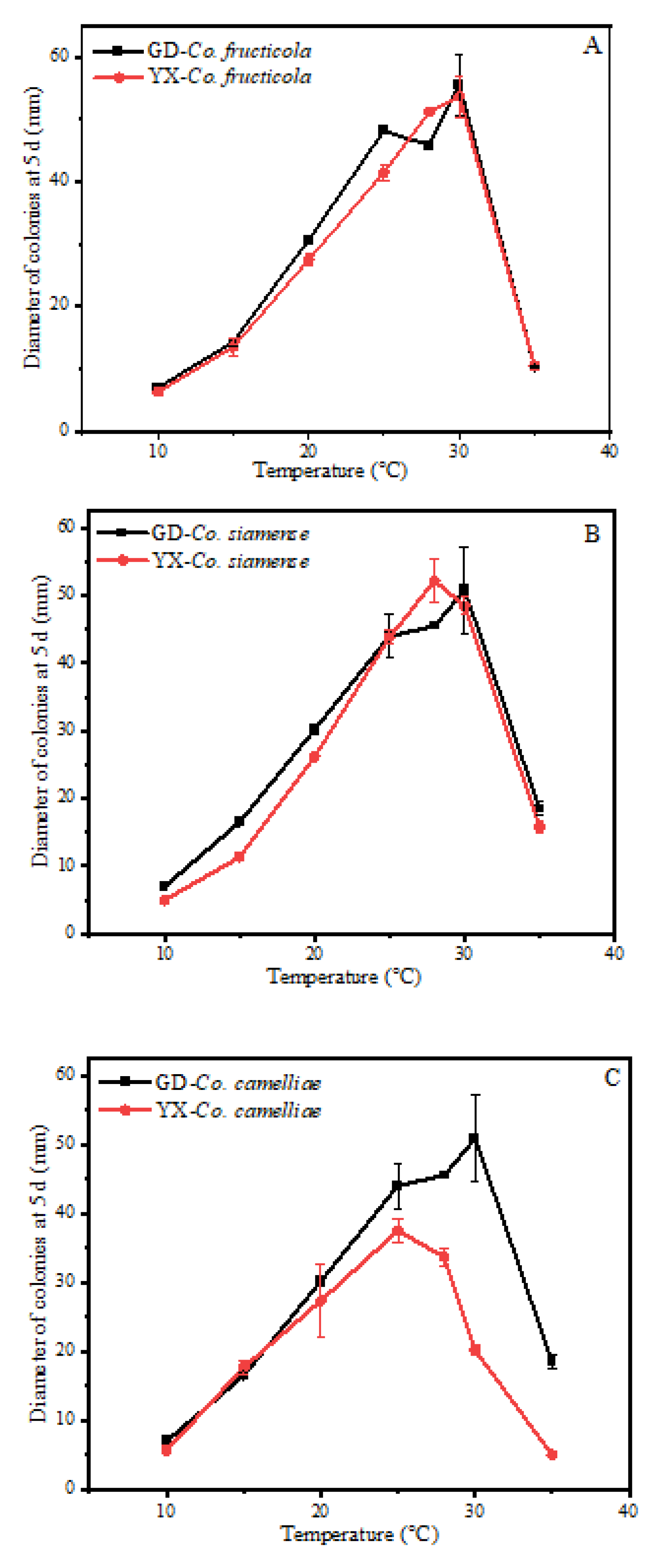
3.5. Effect of Temperature and pH on Mycelial Growth
3.6. Effect of Carbon and Nitrogen Sources on Mycelial Growth
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, Y.Z. Oil Tea Camellia Superior Germplasm Resources; China Forestry Publishing House: Beijing, China, 2008. [Google Scholar]
- Qin, S.; Rong, J.; Zhang, W.; Chen, J. Cultivation history of Camellia oleifera and genetic resources in the Yangtze River Basin. Biodivers. Sci. 2018, 26, 384–395. [Google Scholar] [CrossRef]
- Li, Y.; Ye, T.; Han, C.; Ye, Z.; Zhang, J.; Xiao, S.; Yuan, D. Cytogenetic analysis of interspecific hybridization in oil-tea (Camellia oleifera). Euphytica 2021, 217, 28. [Google Scholar] [CrossRef]
- Shi, T.; Wu, G.; Jin, Q.; Wang, X. Camellia oil authentication: A comparative analysis and recent analytical techniques developed for its assessment. A review. Trends Food Sci. Technol. 2020, 97, 88–99. [Google Scholar] [CrossRef]
- Yang, C.; Liu, X.; Chen, Z.; Lin, Y.; Wang, S. Comparison of Oil Content and Fatty Acid Profile of Ten New Camellia oleifera Cultivars. J. Lipids 2016, 2016, 3982486. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Liang, K.; Wang, A.; Zhang, M.; Qiu, J.; Zhang, L. Physiological Characterization and Transcriptome Analysis of Camellia oleifera Abel. during Leaf Senescence. Forests 2020, 11, 812. [Google Scholar] [CrossRef]
- Chen, Y.; Deng, S.; Chen, L.; Li, M.; He, H.; Wang, X.; Peng, S.; Liu, C.; Wang, R.; Xu, Y.; et al. A new view on the development of oil tea camellia industry. J. Nanjing For. Univ. Nat. Sci. Edit 2020, 44, 1–10. [Google Scholar] [CrossRef]
- Wang, J.C. The total output value of Camellia oleifera Industry in China reached 116 billion yuan. China Food Newspaper, 20 November 2020; 2. [Google Scholar]
- Li, S.Z.; Zhang, S.P.; Li, H. A HOPS protein, CfVps39, is required for appressorium formation, environmental stress response and vacuolar fusion of Colletotrichum fructicola. For. Pathol. 2021, 51, e12692. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, J.; Jiang, S.; Li, H.; Zhou, G. Colletotrichum fructicola STE50 is required for vegetative growth, asexual reproduction, appressorium formation, pathogenicity and the response to external stress. J. Plant. Pathol. 2020, 102, 335–342. [Google Scholar] [CrossRef]
- Jin, A.X.; Zhou, G.Y.; Li, H. Progress, problem and prospect of oil camelliae anthracnose (Colletotrichum gloeosporioides) research. For. Pest. Dis. 2009, 28, 27–31. [Google Scholar]
- Zhu, Y.; Liao, W.; Zou, D.; Wu, Y.; Yan, D. Identification and biological characteristics of the pathogen from Camellia oleifera anthracnose in Guangxi. J. Plant. Prot. 2015, 42, 382–389. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Hao, X.-Y.; Wang, L.; Bin, X.; Wang, X.-C.; Yang, Y.-J. Diverse Colletotrichum species cause anthracnose of tea plants (Camellia sinensis (L.) O. Kuntze) in China. Sci. Rep. 2016, 6, 35287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mongkolporn, O.; Taylor, P.W.J. Chili anthracnose: Colletotrichum taxonomy and pathogenicity. Plant. Pathol. 2018, 67, 1255–1263. [Google Scholar] [CrossRef]
- Hyde, K.D.; Cai, L.; Cannon, P.F.; Crouch, J.A.; Crous, P.W.; Damm, U.; Goodwin, P.H.; Chen, H.; Johnston, P.R.; Jones, E.B.G.; et al. Colletotrichum—Names in current use. Fungal Divers. 2009, 39, 147–182. [Google Scholar]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant. Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhu, D.; Xu, J.; Zhou, G.; Hu, M.; Tian, F. Population genetic structure of Colletotrichum gloeosporioides causing anthracnose of Camellia oleifera in China. Acta Phytopathol. Sin. 2014, 44, 620–628. [Google Scholar] [CrossRef]
- Li, H.; Zhou, G.-Y.; Liu, J.-A.; Xu, J. Population Genetic Analyses of the Fungal Pathogen Colletotrichum fructicola on Tea-Oil Trees in China. PLoS ONE 2016, 11, e0156841. [Google Scholar] [CrossRef] [Green Version]
- Fu, M.; Crous, P.W.; Bai, Q.; Zhang, P.F.; Xiang, J.; Guo, Y.S.; Zhao, F.F.; Yang, M.M.; Hong, N.; Xu, W.X.; et al. Colletotrichum species associated with anthracnose of Pyrus spp. in China. Persoonia 2019, 42, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Wang, Y.; Li, N.; Ni, D.; Yang, Y.; Wang, X. Differences in the Characteristics and Pathogenicity of Colletotrichum camelliae and C. fructicola Isolated From the Tea Plant [Camellia sinensis (L.) O. Kuntze]. Front. Microbiol. 2018, 9, 3060. [Google Scholar] [CrossRef]
- Holtappels, D.; Fortuna, K.; Lavigne, R.; Wagemans, J. The future of phage biocontrol in integrated plant protection for sustainable crop production. Curr. Opin. Biotechnol. 2021, 68, 60–71. [Google Scholar] [CrossRef]
- Savchenko, I.V. Breeding New Varieties and Hybrids of Agricultural Plants. Her. Russ. Acad. Sci. 2017, 87, 104–110. [Google Scholar] [CrossRef]
- Nie, Z.; Huang, X.; Hu, Z.; Li, X.; Yin, H.; Li, J. Characterization of the complete chloroplast genome of Camellia yuhsienensis Hu, a resilient shrub with strong floral fragrance. Mitochondrial DNA B 2020, 5, 3016–3017. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Yao, X.; Ren, H.; Wang, K. Determination of fatty acid composition and metallic element content of four Camellia species used for edible oil extraction in China. J. Consum Prot. Food Saf. 2017, 12, 165–169. [Google Scholar] [CrossRef]
- Denton-Giles, M.; Bradshaw, R.E.; Dijkwel, P.P. Ciborinia camelliae (Sclerotiniaceae) Induces Variable Plant Resistance Responses in Selected Species of Camellia. Phytopathology 2013, 103, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Denton-Giles, M. Characterization of Incompatible and Compatible Camellia-Ciborinia Camelliae Plant-Pathogen Interactions. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 2014. [Google Scholar]
- Yang, G.; Shu, Q.; Duan, L.; Chen, C.; Zheng, H. Resistance of main cultivars of oil tea to Colletotrichum gloeosporioides. J. Anhui Agric. Univ. 2004, 31, 480–483. [Google Scholar] [CrossRef]
- Duan, L.; Yang, G.; Shu, Q.; Hongbing, Z. Relationship of Peel Color with Resistance to Anthracnose in Oiltea Camellia. Nonwood For. Res. 2005, 23, 9–12+20. [Google Scholar] [CrossRef]
- Li, J.; Luo, Z.; Zhang, C.; Qu, X.; Chen, M.; Song, T.; Yuan, J. Seasonal Variation in the Rhizosphere and Non-Rhizosphere Microbial Community Structures and Functions of Camellia yuhsienensis Hu. Microorganisms 2020, 8, 1385. [Google Scholar] [CrossRef]
- Saracchi, M.; Locati, D.; Colombo, E.M.; Pasquali, M. Updates on Ciborinia camelliae, the causal agent of camellia flower blight. J. Plant. Pathol. 2019, 101, 215–223. [Google Scholar] [CrossRef]
- Weir, B.; Johnston, P.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Weir, B.S.; Damm, U.; Crous, P.W.; Wang, Y.; Liu, B.; Wang, M.; Zhang, M.; Cai, L. Unravelling Colletotrichum species associated with Camellia: Employing ApMat and GS loci to resolve species in the C. gloeosporioides complex. Persoonia 2015, 35, 63–86. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; Hyde, K.D.; Taylor, P.W.J.; Weir, B.S.; Waller, J.M.; Abang, M.M.; Zhang, J.Z.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y.; et al. A polyphasic approach for studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Piesik, D.; Rochat, D.; Delaney, K.J.; Marion-Poll, F. Orientation of European corn borer first instar larvae to synthetic green leaf volatiles. J. Appl. Entomol. 2013, 137, 234–240. [Google Scholar] [CrossRef] [Green Version]
- Skoczek, A.; Piesik, D.; Wenda-Piesik, A.; Buszewski, B.; Bocianowski, J.; Wawrzyniak, M. Volatile organic compounds released by maize following herbivory or insect extract application and communication between plants. J. Appl. Entomol. 2017, 141, 630–643. [Google Scholar] [CrossRef]
- Piesik, D.; Rochat, D.; van der Pers, J.; Marion-Poll, F. Pulsed Odors from Maize or Spinach Elicit Orientation in European Corn Borer Neonate Larvae. J. Chem. Ecol. 2009, 35, 1032–1042. [Google Scholar] [CrossRef]
- De Silva, D.D.; Ades, P.K.; Taylor, P.W.J. Pathogenicity of Colletotrichum species causing anthracnose of Capsicum in Asia. Plant. Pathol. 2021, 70, 875–884. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.-Y.; Xu, X.; Cheng, J.; Zheng, L.; Huang, J.; Li, D.-W. Identification and Characterization of Colletotrichum Species Associated with Anthracnose Disease of Camellia oleifera in China. Plant. Dis. 2020, 104, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Moral, J.; Agustí-Brisach, C.; Raya, M.C.; Jurado-Bello, J.; López-Moral, A.; Roca, L.F.; Chattaoui, M.; Rhouma, A.; Nigro, F.; Sergeeva, V.; et al. Diversity of Colletotrichum Species Associated with Olive Anthracnose Worldwide. J. Fungi 2021, 7, 741. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.J.; Kim, W.G.; Kim, H.G.; Choi, H.W.; Lee, Y.K.; Lee, B.D.; Lee, S.Y.; Hong, S.K. Morphology, Molecular Phylogeny and Pathogenicity of Colletotrichum panacicola Causing Anthracnose of Korean Ginseng. Plant. Pathol. J. 2011, 27, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Liu, C.; Liu, J.A.; Zhou, G.Y. First Report of Colletotrichum fructicola Causing Anthracnose on Camellia yuhsienensis Hu in China. Plant. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Pinnaka, A.K.; Shenoy, B.D. Resolving the Colletotrichum siamense species complex using ApMat marker. Fungal Divers. 2015, 71, 247–264. [Google Scholar] [CrossRef]
- Han, Y.C.; Zeng, X.G.; Xiang, F.Y.; Ren, L.; Chen, F.Y.; Gu, Y.C. Distribution and Characteristics of Colletotrichum spp. Associated with Anthracnose of Strawberry in Hubei, China. Plant. Dis. 2016, 100, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Ye, G. A Study on the Peroxidase in Camellia Species and Their Resistance to Anthracnoso Causing Fungus. J. Fujian Coll. For. 1990, 10, 368–375. [Google Scholar] [CrossRef]
- Wang, Y.; Hao, X.; Lu, Q.; Wang, L.; Qian, W.; Li, N.; Ding, C.; Wang, X.; Yang, Y. Transcriptional analysis and histochemistry reveal that hypersensitive cell death and H2O2 have crucial roles in the resistance of tea plant (Camellia sinensis (L.) O. Kuntze) to anthracnose. Hort. Res. 2018, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Lima, N.B.; Lima, W.G.; Tovar-Pedraza, J.M.; Michereff, S.J.; Camara, M.P.S. Comparative epidemiology of Colletotrichum species from mango in northeastern Brazil. Eur. J. Plant. Pathol. 2015, 141, 679–688. [Google Scholar] [CrossRef]
- Xue, L.; Zhang, L.; Yang, X.X.; Huang, X.; Wu, W.; Zhou, X.; White, J.F.; Liu, Y.; Li, C. Characterization, Phylogenetic Analyses, and Pathogenicity of Colletotrichum Species on Morus alba in Sichuan Province, China. Plant. Dis. 2019, 103, 2624–2633. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Tan, G.; Li, Z. Studies on the Biological Characteristics of Collectotrichum gloeosporiodes Strain Related to Virulence and Hypovirulence on Apple Anthracnose. Acta Laser Biol. Sin. 2008, 17, 64–69. [Google Scholar]
- He, Y.Y.; Chen, Q.G.; Shu, C.W.; Yang, M.; Zhou, E.X. Colletotrichum truncatum, a new cause of anthracnose on Chinese flowering cabbage (Brassica parachinensis) in China. Trop. Plant. Pathol. 2016, 41, 183–192. [Google Scholar] [CrossRef]
- Miyara, I.; Shafran, H.; Davidzon, M.; Sherman, A.; Prusky, D. pH Regulation of Ammonia Secretion by Colletotrichum gloeosporioides and Its Effect on Appressorium Formation and Pathogenicity. Mol. Plant.-Microbe Interact. 2010, 23, 304–316. [Google Scholar] [CrossRef] [Green Version]
- Alkan, N.; Meng, X.; Friedlander, G.; Reuveni, E.; Sukno, S.; Sherman, A.; Thon, M.; Fluhr, R.; Prusky, D. Global Aspects of pacC Regulation of Pathogenicity Genes in Colletotrichum gloeosporioides as Revealed by Transcriptome Analysis. Mol. Plant.-Microbe Interact. 2013, 26, 1345–1358. [Google Scholar] [CrossRef] [Green Version]
- Tardi-Ovadia, R.; Linker, R.; Tsror, L. Direct Estimation of Local pH Change at Infection Sites of Fungi in Potato Tubers. Phytopathology 2017, 107, 132–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Costa, D.M.; Chandima, A.A.G. Effect of exogenous pH on development and growth of Colletotrichum musae and development of anthracnose in different banana cultivars in Sri Lanka. J. Natl. Sci. Found. Sri Lanka 2014, 42, 203–214. [Google Scholar] [CrossRef]
- Prusky, D.; McEvoy, J.L.; Leverentz, B.; Conway, W.S. Local Modulation of Host pH by Colletotrichum Species as a Mechanism to Increase Virulence. Mol. Plant.-Microbe Interact. 2001, 14, 1105–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Conidial Size (μm) | Mycelial Growth (mm/d) |
|---|---|---|
| YX2-5-2 | 7.27 ± 0.52 × 1.81 ± 0.31 | 13.9 ± 0.6 |
| 2YX-8-1-2 | 7.27 ± 0.48 × 2.42 ± 0.27 | 13.7 ± 0.3 |
| 2YX-3-1 | 7.27 ± 0.55 × 1.81 ± 0.21 | 13.1 ± 0.8 |
| 2YX-8-1-1 | 9.09 ± 0.45 × 3.64 ± 0.43 | 13.4 ± 0.3 |
| 2YX-5-1 | 9.09 ± 0.36× 3.64 ± 0.39 | 8.5 ± 0.9 |
| Isolate | Gene Name | Genbank Accession Number |
|---|---|---|
| YX2-5-2 | GAPDH | MW398864 |
| YX2-5-2 | ACT | MW398863 |
| YX2-5-2 | CHSI | MW886232 |
| YX2-5-2 | SOD2 | MW398866 |
| YX2-5-2 | ITS | MW398865 |
| 2YX-8-1-2 | GAPDH | MZ224482 |
| 2YX-8-1-2 | ACT | MZ224483 |
| 2YX-8-1-2 | CHSI | OL310498 |
| 2YX-8-1-2 | SOD2 | MZ224480 |
| 2YX-8-1-2 | ITS | MZ224481 |
| 2YX-8-1-1 | GAPDH | MW398861 |
| 2YX-8-1-1 | ACT | MW398860 |
| 2YX-8-1-1 | CHSI | OL310500 |
| 2YX-8-1-1 | SOD2 | MZ048745.1 |
| 2YX-8-1-1 | ITS | MW398862 |
| 2YX-5-1 | GAPDH | MZ048746 |
| 2YX-5-1 | ACT | MW924872 |
| 2YX-5-1 | CHSI | MW924874 |
| 2YX-5-1 | SOD2 | MW924873 |
| 2YX-5-1 | ITS | MW911446 |
| 2YX-3-1 | GAPDH | MW924878 |
| 2YX-3-1 | ACT | MW924879 |
| 2YX-3-1 | CHSI | OL310499 |
| 2YX-3-1 | SOD2 | MW924877 |
| 2YX-3-1 | ITS | MW924880 |
| Isolates | Optimal Temperatures (°C) | |
|---|---|---|
| YX-Colletotrichum spp. | YX-Co. fructicola | 30 |
| YX-Co. siamense | 28 | |
| YX-Co. camelliae | 25 | |
| GD-Colletotrichum spp. | GD-Co. fructicola | 30 |
| GD-Co. siamense | 30 | |
| GD-Co. camelliae | 30 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Jiang, L.; Bao, A.; Liu, C.; Liu, J.; Zhou, G. Molecular Characterization, Pathogenicity and Biological Characterization of Colletotrichum Species Associated with Anthracnose of Camellia yuhsienensis Hu in China. Forests 2021, 12, 1712. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121712
Chen X, Jiang L, Bao A, Liu C, Liu J, Zhou G. Molecular Characterization, Pathogenicity and Biological Characterization of Colletotrichum Species Associated with Anthracnose of Camellia yuhsienensis Hu in China. Forests. 2021; 12(12):1712. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121712
Chicago/Turabian StyleChen, Xinggang, Lingyu Jiang, Anhua Bao, Changlin Liu, Junang Liu, and Guoying Zhou. 2021. "Molecular Characterization, Pathogenicity and Biological Characterization of Colletotrichum Species Associated with Anthracnose of Camellia yuhsienensis Hu in China" Forests 12, no. 12: 1712. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121712