
Metabolic Pathways Involved in the Drought Stress Response of Nitraria tangutorum as Revealed by Transcriptome Analysis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Complementary DNA Library Construction and RNA Sequencing
2.3. Enrichment Analysis of Differentially Expressed Genes
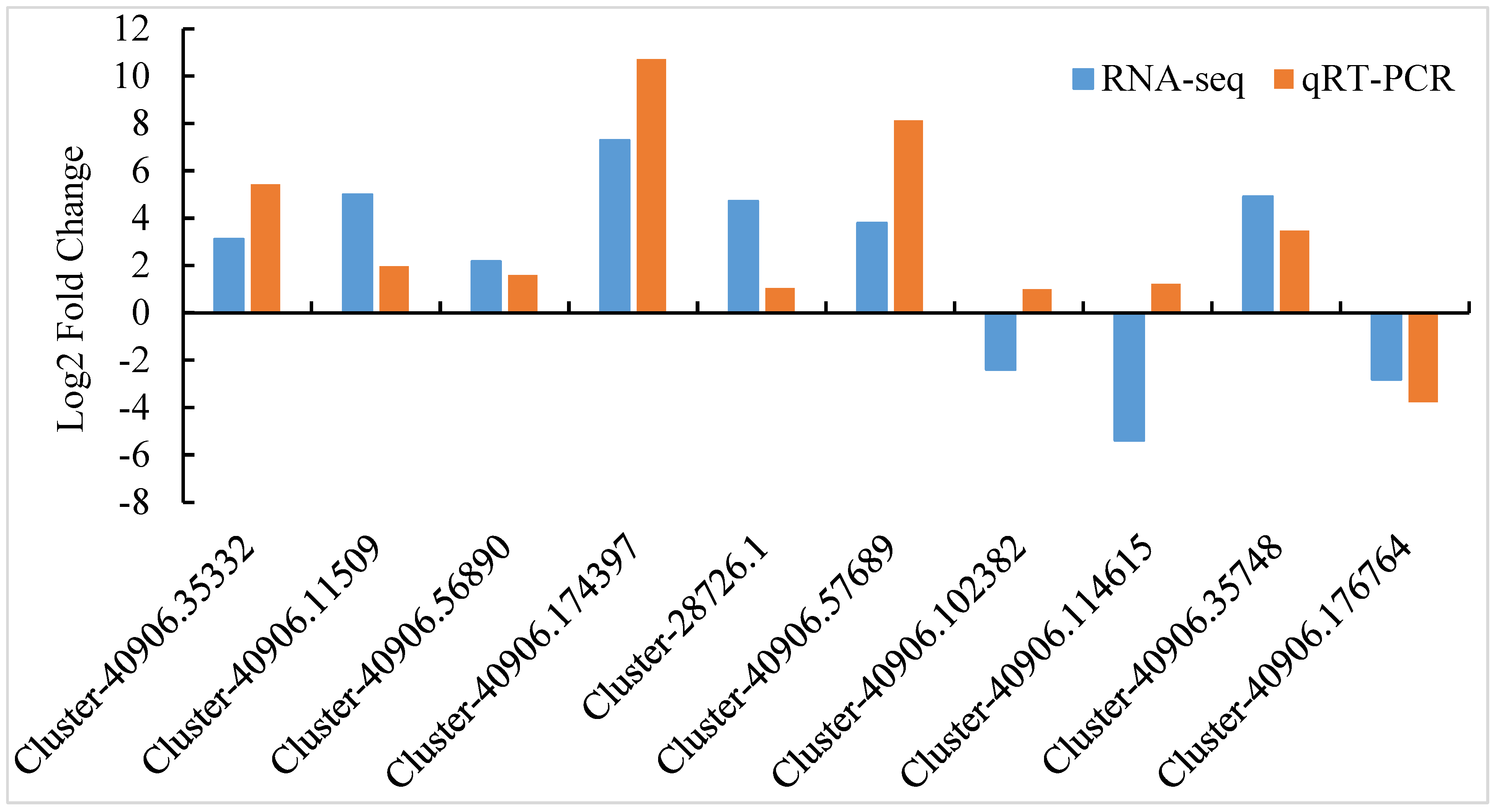
2.4. Verification of RNA-Seq Data by Quantitative Real-Time PCR
3. Results
3.1. RNA-Seq Analysis and Transcript Splicing
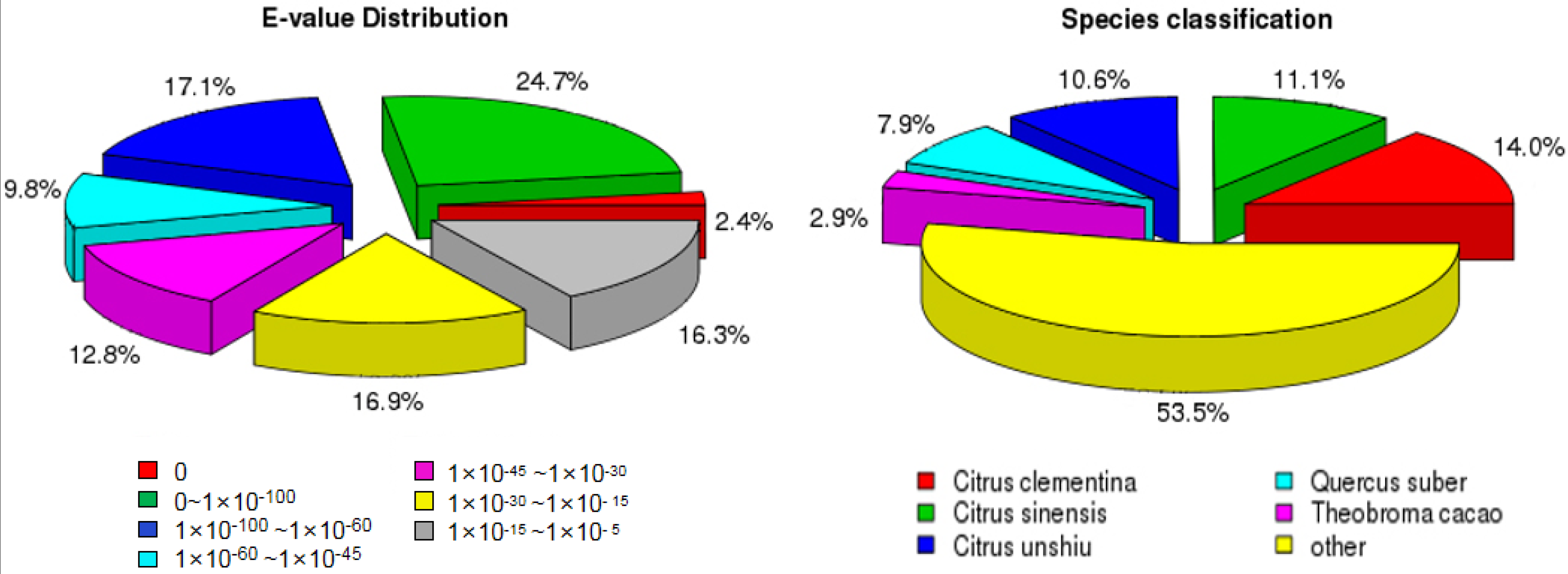
3.2. Gene Function Annotation
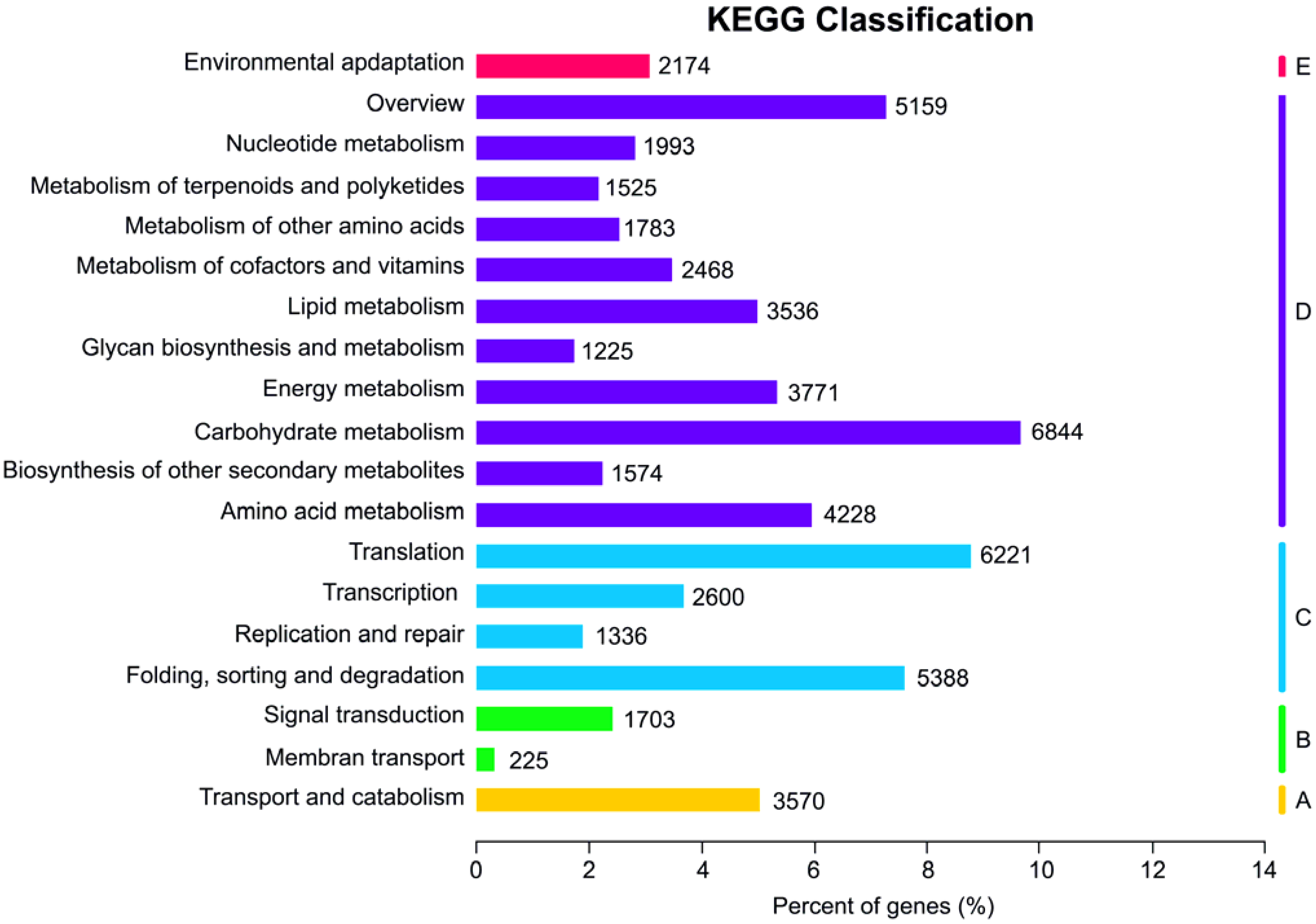
3.3. KEGG Functional Classification
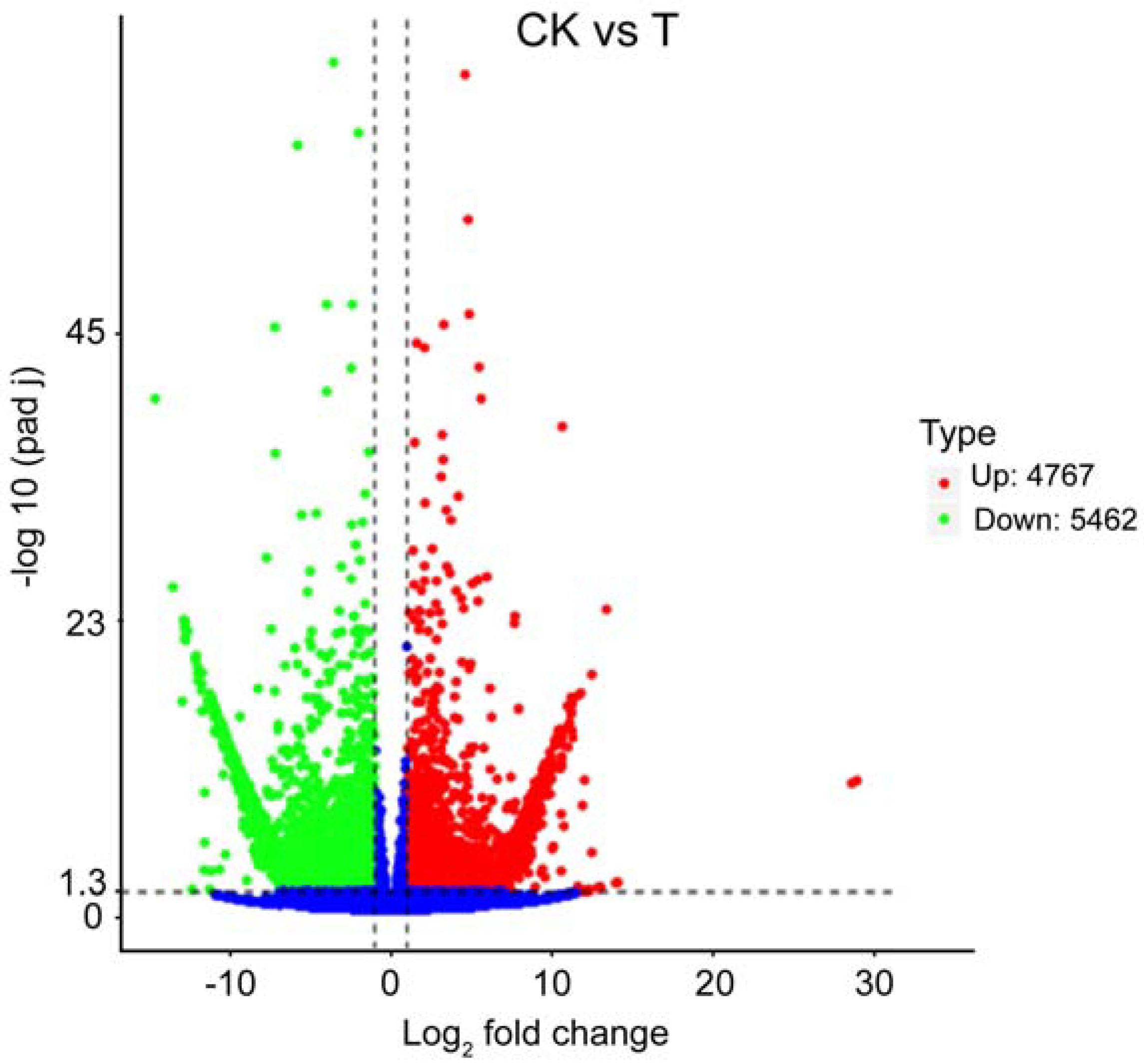
3.4. DEGs Analysis
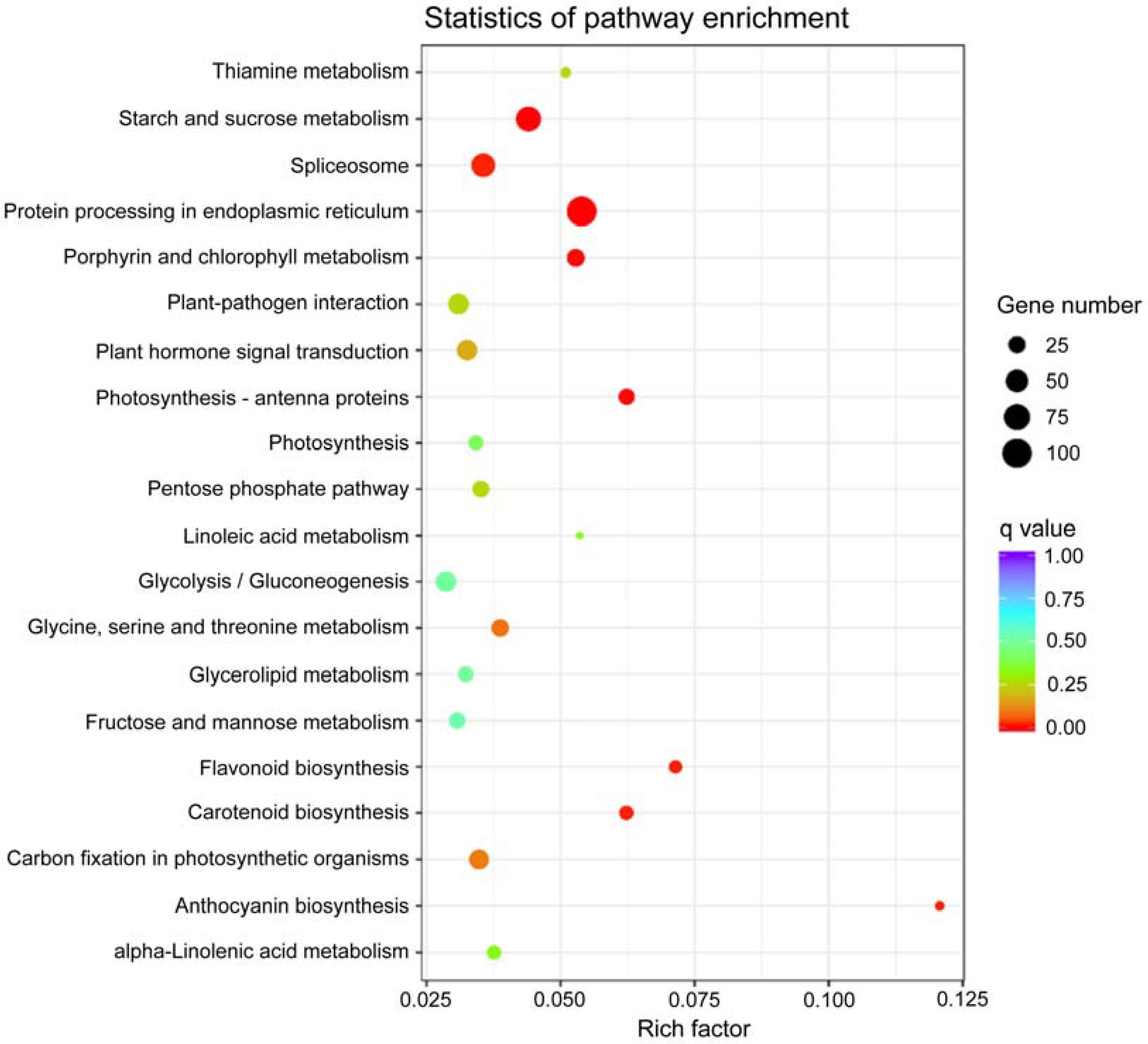
3.5. KEGG Enrichment Analysis
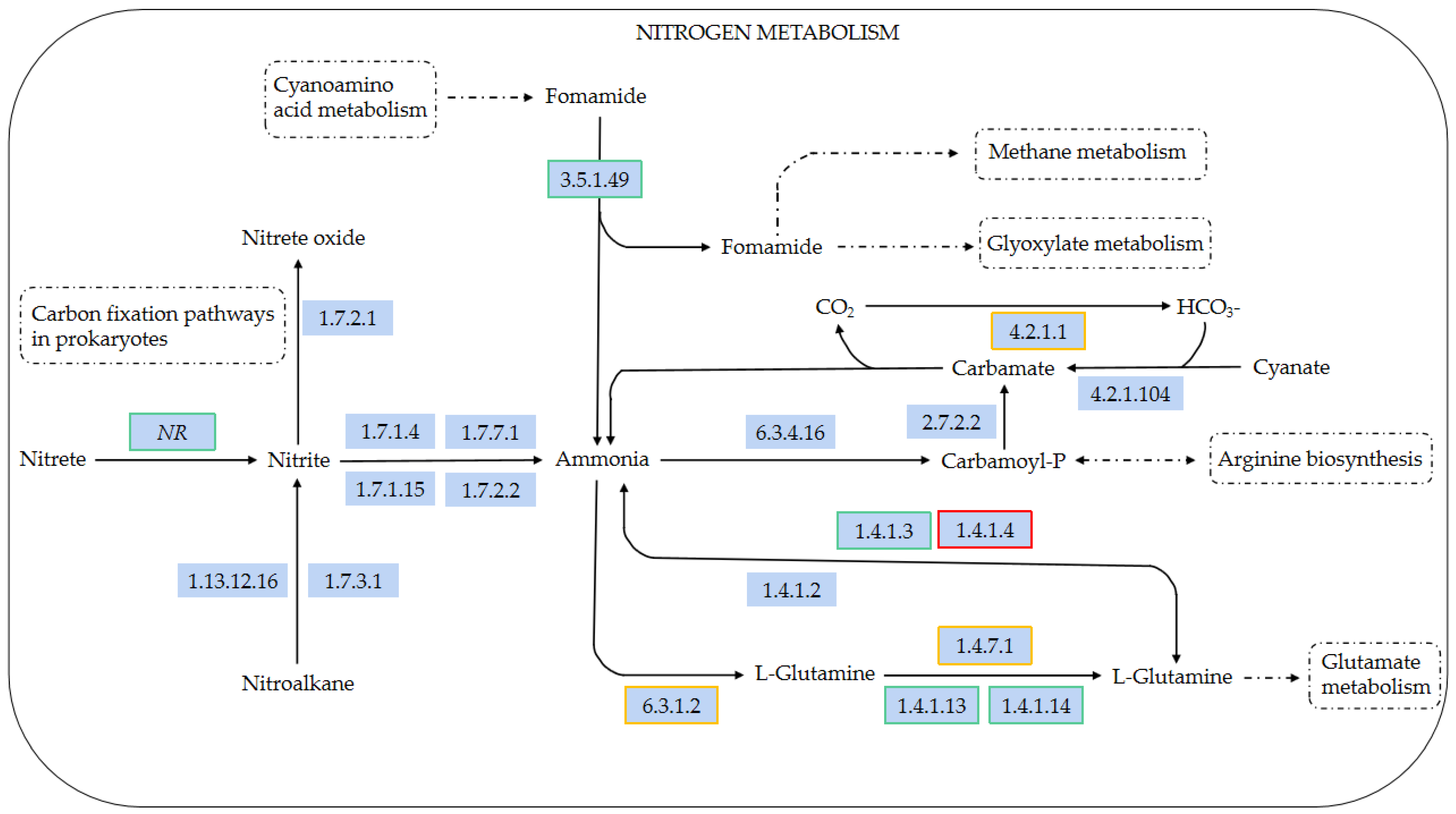
3.6. Nitrogen Metabolism
3.7. Metabolism of Porphyrin and Chlorophyll
3.8. Transcriptome Data Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cheng, H.F.; Hu, Y.A.; Zhao, J.F. Meeting China’s water shortage crisis: Current practices and challenges. Environ. Sci. Technol. 2009, 43, 240–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Chen, F.; Lei, Y.D. Climate change and shifts in cropping systems together exacerbate China’s water scarcity. Environ. Res. Lett. 2020, 15, 104060. [Google Scholar] [CrossRef]
- Huang, J.P.; Yu, H.P.; Guan, X.D.; Wang, G.Y.; Guo, R.X. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2016, 6, 166–171. [Google Scholar] [CrossRef]
- Guo, Y.; Shen, Y. Agricultural water supply/demand changes under projected future climate change in the arid region of northwestern China. J. Hydrol. 2016, 540, 257–273. [Google Scholar] [CrossRef]
- Li, X.Z.; Liu, X.D.; Ma, Z.G. Analysis on the drought characteristics in the main arid regions in the world since recent hundred-odd years. Arid Zone Res. 2004, 21, 97–103. (In Chinese) [Google Scholar]
- Ebeed, H.T.; El-Helely, A.A. Programmed cell death in plants: Insights into developmental and stress-induced cell death. Curr. Protein Pept. Sci. 2021, 22, 873–889. [Google Scholar] [CrossRef]
- Lai, L.M.; Chen, L.J.; Zheng, M.Q.; Jaing, L.H.; Zhou, J.H.; Zheng, Y.R.; Shimizu, H. Seed germination and seedling growth of five desert plants and their relevance to vegetation restoration. Ecol. Evol. 2019, 9, 2160–2170. [Google Scholar] [CrossRef]
- Ebeed, H.T.; Hassan, N.M.; Keshta, M.M.; Hassanin, O.S. Comparative analysis of seed yield and biochemical attributes in different sunflower genotypes under different levels of irrigation and salinity. Egypt. J. Bot. 2019, 59, 339–355. [Google Scholar] [CrossRef]
- Azad, N.; Rezayian, M.; Hassanpour, H.; Niknam, V.; Ebrahimzadeh, H. Physiological mechanism of salicylic acid in Mentha pulegium L. under salinity and drought stress. Braz. J. Bot. 2021, 44, 359–369. [Google Scholar] [CrossRef]
- Hassan, N.; Ebeed, H.; Aljaarany, A. Exogenous application of spermine and putrescine mitigate adversities of drought stress in wheat by protecting membranes and chloroplast ultra-structure. Physiol. Mol. Biol. Plants 2020, 26, 233–245. [Google Scholar] [CrossRef]
- Vanlerberghe, G.C.; Martyn, G.D.; Dahal, K. Alternative oxidase: A respiratory electron transport chain pathway essential for maintaining photosynthetic performance during drought stress. Physiol. Plant. 2016, 157, 322–337. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, L.; Wang, L.; Tan, M.L.; Ogutu, C.O.; Yin, Z.Y.; Zhou, J.; Wang, J.M.; Wang, L.J.; Yan, X.C. Transcriptome analysis and molecular mechanism of linseed (Linum usitatissimum L.) drought tolerance under repeated drought using single-molecule long-read sequencing. BMC Genom. 2021, 22, 109. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.J.; Zhou, J.; Guo, J. Advances in the knowledge of adaptive mechanisms mediating abiotic stress responses in Camellia sinensis. Front. Biosci.-Landmark 2021, 26, 1714–1722. [Google Scholar] [CrossRef] [PubMed]
- Tezara, W.; Mitchell, V.; Driscoll, S.P.; Lawlor, D.W. Effects of water deficit and its interaction with CO2 supply on the biochemistry and physiology of photosynthesis in sunflower. J. Exp. Bot. 2002, 53, 1781–1791. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.B.; Chen, M.; Guo, J.K.; Wang, Y.X.; Min, D.H.; Jiang, Q.Y.; Ji, H.T.; Huang, C.Y.; Wei, W.; Xu, H.J.; et al. Overexpression of the soybean (Glycine max) DRE-binding transcription factor GmDREB1 enhanced drought stress tolerance of transgenic wheat in the field. J. Exp. Bot. 2019, 71, 1842–1857. [Google Scholar] [CrossRef]
- Alam, H.; Khattak, J.Z.K.; Ksiksi, T.S.; Saleem, M.H.; Fahad, S.; SoHail, H.; Ali, Q.; Zamin, M.; El-Esawi, M.A.; Saud, S.; et al. Negative impact of long–term exposure of salinity and drought stress on native Tetraena mandavillei L. Physiol. Plant. 2021, 172, 1336–1351. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.T.; Moutinho-Pereira, J.; Correia, C.M. Drought stress effects and olive tree acclimation under a changing climate. Plants 2019, 8, 232. [Google Scholar] [CrossRef] [Green Version]
- Gessler, A.; Cailleret, M.; Joseph, J.; Schönbeck, L.; Schaub, M.; Lehmann, M.; Treydte, K.; Rigling, A.; Timofeeva, G.; Saurer, M. Drought induced tree mortality–a tree–ring isotope based conceptual model to assess mechanisms and predispositions. New Phytol. 2018, 219, 485490. [Google Scholar] [CrossRef]
- DeSoto, L.; Cailleret, M.; Sterck, F.; Jansen, S.; Kramer, K.; Robert, E.M.R.; Aakala, T.; Amoroso, M.M.; Bigler, C.; Camarero, J.J.; et al. Low growth resilience to drought is related to future mortality risk in trees. Nat. Commun. 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Shao, Y.Y.; Zhang, Y.Q.; Wu, X.Q.; Bourque, C.P.A.; Zhang, J.T.; Qin, S.G.; Wu, B. Relating historical vegetation cover to aridity patterns in the greater desert region of northern China: Implications to planned and existing restoration projects. Ecol. Indic. 2018, 89, 528–537. [Google Scholar] [CrossRef]
- Li, Q.H.; Jiang, Z.P. Research on Nitraria tangutorum; China Forestry Press: Beijing, China, 2011; p. 172. (In Chinese) [Google Scholar]
- Li, S.H.; Mason, J.A.; Xu, Y.H.; Xu, C.; Zheng, G.; Li, J.C.; Yizhaq, H.; Pan, S.; Lu, H.Y.; Xu, Z.W. Biogeomorphology of nebkhas in the Mu Us dune field, north–central China: Chronological and morphological results. Geomorphology 2021, 394, 107979. [Google Scholar] [CrossRef]
- Wei, Y.J.; Dang, X.H.; Wang, J.; Gao, J.L.; Gao, Y. Response of C:N:P in the plant-soil system and stoichiometric homeostasis of Nitraria tangutorum leaves in the oasis-desert ecotone, Northwest China. J. Arid Land 2021, 13, 934–946. [Google Scholar] [CrossRef]
- Zhu, L.M.; Lu, L.; Yang, L.M.; Hao, Z.D.; Chen, J.H.; Cheng, T.L. The full-length transcriptome sequencing and identification of Na+/H+ antiporter genes in halophyte Nitraria tangutorum Bobrov. Genes 2021, 12, 836. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.T.; Zhou, Y.W.; Zhang, J.B.; Peng, Y.; Yang, X.Y.; Hao, Z.D.; Lu, Y.; Wu, W.H.; Cheng, T.L.; Shi, J.S.; et al. Integrative analysis of transcriptome and proteome revealed nectary and nectar traits in the plant-pollinator interaction of Nitraria tangutorum Bobrov. BMC Plant Biol. 2021, 21, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Chen, X.Y.; Zhu, L.M.; Li, M.J.; Zhang, J.B.; Yang, X.Y.; Wang, P.K.; Lu, Y.; Cheng, T.L.; Shi, J.S.; et al. NtCIPK9: A calcineurin B-Like protein-interacting protein kinase from the halophyte Nitraria tangutorum, enhances Arabidopsis salt tolerance. Front. Plant Sci. 2020, 11, 1112. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.Q.; Gao, S.; Li, X.P.; Zhnng, Y.; Ma, B.J.; Wang, Y.C. Exogenous methyl jasmonate promotes salt stress-induced growth inhibition and prioritizes defense response of Nitraria tangutorum Bobr. Physiol. Plant. 2021, 172, 162–175. [Google Scholar] [CrossRef]
- Zhao, J.Q.; Wang, Y.M.; Yang, Y.L.; Zeng, Y.; Wang, Q.L.; Shao, Y.; Mei, L.J.; Shi, Y.P.; Tao, Y.D. Isolation and identification of antioxidant and α-glucosidase inhibitory compounds from fruit juice of Nitraria tangutorum. Food Chem. 2017, 227, 93–101. [Google Scholar] [CrossRef]
- Abla, M.; Zha, X.; Wang, Y.; Wang, X.Y.; Gao, F.; Zhou, Y.J.; Feng, J.C. Characterization of the complete chloroplast genome of Nitraria tangutorum, a desert shrub. J. Genet. 2019, 98, 91. [Google Scholar] [CrossRef]
- Zhan, J.P.; Li, G.S.; Ryu, C.H.; Ma, C.; Zhang, S.S.; Lloyd, A.; Hunter, B.G.; Larkins, B.A. Opaque-2 regulates a complex gene network associated with cell differentiation and storage functions of maize endosperm. Plant Cell 2018, 30, 2425–2446. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.J.; Li, H.W.; Jiang, Y.P.; Tang, Y.L.; Zhao, G.Q.; Zhang, Y.L.; Yang, S.J.; Qiu, H.S.; Wang, Y.C.; Guo, J.M.; et al. Transcriptomic analysis of female panicles reveals gene expression responses to drought stress in maize (Zea mays L.). Agronomy 2020, 10, 313. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.L.; Bui, B.C. Fine mapping for drought tolerance in rice (Oryza sativa L.). Omonrice 2008, 16, 9–15. [Google Scholar]
- Fàbregas, N.; Lozano-Elena, F.; Blasco-Escámez, D.; Tohge, T.; Martínez-Andújar, C.; Albacete, A.; Osorio, S.; Bustamante, M.; Riechmann, J.L.; Nomura, T.; et al. Overexpression of the vascular brassinosteroid receptor BRL3 confers drought resistance without penalizing plant growth. Nat. Commun. 2018, 9, 4680. [Google Scholar] [CrossRef] [Green Version]
- Deyholos, M.K. Making the most of drought and salinity transcriptomics. Plant Cell Environ. 2010, 33, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Costa, V.; Angelini, C.; Feis, I.; Ciccodicola, A. Uncovering the complexity of transcriptomes with RNA–seq. J. Biomed. Biotechnol. 2010, 2010, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Hainaut, M.; Delhomme, N.; Mannapperuma, C.; Immerzeel, P.; Street, N.R.; Henrissat, B.; Mellerowicz, W.J. Poplar carbohydrate-active enzymes: Whole-genome annotation and functional analyses based on RNA expression data. Plant J. 2019, 99, 589–609. [Google Scholar] [CrossRef]
- Yang, J.L.; Wang, H.Z.; Zhao, S.C.; Liu, X.; Zhang, X.; Wu, W.L.; Li, C.H. Overexpression levels of LbDREB6 differentially affect growth, drought, and disease tolerance in poplar. Front. Plant Sci. 2020, 11, 1661. [Google Scholar] [CrossRef]
- Wei, W.; Liang, D.W.; Bian, X.H.; Shen, M.; Xiao, J.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Lv, J.; Chen, X.; et al. GmWRKY54 improves drought tolerance through activating genes in abscisic acid and Ca2+ signaling pathways in transgenic soybean. Plant J. 2019, 100, 384–398. [Google Scholar] [CrossRef]
- Wang, K.; Bu, T.T.; Cheng, Q.; Dong, L.D.; Su, T.; Chen, Z.M.; Kong, F.J.; Gong, Z.Z.; Liu, B.H.; Li, M.N. Two homologous LHY pairs negatively control soybean drought tolerance by repressing the abscisic acid responses. New Phytol. 2020, 229, 2660–2675. [Google Scholar] [CrossRef]
- Danilevskaya, O.N.; Yu, G.X.; Meng, X.; Xu, J.; Stephenson, E.; Estrada, S.; Chilakamarri, S.; Zastrow-Hayes, G.; Thatcher, S. Developmental and transcriptional responses of maize to drought stress under field conditions. Plant Direct 2019, 3, e00129. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.M.; Qu, H.B.; Zhang, H.D.; Liu, S.; Li, Y.; Zhang, C.Q. Hormone and RNA–seq analyses reveal the mechanisms underlying differences in seed vigour at different maize ear positions. Plant Mol. Biol. 2019, 99, 461–476. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.H.; Xin, Z.M.; Gao, T.T.; Wang, S.X.; Xu, J.; Sun, F. Reproductive allocation in four desert species of the genus Nitraria L. Acta. Ecol. Sin. 2012, 32, 5054–5061. (In Chinese) [Google Scholar]
- Kang, J.J.; Zhao, W.Z.; Zhao, M.; Zheng, Y.; Yang, F. NaCl and Na2SiO3 coexistence strengthens growth of the succulent xerophyte Nitraria tangutorum under drought. Plant Growth Regul. 2015, 77, 223–232. [Google Scholar] [CrossRef]
- Ren, Y.; Lu, Q.; Wu, B.; Liu, M.H. Specific leaf area and leaf dry matter content of Nitraria tangutorum in the artificially simulated precipitation. Acta Ecol. Sin. 2015, 35, 4707–4715. (In Chinese) [Google Scholar]
- Kang, J.J.; Zhao, W.Z.; Zhou, H.; Wang, Z.W. The features of main osmolytes, silicon and their coupling effects in improving drought resistance of the typical xerophytes in the desert areas of Northwest China. Land Degrad. Dev. 2020, 31, 2720–2733. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, G.Y.; Ramakrishnan, M.; Vinod, K.K.; Yrjälä, K.; Satheesh, V.; Cho, J.; Fu, Y.; Zhou, M.B. Multi-omics analysis of cellular pathways involved in different rapid growth stages of moso bamboo. Tree Physiol. 2020, 40, 1487–1508. [Google Scholar] [CrossRef]
- Novaes, E.; Drost, D.R.; Farmerie, W.G.; Pappas, G.J., Jr.; Grattapaglia, D.; Sederoff, R.; Kirst, M. High-throughput gene and SNP discovery in Eucalyptus grandis, an uncharacterized genome. BMC Genom. 2008, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.H.; Bao, S.Y.; Liu, J.; Zhuang, Y. De novo sequencing and analysis of the transcriptome of the wild eggplant species Solanum aculeatissimum in response to Verticillium dahliae. Plant Mol. Biol. Rep. 2016, 34, 1193–1203. [Google Scholar] [CrossRef]
- Hou, R.; Bao, Z.M.; Wang, S.; Su, H.L.; Li, Y.; Du, H.X.; Hu, J.J.; Wang, S.; Hu, X.L. Transcriptome sequencing and De Novo analysis for yesso scallop (Patinopecten yessoensis) Using 454 GS FLX. PLoS ONE 2011, 6, e21560. [Google Scholar] [CrossRef] [Green Version]
- Long, Y.; Zhang, J.; Tian, X.; Wu, S.; Zhang, Q.; Zhang, J.; Zhang, D.; Pei, X. De novo assembly of the desert tree Haloxylon ammodendron (C. A. Mey.) based on RNA-seq data provides insight into drought response, gene discovery and marker identification. BMC Genom. 2014, 15, 1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Wei, W.; Pang, X.; Wang, X.; Zhang, H.; Dong, B.; Xing, Y.; Li, X.; Wang, M. Comparative transcriptome profiling of a desert evergreen shrub, Ammopiptanthus mongolicus, in response to drought and cold stresses. BMC Genom. 2014, 15, 671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Na, J.; Nie, H.; Kim, J.; Lee, J.; Kim, S. Comprehensive transcriptome profiling of Caragana microphylla in response to salt condition using de novo assembly. Biotechnol. Lett. 2021, 43, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.S.; Ma, Y.H.; Feng, Z.P.; Zhang, X.F. Transcriptomic analysis of drought stress responses of sea buckthorn (Hippophae rhamnoides subsp. sinensis) by RNA-Seq. PLoS ONE 2018, 13, e0202213. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, T.; Khalid, S.; Abdullah, M.; Ahmed, Z.; Shah, M.K.N.; Ghafoor, A.; Du, X.M. Insights into drought stress signaling in plants and the molecular genetic basis of cotton drought tolerance. Cells 2019, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.X.; Ding, Y.L.; Yang, Y.Q.; Song, C.P.; Wang, B.S.; Yang, S.H.; Guo, Y.; Gong, Z.Z. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 2021, 63, 53–78. [Google Scholar] [CrossRef]
- Chen, Q.C.; Hu, T.; Li, X.H.; Song, Y.W.; Zhu, J.K.; Chen, L.Q.; Zhao, Y. Phosphorylation of SWEET sucrose transporters regulates plant root: Shoot ratio under drought. Nat. Plants 2022, 8, 68–77. [Google Scholar] [CrossRef]
- Li, W.; Fu, L.F.; Geng, Z.W.; Zhao, X.J.; Liu, Q.H.; Jiang, X.Q. Physiological characteristic changes and full-length transcriptome of rose (Rosa chinensis) roots and leaves in response to drought stress. Plant Cell Physiol. 2020, 61, 2153–2166. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef]
- Zhang, C.X.; Meng, S.; Li, M.J.; Zhao, Z. Transcriptomic insight into nitrogen uptake and metabolism of Populus simonii in response to drought and low nitrogen stresses. Tree Physiol. 2018, 38, 1672–1684. [Google Scholar] [CrossRef]
- Cai, H.M.; Xiao, J.H.; Zhang, Q.F.; Lian, X.M. Co–suppressed glutamine synthase 2 gene modifies nitrogen metabolism and plant growth in rice. Chin. Sci. Bull. 2010, 55, 871–882. [Google Scholar] [CrossRef]
- Xu, X.; Fu, X.; Liao, H. Advances in study of ammonium assimilation and its regulatory mechanism in plants. Bull. Bot. 2016, 51, 152–166. (In Chinese) [Google Scholar]
- Campbell, W.H. Nitrate reductase and its role in nitrate assimilation in plants. Physiol. Plant. 1988, 74, 214–219. [Google Scholar] [CrossRef]
- Caravaca, F.; Figueroa, D.; Barea, J.M.; Azcón-Aguilar, C.; Roldán, A. Effect of mycorrhizal inoculation on nutrient acquisition, gas exchange, and nitrate reductase activity of two Mediterranean-autochthonous shrub species under drought stress. J. Plant Nutr. 2004, 27, 57–74. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate reductase regulates plant nitric oxide homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef]
- Han, R.C.; LI, C.Y.; Rasheed, A.; Pan, X.H.; Shi, Q.H.; Wu, Z.M. Reducing phosphorylation of nitrate reductase improves nitrate assimilation in rice. J. Integr. Agric. 2022, 21, 15–25. [Google Scholar] [CrossRef]
- Han, M.L.; Lv, Q.Y.; Zhang, J.; Wang, T.; Zhang, C.X.; Tan, R.J.; Wang, T.L.; Zhong, L.Y.; Gao, Y.Q.; Chao, Z.F.; et al. Decreasing nitrogen assimilation under drought stress by suppressing DST-mediated activation of nitrate reductase 1.2 in rice. Mol. Plant 2021, 15, 167–178. [Google Scholar] [CrossRef]
- Yadav, R.S.; Sharwa, R.L.; Pandey, U.K. Effects of various water potential treatment on nitrogen reductase activity in wheat genotypes. Agric. Sci. Digest. 1998, 18, 73–75. [Google Scholar]
- Robredo, A.; Perez-Lopez, U.; Miranda-Apodaca, J.; Lacuesta, M.; Mena-Petite, A.; Munoz-Rueda, A. Elevated CO2 reduces the drought effect on nitrogen metabolism in barley plants during drought and subsequent recovery. J. Exp. Bot. 2011, 71, 399–408. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, E.; Rubio-Wilhelmi, M.d.M.; Rios, J.J.; Blasco, B.; Rosales, M.A.; Melgarejo, R.; Romero, L.; Ruiz, J.M. Ammonia production and assimilation: Its importance as a tolerance mechanism during moderate water deficit in tomato plants. J. Plant Physiol. 2011, 168, 816–823. [Google Scholar] [CrossRef]
- Meng, S.; Zhang, C.X.; Su, L.; Li, Y.M.; Zhao, Z. Nitrogen uptake and metabolism of Populus simonii in response to PEG-induced drought stress. Environ. Exp. Bot. 2016, 123, 78–87. [Google Scholar] [CrossRef]
- Liu, Y.L.; Duan, X.L.; Zhao, X.D.; Ding, W.L.; Wang, Y.W.; Xiong, Y. Diverse nitrogen signals activate convergent ROP2-TOR signaling in Arabidopsis. Dev. Cell 2021, 56, 1283–1295.e5. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Wilhelmi, M.d.M.; Sanchez-Rodriguez, E.; Leyva, R.; Blasco, B.; Romero, L.; Blumwald, E.; Ruiz, J.M. Response of carbon and nitrogen-rich metabolites to nitrogen deficiency in PSARK:: IPT tobacco plants. Plant Physiol. Biochem. 2012, 57, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Rana, N.K.; Mohanpuria, P.; Yadav, S.K. Expression of tea cytosolic glutamine synthetase is tissue specific and induced by cadmium and salt stress. Biol. Plant. 2008, 52, 361–364. [Google Scholar] [CrossRef]
- Goel, P.; Singh, A.K. Abiotic stresses downregulate key genes involved in nitrogen uptake and assimilation in Brassica juncea L. PLoS ONE 2015, 10, e0143645. [Google Scholar]
- Bowsher, C.G.; Lacey, A.E.; Hanke, G.T.; Clarkson, D.T.; Saker, L.R.; Stulen, L.; Emes, M.J. The effect of Glc6P uptake and its subsequent oxidation within pea root plastids on nitrite reduction and glutamate synthesis. J. Exp. Bot. 2007, 58, 1109–1118. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Nian, J.Q.; Xie, X.Z.; Yu, H.; Zhang, J.; Bai, J.T.; Dong, G.J.; Hu, J.; Bo, B.; Chen, L.C.; et al. Genetic variations in ARE1 mediate grain yield by modulating nitrogen utilization in rice. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yoneyama, T.; Suzuki, A. Exploration of nitrate-to-glutamate assimilation in non-photosynthetic roots of higher plants by studies of 15N-tracing, enzymes involved, reductant supply, and nitrate signaling: A review and synthesis. Plant Physiol. Biochem. 2019, 136, 245–254. [Google Scholar] [CrossRef]
- Borsani, O.; Diaz, P.; Monza, J. Proline is involved in water stress responses of Lotus corniculatus nitrogen fixing and nitrate fed plants. J. Plant Physiol. 1999, 155, 269–273. [Google Scholar] [CrossRef]
- Martinelli, T.; Whittaker, A.; Bochicchio, A.; Vazzana, C.; Suzuki, A.; Masclaux-Daubresse, C. Amino acid pattern and glutamate metabolism during dehydration stress in the ‘resurrection’ plant Sporobolus stapfianus: A comparison between desiccation-sensitive and desiccation-tolerant leaves. J. Exp. Bot. 2007, 58, 3037–3046. [Google Scholar] [CrossRef] [Green Version]
- Curtis, T.Y.; Bo, V.; Tucker, A.; Halford, N. Construction of a network describing asparagine metabolism in plants and its application to the identification of genes affecting asparagine metabolism in wheat under drought and nutritional stress. Food Energy Secur. 2018, 7, e00126. [Google Scholar] [CrossRef] [PubMed]
- Bota, J.; Medrano, H.; Flexas, J. Is photosynthesis limited by decreased RuBisCo activity and RuBP content under progressive water stress? New Phytol. 2010, 162, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Lang, Y.; Wang, M.; Xia, J.B.; Zhao, Q.K. Effects of soil drought stress on photosynthetic gas exchange traits and chlorophyll fluorescence in Forsythia suspensa. J. For. Res. 2018, 29, 45–53. [Google Scholar] [CrossRef]
- Zhuang, J.; Wang, Y.L.; Chi, Y.G.; Zhou, L.; Chen, J.J.; Zhou, W.; Song, J.; Zhao, N.; Ding, J.X. Drought stress strengthens the link between chlorophyll fluorescence parameters and photosynthetic traits. PeerJ 2020, 8, e10046. [Google Scholar] [CrossRef] [PubMed]
- Cornah, J.E.; Terry, M.J.; Smith, A.G. Green or red: What stops the traffic in the tetrapyrrole pathway. Trends Plant Sci. 2003, 8, 224–230. [Google Scholar] [CrossRef]
- Richter, A.; Peter, E.; Pors, Y.; Lorenzen, S.; Grimm, B.; Czarnecki, O. Rapid dark repression of 5-aminolevulinic acid synthesis in green barley leaves. Plant Cell Physiol. 2010, 51, 670–681. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.V. Regulation of chlorophyll biosynthesis and degradation by silt stress in sunflower leaves. Sci. Hortic. 2004, 103, 93–99. [Google Scholar] [CrossRef]
- Wang, Y.X.; Li, X.Y.; Liu, N.N.; Wei, S.M.; Wang, J.N.; Qin, F.J.; Suo, B. The iTRAQ-based chloroplast proteomic analysis of Triticum aestivum L. leaves subjected to drought stress and 5-aminolevulinic acid alleviation reveals several proteins involved in the protection of photosynthesis. BMC Plant Biol. 2020, 20, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Beale, S. Green genes gleaned. Trends Plant Sci. 2005, 10, 309–312. [Google Scholar] [CrossRef]
- Ma, Q.L.; Xu, X.; Xie, Y.Z.; Huang, T.; Wang, W.J.; Zhao, L.J. Comparative metabolomic analysis of the metabolism pathways under drought stress in alfalfa leaves. Environ. Exp. Bot. 2021, 183, 104329. [Google Scholar] [CrossRef]
- Merewitz, E.; Xu, Y.; Huang, B. Differentially expressed genes associated with improved drought tolerance in creeping bentgrass overexpressing a gene for cytokinin biosynthesis. PLoS ONE 2016, 11, e0166676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Chang, E.J.; Yu, X.X.; Chen, Y.H.; Yang, Q.S.; Cao, Y.T.; Li, X.K.; Wang, Y.H.; Fu, A.G.; Xu, M. Molecular characterization of Magnesium Chelatase in soybean [Glycine max (L.) Merr.]. Front Plant Sci. 2018, 9, 720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, M.J.; Kendrick, R.E. Feedback inhibition of chlorophyll synthesis in the phytochrome chromophore-deficient aurea and yellow-green 2 mutants of tomato. Plant Physiol. 1999, 119, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phung, T.H.; Jung, H.; Park, J.H.; Kim, J.G.; Back, K.; Jung, S. Porphyrin biosynthesis control under water stress: Sustained porphyrin status correlates with drought tolerance in transgenic rice. Plant Physiol. 2011, 157, 1746–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.L.; Lin, N.; Liu, X.Y.; Yang, S.; Wang, W.; Wan, X.C. From chloroplast biogenesis to chlorophyll accumulation: The interplay of light and hormones on gene expression in Camellia sinensis cv. Shuchazao Leaves. Front. Plant Sci. 2020, 11, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaleel, C.A.; Gopi, R.; Sankar, B.; Gomathinayagam, M.; Panneerselvam, R. Differential responses in water use efficiency in two varieties of Catharanthus roseus under drought stress. Comptes Rendus Biol. 2008, 331, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Asthir, B. Molecular responses to drought stress in plants. Biol. Plant. 2017, 61, 201–209. [Google Scholar] [CrossRef]
- Sato, Y.; Morita, R.; Nishimura, M.; Yamaguchi, H.; Kusaba, M. Mendel’s green cotyledon gene encodes a positive regulator of the chlorophyll-degrading pathway. Proc. Natl. Acad. Sci. USA 2007, 104, 14169–14174. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| Cluster-40906.35332 | TTCCTCCCCACGCACTTTTC | CCAGTTCCGGCCTGTCATTC |
| Cluster-40906.11509 | AACTCTGAGCAGTAACCAAGCAA | ACTCTGAATTAGAATATGCAACCGA |
| Cluster-40906.56890 | ATCCAAGCGACATGGCGAA | TTCTGCAGGAAACTTGAAGTCGTAG |
| Cluster-40906.174397 | CCAATGATACCGCATCCAAT | ATGATCTCGGAAAAGGTGGAC |
| Cluster-28726.1 | CCATCATCACCACAGAATGTAAAA | TCCAGATCGGAGTACAAAATTCAC |
| Cluster-40906.57689 | TTGCTACCCCAAAACTGACTTC | CATTTCCTCTTCCGCCTCAC |
| Cluster-40906.102382 | ATTCAACCCAATTCTCACTCTTCTT | ATAACGCAGGAGACGCACCA |
| Cluster-40906.114615 | ACATCCTTCTTTCCATCCTCCA | GAATGATGTATGACCGACCACC |
| Cluster-40906.35748 | TCCGGGAAATTGAGAGTTGC | AAGGGGGTGAGGATGAGAAA |
| Cluster-40906.176764 | AGAAGCTGGGGAAAATGGGTAT | GAGTTGGGGGAAGTTGAGGAC |
| NsActin | GGAATCCACGAGACCACCTACA | GATTGATCCTCCGATCCAGACA |
| Database Name | Unigene Numbers | Ratio (%) |
|---|---|---|
| Nr (NCBI, Non-redundant Protein Sequences Database) | 176,393 | 63.81 |
| Nt (NCBI, Nucleotide Sequences Database) | 116,688 | 42.21 |
| KO (KEGG Orthology) | 70,817 | 25.61 |
| Swiss-Prot (Annotated Protein Sequence Database) | 129,949 | 47.01 |
| PFAM (Family Protein Database) | 122,945 | 44.47 |
| GO (Gene Ontology Database) | 122,945 | 44.47 |
| KOG (euKaryotic Ortholog Groups) | 48,287 | 17.46 |
| All databases | 25,380 | 9.18 |
| At least one database | 195,214 | 70.62 |
| Total unigenes | 276,423 | 100 |
| Gene ID | log2 Fold Change | p-Value | Description |
|---|---|---|---|
| Cluster-40906.113941 | −3.5898 | 7.38 × 10−74 | Oxidoreductase activity//chlorophyllide a oxygenase (overall) activity//2 iron, 2 sulfur cluster binding |
| Cluster-40906.41614 | 4.6013 | 1.57 × 10−72 | Unfolded protein binding//ATP binding |
| Cluster-40906.93310 | −2.0399 | 1.15 × 10−67 | Protein binding |
| Cluster-40906.110044 | −5.814 | 1.43 × 10−66 | Calcium ion binding |
| Cluster-40906.175859 | 4.7681 | 1.49 × 10−60 | DNA binding//transposase activity//ATP binding//unfolded protein binding |
| Cluster-40906.104561 | −4.0125 | 1.19 × 10−53 | Transporter activity |
| Cluster-40906.116353 | −2.4038 | 1.41 × 10−53 | Chitinase activity |
| Cluster-40906.83443 | 4.8401 | 9.86 × 10−53 | Unfolded protein binding//ATP binding |
| Cluster-40906.62892 | 3.2606 | 7.27 × 10−52 | Hypothetical protein CUMW_183970 |
| Cluster-40906.109750 | −7.2303 | 1.29 × 10−51 | -- |
| Cluster-40906.123767 | 1.5802 | 2.95 × 10−50 | DnaJ homolog subfamily B member 7 isoform X2 |
| Cluster-40906.107883 | 2.056 | 7.53 × 10−50 | Chlorophyll a-b-binding protein CP26, chloroplastic-like |
| Cluster-40906.175857 | 5.4749 | 2.58 × 10−48 | Heat shock protein 90-1 |
| Cluster-40906.108887 | −2.4863 | 3.24 × 10−48 | Transmembrane transport |
| Cluster-40906.121414 | −4.0105 | 2.51 × 10−46 | Phosphatidylinositol-4-phosphate binding |
| Cluster-40906.112950 | −14.664 | 1.01 × 10−45 | Zinc ion binding |
| Cluster-40906.115294 | 5.5761 | 1.08 × 10−45 | Heme binding//peroxidase activity |
| Cluster-40906.112156 | 10.614 | 2.10 × 10−43 | Hydrolase activity, hydrolyzing O-glycosyl compounds |
| Cluster-40906.147611 | 3.1736 | 1.04 × 10−42 | Catalytic activity//oxidoreductase activity//3-hydroxyacyl-CoA dehydrogenase activity//peptidase activity//hydrolyase activity |
| Cluster-40906.119123 | 1.4464 | 4.56 × 10−42 | Protein binding |
| Cluster-40906.111055 | −1.3917 | 2.64 × 10−41 | ATP binding//GTP binding//GTPase activity//cytidylate kinase activity//ATPase activity |
| Cluster-40906.117956 | −7.1941 | 3.62 × 10−41 | Adenyl-nucleotide exchange factor activity//transcription factor activity, sequence-specific DNA binding//exodeoxyribonuclease VII activity//protein homodimerization activity//unfolded protein binding//chaperone binding//protein binding, bridging//protein tag//motor activity//structural molecule activity//acid–amino acid ligase activity//receptor binding |
| Cluster-40906.105679 | 3.2201 | 1.26 × 10−40 | 4-Hydroxy-3-methylbut-2-en-1-yl diphosphate synthase activity |
| Cluster-40906.35332 | 3.1073 | 2.86 × 10−39 | Protein binding//catalytic activity |
| Cluster-40906.111273 | −1.6128 | 7.15 × 10−38 | Transcription factor activity, sequence-specific DNA binding |
| Cluster-40906.111303 | 4.1859 | 1.28 × 10−37 | Hypothetical protein B456_006G088300 |
| Cluster-40906.118097 | 2.0982 | 4.43 × 10−37 | -- |
| Cluster-40906.89392 | 3.4364 | 1.82 × 10−36 | Protein dimerization activity |
| Cluster-40906.129546 | −4.631 | 3.31 × 10−36 | Alcohol dehydrogenase 1 |
| Cluster-40906.49184 | −5.5647 | 4.42 × 10−36 | Structural constituent of ribosome |
| KEGG Pathway | ID | DEG Number | p-Value |
|---|---|---|---|
| Ribosome | ko03010 | 177 | 1.26 × 10−7 |
| Plant hormone signal transduction | ko04075 | 101 | 1.86 × 10−6 |
| Protein processing in endoplasmic reticulum | ko04141 | 139 | 1.69 × 10−5 |
| Porphyrin and chlorophyll metabolism | ko00860 | 44 | 0.001141408 |
| Spliceosome | ko03040 | 110 | 0.001860469 |
| Phenylalanine, tyrosine, and tryptophan biosynthesis | ko00400 | 34 | 0.005868948 |
| Starch and sucrose metabolism | ko00500 | 98 | 0.006867847 |
| Anthocyanin biosynthesis | ko00942 | 8 | 0.011943248 |
| Flavonoid biosynthesis | ko00941 | 17 | 0.013126286 |
| Alpha-linolenic acid metabolism | ko00592 | 29 | 0.032232635 |
| Taurine and hypotaurine metabolism | ko00430 | 12 | 0.034211087 |
| Photosynthesis (antenna proteins) | ko00196 | 26 | 0.035911307 |
| Limonene and pinene degradation | ko00903 | 14 | 0.038622556 |
| Stilbenoid, diarylheptanoid, and gingerol biosynthesis | ko00945 | 14 | 0.046974746 |
| Enzyme | Gene Name | Gene ID | Up/Downregulated |
|---|---|---|---|
| 6.1.1.17 | Glutamine tRNA synthetase | Cluster-40906.71722 | Down |
| Cluster-40906.54653 | Down | ||
| 1.2.1.70 | Glutamyl-tNRA reductase | Cluster-40906.114395 | Up |
| Cluster-40906.129403 | Up | ||
| 5.4.3.8 | Glutamate-1-hemialdehyde transaminase | Cluster-40906.192599 | Up |
| Cluster-40906.119289 | Down | ||
| 4.2.1.24 | Bile pigment synthase | Cluster-40906.85742 | Up |
| Cluster-40906.85733 | Up | ||
| Cluster-40906.85731 | Up | ||
| 4.2.1.75 | Uroporphyrinogen decarboxylase | Cluster-40906.104102 | Up |
| 1.3.3.4/1.3.3.15 | Protoporphyrinogen oxidase | Cluster-40906.34049 | Up |
| 4.99.1.1/4.99.1.9 | Ferrochelatase | Cluster-40906.121603 | Up |
| Cluster-40906.121607 | Up | ||
| Cluster-40906.121031 | Up | ||
| Cluster-40906.100839 | Down | ||
| Cluster-40906.93744 | Down | ||
| Cluster-40906.100841 | Down | ||
| 2.5.1.- | Heme O ferric synthetase | Cluster-40906.118608 | Up |
| COX15 | Cluster-40906.59147 | Up | |
| 6.6.1.1 | Magnesium chelatase H subgroup | Cluster-40906.117245 | Up |
| Cluster-40906.137917 | Up | ||
| Cluster-40906.157365 | Up | ||
| Cluster-40906.106867 | Up | ||
| Cluster-40906.93092 | Up | ||
| Cluster-40906.118198 | Up | ||
| Cluster-40906.118566 | Up | ||
| Cluster-40906.110295 | Up | ||
| Cluster-40906.121612 | Down | ||
| Cluster-40906.103817 | Down | ||
| Cluster-40906.106857 | Down | ||
| 1.3.1.75 | Divinyl reductase | Cluster-40906.109654 | Down |
| 1.3.1.33 | Prochlorophyll reductase | Cluster-40906.168882 | Up |
| Cluster-40906.99070 | |||
| 2.5.1.62 | Chlorophyll synthase | Cluster-40906.89493 | Down |
| 1.1.1.294 | Chlorophyll b reductase | Cluster-40906.133380 | Up |
| Cluster-40906.59982 | Up | ||
| Cluster-40906.59985 | Down | ||
| Cluster-40906.27543 | Down | ||
| 1.17.7.2 | 7-Hydroxymethyl chlorophyll a reductase | Cluster-40906.185694 | Up |
| 3.1.1.14 | Chlorophyllase | Cluster-40906.151688 | Down |
| 1.14.1517 | PAO | Cluster-40906.92868 | Up |
| 2.5.1.133 | Chlorophyll synthase | Cluster-40906.89493 | Down |
| 1.3.1.111 | CHIP | Cluster-40906.113828 | Up |
| Cluster-40906.113825 | Down | ||
| Cluster-40906.113826 | Down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Duan, N.; Chen, X.; Li, H.; Zhao, X.; Duo, P.; Wang, J.; Li, Q. Metabolic Pathways Involved in the Drought Stress Response of Nitraria tangutorum as Revealed by Transcriptome Analysis. Forests 2022, 13, 509. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040509
Liu C, Duan N, Chen X, Li H, Zhao X, Duo P, Wang J, Li Q. Metabolic Pathways Involved in the Drought Stress Response of Nitraria tangutorum as Revealed by Transcriptome Analysis. Forests. 2022; 13(4):509. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040509
Chicago/Turabian StyleLiu, Chenggong, Na Duan, Xiaona Chen, Huiqing Li, Xiulian Zhao, Puzeng Duo, Ji Wang, and Qinghe Li. 2022. "Metabolic Pathways Involved in the Drought Stress Response of Nitraria tangutorum as Revealed by Transcriptome Analysis" Forests 13, no. 4: 509. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040509