
Development of Genotype-Specific Anti-Bovine Rotavirus A Immunoglobulin Yolk Based on a Current Molecular Epidemiological Analysis of Bovine Rotaviruses A Collected in Japan during 2017–2020

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples, RNA Extraction, and RT-PCR Amplification
2.2. Sequence and Phylogenetic Analyses
2.3. Virus Isolation
2.4. Production of Anti-Bovine Rotavirus A Immunoglobulin Y and Control Immunoglobulin Y
2.5. Neutralization Assay
2.6. Statistical Analysis
3. Results
3.1. Sequence and Phylogenetic Analysis of VP7
3.2. Sequence and Phylogenetic Analysis of VP4
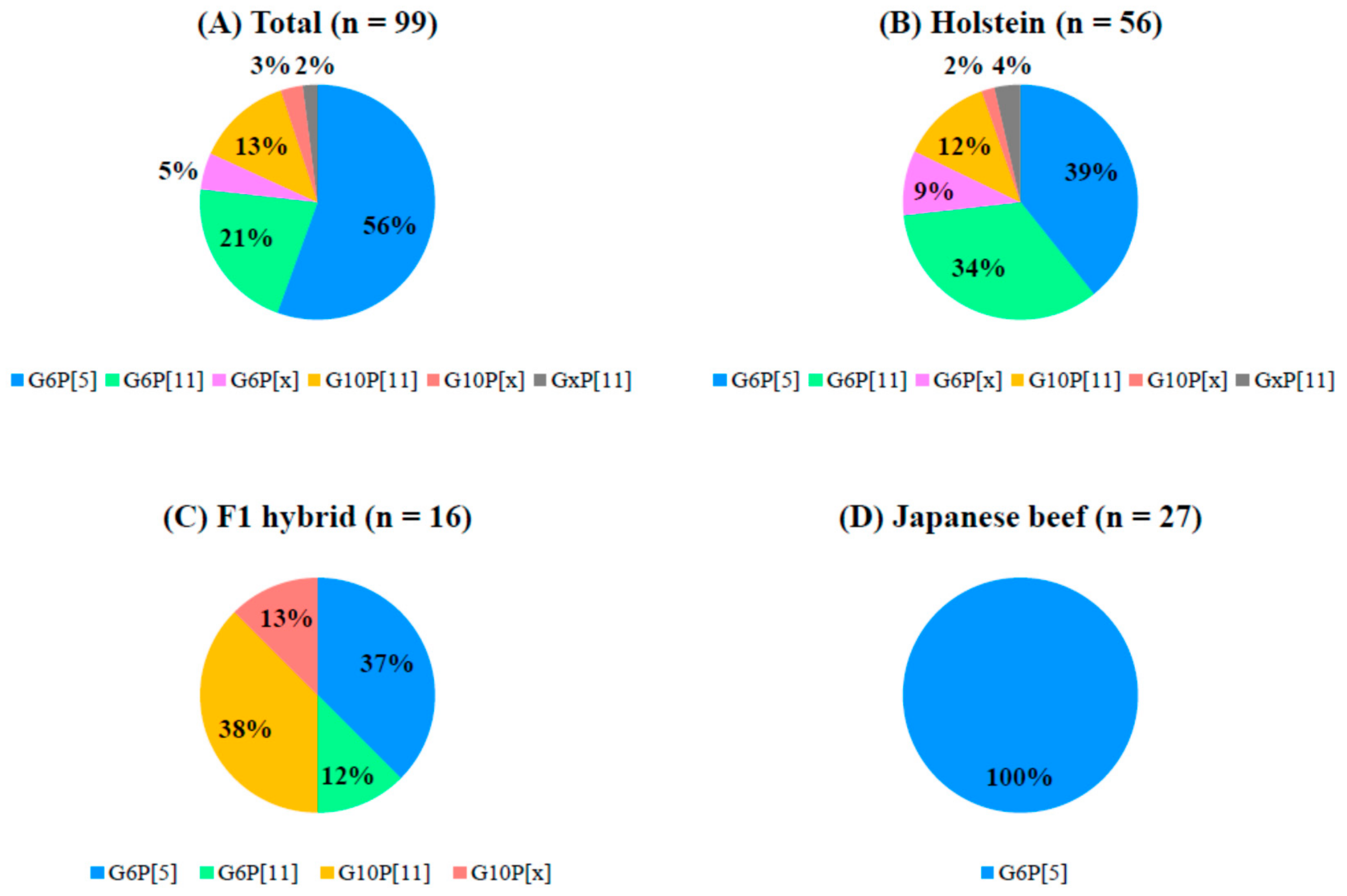
3.3. Analysis of the Combination of G and P Genotypes
3.4. Production of Anti-Bovine Rotavirus A Immunoglobulin Y and Control Immunoglobulin Y
3.5. Neutralization Assay
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Annual Case Report for Livestock Disease from the Ministry of Agriculture, Forestry and Fisheries of Japan. Available online: https://www.maff.go.jp/j/tokei/kouhyou/katiku_kyosai/ (accessed on 28 November 2020).
- De Rycke, J.; Bernard, S.; Laporte, J.; Naciri, M.; Popoff, M.R.; Rodolakis, A. Prevalence of various enteropathogens in the feces of diarrheic and healthy calves. Ann. Rech. Vet. 1986, 17, 159–168. [Google Scholar]
- Svensson, C. Peripartal excretion of Eimeria oocyst by cows on Swedish dairy farms and the age of calves at first excretion. Acta Vet. Scand. 1993, 34, 77–81. [Google Scholar]
- De La Fuente, R.; Luzón, M.; Ruiz-Santa-Quiteria, J.A.; García, A.; Cid, D.; Orden, J.A.; García, S.; Sanz, R.; Gómez-Bautista, M. Cryptosporidium and concurrent infections with other major enterophatogens in 1 to 30-day-old diarrheic dairy calves in central Spain. Vet. Parasitol. 1999, 80, 179–185. [Google Scholar] [CrossRef]
- Mawatari, T.; Hirano, K.; Ikeda, H.; Tsunemitsu, H.; Suzuki, T. Surveillance of diarrhea-causing pathogens in dairy and beef cows in Yamagata Prefecture, Japan from 2002 to 2011. Microbiol. Immunol. 2014, 58, 530–535. [Google Scholar] [CrossRef]
- Estes, M.; Kapikian, A. Rotaviruses. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 1917–1974. [Google Scholar]
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Parashar, U.D.; Agocs, M.; Serhan, F.; De Oliveira, L.; Mwenda, J.M.; Mihigo, R.; Ranjan, W.P.; et al. Global, Regional, and National Estimates of Rotavirus Mortality in Children <5 Years of Age, 2000–2013. Clin. Infect. Dis. 2016, 62, S96–S105. [Google Scholar] [CrossRef] [Green Version]
- Matthijnssens, J.; Otto, P.H.; Ciarlet, M.; Desselberger, U.; van Ranst, M.; Johne, R. VP6-sequence-based cutoff values as a criterion for rotavirus species demarcation. Arch. Virol. 2012, 157, 1177–1182. [Google Scholar] [CrossRef]
- Mihalov-Kovács, E.; Gellért, Á.; Marton, S.; Farkas, S.L.; Fehér, E.; Oldal, M.; Jakab, F.; Martella, V.; Bányai, K. Candidate new Rotavirus species in sheltered Dogs, Hungary. Emerg. Infect. Dis. 2015, 21, 660–663. [Google Scholar] [CrossRef]
- Bányai, K.; Kemenesi, G.; Budinski, I.; Földes, F.; Zana, B.; Marton, S.; Varga-Kugler, R.; Oldal, M.; Kurucz, K.; Jakab, F. Candidate new rotavirus species in Schreiber’s bats, Serbia. Infect. Genet. Evol. 2017, 48, 19–26. [Google Scholar] [CrossRef]
- Johne, R.; Tausch, S.H.; Grützke, J.; Falkenhagen, A.; Patzina-Mehling, C.; Beer, M.; Höper, D.; Ulrich, R.G. Distantly related rotaviruses in common shrews, Germany, 2004–2014. Emerg. Infect. Dis. 2019, 25, 2310–2314. [Google Scholar] [CrossRef]
- Pesavento, J.B.; Crawford, S.E.; Estes, M.K.; Prasad, B.V. Rotavirus proteins: Structure and assembly. Curr. Top. Microbiol. Immunol. 2006, 309, 189–219. [Google Scholar] [CrossRef]
- Ghosh, S.; Varghese, V.; Samajdar, S.; Sinha, M.; Naik, T.N.; Kobayashi, N. Evidence for bovine origin of VP4 and VP7 genes of human group A rotavirus G6P[14] and G10P[14] strains. J. Clin. Microbiol. 2007, 45, 2751–2753. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Duan, Z.; Zhang, Q.; Liu, N.; Xie, Z.; Jiang, B.; Steele, D.; Jiang, X.; Wang, Z.; Fang, Z. Molecular characterization of unusual human G5P[6] rotaviruses identified in China. J. Clin. Virol. 2008, 42, 141–148. [Google Scholar] [CrossRef]
- Nguyen, T.A.; Khamrin, P.; Trinh, Q.D.; Phan, T.G.; Pham, L.D.; Hoang, L.P.; Hoang, K.T.; Yagyu, F.; Okitsu, S.; Ushijima, H. Sequence analysis of Vietnamese P[6] rotavirus strains suggests evidence of interspecies transmission. J. Med. Virol. 2007, 79, 1959–1965. [Google Scholar] [CrossRef]
- Parra, G.I.; Vidales, G.; Gomez, J.A.; Fernandez, F.M.; Parreño, V.; Bok, K. Phylogenetic analysis of porcine rotavirus in Argentina: Increasing diversity of G4 strains and evidence of interspecies transmission. Vet. Microbiol. 2008, 126, 243–250. [Google Scholar] [CrossRef]
- Rahman, M.; Matthijnssens, J.; Yang, X.; Delbeke, T.; Arijs, I.; Taniguchi, K.; Iturriza-Gómara, M.; Iftekharuddin, N.; Azim, T.; Van Ranst, M. Evolutionary History and Global Spread of the Emerging G12 Human Rotaviruses. J. Virol. 2007, 81, 2382–2390. [Google Scholar] [CrossRef] [Green Version]
- Isegawa, Y.; Nakagomi, O.; Brüssow, H.; Minamoto, N.; Nakagomi, T.; Ueda, S. A Unique VP4 Gene Allele Carried by an Unusual Bovine Rotavirus Strain, 993/83. Virology 1994, 198, 366–369. [Google Scholar] [CrossRef]
- Mitake, H.; Ito, N.; Okadera, K.; Okada, K.; Nakagawa, K.; Tanaka, T.; Katsuragi, K.; Kasahara, K.; Nihongi, T.; Tsunemitsu, H.; et al. Detection of Avian-Like rotavirus a VP4 from a calf in Japan. J. Vet. Med. Sci. 2015, 77, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Abe, M.; Ito, N.; Morikawa, S.; Takasu, M.; Murase, T.; Kawashima, T.; Kawai, Y.; Kohara, J.; Sugiyama, M. Molecular epidemiology of rotaviruses among healthy calves in Japan: Isolation of a novel bovine rotavirus bearing new P and G genotypes. Virus Res. 2009, 144, 250–257. [Google Scholar] [CrossRef]
- Abe, M.; Ito, N.; Masatani, T.; Nakagawa, K.; Yamaoka, S.; Kanamaru, Y.; Suzuki, H.; Shibano, K.I.; Arashi, Y.; Sugiyama, M. Whole genome characterization of new bovine rotavirus G21P[29] and G24P[33] strains provides evidence for interspecies transmission. J. Gen. Virol. 2011, 92, 952–960. [Google Scholar] [CrossRef]
- Larsson, A.; Carlander, D. Oral immunotherapy with yolk antibodies to prevent infections in humans and animals. Upsala J. Med. Sci. 2003, 108, 129–140. [Google Scholar]
- Nguyen, S.V.; Umeda, K.; Yokoyama, H.; Tohya, Y.; Kodama, Y. Passive protection of dogs against clinical disease due to Canine parvovirus-2 by specific antibody from chicken egg yolk. Can. J. Vet. Res. 2006, 70, 62–64. [Google Scholar]
- Suzuki, H.; Nomura, S.; Masaoka, T.; Goshima, H.; Kamata, N.; Kodama, Y.; Ishii, H.; Kitajima, M.; Nomoto, K.; Hibi, T. Effect of dietary anti-Helicobacter pylori-urease immunoglobulin Y on Helicobacter pylori infection. Aliment. Pharmacol. Ther. 2004, 20, 185–192. [Google Scholar] [CrossRef]
- Hiraga, C.; Kodama, Y.; Sugiyama, T.; Ichikawa, Y. Prevention of human rotavirus infection with chicken egg yolk immunoglobulins containing rotavirus antibody in cat. Kansenshogaku Zasshi 1990, 64, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Hatta, H.; Tsuda, K.; Akachi, S.; Kim, M.; Yamamoto, T.; Ebina, T. Oral Passive Immulnization Effect of Anti-Human Rotavirus IgY and Its Behavior Against Proteolytic Enzymes. Biosci. Biotechnol. Biochem. 1993, 57, 1077–1081. [Google Scholar] [CrossRef]
- Ebina, T.; Tsukada, K.; Umezu, K.; Nose, M.; Tsuda, K.; Hatta, H.; Kim, M.; Yamamoto, T. Gastroenteritis in Suckling Mice Caused by Human Rotavirus Can Be Prevented with Egg Yolk Immunoglobulin (IgY) and Treated with a Protein-Bound Polysaccharide Preparation (PSK). Microbiol. Immunol. 1990, 34, 617–629. [Google Scholar] [CrossRef]
- Kuroki, M.; Ohta, M.; Ikemori, Y.; Icatlo, F.C.; Kobayashi, C.; Yokoyama, H.; Kodama, Y. Field evaluation of chicken egg yolk immunoglobulins specific for bovine rotavirus in neonatal calves. Arch. Virol. 1997, 142, 843–851. [Google Scholar] [CrossRef]
- Fujii, Y.; Shimoike, T.; Takagi, H.; Murakami, K.; Todaka-Takai, R.; Park, Y.; Katayama, K. Amplification of all 11 RNA segments of group A rotaviruses based on reverse transcription polymerase chain reaction. Microbiol. Immunol. 2012, 56, 630–638. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Matthijnssens, J.; Ciarlet, M.; Heiman, E.; Arijs, I.; Delbeke, T.; McDonald, S.M.; Palombo, E.A.; Iturriza-Gómara, M.; Maes, P.; Patton, J.T.; et al. Full Genome-Based Classification of Rotaviruses Reveals a Common Origin between Human Wa-Like and Porcine Rotavirus Strains and Human DS-1-Like and Bovine Rotavirus Strains. J. Virol. 2008, 82, 3204–3219. [Google Scholar] [CrossRef] [Green Version]
- Murakami, Y.; Nishioka, N.; Hashiguchi, Y.; Kuniyasu, C. Primary isolation of cytopathic bovine rotaviruses on fetal rhesus monkey kidney cells. Vet. Microbiol. 1983, 8, 135–139. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Percent Endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Nguyen, S.V.; Icatlo, F.C.; Nakano, T.; Isogai, E.; Hirose, K.; Mizugai, H.; Kobayashi-Sakamoto, M.; Isogai, H.; Chiba, I. Anti-cell-associated glucosyltransferase immunoglobulin Y suppression of salivary mutans streptococci in healthy young adults. J. Am. Dent. Assoc. 2011, 142, 943–949. [Google Scholar] [CrossRef]
- Ibrahim, E.S.M.; Rahman, A.K.M.S.; Isoda, R.; Umeda, K.; Nguyen, S.V.; Kodama, Y. In vitro and in vivo effectiveness of egg yolk antibody against Candida albicans (anti-CA IgY). Vaccine 2008, 26, 2073–2080. [Google Scholar] [CrossRef]
- Rahman, A.K.M.S.; Ibrahim, E.S.M.; Isoda, R.; Umeda, K.; Nguyen, S.V.; Kodama, Y. Effect of passive immunization by anti-gingipain IgY on periodontal health of dogs. Vet. Sci. Dev. 2011, 1, e8. [Google Scholar] [CrossRef]
- Kuroki, M.; Ikemori, Y.; Yokoyama, H.; Peralta, R.C.; Icatlo, F.C.; Kodama, Y. Passive protection against bovine rotavirus-induced diarrhea in murine model by specific immunoglobulins from chicken egg yolk. Vet. Microbiol. 1993, 37, 135–146. [Google Scholar] [CrossRef]
- Masuda, T.; Nagai, M.; Yamasato, H.; Tsuchiaka, S.; Okazaki, S.; Katayama, Y.; Oba, M.; Nishiura, N.; Sassa, Y.; Omatsu, T.; et al. Identification of novel bovine group A rotavirus G15P[14] strain from epizootic diarrhea of adult cows by de novo sequencing using a next-generation sequencer. Vet. Microbiol. 2014, 171, 66–73. [Google Scholar] [CrossRef]
- Sato, M.; Nakagomi, T.; Tajima, K.; Ezura, K.; Akashi, H.; Nakagomi, O. Isolation of serotype G8, P6[1] bovine rotavirus from adult cattle with diarrhea. J. Clin. Microbiol. 1997, 35, 1266–1268. [Google Scholar] [CrossRef] [Green Version]
- Fukai, K.; Sakai, T.; Hirose, M.; Itou, T. Prevalence of calf diarrhea caused by bovine group: A rotavirus carrying G serotype 8 specificity. Vet. Microbiol. 1999, 66, 301–311. [Google Scholar] [CrossRef]
- Fukai, K.; Maeda, Y.; Fujimoto, K.; Itou, T.; Sakai, T. Changes in the prevalence of rotavirus G and P types in diarrheic calves from the Kagoshima prefecture in Japan. Vet. Microbiol. 2002, 86, 343–349. [Google Scholar] [CrossRef]
- Fukai, K.; Saito, T.; Inoue, K.; Sato, M. Molecular characterization of novel P[14],G8 bovine group A rotavirus, Sun9, isolated in Japan. Virus Res. 2004, 105, 101–106. [Google Scholar] [CrossRef]
- Okada, N.; Matsumoto, Y. Bovine rotavirus G and P types and sequence analysis of the VP7 gene of two G8 bovine rotaviruses from JPN. Vet. Microbiol. 2002, 84, 297–305. [Google Scholar] [CrossRef]
- Abe, S.; Watanabe, D.; Itagaki, S. The preventive and therapeutic effects of administration of the mixture feedstuff including immunoglobulin against the calf diarrhea. J. Livest. Med. 1998, 423, 617–623. [Google Scholar]
- Kuroki, M.; Ohta, M.; Ikemori, Y.; Peralta, R.C.; Yokoyama, H.; Kodama, Y. Passive protection against bovine rotavirus in calves by specific immunoglobulins from chicken egg yolk. Arch. Virol. 1994, 138, 143–148. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Collection Area | Collection Data | Age | Cattle Type | VP7 | VP4 | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Open Reading Frame (Length) | G Genotype a | Accession Number | Open Reading Frame (Length) | P Genotype b | Accession Number | |||||
| HKD1 | Hokkaido | 2017.4 | 5 | Holstein | 981 | G6 | LC590915 | 2136 | P[5] | LC590916 |
| HKD2 | Hokkaido | 2017.4 | 10 | Holstein | 981 | G6 | LC590917 | 2226 | P[11] | LC590918 |
| HKD3 | Hokkaido | 2017.3 | 8 | Holstein | 981 | G6 | LC590919 | 2277 | P[11] | LC590920 |
| HKD5 | Hokkaido | 2017.3 | 7 | Holstein | 981 | G6 | LC590921 | 2283 | P[5] | LC590922 |
| HKD6 | Hokkaido | 2017.3 | 7 | Holstein | 981 | G6 | LC590923 | 2202 | P[11] | LC590924 |
| HKD7 | Hokkaido | 2017.3 | 9 | Holstein | 981 | G6 | LC590925 | 2199 | P[11] | LC590926 |
| HKD8 | Hokkaido | 2017.3 | 8 | Holstein | 981 | G6 | LC590927 | N.D. | P[x] | - |
| HKD9 | Hokkaido | 2017.3 | 7 | Holstein | 981 | G6 | LC590928 | 2196 | P[11] | LC590929 |
| HKD10 | Hokkaido | 2017.5 | 4 | Holstein | 981 | G6 | LC590930 | 2286 | P[5] | LC590931 |
| HKD11 | Hokkaido | 2017.2 | 6 | Holstein | 981 | G6 | LC590932 | N.D. | P[x] | - |
| HKD13 | Hokkaido | 2018.7 | <7 | Holstein | 981 | G6 | LC590933 | 2274 | P[11] | LC590934 |
| HKD14 | Hokkaido | 2018.8 | <7 | Holstein | 981 | G6 | LC590935 | 2274 | P[11] | LC590936 |
| HKD15 | Hokkaido | 2018.8 | <7 | Holstein | 981 | G6 | LC590937 | 2274 | P[11] | LC590938 |
| HKD16 | Hokkaido | 2017.12 | 10 | Holstein | 981 | G6 | LC590939 | 2268 | P[5] | LC590940 |
| HKD17 | Hokkaido | 2017.12 | 12 | Holstein | 981 | G6 | LC590941 | 2235 | P[11] | LC590942 |
| HKD18 | Hokkaido | 2018.3 | <7 | Holstein | 981 | G6 | LC590943 | 2274 | P[5] | LC590944 |
| HKD19 | Hokkaido | 2018.3 | <7 | Holstein | 981 | G6 | LC590945 | 2277 | P[5] | LC590946 |
| HKD20 | Hokkaido | 2018.3 | <7 | Holstein | 981 | G6 | LC590947 | 2277 | P[5] | LC590948 |
| HKD21 | Hokkaido | 2018.3 | <7 | Holstein | 981 | G6 | LC590949 | 2283 | P[5] | LC590950 |
| HKD22 | Hokkaido | 2018.3 | 10 | Holstein | 981 | G6 | LC590951 | 2274 | P[11] | LC590952 |
| HKD23 | Hokkaido | 2018.3 | 8 | Holstein | 981 | G6 | LC590953 | 2280 | P[5] | LC590954 |
| HKD24 | Hokkaido | 2018.3 | <7 | Japanese beef | 981 | G6 | LC590955 | 2265 | P[5] | LC590956 |
| HKD25 | Hokkaido | 2018.3 | <7 | Japanese beef | 981 | G6 | LC590957 | 2271 | P[5] | LC590958 |
| HKD27 | Hokkaido | 2018.3 | <7 | Japanese beef | 981 | G6 | LC590959 | 2274 | P[5] | LC590960 |
| HKD28 | Hokkaido | 2018.4 | 9 | Holstein | 981 | G6 | LC590961 | 2277 | P[5] | LC590962 |
| HKD29 | Hokkaido | 2018.4 | 9 | Japanese beef | 981 | G6 | LC590963 | 2277 | P[5] | LC590964 |
| SZO30 | Shizuoka | 2017.12 | 14 | F1 hybrid | 981 | G10 | LC590965 | 2265 | P[11] | LC590966 |
| OKY31 | Okayama | 2017.10 | 9 | F1 hybrid | 981 | G10 | LC590967 | 2268 | P[11] | LC590968 |
| OKY32 | Okayama | 2017.11 | 91 | Holstein | 981 | G10 | LC590969 | 2268 | P[11] | LC590970 |
| OKY33 | Okayama | 2018.1 | 62 | F1 hybrid | 981 | G10 | LC590971 | 2274 | P[11] | LC590972 |
| SMN34 | Shimane | 2017.11 | 10 | F1 hybrid | 981 | G6 | LC590973 | 2277 | P[5] | LC590974 |
| SMN35 | Shimane | 2018.1 | 13 | Japanese beef | 981 | G6 | LC590975 | 2286 | P[5] | LC590976 |
| SMN37 | Shimane | 2018.3 | 7 | Japanese beef | 981 | G6 | LC590977 | 2262 | P[5] | LC590978 |
| KGW39 | Kagawa | 2017.6 | 6 | F1 hybrid | 981 | G10 | LC590979 | 2268 | P[11] | LC590980 |
| KGW40 | Kagawa | 2017.7 | 9 | F1 hybrid | 981 | G10 | LC590981 | N.D. | P[x] | - |
| OKY41 | Okayama | 2018.4 | 5 | F1 hybrid | 981 | G6 | LC590982 | 2271 | P[5] | LC590983 |
| OKY42 | Okayama | 2019.1 | 14 | Holstein | 981 | G6 | LC590984 | 2277 | P[11] | LC590985 |
| OKY44 | Okayama | 2019.2 | 15 | F1 hybrid | 981 | G6 | LC590986 | 2277 | P[11] | LC590987 |
| OKY45 | Okayama | 2019.1 | 10 | Holstein | 981 | G6 | LC590988 | 2277 | P[11] | LC590989 |
| OKY46 | Okayama | 2019.1 | 16 | Holstein | 981 | G6 | LC590990 | 2277 | P[11] | LC590991 |
| OKY47 | Okayama | 2019.1 | 20 | F1 hybrid | 981 | G10 | LC590992 | 2277 | P[11] | LC590993 |
| OKY48 | Okayama | 2019.2 | 39 | F1 hybrid | 981 | G6 | LC590994 | 2277 | P[11] | LC590995 |
| OKY49 | Okayama | 2019.2 | 13 | Holstein | 981 | G6 | LC590996 | 2277 | P[11] | LC590997 |
| HKD50 | Hokkaido | 2019.1 | 17 | Holstein | 981 | G6 | LC590998 | 2271 | P[11] | LC590999 |
| HKD51 | Hokkaido | 2019.1 | 15 | Holstein | 981 | G6 | LC591000 | 2271 | P[11] | LC591001 |
| HKD52 | Hokkaido | 2019.6 | 17 | Holstein | 975 | G10 | LC591002 | 2271 | P[11] | LC591003 |
| HKD53 | Hokkaido | 2019.2 | 9-11 | Holstein | 981 | G6 | LC591004 | 2271 | P[11] | LC591005 |
| HKD54 | Hokkaido | 2019.2 | 9-11 | Holstein | 981 | G6 | LC591006 | 2271 | P[11] | LC591007 |
| HKD56 | Hokkaido | 2019.3 | 8 | Japanese beef | 981 | G6 | LC591008 | 2289 | P[5] | LC591009 |
| HKD57 | Hokkaido | 2019.3 | 24 | Japanese beef | 969 | G6 | LC591010 | 2289 | P[5] | LC591011 |
| HKD58 | Hokkaido | 2019.3 | 14 | Holstein | 981 | G6 | LC591012 | N.D. | P[x] | - |
| HKD59 | Hokkaido | 2019.3 | 10 | Holstein | 981 | G6 | LC591013 | 2271 | P[11] | LC591014 |
| HKD60 | Hokkaido | 2019.4 | 8 | Holstein | 981 | G6 | LC591015 | 2289 | P[5] | LC591016 |
| HKD61 | Hokkaido | 2019.4 | 8 | Holstein | 981 | G6 | LC591017 | 2289 | P[5] | LC591018 |
| HKD62 | Hokkaido | 2019.4 | 7 | Holstein | 981 | G6 | LC591019 | 2289 | P[5] | LC591020 |
| HKD63 | Hokkaido | 2019.5 | <7 | Holstein | 981 | G10 | LC591021 | 2271 | P[11] | LC591022 |
| HKD64 | Hokkaido | 2019.5 | <7 | Holstein | 981 | G10 | LC591023 | N.D. | P[x] | - |
| HKD65 | Hokkaido | 2019.5 | <7 | Holstein | 981 | G6 | LC591024 | 2289 | P[5] | LC591025 |
| HKD66 | Hokkaido | 2019.7 | <7 | Japanese beef | 981 | G6 | LC591026 | 2289 | P[5] | LC591027 |
| HKD67 | Hokkaido | 2019.7 | <7 | Japanese beef | 981 | G6 | LC591028 | 2289 | P[5] | LC591029 |
| HKD68 | Hokkaido | 2019.7 | <7 | Japanese beef | 981 | G6 | LC591030 | 2289 | P[5] | LC591031 |
| FKS69 | Fukushima | 2019.8 | 10 | Holstein | 981 | G6 | LC591032 | 2289 | P[5] | LC591033 |
| FKS70 | Fukushima | 2019.8 | 10 | Holstein | 981 | G6 | LC591034 | 2289 | P[5] | LC591035 |
| FKS71 | Fukushima | 2019.8 | 10 | Holstein | 981 | G6 | LC591036 | 2289 | P[5] | LC591037 |
| FKS72 | Fukushima | 2019.8 | 8 | F1 hybrid | 981 | G6 | LC591038 | 2289 | P[5] | LC591039 |
| FKS73 | Fukushima | 2019.8 | 10 | Japanese beef | 981 | G6 | LC591040 | 2289 | P[5] | LC591041 |
| OKY75 | Okayama | 2019.7 | 13 | F1 hybrid | 981 | G10 | LC591042 | 2271 | P[11] | LC591043 |
| OKY76 | Okayama | 2019.8 | 10 | Holstein | 975 | G10 | LC591044 | 2271 | P[11] | LC591045 |
| OKY77 | Okayama | 2019.8 | 13 | Holstein | 975 | G10 | LC591046 | 2271 | P[11] | LC591047 |
| OKY78 | Okayama | 2019.9 | 9 | Holstein | 981 | G10 | LC591048 | 2271 | P[11] | LC591049 |
| HKD79 | Hokkaido | 2019.10 | 5 | Holstein | 981 | G6 | LC591050 | 2289 | P[5] | LC591051 |
| HKD80 | Hokkaido | 2019.10 | 5 | Holstein | 981 | G6 | LC591052 | 2289 | P[5] | LC591053 |
| HKD81 | Hokkaido | 2019.10 | 5 | Holstein | 981 | G6 | LC591054 | 2289 | P[5] | LC591055 |
| HKD82 | Hokkaido | 2019.10 | 5 | Holstein | 981 | G6 | LC591056 | 2289 | P[5] | LC591057 |
| HKD83 | Hokkaido | 2019.11 | 13 | Japanese beef | 981 | G6 | LC591058 | 2283 | P[5] | LC591059 |
| HKD84 | Hokkaido | 2020.1 | 7 | Japanese beef | 981 | G6 | LC591060 | 2280 | P[5] | LC591061 |
| HKD85 | Hokkaido | 2019.12 | 5 | Japanese beef | 981 | G6 | LC591062 | 2247 | P[5] | LC591063 |
| HKD86 | Hokkaido | 2020.1 | 7 | Japanese beef | 981 | G6 | LC591064 | 2289 | P[5] | LC591065 |
| HKD87 | Hokkaido | 2020.2 | 12 | Japanese beef | 981 | G6 | LC591066 | 2289 | P[5] | LC591067 |
| HKD88 | Hokkaido | 2020.2 | 13 | Holstein | 981 | G10 | LC591068 | 2268 | P[11] | LC591069 |
| HKD89 | Hokkaido | 2020.2 | 4 | Holstein | 981 | G6 | LC591070 | 2289 | P[5] | LC591071 |
| HKD91 | Hokkaido | 2020.5 | 14 | F1 hybrid | 981 | G6 | LC591072 | 2292 | P[5] | LC591073 |
| FKS92 | Fukushima | 2020.2 | 9 | F1 hybrid | 957 | G6 | LC591074 | 2292 | P[5] | LC591075 |
| FKS93 | Fukushima | 2020.2 | 12 | Holstein | N.D. | Gx | - | 2235 | P[11] | LC591076 |
| FKS94 | Fukushima | 2020.3 | 10 | F1 hybrid | 981 | G10 | LC591077 | N.D. | P[x] | - |
| FKS95 | Fukushima | 2020.3 | 8 | F1 hybrid | 954 | G6 | LC591078 | 2136 | P[5] | LC591079 |
| MYZ96 | Miyazaki | 2020.2 | 8 | Japanese beef | 981 | G6 | LC591080 | 2220 | P[5] | LC591081 |
| MYZ97 | Miyazaki | 2020.2 | 10 | Japanese beef | 981 | G6 | LC591082 | 2292 | P[5] | LC591083 |
| KGS98 | Kagoshima | 2020.3 | 5 | Japanese beef | 981 | G6 | LC591084 | 2280 | P[5] | LC591085 |
| OKY100 | Okayama | 2019.12 | 7 | Japanese beef | 981 | G6 | LC591086 | 2286 | P[5] | LC591087 |
| OKY101 | Okayama | 2020.2 | 9 | Holstein | N.D. | Gx | - | 2200 | P[11] | LC591088 |
| OKY102 | Okayama | 2020.4 | 21 | Japanese beef | 981 | G6 | LC591089 | 2154 | P[5] | LC591090 |
| OKY103 | Okayama | 2020.4 | 6 | Japanese beef | 981 | G6 | LC591091 | 2292 | P[5] | LC591092 |
| OKY104 | Okayama | 2020.4 | 5 | Japanese beef | 981 | G6 | LC591093 | 2286 | P[5] | LC591094 |
| OKY105 | Okayama | 2020.4 | 14 | Holstein | 981 | G6 | LC591095 | N.D. | P[x] | - |
| OKY106 | Okayama | 2020.4 | 5 | Japanese beef | 981 | G6 | LC591096 | 2292 | P[5] | LC591097 |
| OKY107 | Okayama | 2020.4 | 6 | Japanese beef | 981 | G6 | LC591098 | 2286 | P[5] | LC591099 |
| OKY108 | Okayama | 2020.5 | 10 | Japanese beef | 981 | G6 | LC591100 | 2286 | P[5] | LC591101 |
| OKY109 | Okayama | 2020.5 | 6 | Holstein | 921 | G6 | LC591102 | N.D. | P[x] | - |
| IgY | Genotype | Neutralization Antibody titer of Three IgYs against Bovine RVA with Different Genotypes a | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SMN-1 | HKD18 | SMN35 | HKD6 | HKD7 | HKD17 | KK-3 | OKY31 | MYG-1 | Dai-10 | ||
| G6P[1] | G6P[5] | G6P[5] | G6P[11] | G6P[11] | G6P[11] | G10P[11] | G10P[11] | G8P[14] | G24P[33] | ||
| Anti-SMN35 IgY | G6P[5] | 2560 | 2560 | 5120 | 1280 | 1280 | 1280 | 160 | 80 | 80 | <80 |
| Anti-OKY31 IgY | G10P[11] | <160 | 320 | 160 | 640 | 640 | 640 | 2560 | 2560 | <80 | <80 |
| Control IgY | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Odagiri, K.; Yoshizawa, N.; Sakihara, H.; Umeda, K.; Rahman, S.; Nguyen, S.V.; Suzuki, T. Development of Genotype-Specific Anti-Bovine Rotavirus A Immunoglobulin Yolk Based on a Current Molecular Epidemiological Analysis of Bovine Rotaviruses A Collected in Japan during 2017–2020. Viruses 2020, 12, 1386. https://0-doi-org.brum.beds.ac.uk/10.3390/v12121386
Odagiri K, Yoshizawa N, Sakihara H, Umeda K, Rahman S, Nguyen SV, Suzuki T. Development of Genotype-Specific Anti-Bovine Rotavirus A Immunoglobulin Yolk Based on a Current Molecular Epidemiological Analysis of Bovine Rotaviruses A Collected in Japan during 2017–2020. Viruses. 2020; 12(12):1386. https://0-doi-org.brum.beds.ac.uk/10.3390/v12121386
Chicago/Turabian StyleOdagiri, Koki, Nobuki Yoshizawa, Hisae Sakihara, Koji Umeda, Shofiqur Rahman, Sa Van Nguyen, and Tohru Suzuki. 2020. "Development of Genotype-Specific Anti-Bovine Rotavirus A Immunoglobulin Yolk Based on a Current Molecular Epidemiological Analysis of Bovine Rotaviruses A Collected in Japan during 2017–2020" Viruses 12, no. 12: 1386. https://0-doi-org.brum.beds.ac.uk/10.3390/v12121386